
Litters of Various-Sized Mummies (LVSM) and Stillborns after Porcine Reproductive and Respiratory Syndrome Virus Type 1 Infection—A Case Report

 , ,
, , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Case Presentation
2.1. Anamnesis and Physical Findings
2.2. Diagnostic Methods and Laboratory Findings
2.3. Further Steps and Outcome of the Case
3. Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dunne, H.W.; Gobble, J.L.; Hokanson, J.F.; Kradel, D.C.; Bubash, G.R. Porcine reproductive failure associated with a newly identified “SMEDI” group of picorna viruses. Am. J. Vet. Res. 1965, 26, 1284–1297. [Google Scholar] [PubMed]
- Streck, A.F.; Truyen, U. Porcine parvovirus. Curr. Issues Mol. Biol. 2020, 37, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Madson, D.M.; Patterson, A.R.; Ramamoorthy, S.; Pal, N.; Meng, X.J.; Opriessnig, T. Reproductive failure experimentally induced in sows via artificial insemination with semen spiked with porcine circovirus type 2. Vet. Pathol. 2009, 46, 707–716. [Google Scholar] [CrossRef]
- Love, R.J.; Grewal, A.S. Reproductive failure in pigs caused by encephalomyocarditis virus. Aust. Vet. J. 1986, 63, 128–129. [Google Scholar] [CrossRef] [PubMed]
- Arent, Z.J.; Ellis, W.A. Leptospirosis. In Diseases of Swine; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; Wiley: Hoboken, NY, USA, 2019; pp. 854–862. ISBN 9781119350859. [Google Scholar]
- Arthur, G.H. Arthur’s Veterinary Reproduction and Obstetrics; Elsevier: Amsterdam, The Netherlands, 2001; ISBN 9780702025563. [Google Scholar]
- Carr, J.; Chen, S.; Conner, J.F.; Kirkwood, R.N.; Segalés, J. Pig Health; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2018; ISBN 978-1-4987-0472-4. [Google Scholar]
- Christianson, W.T. Stillbirths, mummies, abortions, and early embryonic death. Vet. Clin. N. Am. Food Anim. Pract. 1992, 8, 623–639. [Google Scholar] [CrossRef]
- Van der Lende, T.; van Rens, B. Critical periods for foetal mortality in gilts identified by analysing the length distribution of mummified foetuses and frequency of non-fresh stillborn piglets. Anim. Reprod. Sci. 2003, 75, 141–150. [Google Scholar] [CrossRef]
- Neumann, E.J.; Kliebenstein, J.B.; Johnson, C.D.; Mabry, J.W.; Bush, E.J.; Seitzinger, A.H.; Green, A.L.; Zimmerman, J.J. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J. Am. Vet. Med. Assoc. 2005, 227, 385–392. [Google Scholar] [CrossRef]
- Zimmerman, J.J.; Dee, S.A.; Holtkamp, D.J.; Murtaugh, M.P.; Stadejek, T.; Stevenson, G.W.; Torremorell, M.; Yang, H.; Zhang, J. Porcine reproductive and respiratory syndrome viruses (porcine arteriviruses). In Diseases of Swine; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; Wiley: Hoboken, NY, USA, 2019; pp. 685–708. ISBN 9781119350859. [Google Scholar]
- Prieto, C.; Suarez, P.; Simarro, I.; Garcia, C.; Fernandez, A.; Castro, J.M. Transplacental infection following exposure of gilts to porcine reproductive and respiratory syndrome virus at the onset of gestation. Vet. Microbiol. 1997, 57, 301–311. [Google Scholar] [CrossRef]
- Sinn, L.J.; Klingler, E.; Lamp, B.; Brunthaler, R.; Weissenböck, H.; Rümenapf, T.; Ladinig, A. Emergence of a virulent porcine reproductive and respiratory syndrome virus (PRRSV) 1 strain in Lower Austria. Porc. Health Manag. 2016, 2, 28. [Google Scholar] [CrossRef]
- Terpstra, C.; Wensvoort, G.; Pol, J.M. Experimental reproduction of porcine epidemic abortion and respiratory syndrome (mystery swine disease) by infection with Lelystad virus: Koch’s postulates fulfilled. Vet. Q. 1991, 13, 131–136. [Google Scholar] [CrossRef]
- Ladinig, A.; Foxcroft, G.; Ashley, C.; Lunney, J.K.; Plastow, G.; Harding, J.C.S. Birth weight, intrauterine growth retardation and fetal susceptibility to porcine reproductive and respiratory syndrome virus. PLoS ONE 2014, 9, e109541. [Google Scholar] [CrossRef] [PubMed]
- Kreutzmann, H.; Stadler, J.; Knecht, C.; Sassu, E.L.; Ruczizka, U.; Zablotski, Y.; Vatzia, E.; Balka, G.; Zaruba, M.; Chen, H.-W.; et al. Phenotypic Characterization of a Virulent PRRSV-1 Isolate in a Reproductive Model with and Without Prior Heterologous Modified Live PRRSV-1 Vaccination. Front. Vet. Sci. 2022, 9, 820233. [Google Scholar] [CrossRef] [PubMed]
- Ladinig, A.; Ashley, C.; Detmer, S.E.; Wilkinson, J.M.; Lunney, J.K.; Plastow, G.; Harding, J.C.S. Maternal and fetal predictors of fetal viral load and death in third trimester, type 2 porcine reproductive and respiratory syndrome virus infected pregnant gilts. Vet. Res. 2015, 46, 107. [Google Scholar] [CrossRef] [PubMed]
- Scortti, M.; Prieto, C.; Simarro, I.; Castro, J.M. Reproductive performance of gilts following vaccination and subsequent heterologous challenge with European strains of porcine reproductive and respiratory syndrome virus. Theriogenology 2006, 66, 1884–1893. [Google Scholar] [CrossRef] [PubMed]
- Kranker, S.; Nielsen, J.; Bille-Hansen, V.; Bøtner, A. Experimental inoculation of swine at various stages of gestation with a Danish isolate of porcine reproductive and respiratory syndrome virus (PRRSV). Vet. Microbiol. 1998, 61, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Christianson, W.T.; Choi, C.-S.; Collins, J.E.; Molitor, T.W.; Morrison, R.B.; Joo, H.-S. Pathogenesis of porcine reproductive and respiratory syndrome virus infection in mid-gestation sows and fetuses. Can. J. Vet. Res. 1993, 57, 262. [Google Scholar] [PubMed]
- Holtkamp, D.; Torremorell, M.; Corzo, C.; Linhares, D.; Nunes de Almeida, M.; Polson, D.; Snelson, H.; Silva, G.; Sanhueza, J.; Vilalta, C.; et al. Proposed modifications to porcine reproductive and respiratory syndrome virus herd classification. J. Swine Health Prod. 2021, 29, 261–270. [Google Scholar]
- Opriessnig, T.; Shen, H.G.; Pal, N.; Ramamoorthy, S.; Huang, Y.W.; Lager, K.M.; Beach, N.M.; Halbur, P.G.; Meng, X.J. A live-attenuated chimeric porcine circovirus type 2 (PCV2) vaccine is transmitted to contact pigs but is not upregulated by concurrent infection with porcine parvovirus (PPV) and porcine reproductive and respiratory syndrome virus (PRRSV) and is efficacious in a PCV2b-PRRSV-PPV challenge model. Clin. Vaccine Immunol. 2011, 18, 1261–1268. [Google Scholar] [CrossRef]
- Streck, A.F.; Hergemöller, F.; Rüster, D.; Speck, S.; Truyen, U. A TaqMan qPCR for quantitation of Ungulate protoparvovirus 1 validated in several matrices. J. Virol. Methods 2015, 218, 46–50. [Google Scholar] [CrossRef]
- Wilhelm, S.; Zimmermann, P.; Selbitz, H.J.; Truyen, U. Real-time PCR protocol for the detection of porcine parvovirus in field samples. J. Virol. Methods 2006, 134, 257–260. [Google Scholar] [CrossRef]
- Opriessnig, T.; Yu, S.; Gallup, J.M.; Evans, R.B.; Fenaux, M.; Pallares, F.; Thacker, E.L.; Brockus, C.W.; Ackermann, M.R.; Thomas, P.; et al. Effect of vaccination with selective bacterins on conventional pigs infected with type 2 porcine circovirus. Vet. Pathol. 2003, 40, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Palinski, R.; Piñeyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B.M. A Novel Porcine Circovirus Distantly Related to Known Circoviruses Is Associated with Porcine Dermatitis and Nephropathy Syndrome and Reproductive Failure. J. Virol. 2017, 91, e01879-16. [Google Scholar] [CrossRef] [PubMed]
- Pérez, L.J.; Díaz de Arce, H. A RT-PCR assay for the detection of encephalomycarditis virus infections in pigs. Braz. J. Microbiol. 2009, 40, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Palmquist, J.M.; Munir, S.; Taku, A.; Kapur, V.; Goyal, S.M. Detection of porcine teschovirus and enterovirus type II by reverse transcription-polymerase chain reaction. J. Vet. Diagn. Investig. 2002, 14, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Letocart, M.; Baranton, G.; Perolat, P. Rapid identification of pathogenic Leptospira species (Leptospira interrogans, L. borgpetersenii, and L. kirschneri) with species-specific DNA probes produced by arbitrarily primed PCR. J. Clin. Microbiol. 1997, 35, 248–253. [Google Scholar] [CrossRef]
- Blumer, S.; Greub, G.; Waldvogel, A.; Hässig, M.; Thoma, R.; Tschuor, A.; Pospischil, A.; Borel, N. Waddlia, Parachlamydia and Chlamydiaceae in bovine abortion. Vet. Microbiol. 2011, 152, 385–393. [Google Scholar] [CrossRef]
- Fetzer, C.; Pesch, S.; Ohlinger, V.F. High risk of false positive results in a widely used diagnostic test for detection of the porcine reproductive and respiratory syndrome virus (PRRSV). Vet. Microbiol. 2006, 115, 21–31. [Google Scholar] [CrossRef]
- Lagumdzic, E.; Pernold, C.P.S.; Ertl, R.; Palmieri, N.; Stadler, M.; Sawyer, S.; Stas, M.R.; Kreutzmann, H.; Rümenapf, T.; Ladinig, A.; et al. Gene expression of peripheral blood mononuclear cells and CD8+ T cells from gilts after PRRSV infection. Front. Immunol. 2023, 14, 1159970. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Karniychuk, U.U.; Nauwynck, H.J. Pathogenesis and prevention of placental and transplacental porcine reproductive and respiratory syndrome virus infection. Vet. Res. 2013, 44, 95. [Google Scholar] [CrossRef]
- Li, X.; Galliher-Beckley, A.; Pappan, L.; Trible, B.; Kerrigan, M.; Beck, A.; Hesse, R.; Blecha, F.; Nietfeld, J.C.; Rowland, R.R.; et al. Comparison of host immune responses to homologous and heterologous type II porcine reproductive and respiratory syndrome virus (PRRSV) challenge in vaccinated and unvaccinated pigs. Biomed Res. Int. 2014, 2014, 416727. [Google Scholar] [CrossRef] [PubMed]
- Martelli, P.; Gozio, S.; Ferrari, L.; Rosina, S.; De Angelis, E.; Quintavalla, C.; Bottarelli, E.; Borghetti, P. Efficacy of a modified live porcine reproductive and respiratory syndrome virus (PRRSV) vaccine in pigs naturally exposed to a heterologous European (Italian cluster) field strain: Clinical protection and cell-mediated immunity. Vaccine 2009, 27, 3788–3799. [Google Scholar] [CrossRef] [PubMed]
- Murtaugh, M.P.; Genzow, M. Immunological solutions for treatment and prevention of porcine reproductive and respiratory syndrome (PRRS). Vaccine 2011, 29, 8192–8204. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Kang, I.; Kim, S.; Park, S.-J.; Park, K.H.; Oh, T.; Yang, S.; Chae, C. A modified-live porcine reproductive and respiratory syndrome virus (PRRSV)-1 vaccine protects late-term pregnancy gilts against heterologous PRRSV-1 but not PRRSV-2 challenge. Transbound. Emerg. Dis. 2018, 65, 1227–1234. [Google Scholar] [CrossRef]
- Bitsouni, V.; Lycett, S.; Opriessnig, T.; Doeschl-Wilson, A. Predicting vaccine effectiveness in livestock populations: A theoretical framework applied to PRRS virus infections in pigs. PLoS ONE 2019, 14, e0220738. [Google Scholar] [CrossRef]




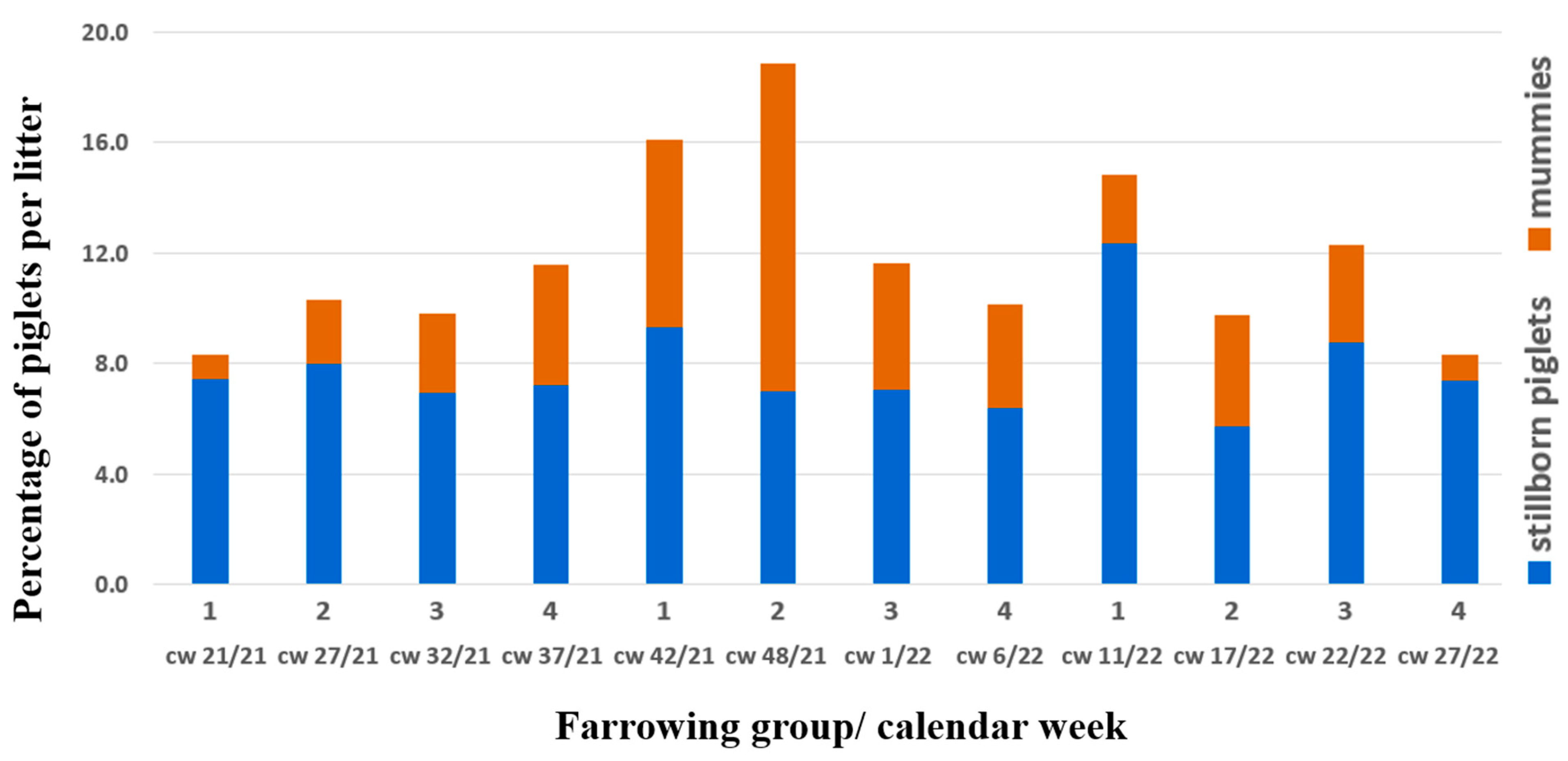{kind=link}
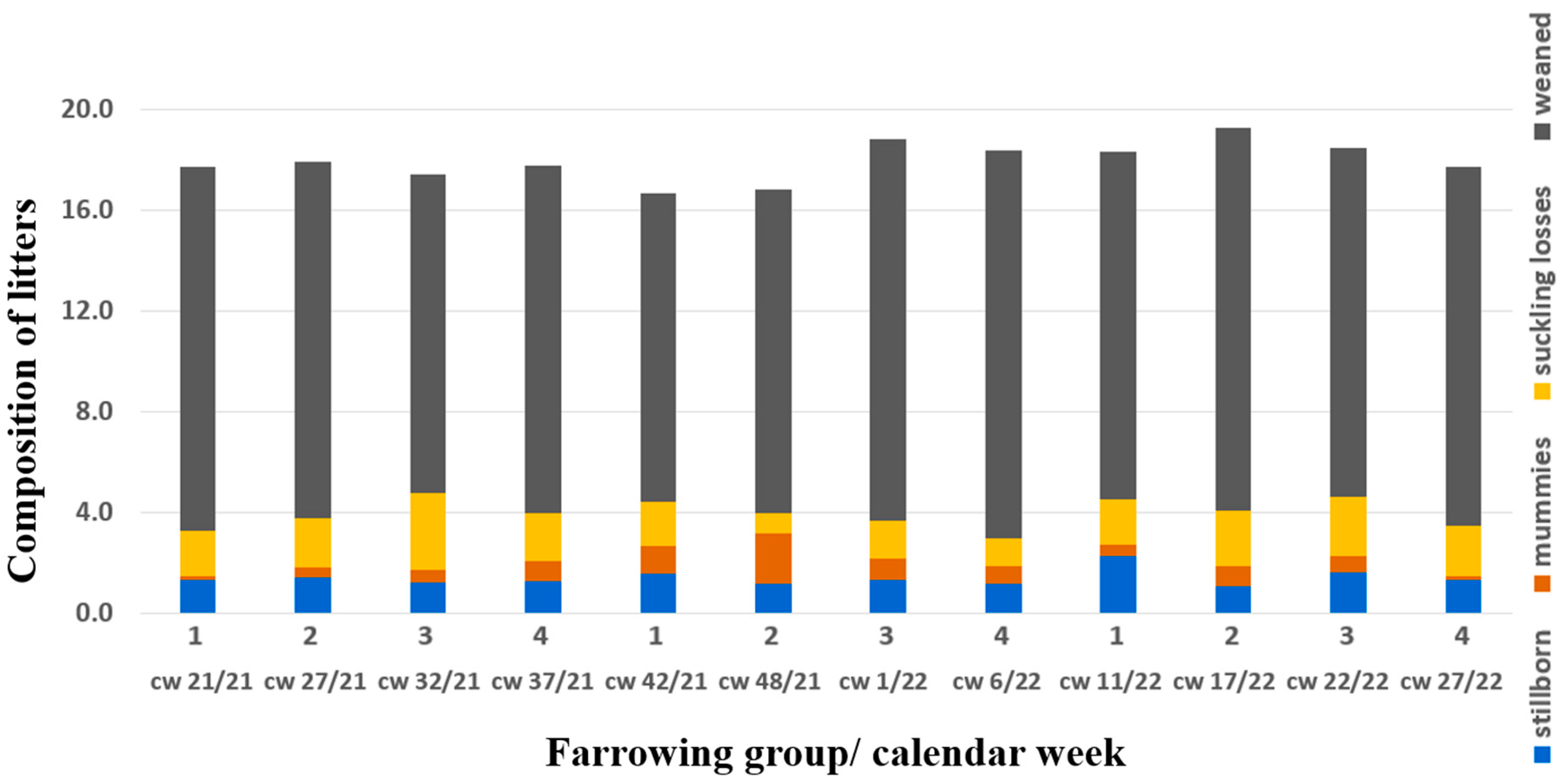{kind=link}
{kind=link}
| Sow ID | Parity | Nr of Total PRRSV Vaccinations * | Days of Gestation | Composition of Following Litters ** | ||
|---|---|---|---|---|---|---|
| Live-Born | Mummies | Stillborns | ||||
| Litter A | 3 | 6 | 115 | 20 | 2 | 1 |
| Litter B | 2 | 5 | 115 | 18 | 0 | 2 |
| Litter C | 5 | 8 | 116 | 17 | 1 | 2 |
| Litter | Crown–Rump Length in mm | Nr of Live-Borns * | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | 280 | 255 | 250 | 265 | 230 | 210 | 130 | 100 | 8 | ||||||||
| B | 265 | 245 | 240 | 200 | 185 | 175 | 135 | 135 | 130 | 125 | 110 | 95 | 90 | 70 | 60 | 60 | 6 |
| C | 300 | 290 | 255 | 220 | 220 | 210 | 205 | 200 | 150 | 130 | 120 | 110 | 80 | 80 | 5 | ||
| Pathogen | Tested Organ | Method | Result | PCR Assay According to |
|---|---|---|---|---|
| PPV1 PPV in general | Placenta, liver, lung, kidney and umbilical cord | PCR | Negative | [22] |
| PCR | Negative | [23] | ||
| qPCR | Negative | [24] | ||
| Virus isolation | Negative | |||
| PTV, PEV | Placenta, liver, lung, kidney and umbilical cord | RT-PCR | Negative | [28] |
| PCV2 | Heart | qPCR | Negative | [25] |
| PCV3 | Heart | qPCR | Negative | [26] |
| EMCV | Heart | RT-PCR | Negative | [27] |
| Leptospira interrogans | Lung, liver, kidney, placenta | PCR | Negative | [29] |
| Chlamydia spp. | Lung, liver, kidney, placenta | qPCR | Negative | [30] |
| PRRSV | Thymus | RT-PCR | Positive | [31] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Unterweger, C.; Kreutzmann, H.; Buenger, M.; Klingler, E.; Auer, A.; Rümenapf, T.; Truyen, U.; Ladinig, A. Litters of Various-Sized Mummies (LVSM) and Stillborns after Porcine Reproductive and Respiratory Syndrome Virus Type 1 Infection—A Case Report. Vet. Sci. 2023, 10, 494. https://doi.org/10.3390/vetsci10080494
Unterweger C, Kreutzmann H, Buenger M, Klingler E, Auer A, Rümenapf T, Truyen U, Ladinig A. Litters of Various-Sized Mummies (LVSM) and Stillborns after Porcine Reproductive and Respiratory Syndrome Virus Type 1 Infection—A Case Report. Veterinary Sciences. 2023; 10(8):494. https://doi.org/10.3390/vetsci10080494
Chicago/Turabian StyleUnterweger, Christine, Heinrich Kreutzmann, Moritz Buenger, Eva Klingler, Angelika Auer, Till Rümenapf, Uwe Truyen, and Andrea Ladinig. 2023. "Litters of Various-Sized Mummies (LVSM) and Stillborns after Porcine Reproductive and Respiratory Syndrome Virus Type 1 Infection—A Case Report" Veterinary Sciences 10, no. 8: 494. https://doi.org/10.3390/vetsci10080494
APA StyleUnterweger, C., Kreutzmann, H., Buenger, M., Klingler, E., Auer, A., Rümenapf, T., Truyen, U., & Ladinig, A. (2023). Litters of Various-Sized Mummies (LVSM) and Stillborns after Porcine Reproductive and Respiratory Syndrome Virus Type 1 Infection—A Case Report. Veterinary Sciences, 10(8), 494. https://doi.org/10.3390/vetsci10080494