

A Review of Effect of Saponins on Ruminal Fermentation, Health and Performance of Ruminants


Abstract
Simple Summary
Abstract
1. Introduction
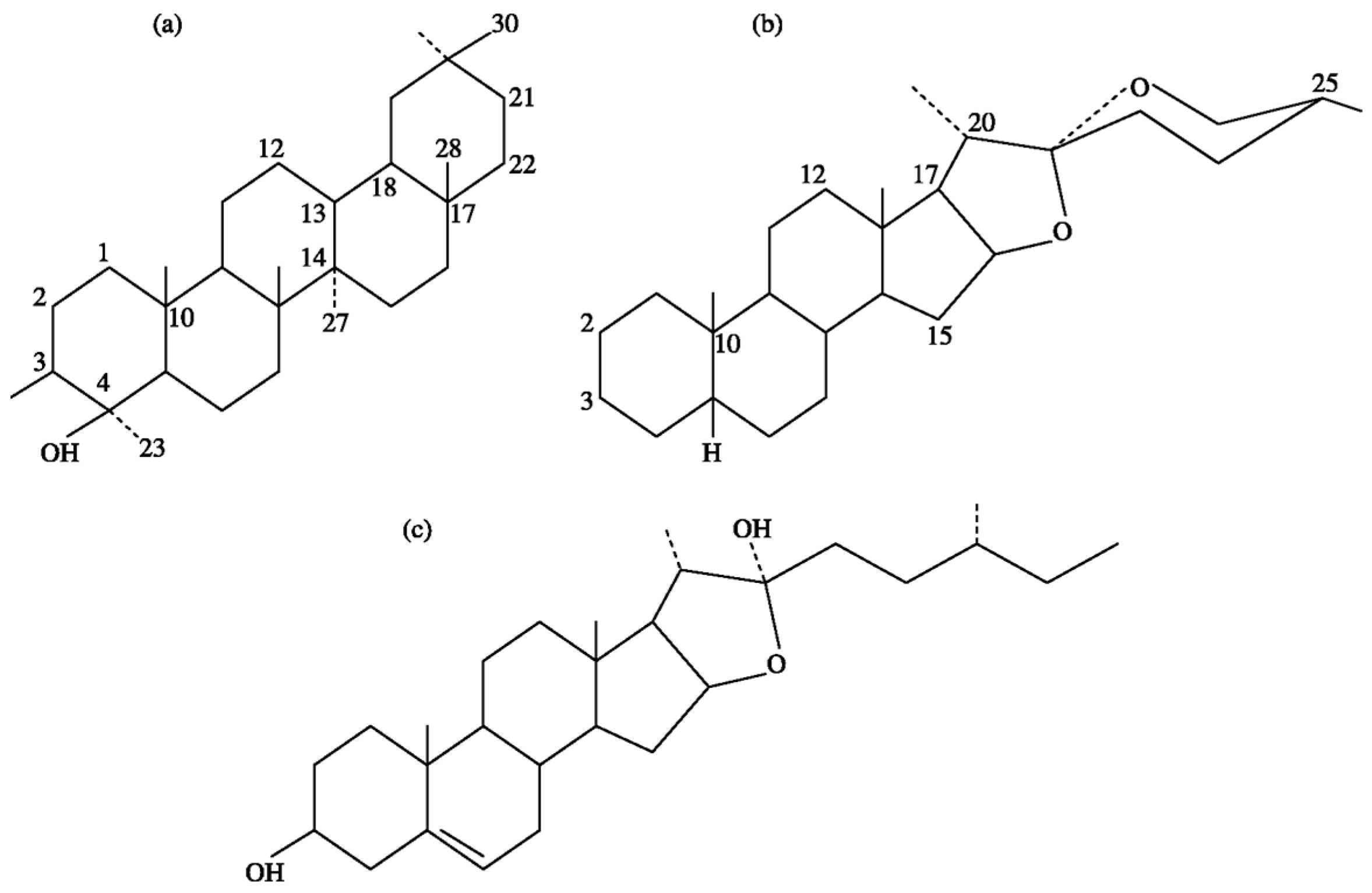
2. Chemistry of Saponins
3. Occurrence and Roles of Saponins in Plants
4. Saponins and Rumen Microbiota Population
4.1. Ciliate Protozoa
4.2. Bacteria
4.3. Fungi
4.4. Archaea
5. Microbial Adaptation and Saponins Metabolism
6. Ruminal Degradation of Saponins
7. Effects of Saponins on Feed Digestion and Rumen Fermentation
7.1. Ruminal Enzyme Activity and Digestion of Feeds
7.2. Microbial Protein Synthesis
7.3. Ammonia-N and N Utilization
7.4. Volatile Fatty Acids
7.5. Gas and Methane Productions
8. Effects of Saponins on Blood Parameters
9. Effects of Saponins on Animal Performance
9.1. Growth Performance
9.2. Lactation Performance
10. Saponins as Anthelmintics
11. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kholif, A.E.; Olafadehan, O.A. Essential Oils and Phytogenic Feed Additives in Ruminant Diet: Chemistry, Ruminal Microbiota and Fermentation, Feed Utilization and Productive Performance. Phytochem. Rev. 2021, 20, 1087–1108. [Google Scholar] [CrossRef]
- Ramdani, D.; Yuniarti, E.; Jayanegara, A.; Chaudhry, A.S. Roles of Essential Oils, Polyphenols, and Saponins of Medicinal Plants as Natural Additives and Anthelmintics in Ruminant Diets: A Systematic Review. Animals 2023, 13, 767. [Google Scholar] [CrossRef] [PubMed]
- Khattab, M.S.A.; Kholif, A.E.; Abd El Tawab, A.M.; Shaaban, M.M.; Hadhoud, F.I.; El-Fouly, H.A.; Olafadehan, O.A. Effect of Replacement of Antibiotics with Thyme and Celery Seed Mixture on the Feed Intake and Digestion, Ruminal Fermentation, Blood Chemistry, and Milk Lactation of Lactating Barki Ewes. Food Funct. 2020, 11, 6889–6898. [Google Scholar] [CrossRef]
- Shaaban, M.M.; Kholif, A.E.; Abd El Tawab, A.M.; Radwan, M.A.; Hadhoud, F.I.; Khattab, M.S.A.; Saleh, H.M.; Anele, U.Y. Thyme and Celery as Potential Alternatives to Ionophores Use in Livestock Production: Their Effects on Feed Utilization, Growth Performance and Meat Quality of Barki Lambs. Small Rumin. Res. 2021, 200, 106400. [Google Scholar] [CrossRef]
- Wanapat, M.; Phesatcha, K.; Viennasay, B.; Phesatcha, B.; Ampapon, T.; Kang, S. Strategic Supplementation of Cassava Top Silage to Enhance Rumen Fermentation and Milk Production in Lactating Dairy Cows in the Tropics. Trop. Anim. Health Prod. 2018, 50, 1539–1546. [Google Scholar] [CrossRef]
- Kim, H.; Lee, H.G.; Baek, Y.C.; Lee, S.; Seo, J. The Effects of Dietary Supplementation with 3-Nitrooxypropanol on Enteric Methane Emissions, Rumen Fermentation, and Production Performance in Ruminants: A Meta-Analysis. J. Anim. Sci. Technol. 2020, 62, 31–42. [Google Scholar] [CrossRef]
- Gunun, P.; Gunun, N.; Khejornsart, P.; Ouppamong, T.; Cherdthong, A.; Wanapat, M.; Sirilaophaisan, S.; Yuangklang, C.; Polyorach, S.; Kenchaiwong, W.; et al. Effects of Antidesma thwaitesianum Muell. Arg. Pomace as a Source of Plant Secondary Compounds on Digestibility, Rumen Environment, Hematology, and Milk Production in Dairy Cows. Anim. Sci. J. 2019, 90, 372–381. [Google Scholar] [CrossRef]
- Demirtaş, A.; Öztürk, H.; Pişkin, I. Overview of Plant Extracts and Plant Secondary Metabolites as Alternatives to Antibiotics for Modification of Ruminal Fermentation. Ank. Üniversitesi Vet. Fakültesi Derg. 2018, 65, 213–217. [Google Scholar] [CrossRef]
- Patra, A.K.; Saxena, J. The Effect and Mode of Action of Saponins on the Microbial Populations and Fermentation in the Rumen and Ruminant Production. Nutr. Res. Rev. 2009, 22, 204–219. [Google Scholar] [CrossRef]
- Kozłowska, M.; Cieślak, A.; Jóźwik, A.; El-Sherbiny, M.; Stochmal, A.; Oleszek, W.; Kowalczyk, M.; Filipiak, W.; Szumacher-Strabel, M. The Effect of Total and Individual Alfalfa Saponins on Rumen Methane Production. J. Sci. Food Agric. 2020, 100, 1922–1930. [Google Scholar] [CrossRef]
- Jafari, S.; Ebrahimi, M.; Goh, Y.M.; Rajion, M.A.; Jahromi, M.F.; Al-Jumaili, W.S. Manipulation of Rumen Fermentation and Methane Gas Production by Plant Secondary Metabolites (Saponin, Tannin and Essential Oil)—A Review of Ten-Year Studies. Ann. Anim. Sci. 2019, 19, 3–29. [Google Scholar] [CrossRef]
- Fleck, J.D.; Betti, A.H.; da Silva, F.P.; Troian, E.A.; Olivaro, C.; Ferreira, F.; Verza, S.G. Saponins from Quillaja saponaria and Quillaja brasiliensis: Particular Chemical Characteristics and Biological Activities. Molecules 2019, 24, 171. [Google Scholar] [CrossRef]
- Ridla, M.; Laconi, E.B.; Jayanegara, A. Effects of Saponin on Enteric Methane Emission and Nutrient Digestibility of Ruminants: An in Vivo Meta-Analysis. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Surakarta, Indonesia, 24–25 August 2021; Volume 788, p. 012028. [Google Scholar]
- Jayanegara, A.; Yogianto, Y.; Wina, E.; Sudarman, A.; Kondo, M.; Obitsu, T.; Kreuzer, M. Combination Effects of Plant Extracts Rich in Tannins and Saponins as Feed Additives for Mitigating In Vitro Ruminal Methane and Ammonia Formation. Animals 2020, 10, 1531. [Google Scholar] [CrossRef]
- Ashour, A.S.; El Aziz, M.M.A.; Gomha Melad, A.S. A Review on Saponins from Medicinal Plants: Chemistry, Isolation, and Determination. J. Nanomed. Res. 2019, 7, 282–288. [Google Scholar] [CrossRef]
- Tedeschi, L.O.; Muir, J.P.; Naumann, H.D.; Norris, A.B.; Ramírez-Restrepo, C.A.; Mertens-Talcott, S.U. Nutritional Aspects of Ecologically Relevant Phytochemicals in Ruminant Production; Frontiers Media S.A.: Lausanne, Switzerland, 2021; Volume 8, p. 155. [Google Scholar]
- Awulachew, M.T. A Review of Anti-Nutritional Factors in Plant Based Foods. Adv. Nutr. Food Sci. 2021, 7, 223–236. [Google Scholar] [CrossRef]
- Carter, J.P.; Spink, J.; Cannon, P.F.; Daniels, M.J.; Osbourn, A.E. Isolation, Characterization, and Avenacin Sensitivity of a Diverse Collection of Cereal-Root-Colonizing Fungi. Appl Environ Microbiol 1999, 65, 3364–3372. [Google Scholar] [CrossRef]
- Rupasinghe, H.P.V.; Jackson, C.J.C.; Poysa, V.; Di Berardo, C.; Bewley, J.D.; Jenkinson, J. Soyasapogenol A and B Distribution in Soybean (Glycine max L. Merr.) in Relation to Seed Physiology, Genetic Variability, and Growing Location. J. Agric. Food Chem. 2003, 51, 5888–5894. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Morales, E.; de la Fuente, G.; Nash, R.J.; Braganca, R.; Duval, S.; Bouillon, M.E.; Lahmann, M.; Newbold, C.J. Improving the Antiprotozoal Effect of Saponins in the Rumen by Combination with Glycosidase Inhibiting Iminosugars or by Modification of Their Chemical Structure. PLoS ONE 2017, 12, e0184517. [Google Scholar] [CrossRef]
- Belanche, A.; Pinloche, E.; Preskett, D.; Newbold, C.J. Effects and Mode of Action of Chitosan and Ivy Fruit Saponins on the Microbiome, Fermentation and Methanogenesis in the Rumen Simulation Technique. FEMS Microbiol. Ecol. 2016, 92, fiv160. [Google Scholar] [CrossRef] [PubMed]
- Newbold, C.J.; de la Fuente, G.; Belanche, A.; Ramos-Morales, E.; McEwan, N.R. The Role of Ciliate Protozoa in the Rumen. Front. Microbiol. 2015, 6, 1313. [Google Scholar] [CrossRef] [PubMed]
- Weimer, P.J. Degradation of Cellulose and Hemicellulose by Ruminal Microorganisms. Microorganisms 2022, 10, 2345. [Google Scholar] [CrossRef] [PubMed]
- Gunun, P.; Cherdthong, A.; Khejornsart, P.; Wanapat, M.; Polyorach, S.; Kang, S.; Kaewwongsa, W.; Gunun, N. The Effect of Phytonutrients in Terminalia chebula Retz. on Rumen Fermentation Efficiency, Nitrogen Utilization, and Protozoal Population in Goats. Animals 2022, 12, 2022. [Google Scholar] [CrossRef] [PubMed]
- Widyarini, S.; Nagari, F.S.; Hanim, C.; Bachruddin, Z.; Muhlisin, M.; Mira Yusiati, L. Effect of Nigella sativa L. as Saponin Sources on In Vitro Rumen Fermentation, Enzyme Activity and Nutrients Digestibility. Adv. Anim. Vet. Sci. 2021, 9, 2247–2257. [Google Scholar] [CrossRef]
- Guo, Y.Q.; Liu, J.X.; Lu, Y.; Zhu, W.Y.; Denman, S.E.; McSweeney, C.S. Effect of Tea Saponin on Methanogenesis, Microbial Community Structure and Expression of McrA Gene, in Cultures of Rumen Micro-Organisms. Lett. Appl. Microbiol. 2008, 47, 421–426. [Google Scholar] [CrossRef]
- Hu, W.L.; Liu, J.X.; Ye, J.A.; Wu, Y.M.; Guo, Y.Q. Effect of Tea Saponin on Rumen Fermentation In Vitro. Anim. Feed Sci. Technol. 2005, 120, 333–339. [Google Scholar] [CrossRef]
- Wallace, R.J.; Arthaud, L.; Newbold, C.J. Influence of Yucca shidigera Extract on Ruminal Ammonia Concentrations and Ruminal Microorganisms. Appl. Environ. Microbiol. 1994, 60, 1762–1767. [Google Scholar] [CrossRef] [PubMed]
- Hristov, A.N.; Ivan, M.; Neill, L.; McAllister, T.A. Evaluation of Several Potential Bioactive Agents for Reducing Protozoal Activity in Vitro. Anim Feed Sci Technol 2003, 105, 163–184. [Google Scholar] [CrossRef]
- Kim, H.; Kim, B.-W.; Yoo, D.; Moon, J.; Kwon, I.; Lee, Y.; Seo, J. In Vitro Evaluation of Aloe saponaria as a Potential Feed Additive to Modulate Ruminal Fermentation and Microbial Diversity. J. Appl. Anim. Res. 2023, 51, 115–122. [Google Scholar] [CrossRef]
- Santoso, B.; Kilmaskossu, A.; Sambodo, P. Effects of Saponin from Biophytum Petersianum Klotzsch on Ruminal Fermentation, Microbial Protein Synthesis and Nitrogen Utilization in Goats. Anim. Feed Sci. Technol. 2007, 137, 58–68. [Google Scholar] [CrossRef]
- Moheghi, M.M.; Ghoryar, M.A.; Ataei, A.H. The Impact of Saponin and Tannin on Performance, Blood Constituents, and Carcass Fatty Acids in Baluchi Lambs. J. Livest. Sci. Technol. 2022, 10, 11–18. [Google Scholar] [CrossRef]
- Holtshausen, L.; Chaves, A.V.; Beauchemin, K.A.; McGinn, S.M.; McAllister, T.A.; Odongo, N.E.; Cheeke, P.R.; Benchaar, C. Feeding Saponin-Containing Yucca schidigera and Quillaja saponaria to Decrease Enteric Methane Production in Dairy Cows. J. Dairy Sci. 2009, 92, 2809–2821. [Google Scholar] [CrossRef]
- Singer, M.D.; Robinson, P.H.; Salem, A.Z.M.; De Peters, E.J. Impacts of Rumen Fluid Modified by Feeding Yucca schidigera to Lactating Dairy Cows on In Vitro Gas Production of 11 Common Dairy Feedstuffs, as Well as Animal Performance. Anim. Feed Sci. Technol. 2008, 146, 242–258. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, T.; Chen, D.; Zhang, N.; Si, B.; Deng, K.; Tu, Y.; Diao, Q. Effects of Tea Saponin Supplementation on Nutrient Digestibility, Methanogenesis, and Ruminal Microbial Flora in Dorper Crossbred Ewe. Animals 2019, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Lovett, D.K.; Stack, L.; Lovell, S.; Callan, J.; Flynn, B.; Hawkins, M.; O’Mara, F.P. Effect of Feeding Yucca schidigera Extract on Performance of Lactating Dairy Cows and Ruminal Fermentation Parameters in Steers. Livest. Sci. 2006, 102, 23–32. [Google Scholar] [CrossRef]
- Wang, C.J.; Wang, S.P.; Zhou, H. Influences of Flavomycin, Ropadiar, and Saponin on Nutrient Digestibility, Rumen Fermentation, and Methane Emission from Sheep. Anim. Feed Sci. Technol. 2009, 148, 157–166. [Google Scholar] [CrossRef]
- Hussain, I.; Cheeke, P.R.R. Effect of Dietary Yucca schidigera Extract on Rumen and Blood Profiles of Steers Fed Concentrate- or Roughage-Based Diets. Anim. Feed Sci. Technol. 1995, 51, 231–242. [Google Scholar] [CrossRef]
- Benchaar, C.; McAllister, T.A.; Chouinard, P.Y. Digestion, Ruminal Fermentation, Ciliate Protozoal Populations, and Milk Production from Dairy Cows Fed Cinnamaldehyde, Quebracho Condensed Tannin, or Yucca schidigera Saponin Extracts. J. Dairy Sci. 2008, 91, 4765–4777. [Google Scholar] [CrossRef]
- Hristov, A.N.; McAllister, T.A.; van Herk, F.H.; Cheng, K.J.; Newbold, C.J.; Cheeke, P.R. Effect of Yucca schidigera on Ruminal Fermentation and Nutrient Digestion in Heifers. J. Anim. Sci. 1999, 77, 2554–2563. [Google Scholar] [CrossRef]
- Lu, C.D.; Jorgensen, N.A. Alfalfa Saponins Affect Site and Extent of Nutrient Digestion in Ruminants. J. Nutr. 1987, 117, 919–927. [Google Scholar] [CrossRef]
- Hess, H.D.; Kreuzer, M.; Díaz, T.E.; Lascano, C.E.; Carulla, J.E.; Soliva, C.R.; Machmüller, A. Saponin Rich Tropical Fruits Affect Fermentation and Methanogenesis in Faunated and Defaunated Rumen Fluid. Anim. Feed Sci. Technol. 2003, 109, 79–94. [Google Scholar] [CrossRef]
- Soltan, Y.A.; Patra, A.K. Ruminal Microbiome Manipulation to Improve Fermentation Efficiency in Ruminants. In Animal Feed Science and Nutrition—Production, Health and Environment; Patra, A.K., Payan-Carreira, R., Eds.; IntechOpen Publisher: London, UK, 2022; pp. 1–20. [Google Scholar]
- Odenyo, A.A.; Osuji, P.O.; Karanfil, O. Effect of Multipurpose Tree (MPT) Supplements on Ruminal Ciliate Protozoa. Anim. Feed Sci. Technol. 1997, 67, 169–180. [Google Scholar] [CrossRef]
- Newbold, C.J.; El Hassan, S.M.; Wang, J.; Ortega, M.E.; Wallace, R.J. Influence of Foliage from African Multipurpose Trees on Activity of Rumen Protozoa and Bacteria. Br. J. Nutr. 1997, 78, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Hua, D.; Hendriks, W.H.; Xiong, B.; Pellikaan, W.F. Starch and Cellulose Degradation in the Rumen and Applications of Metagenomics on Ruminal Microorganisms. Animals 2022, 12, 3020. [Google Scholar] [CrossRef] [PubMed]
- Hess, H.D.; Beuret, R.A.; Lötscher, M.; Hindrichsen, I.K.; Machmüller, A.; Carulla, J.E.; Lascano, C.E.; Kreuzer, M. Ruminal Fermentation, Methanogenesis and Nitrogen Utilization of Sheep Receiving Tropical Grass Hay-Concentrate Diets Offered with Sapindus saponaria Fruits and Cratylia argentea Foliage. Anim. Sci. 2004, 79, 177–189. [Google Scholar] [CrossRef]
- Wang, Y.; McAllister, T.A.; Yanke, L.J.; Cheeke, P.R. Effect of Steroidal Saponin from Yucca schidigera Extract on Ruminal Microbes. J. Appl. Microbiol. 2000, 88, 887–896. [Google Scholar] [CrossRef]
- Wang, B.; Ma, M.P.; Diao, Q.Y.; Tu, Y. Saponin-Induced Shifts in the Rumen Microbiome and Metabolome of Young Cattle. Front. Microbiol. 2019, 10, 356. [Google Scholar] [CrossRef]
- Avato, P.; Bucci, R.; Tava, A.; Vitali, C.; Rosato, A.; Bialy, Z.; Jurzysta, M. Antimicrobial Activity of Saponins from Medicago sp.: Structure-Activity Relationship. Phytother. Res. 2006, 20, 454–457. [Google Scholar] [CrossRef]
- Muetzel, S. Supplementation of Barley Straw with Sesbania pachycarpa Leaves In Vitro: Effects on Fermentation Variables and Rumen Microbial Population Structure Quantified by Ribosomal RNA-Targeted Probes. Br. J. Nutr. 2003, 89, 445–453. [Google Scholar] [CrossRef]
- Li, Y.; Du, Y.; Zou, C. Effects of pH on Antioxidant and Antimicrobial Properties of Tea Saponins. Eur. Food Res. Technol. 2009, 228, 1023–1028. [Google Scholar] [CrossRef]
- Wina, E.; Muetzel, S.; Becker, K. The Dynamics of Major Fibrolytic Microbes and Enzyme Activity in the Rumen in Response to Short- and Long-Term Feeding of Sapindus rarak Saponins. J. Appl. Microbiol. 2006, 100, 114–122. [Google Scholar] [CrossRef]
- Wina, E.; Muetzel, S.; Hoffmann, E.; Makkar, H.P.S.; Becker, K. Saponins Containing Methanol Extract of Sapindus rarak Affect Microbial Fermentation, Microbial Activity and Microbial Community Structure In Vitro. Anim. Feed Sci. Technol. 2005, 121, 159–174. [Google Scholar] [CrossRef]
- Barile, E.; Bonanomi, G.; Antignani, V.; Zolfaghari, B.; Sajjadi, S.E.; Scala, F.; Lanzotti, V. Saponins from Allium minutiflorum with Antifungal Activity. Phytochemistry 2007, 68, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Oleszek, W.A. Saponins. Natural Food Antimicrobial Systems; Naidu, A., Ed.; CRC Press: Boca Raton, FL, USA, 2000; pp. 295–324. ISBN 978-0-8493-2047-7. [Google Scholar]
- Liwiński, B.J.; Soliva, C.R.; Machmüller, A.; Kreuzer, M. Efficacy of Plant Extracts Rich in Secondary Constituents to Modify Rumen Fermentation. Anim. Feed Sci. Technol. 2002, 101, 101–114. [Google Scholar] [CrossRef]
- Śliwiński, B.J.; Kreuzer, M.; Wettstein, H.-R.; Machmüller, A. Rumen Fermentation and Nitrogen Balance of Lambs Fed Diets Containing Plant Extracts Rich in Tannins and Saponins, and Associated Emissions of Nitrogen and Methane. Arch. Anim. Nutr. 2002, 56, 379–392. [Google Scholar] [CrossRef]
- Lila, Z.A.; Mohammed, N.; Kanda, S.; Kamada, T.; Itabashi, H. Effect of Sarsaponin on Ruminal Fermentation with Particular Reference to Methane Production In Vitro. J. Dairy Sci. 2003, 86, 3330–3336. [Google Scholar] [CrossRef]
- Meagher, L.P.; Smith, B.L.; Wilkins, A.L. Metabolism of Diosgenin-Derived Saponins: Implications for Hepatogenous Photosensitization Diseases in Ruminants. Anim. Feed Sci. Technol. 2001, 91, 157–170. [Google Scholar] [CrossRef]
- Flåøyen, A.; Wilkins, A.L. Metabolism of Saponins from Narthecium ossifragum—A Plant Implicated in the Aetiology of Alveld, a Hepatogenous Photosensitization of Sheep. Vet. Res. Commun. 1997, 21, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Wisløff, H.; Wilkins, A.L.; Scheie, E.; Flåøyen, A. Accumulation of Sapogenin Conjugates and Histological Changes in the Liver and Kidneys of Lambs Suffering from Alveld, a Hepatogenous Photosensitization Disease of Sheep Grazing Narthecium ossifragum. Vet. Res. Commun. 2002, 26, 381–396. [Google Scholar] [CrossRef]
- Wang, Y.; McAllister, T.A.; Newbold, C.J.; Rode, L.M.; Cheeke, P.R.; Cheng, K.J. Effects of Yucca schidigera Extract on Fermentation and Degradation of Steroidal Saponins in the Rumen Simulation Technique (RUSITEC). Anim Feed Sci Technol 1998, 74, 143–153. [Google Scholar] [CrossRef]
- Patel, S. Role of Rumen Protozoa: Metabolic and Fibrolytic. Adv. Biotechnol. Microbiol. 2018, 10, 1–6. [Google Scholar] [CrossRef]
- Hanh, T.T.H.; Dang, N.H.; Dat, N.T. α-Amylase and α-Glucosidase Inhibitory Saponins from Polyscias fruticosa Leaves. J. Chem. 2016, 2016, 1–5. [Google Scholar] [CrossRef]
- Samtiya, M.; Aluko, R.E.; Dhewa, T. Plant Food Anti-Nutritional Factors and Their Reduction Strategies: An Overview. Food Prod. Process. Nutr. 2020, 2, 1–14. [Google Scholar] [CrossRef]
- Klita, P.T.; Mathison, G.W.; Fenton, T.W.; Hardin, R.T. Effects of Alfalfa Root Saponins on Digestive Function in Sheep. J. Anim. Sci. 1996, 74, 1144–1156. [Google Scholar] [CrossRef] [PubMed]
- Taiwo, G.; Sidney, T.; Idowu, M.; Eichie, F.; Karnezos, T.P.; Ogunade, I.M. Dietary Fenugreek Seed Extract Improves Dry Matter Intake, Apparent Total Tract Nutrient Digestibility, and Alters Whole Blood Transcriptome of Holstein Dairy Heifers. Transl. Anim. Sci. 2022, 6, 1–7. [Google Scholar] [CrossRef]
- Emerson, E.L.; Weimer, P.J. Fermentation of Model Hemicelluloses by Prevotella Strains and Butyrivibrio Fibrisolvens in Pure Culture and in Ruminal Enrichment Cultures. Appl. Microbiol. Biotechnol. 2017, 101, 4269–4278. [Google Scholar] [CrossRef]
- Supapong, C.; Cherdthong, A.; Seankamsorn, A.; Khonkhaeng, B.; Wanapat, M.; Gunun, N.; Gunun, P.; Chanjula, P.; Polyorach, S. Effect of Delonix regia Seed Meal Supplementation in Thai Native Beef Cattle on Feed Intake, Rumen Fermentation Characteristics and Methane Production. Anim. Feed Sci. Technol. 2017, 232, 40–48. [Google Scholar] [CrossRef]
- Cardozo, P.W.; Calsamiglia, S.; Ferret, A.; Kamel, C. Screening for the Effects of Natural Plant Extracts at Different pH on In Vitro Rumen Microbial Fermentation of a High-Concentrate Diet for Beef Cattle. J. Anim. Sci. 2005, 83, 2572–2579. [Google Scholar] [CrossRef]
- Samal, L.; Kumar Dash, S. Nutritional Interventions to Reduce Methane Emissions in Ruminants. In Animal Feed Science and Nutrition—Production, Health and Environment; Patra, A.K., Payan-Carreira, R., Eds.; IntechOpen: London, UK, 2022; pp. 1–19. ISBN 978-1-83969-861-3. [Google Scholar]
- Guyader, J.; Eugène, M.; Doreau, M.; Morgavi, D.P.P.; Gérard, C.; Martin, C. Tea Saponin Reduced Methanogenesis In Vitro but Increased Methane Yield in Lactating Dairy Cows. J. Dairy Sci. 2017, 100, 1845–1855. [Google Scholar] [CrossRef]
- Wang, X.F.; Mao, S.Y.; Liu, J.H.; Zhang, L.L.; Cheng, Y.F.; Jin, W.; Zhu, W.Y. Effect of the Gynosaponin on Methane Production and Microbe Numbers in a Fungus-Methanogen Co-Culture. J Anim Feed Sci 2011, 20, 272–284. [Google Scholar] [CrossRef]
- Goel, G.; Makkar, H.P.S. Methane Mitigation from Ruminants Using Tannins and Saponins. Trop. Anim. Health Prod. 2012, 44, 729–739. [Google Scholar] [CrossRef]
- Śliwiński, B.J.; Kreuzer, M.; Sutter, F.; Machmüller, A.; Wettstein, H.R. Performance, Body Nitrogen Conversion and Nitrogen Emission from Manure of Dairy Cows Fed Diets Supplemented with Different Plant Extracts. J. Anim. Feed Sci. 2004, 13, 73–91. [Google Scholar] [CrossRef]
- Abdullah, R.H.; Al-Galbi, H.A.J. Impact of Feeding Tea Leaves Saponins on Some Productive and Serum Biochemical Parameters of Awassi Lambs. Basrah J. Agric. Sci. 2020, 33, 17–25. [Google Scholar] [CrossRef]
- Görgülü, M.; Yurtseven, S.; Ünsal, İ.; Kutlu, H.R. Effect of Dietary Supplemental Yucca schidigera Powder on Fattening Performance of Male Lambs. J. Appl. Anim. Res. 2004, 25, 33–36. [Google Scholar] [CrossRef]
- Botura, M.B.; dos Santos, J.D.G.; da Silva, G.D.; de Lima, H.G.; de Oliveira, J.V.A.; de Almeida, M.A.O.; Batatinha, M.J.M.; Branco, A. In Vitro Ovicidal and Larvicidal Activity of Agave sisalana Perr. (Sisal) on Gastrointestinal Nematodes of Goats. Vet. Parasitol. 2013, 192, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Botura, M.B.; Silva, G.D.; Lima, H.G.; Oliveira, J.V.A.; Souza, T.S.; Santos, J.D.G.; Branco, A.; Moreira, E.L.T.; Almeida, M.A.O.; Batatinha, M.J.M. In Vivo Anthelmintic Activity of an Aqueous Extract from Sisal Waste (Agave sisalana Perr.) against Gastrointestinal Nematodes in Goats. Vet. Parasitol. 2011, 177, 104–110. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Saponin Source 2 | Animal | Dose | Feed (Forage:Concentrate Ratio) | Protozoa | Ammonia | Volatile Fatty Acid | Acetate: Propionate | Microbial Protein Synthesis | Methane | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Aloe saponaria | Holstein steers | 1 and 2% of total diet | 30.5:69.5 | NR | = | + | + | NR | = | [30] |
| Antidesma thwaitesianum | Cow | 9.8, 19.6 and 29.4 g/cow/d | 0:100 | − | − | = | − | NR | − | [7] |
| Biophytum petersianum | Goats | 13 mg/kg BW | 70:30 | − | − | − | − | = | NR | [31] |
| Biophytum petersianum | Goats | 26 mg/kg BW | 70:30 | − | − | = | − | + | NR | [31] |
| Biophytum petersianum | Goats | 19.5 mg/kg BW | 70:30 | − | − | − | − | + | NR | [31] |
| Crude saponins | Lambs | 150 mg/kg DM feed | 30:70 | NR | = | NR | NR | NR | NR | [32] |
| Medicago sativa L. | Cow | 2.4 mg | 60:40 | − | − | NR | NR | NR | − | [10] |
| Nigella sativa | Cattle | 0%, 0.2%, 0.4%, and 0.6% saponin | 100:0 | + | − | = | = | − | − | [25] |
| Quillaja saponaria plant | Dairy cows | 10 | 51:49 | = | = | = | = | NR | = | [33] |
| Sapindus rarak | Cow | 2 mg/mL medium | 30:70 | − | − | + | − | NR | − | [14] |
| Sapindus Saponaria fruit | Sheep | 5 g/kg BW0.75 | 67:33 | − | = | + | − | NR | − | [26] |
| Sarsaponin | Dairy cows | 0.2, 0.41, 0.62 | 36:64 | = | − | = | = | NR | NR | [34] |
| Tea | Sheep | 2 g/ewe/day | 68.7:31.3 | − | − | + | − | = | NR | [35] |
| Terminalia chebula | Goat | 0, 8, 16, and 24 g/kg of total DM intake | 50:50 | NR | − | = | = | NR | NR | [24] |
| Yucca schidigera extract | Steers | 2.56 | 63:37 | − | NR | − | = | NR | NR | [36] |
| Yucca schidigera extract | Sheep | 0.13 | 75:25 | NR | − | + | = | NR | − | [37] |
| Yucca schidigera extract | Steers | 0.075 | 92:8, 96:4, 45:55, 48:52 | NR | = | = | = | NR | NR | [38] |
| Yucca schidigera extract | Dairy cows | 2.8 | 40:60 | = | = | = | = | NR | NR | [39] |
| Yucca schidigera plant | Heifers | 1.96 and 5.83 | 61:39 | − | = | = | − | = | NR | [40] |
| Yucca schidigera plant | Dairy cows | 10 | 51:49 | = | − | = | = | NR | = | [33] |
| Saponin Source 2 | Animal | Dose | Feed (Forage: Concentrate Ratio) | DM Intake | Average Daily Gain | Milk Production | Reference |
|---|---|---|---|---|---|---|---|
| Antidesma thwaitesianum | Cow | 9.8, 19.6, and 29.4 g/cow/d | 0:100 | = | NR | = | [7] |
| Crude saponins | Lambs | 150 mg/kg DM | 30:70 | = | = | NR | [32] |
| Quillaja saponaria plant | Dairy cows | 10 | 51:49 | + | = | = | [33] |
| Sapindus Saponaria fruit | Sheep | 5 g/kg BW0.75 | 67:33 | = | + | NR | [26] |
| Sarsaponin | Dairy cows | 0.2, 0.41, and 0.62 | 36:64 | = | = | = | [34] |
| Tea leaves | Lambs | 180 mg/kg DM feed | NR | = | + | NR | [77] |
| Yucca schidigera extract | Dairy cows | 1.46 and 3.1 | 63:37 | = | NR | = | [36] |
| Yucca schidigera extract | Steers | 1.25 and 2.56 | 63:37 | = | NR | = | [36] |
| Yucca schidigera extract | Sheep | 0.13 | 75:25 | = | NR | NR | [37] |
| Yucca schidigera extract | Dairy cows | 2.8 | 40:60 | NR | NR | = | [39] |
| Yucca schidigera plant | Dairy cows | 10 | 51:49 | + | = | = | [33] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kholif, A.E. A Review of Effect of Saponins on Ruminal Fermentation, Health and Performance of Ruminants. Vet. Sci. 2023, 10, 450. https://doi.org/10.3390/vetsci10070450
Kholif AE. A Review of Effect of Saponins on Ruminal Fermentation, Health and Performance of Ruminants. Veterinary Sciences. 2023; 10(7):450. https://doi.org/10.3390/vetsci10070450
Chicago/Turabian StyleKholif, Ahmed E. 2023. "A Review of Effect of Saponins on Ruminal Fermentation, Health and Performance of Ruminants" Veterinary Sciences 10, no. 7: 450. https://doi.org/10.3390/vetsci10070450
APA StyleKholif, A. E. (2023). A Review of Effect of Saponins on Ruminal Fermentation, Health and Performance of Ruminants. Veterinary Sciences, 10(7), 450. https://doi.org/10.3390/vetsci10070450