
Complete Genomic Characterization of Lumpy Skin Disease Virus Isolates from Beef Cattle in Lopburi Province, Central Thailand, during 2021–2022

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sample Preparation
2.3. Real-Time PCR
2.4. LSDV Isolation and Identification
2.5. Whole Genome Sequencing (WGS) for LSDV
2.6. DNA Sequencing, ORF023
2.7. Nucleotide Mutation and Phylogenetic Analysis
3. Results
3.1. Animal and Sample Data
3.2. Virus Detection
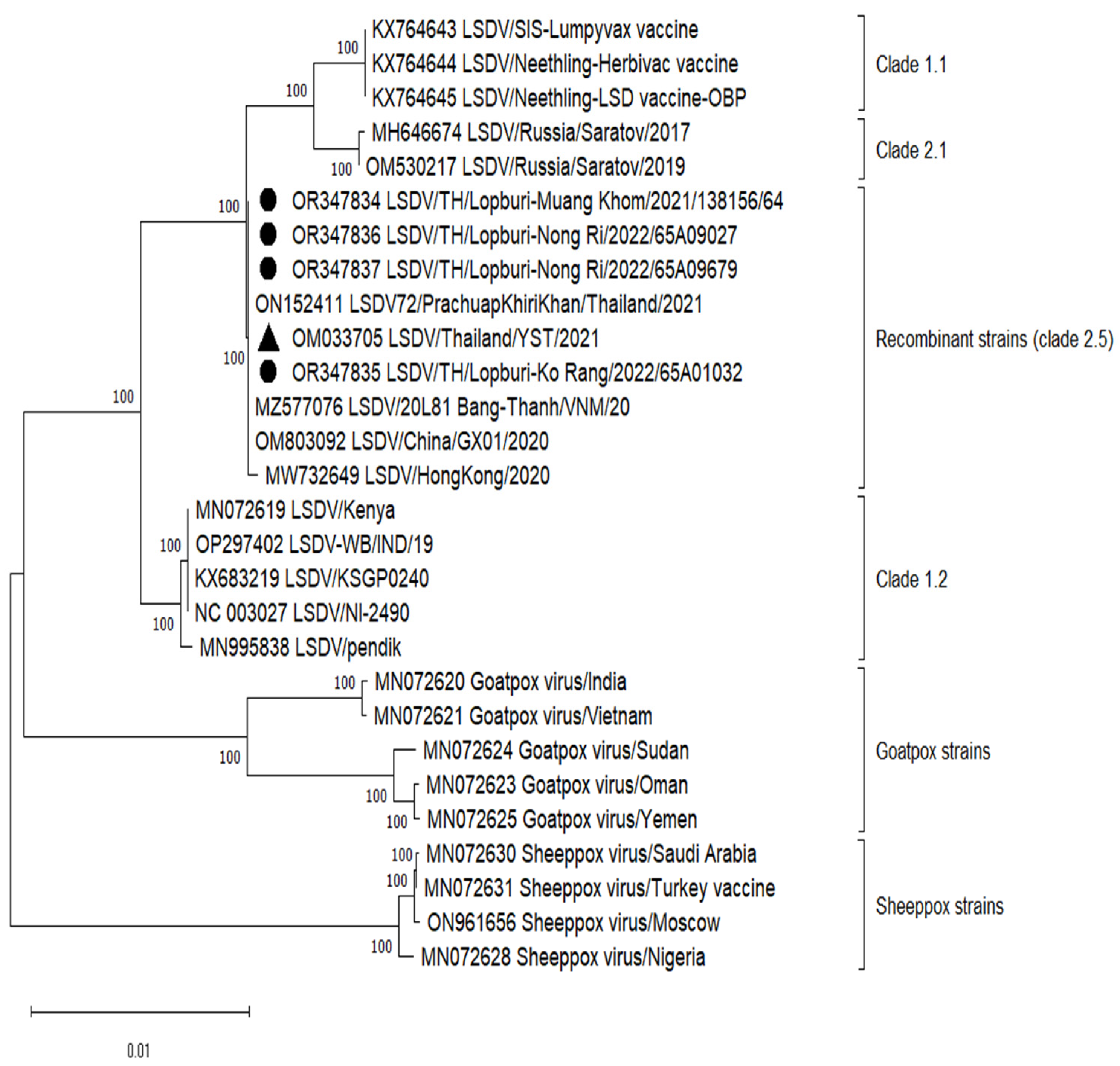
3.3. Whole Genome Sequencing and Phylogenetic Analysis
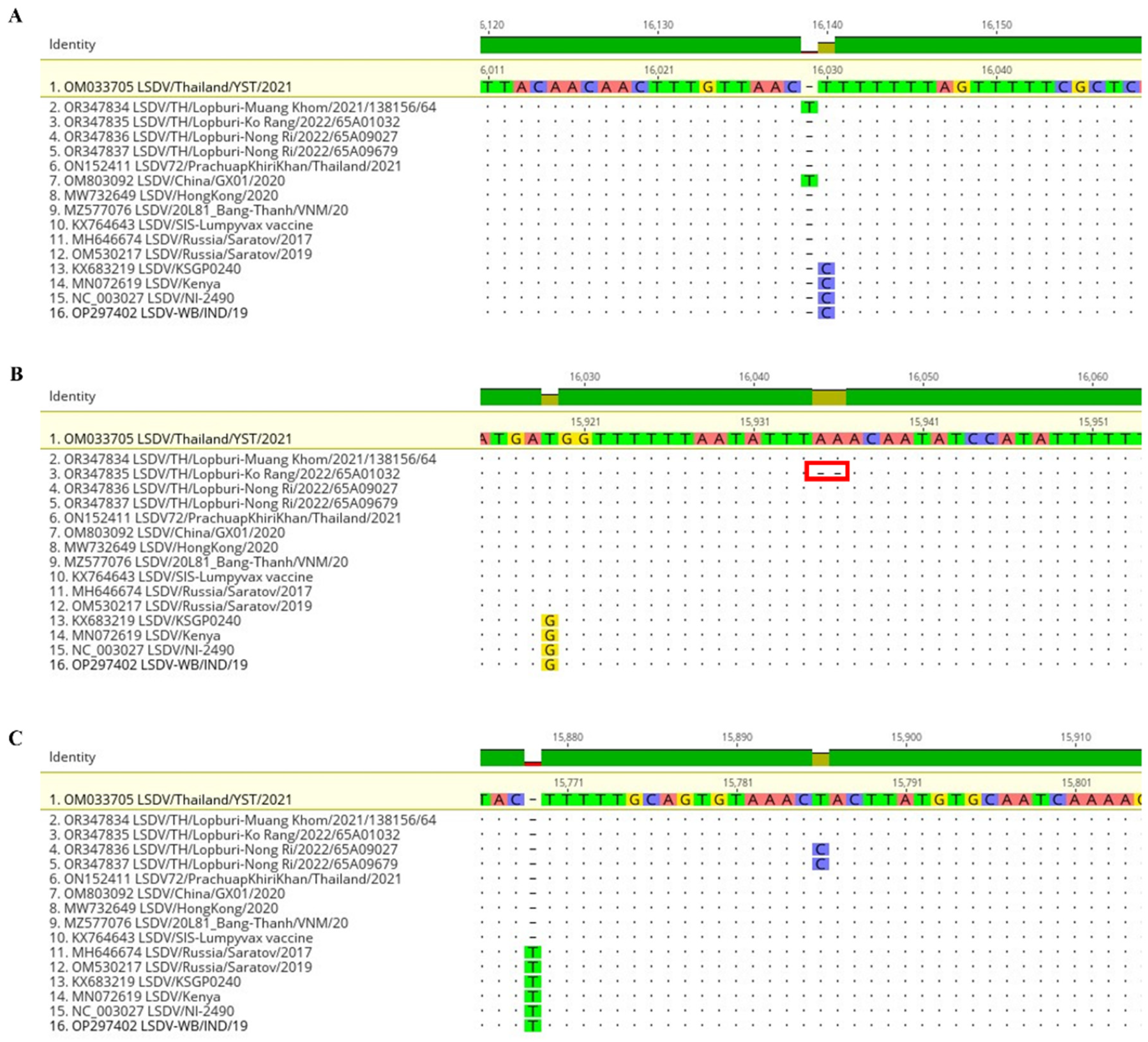
3.4. Mutation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Das, M.; Chowdhury, M.S.R.; Akter, S.; Mondal, A.K.; Jamal, M. An updated review on lumpy skin disease: Perspective of Southeast Asian countries. J. Adv. Biotechnol. Exp. Ther. 2021, 4, 322–333. [Google Scholar] [CrossRef]
- Dao, T.D.; Tran, L.H.; Nguyen, H.D.; Hoang, T.T.; Nguyen, G.H.; Tran, K.V.D.; Nguyen, H.X.; Van Dong, H.; Bui, A.N.; Bui, V.N. Characterization of Lumpy skin disease virus isolated from a giraffe in Vietnam. Transbound. Emerg. Dis. 2022, 69, e3268–e3272. [Google Scholar] [CrossRef] [PubMed]
- Sudhakar, S.B.; Mishra, N.; Kalaiyarasu, S.; Ahirwar, K.; Chatterji, S.; Parihar, O.; Singh, V.P.; Sanyal, A. Lumpy Skin Disease Virus Infection in Free-Ranging Indian Gazelles (Gazella bennettii), Rajasthan, India. Emerg. Infect. Dis. 2023, 29, 1407. [Google Scholar] [CrossRef] [PubMed]
- Suwankitwat, N.; Songkasupa, T.; Boonpornprasert, P.; Sripipattanakul, P.; Theerawatanasirikul, S.; Deemagarn, T.; Suwannaboon, M.; Arjkumpa, O.; Buamithup, N.; Hongsawat, A.; et al. Rapid Spread and Genetic Characterisation of a Recently Emerged Recombinant Lumpy Skin Disease Virus in Thailand. Vet. Sci. 2022, 9, 542. [Google Scholar] [CrossRef] [PubMed]
- Tageldin, M.H.; Wallace, D.B.; Gerdes, G.H.; Putterill, J.F.; Greyling, R.R.; Phosiwa, M.N.; Al Busaidy, R.M.; Al Ismaaily, S.I. Lumpy skin disease of cattle: An emerging problem in the Sultanate of Oman. Trop. Anim. Health Prod. 2014, 46, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Mathivanan, E.; Raju, K.; Murugan, R. Outbreak of lumpy skin disease in India 2022—An emerging threat to livestock & livelihoods. Glob. Biosecurity 2023, 5. [Google Scholar] [CrossRef]
- Amanuel, S.; Kassa, T. Study on major ectoparasites and associated skin diseases of ruminants. Glob. J. Anim. Sci. Res. 2023, 11, 62–75. [Google Scholar]
- Selim, A.; Manaa, E.; Khater, H. Seroprevalence and risk factors for lumpy skin disease in cattle in Northern Egypt. Trop. Anim. Health Prod. 2021, 53, 350. [Google Scholar] [CrossRef]
- Aleksandr, K.; Olga, B.; David, W.B.; Pavel, P.; Yana, P.; Svetlana, K.; Alexander, N.; Vladimir, R.; Dmitriy, L.; Alexander, S. Non-vector-borne transmission of lumpy skin disease virus. Sci. Rep. 2020, 10, 7436. [Google Scholar] [CrossRef]
- Haegeman, A.; Sohier, C.; Mostin, L.; De Leeuw, I.; Van Campe, W.; Philips, W.; De Regge, N.; De Clercq, K. Evidence of Lumpy Skin Disease Virus Transmission from Subclinically Infected Cattle by Stomoxys calcitrans. Viruses 2023, 15, 1285. [Google Scholar] [CrossRef]
- Sprygin, A.; Pestova, Y.; Wallace, D.B.; Tuppurainen, E.; Kononov, A.V. Transmission of lumpy skin disease virus: A short review. Virus Res. 2019, 269, 197637. [Google Scholar] [CrossRef] [PubMed]
- Namazi, F.; Khodakaram Tafti, A. Lumpy skin disease, an emerging transboundary viral disease: A review. Vet. Med. Sci. 2021, 7, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Limon, G.; Gamawa, A.A.; Ahmed, A.I.; Lyons, N.A.; Beard, P.M. Epidemiological Characteristics and Economic Impact of Lumpy Skin Disease, Sheeppox and Goatpox Among Subsistence Farmers in Northeast Nigeria. Front. Vet. Sci. 2020, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Tuppurainen, E.; Dietze, K.; Wolff, J.; Bergmann, H.; Beltran-Alcrudo, D.; Fahrion, A.; Lamien, C.E.; Busch, F.; Sauter-Louis, C.; Conraths, F.J.; et al. Review: Vaccines and Vaccination against Lumpy Skin Disease. Vaccines 2021, 9, 1136. [Google Scholar] [CrossRef] [PubMed]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Kutish, G.F.; Rock, D.L. Genome of lumpy skin disease virus. J. Virol. 2001, 75, 7122–7130. [Google Scholar] [CrossRef]
- WOAH. Chapter 3.4.12 Lumpy Skin Disease. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.04.12_LSD.pdf (accessed on 17 August 2023).
- Sprygin, A.; Babin, Y.; Pestova, Y.; Kononova, S.; Wallace, D.B.; Van Schalkwyk, A.; Byadovskaya, O.; Diev, V.; Lozovoy, D.; Kononov, A. Analysis and insights into recombination signals in lumpy skin disease virus recovered in the field. PLoS ONE 2018, 13, e0207480. [Google Scholar] [CrossRef]
- Ma, J.; Yuan, Y.; Shao, J.; Sun, M.; He, W.; Chen, J.; Liu, Q. Genomic characterization of lumpy skin disease virus in southern China. Transbound. Emerg. Dis. 2021, 69, 1–12. [Google Scholar] [CrossRef]
- Maw, M.T.; Khin, M.M.; Hadrill, D.; Meki, I.K.; Settypalli, T.B.K.; Kyin, M.M.; Myint, W.W.; Thein, W.Z.; Aye, O.; Palamara, E. First Report of Lumpy Skin Disease in Myanmar and Molecular Analysis of the Field Virus Isolates. Microorganisms 2022, 10, 897. [Google Scholar] [CrossRef]
- Tran, H.T.T.; Truong, A.D.; Dang, A.K.; Ly, D.V.; Nguyen, C.T.; Chu, N.T.; Hoang, T.V.; Nguyen, H.T.; Nguyen, V.T.; Dang, H.V. Lumpy skin disease outbreaks in Vietnam, 2020. Transbound. Emerg. Dis. 2021, 68, 977–980. [Google Scholar] [CrossRef]
- Vandenbussche, F.; Mathijs, E.; Philips, W.; Saduakassova, M.; De Leeuw, I.; Sultanov, A.; Haegeman, A.; De Clercq, K. Recombinant LSDV Strains in Asia: Vaccine Spillover or Natural Emergence? Viruses 2022, 14, 1429. [Google Scholar] [CrossRef]
- Kumar, N.; Barua, S.; Kumar, R.; Khandelwal, N.; Kumar, A.; Verma, A.; Singh, L.; Godara, B.; Chander, Y.; Kumar, G. Evaluation of the safety, immunogenicity and efficacy of a new live-attenuated lumpy skin disease vaccine in India. Virulence 2023, 14, 2190647. [Google Scholar] [CrossRef] [PubMed]
- Sariya, L.; Paungpin, W.; Chaiwattanarungruengpaisan, S.; Thongdee, M.; Nakthong, C.; Jitwongwai, A.; Taksinoros, S.; Sutummaporn, K.; Boonmasawai, S.; Kornmatitsuk, B. Molecular detection and characterization of lumpy skin disease viruses from outbreaks in Thailand in 2021. Transbound. Emerg. Dis. 2022, 69, e2145–e2152. [Google Scholar] [CrossRef] [PubMed]
- Singhla, T.; Boonsri, K.; Kreausukon, K.; Modethed, W.; Pringproa, K.; Sthitmatee, N.; Punyapornwithaya, V.; Vinitchaikul, P. Molecular Characterization and Phylogenetic Analysis of Lumpy Skin Disease Virus Collected from Outbreaks in Northern Thailand in 2021. Vet. Sci. 2022, 9, 194. [Google Scholar] [CrossRef] [PubMed]
- Arjkumpa, O.; Suwannaboon, M.; Boonrawd, M.; Punyawan, I.; Laobannu, P.; Yantaphan, S.; Bungwai, A.; Ponyium, V.; Suwankitwat, N.; Boonpornprasert, P.; et al. First emergence of lumpy skin disease in cattle in Thailand, 2021. Transboundary and Emerging Diseases 2021, 68, 3002–3004. [Google Scholar] [CrossRef] [PubMed]
- Suwankitwat, N.; Deemagarn, T.; Bhakha, K.; Songkasupa, T.; Lekcharoensuk, P.; Arunvipas, P. Monitoring of genetic alterations of lumpy skin disease virus in cattle after vaccination in Thailand. J. Anim. Sci. Technol. 2023. [Google Scholar] [CrossRef]
- Bowden, T.R.; Babiuk, S.L.; Parkyn, G.R.; Copps, J.S.; Boyle, D.B. Capripoxvirus tissue tropism and shedding: A quantitative study in experimentally infected sheep and goats. Virology 2008, 371, 380–393. [Google Scholar] [CrossRef] [PubMed]
- Suwankitwat, N.; Bhakha, K.; Molee, L.; Songkasupa, T.; Puangjinda, K.; Chamchoy, T.; Arjkumpa, O.; Nuansrichay, B.; Srisomrun, S.; Pongphitcha, P. Long-term monitoring of immune response to recombinant lumpy skin disease virus in dairy cattle from small-household farms in western Thailand. Comp. Immunol. Microbiol. Infect. Dis. 2023, 99, 102008. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Breman, F.C.; Haegeman, A.; Krešić, N.; Philips, W.; De Regge, N. Lumpy Skin Disease Virus Genome Sequence Analysis: Putative Spatio-Temporal Epidemiology, Single Gene versus Whole Genome Phylogeny and Genomic Evolution. Viruses 2023, 15, 1471. [Google Scholar] [CrossRef]
- Han, G.-Z.; Liu, X.-P.; Li, S.-S. Caution about Newcastle disease virus-based live attenuated vaccine. J. Virol. 2008, 82, 6782. [Google Scholar] [CrossRef]
- He, D.C.; He, C.-Q. Discovery of vaccine-like recombinant SARS-CoV-2 circulating in human. Virol. J. 2022, 19, 209. [Google Scholar] [CrossRef] [PubMed]
- van Schalkwyk, A.; Kara, P.; Ebersohn, K.; Mather, A.; Annandale, C.H.; Venter, E.H.; Wallace, D.B. Potential link of single nucleotide polymorphisms to virulence of vaccine-associated field strains of lumpy skin disease virus in South Africa. Transbound. Emerg. Dis. 2020, 67, 2946–2960. [Google Scholar] [CrossRef] [PubMed]
- DLD. Cattle in Thailand 2022. Available online: https://ict.dld.go.th/webnew/images/stories/stat_web/yearly/2565/province/T2-1-Cattle.pdf (accessed on 20 September 2023).
- Pivova, E.Y.; Vlasov, M.E.; Sevskikh, T.A.; Povolyaeva, O.S.; Zhivoderov, S.P. A Study of the Susceptibility of Laboratory Animals to the Lumpy Skin Disease Virus. Life 2023, 13, 1489. [Google Scholar] [CrossRef] [PubMed]
- Debing, Y.; Ramière, C.; Dallmeier, K.; Piorkowski, G.; Trabaud, M.-A.; Lebossé, F.; Scholtès, C.; Roche, M.; Legras-Lachuer, C.; De Lamballerie, X. Hepatitis E virus mutations associated with ribavirin treatment failure result in altered viral fitness and ribavirin sensitivity. J. Hepatol. 2016, 65, 499–508. [Google Scholar] [CrossRef]
- Bazid, A.-H.; Wasfy, M.; Fawzy, M.; Nayel, M.; Abdelmegeid, M.; Thabet, R.Y.; Yong, H.S.; El-Sayed, M.M.; Magouz, A.; Badr, Y. Emergency vaccination of cattle against lumpy skin disease: Evaluation of safety, efficacy, and potency of MEVAC® LSD vaccine containing Neethling strain. Vet. Res. Commun. 2023, 47, 767–777. [Google Scholar] [CrossRef]
- Molla, W.; de Jong, M.C.M.; Gari, G.; Frankena, K. Economic impact of lumpy skin disease and cost effectiveness of vaccination for the control of outbreaks in Ethiopia. Prev. Vet. Med. 2017, 147, 100–107. [Google Scholar] [CrossRef]
- Vinitchaikul, P.; Punyapornwithaya, V.; Seesupa, S.; Phuykhamsingha, S.; Arjkumpa, O.; Sansamur, C.; Jarassaeng, C. The first study on the impact of lumpy skin disease outbreaks on monthly milk production on dairy farms in Khon Kaen, Thailand. Vet. World 2023, 16, 687. [Google Scholar] [CrossRef]
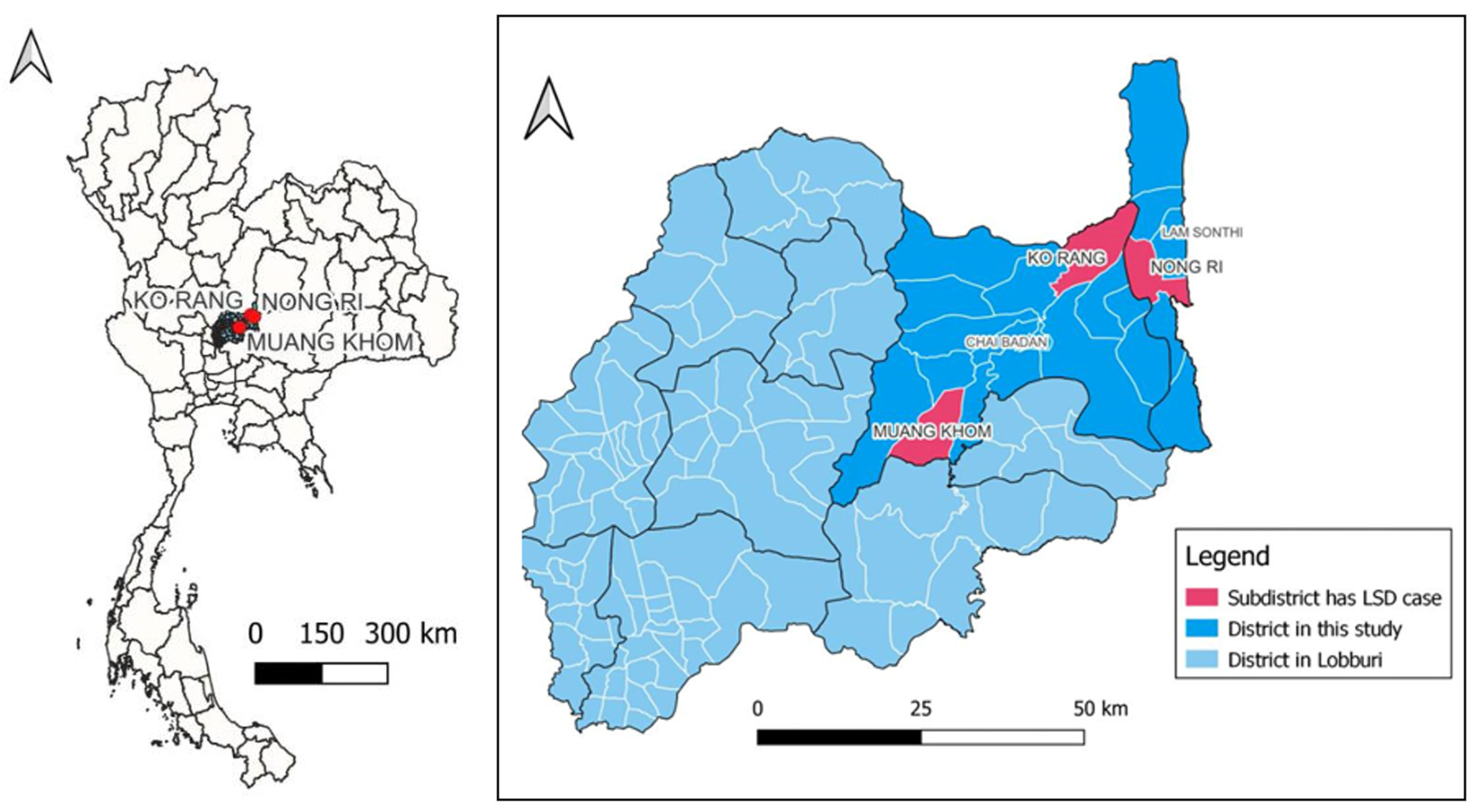

{kind=link}
{kind=link}
{kind=link}
| No. | Sample Received Date | Animal | Age (Months) | Morbidity Rate (%) | Mortality Rate (%) | Subdistrict | District | Initial Ct | Passage for WGS |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 28 September 2021 | Beef cattle | 2 | 4 | 0 | Muang Khom | Chai Badan | 19.4 | 5 |
| 2 | 15 February 2022 | Beef cattle | 2 | 50 | 0 | Ko Rang | Chai Badan | 19.4 | 6 |
| 3 | 10 June 2022 | Beef cattle | 5 | 11.76 | 5.88 | Nong Ri | Lam Sonthi | 20.9 | 4 |
| 4 | 21 June 2022 | Beef cattle | 5 | 11.76 | 5.88 | Nong Ri | Lam Sonthi | 22.5 | 3 |
| No. | Number of Reads | Number of Trimmed Reads (Q30/L20) | Size of Largest Contig or Scaffold (bp) | Number of Trimmed Reads * | Mean of Coverage Depth * | Genome Size (bp) | % Identity to YST/2021 | Number of Differences in Nucleotide | Affected ORF (Protein) | SNPs (Position) | Mutation Type | Amino Acid Change | GenBank Accession No. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 9,048,280 | 7,540,224 | 146,141 | 399,550 399,549 | 415.6 414.7 | 150,857 | 99.9993 | 1 | 023 (hypothetical protein) | - → T (16,030) | Insertion | V → S*stop (del 64 aa) | OR347834 |
| 2 | 26,229,520 | 22,445,168 | 145,884 | 441,854 537,976 | 335.6 523.8 | 150,854 | 99.9987 | 2 | 023 (hypothetical protein) | AA → -- (15935–15936) | Deletion | LNI → KY*stop (del 32 aa) | OR347835 |
| 3 | 9,278,280 | 7,587,054 | 145,886 | 377,662 377,661 | 388.5 388.1 | 150,856 | 99.9993 | 1 | UTR | T → C (15,786) | Transition | - | OR347836 |
| 4 | 15,513,540 | 13,656,538 | 145,886 | 333,454 333,454 | 389.8 389.3 | 150,856 | 99.9993 | 1 | UTR | T → C (15.786) | Transition | - | OR347837 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suwankitwat, N.; Deemagarn, T.; Bhakha, K.; Songkasupa, T.; Vitoonpong, R.; Trakunjaroonkit, P.; Rodphol, S.; Nuansrichay, B.; Chintapitaksakul, L.; Wongsarattanasin, K.; et al. Complete Genomic Characterization of Lumpy Skin Disease Virus Isolates from Beef Cattle in Lopburi Province, Central Thailand, during 2021–2022. Vet. Sci. 2024, 11, 10. https://doi.org/10.3390/vetsci11010010
Suwankitwat N, Deemagarn T, Bhakha K, Songkasupa T, Vitoonpong R, Trakunjaroonkit P, Rodphol S, Nuansrichay B, Chintapitaksakul L, Wongsarattanasin K, et al. Complete Genomic Characterization of Lumpy Skin Disease Virus Isolates from Beef Cattle in Lopburi Province, Central Thailand, during 2021–2022. Veterinary Sciences. 2024; 11(1):10. https://doi.org/10.3390/vetsci11010010
Chicago/Turabian StyleSuwankitwat, Nutthakarn, Taweewat Deemagarn, Kultyarat Bhakha, Tapanut Songkasupa, Ratchaneekorn Vitoonpong, Pannaporn Trakunjaroonkit, Sureenipa Rodphol, Bandit Nuansrichay, Lerdchai Chintapitaksakul, Khanin Wongsarattanasin, and et al. 2024. "Complete Genomic Characterization of Lumpy Skin Disease Virus Isolates from Beef Cattle in Lopburi Province, Central Thailand, during 2021–2022" Veterinary Sciences 11, no. 1: 10. https://doi.org/10.3390/vetsci11010010
APA StyleSuwankitwat, N., Deemagarn, T., Bhakha, K., Songkasupa, T., Vitoonpong, R., Trakunjaroonkit, P., Rodphol, S., Nuansrichay, B., Chintapitaksakul, L., Wongsarattanasin, K., Kwon, O.-K., Kang, H.-E., & Shin, Y.-K. (2024). Complete Genomic Characterization of Lumpy Skin Disease Virus Isolates from Beef Cattle in Lopburi Province, Central Thailand, during 2021–2022. Veterinary Sciences, 11(1), 10. https://doi.org/10.3390/vetsci11010010