
Exploring the Enteric Virome of Cats with Acute Gastroenteritis

 ,
,  , ,
, , 
 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Virological Investigation
2.3. Statistical Analysis
2.4. SISPA, ONT Library Preparation and Sequencing
3. Results
3.1. Molecular Screening for Selected Viral Pathogens
3.2. Sanger Sequencing
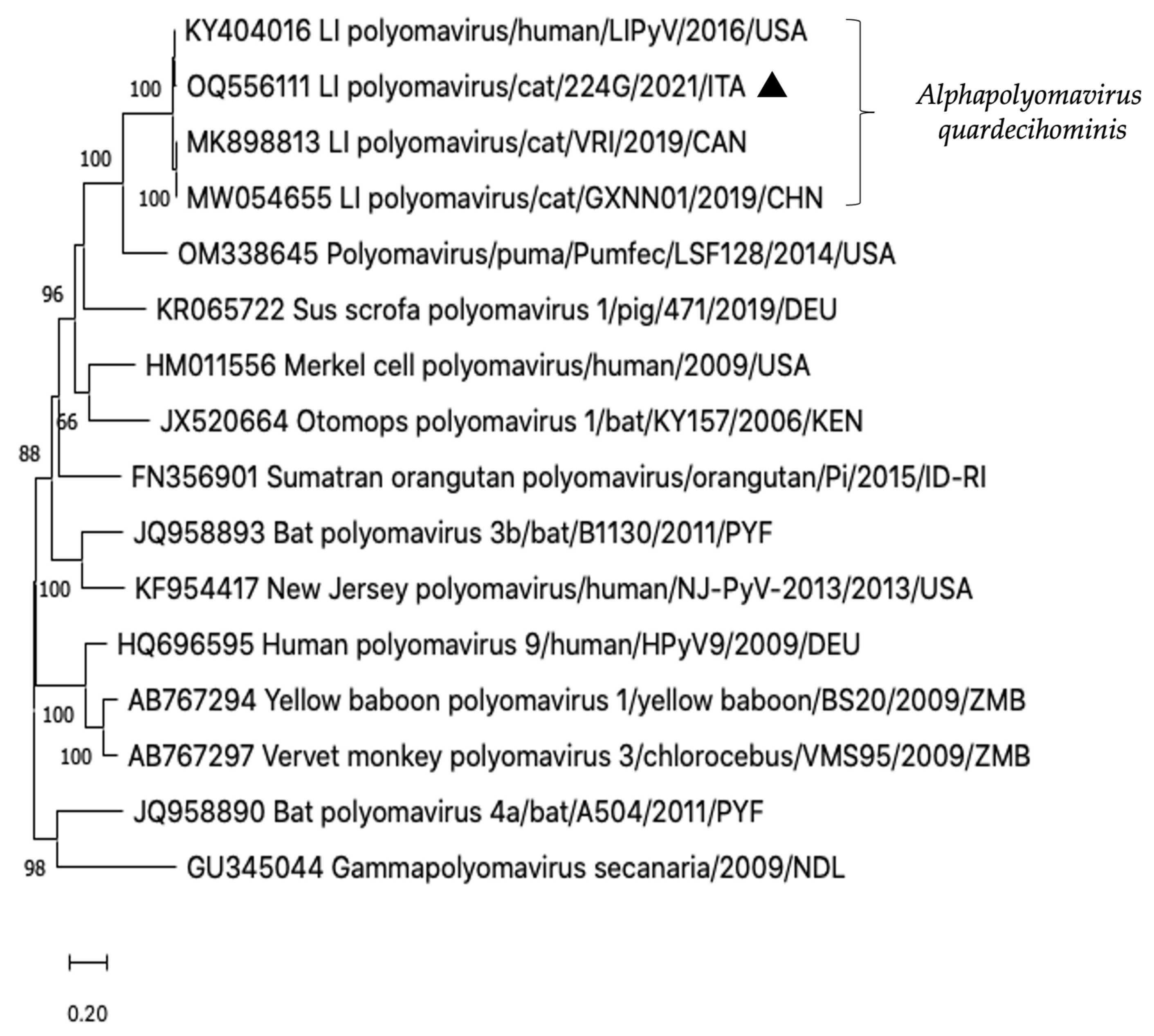
3.3. ONT Sequencing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pesavento, P.A.; Murphy, B.G. Common and Emerging Infectious Diseases in the Animal Shelter. Vet. Pathol. 2014, 51, 478–491. [Google Scholar] [CrossRef] [PubMed]
- Stuetzer, B.; Hartmann, K. Feline Parvovirus Infection and Associated Diseases. Vet. J. 2014, 201, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Barrs, V.R. Feline Panleukopenia: A Re-Emergent Disease. Vet. Clin. N. Am. Small Anim. Pract. 2019, 49, 651–670. [Google Scholar] [CrossRef]
- Lau, S.K.P.; Woo, P.C.Y.; Yeung, H.C.; Teng, J.L.L.; Wu, Y.; Bai, R.; Fan, R.Y.Y.; Chan, K.-H.; Yuen, K.-Y. Identification and Characterization of Bocaviruses in Cats and Dogs Reveals a Novel Feline Bocavirus and a Novel Genetic Group of Canine Bocavirus. J. Gen. Virol. 2012, 93, 1573–1582. [Google Scholar] [CrossRef]
- Pinto, P.; Wang, Q.; Chen, N.; Dubovi, E.J.; Daniels, J.B.; Millward, L.M.; Buonavoglia, C.; Martella, V.; Saif, L.J. Discovery and Genomic Characterization of Noroviruses from a Gastroenteritis Outbreak in Domestic Cats in the US. PLoS ONE 2012, 7, e32739. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.P.; Woo, P.C.Y.; Yip, C.C.Y.; Choi, G.K.Y.; Wu, Y.; Bai, R.; Fan, R.Y.Y.; Lai, K.K.Y.; Chan, K.-H.; Yuen, K.-Y. Identification of a Novel Feline Picornavirus from the Domestic Cat. J. Virol. 2012, 86, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.F.F.; Mesquita, J.R.; Nascimento, M.S.J.; Kondov, N.O.; Wong, W.; Reuter, G.; Knowles, N.J.; Vega, E.; Esona, M.D.; Deng, X.; et al. Feline Fecal Virome Reveals Novel and Prevalent Enteric Viruses. Vet. Microbiol. 2014, 171, 102–111. [Google Scholar] [CrossRef]
- Li, Y.; Gordon, E.; Idle, A.; Altan, E.; Seguin, M.A.; Estrada, M.; Deng, X.; Delwart, E. Virome of a Feline Outbreak of Diarrhea and Vomiting Includes Bocaviruses and a Novel Chapparvovirus. Viruses 2020, 12, 506. [Google Scholar] [CrossRef]
- Fahsbender, E.; Altan, E.; Estrada, M.; Seguin, M.A.; Young, P.; Leutenegger, C.M.; Delwart, E. Lyon-IARC Polyomavirus DNA in Feces of Diarrheic Cats. Microbiol. Resour. Announc. 2019, 8, e00550-19. [Google Scholar] [CrossRef]
- Harbour, D.A.; Ashley, C.R.; Williams, P.D.; Gruffydd-Jones, T.J. Natural and Experimental Astrovirus Infection of Cats. Vet. Rec. 1987, 120, 555–557. [Google Scholar] [CrossRef]
- Takano, T.; Kusuhara, H.; Kuroishi, A.; Takashina, M.; Doki, T.; Nishinaka, T.; Hohdatsu, T. Molecular Characterization and Pathogenicity of a Genogroup GVI Feline Norovirus. Vet. Microbiol. 2015, 178, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Hiramatsu, K.; Matsuyama, M.; Mutoh, K.; Matsumoto, Y.; Fukushima, T.; Doki, T.; Kusuhara, H.; Hohdatsu, T. Viral Shedding and Clinical Status of Feline-Norovirus-Infected Cats after Reinfection with the Same Strain. Arch. Virol. 2018, 163, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Castro, T.X.; Cubel Garcia, R.d.C.N.; Fumian, T.M.; Costa, E.M.; Mello, R.; White, P.A.; Leite, J.P.G. Detection and Molecular Characterization of Caliciviruses (Vesivirus and Norovirus) in an Outbreak of Acute Diarrhea in Kittens from Brazil. Vet. J. 2015, 206, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, B.; Di Profio, F.; Melegari, I.; Marsilio, F.; Martella, V. Detection of Feline Kobuviruses in Diarrhoeic Cats, Italy. Vet. Microbiol. 2015, 176, 186–189. [Google Scholar] [CrossRef]
- Di Martino, B.; Di Profio, F.; Melegari, I.; Sarchese, V.; Cafiero, M.A.; Robetto, S.; Aste, G.; Lanave, G.; Marsilio, F.; Martella, V. A Novel Feline Norovirus in Diarrheic Cats. Infect. Genet. Evol. 2016, 38, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, B.; Lanave, G.; Di Profio, F.; Melegari, I.; Marsilio, F.; Camero, M.; Catella, C.; Capozza, P.; Bányai, K.; Barrs, V.R.; et al. Identification of Feline Calicivirus in Cats with Enteritis. Transbound. Emerg. Dis. 2020, 67, 2579–2588. [Google Scholar] [CrossRef]
- Piewbang, C.; Kasantikul, T.; Pringproa, K.; Techangamsuwan, S. Feline Bocavirus-1 Associated with Outbreaks of Hemorrhagic Enteritis in Household Cats: Potential First Evidence of a Pathological Role, Viral Tropism and Natural Genetic Recombination. Sci. Rep. 2019, 9, 16367. [Google Scholar] [CrossRef]
- Di Profio, F.; Sarchese, V.; Palombieri, A.; Fruci, P.; Massirio, I.; Martella, V.; Fulvio, M.; Di Martino, B. Feline Chaphamaparvovirus in Cats with Enteritis and Upper Respiratory Tract Disease. Transbound. Emerg. Dis. 2022, 69, 660–668. [Google Scholar] [CrossRef]
- Van Brussel, K.; Wang, X.; Shi, M.; Carrai, M.; Feng, S.; Li, J.; Holmes, E.C.; Beatty, J.A.; Barrs, V.R. The Enteric Virome of Cats with Feline Panleukopenia Differs in Abundance and Diversity from Healthy Cats. Transbound. Emerg. Dis. 2022, 69, e2952–e2966. [Google Scholar] [CrossRef] [PubMed]
- Buonavoglia, C.; Martella, V.; Pratelli, A.; Tempesta, M.; Cavalli, A.; Buonavoglia, D.; Bozzo, G.; Elia, G.; Decaro, N.; Carmichael, L. Evidence for Evolution of Canine Parvovirus Type 2 in Italy. J. Gen. Virol. 2001, 82, 3021–3025. [Google Scholar] [CrossRef]
- Martella, V.; Lanave, G.; Mihalov-Kovács, E.; Marton, S.; Varga-Kugler, R.; Kaszab, E.; Di Martino, B.; Camero, M.; Decaro, N.; Buonavoglia, C.; et al. Novel Parvovirus Related to Primate Bufaviruses in Dogs. Emerg. Infect. Dis. 2018, 24, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Gheit, T.; Dutta, S.; Oliver, J.; Robitaille, A.; Hampras, S.; Combes, J.-D.; McKay-Chopin, S.; Le Calvez-Kelm, F.; Fenske, N.; Cherpelis, B.; et al. Isolation and Characterization of a Novel Putative Human Polyomavirus. Virology 2017, 506, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.W.; Leung, C.Y.H.; Gilbert, M.; Joyner, P.H.; Ng, E.M.; Tse, T.M.; Guan, Y.; Peiris, J.S.M.; Poon, L.L.M. Avian Coronavirus in Wild Aquatic Birds. J. Virol. 2011, 85, 12815–12820. [Google Scholar] [CrossRef] [PubMed]
- Gunn-Moore, D.A.; Gruffydd-Jones, T.J.; Harbour, D.A. Detection of Feline Coronaviruses by Culture and Reverse Transcriptase-Polymerase Chain Reaction of Blood Samples from Healthy Cats and Cats with Clinical Feline Infectious Peritonitis. Vet. Microbiol. 1998, 62, 193–205. [Google Scholar] [CrossRef]
- Marsilio, F.; Di Martino, B.; Decaro, N.; Buonavoglia, C. A Novel Nested PCR for the Diagnosis of Calicivirus Infections in the Cat. Vet. Microbiol. 2005, 105, 1–7. [Google Scholar] [CrossRef]
- Vennema, H.; de Bruin, E.; Koopmans, M. Rational Optimization of Generic Primers Used for Norwalk-like Virus Detection by Reverse Transcriptase Polymerase Chain Reaction. J. Clin. Virol. 2002, 25, 233–235. [Google Scholar] [CrossRef]
- Chu, D.K.W.; Poon, L.L.M.; Guan, Y.; Peiris, J.S.M. Novel Astroviruses in Insectivorous Bats. J. Virol. 2008, 82, 9107–9114. [Google Scholar] [CrossRef]
- Chung, J.-Y.; Kim, S.-H.; Kim, Y.-H.; Lee, M.-H.; Lee, K.-K.; Oem, J.-K. Detection and Genetic Characterization of Feline Kobuviruses. Virus Genes 2013, 47, 559–562. [Google Scholar] [CrossRef]
- Palombieri, A.; Fruci, P.; Sarchese, V.; Robetto, S.; Orusa, R.; Arbuatti, A.; Martella, V.; Di Martino, B.; Di Profio, F. Detection and Characterization of a Novel Picornavirus in European Badger (Meles Meles). Vet. Sci. 2022, 9, 645. [Google Scholar] [CrossRef]
- Allander, T.; Tammi, M.T.; Eriksson, M.; Bjerkner, A.; Tiveljung-Lindell, A.; Andersson, B. Cloning of a Human Parvovirus by Molecular Screening of Respiratory Tract Samples. Proc. Natl. Acad. Sci. USA 2005, 102, 12891–12896. [Google Scholar] [CrossRef]
- Djikeng, A.; Halpin, R.; Kuzmickas, R.; Depasse, J.; Feldblyum, J.; Sengamalay, N.; Afonso, C.; Zhang, X.; Anderson, N.G.; Ghedin, E.; et al. Viral Genome Sequencing by Random Priming Methods. BMC Genom. 2008, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Vilsker, M.; Moosa, Y.; Nooij, S.; Fonseca, V.; Ghysens, Y.; Dumon, K.; Pauwels, R.; Alcantara, L.C.; Vanden Eynden, E.; Vandamme, A.-M.; et al. Genome Detective: An Automated System for Virus Identification from High-Throughput Sequencing Data. Bioinformatics 2019, 35, 871–873. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Campolo, M.; Lorusso, E.; Cavicchio, P.; Camero, M.; Bellacicco, A.L.; Decaro, N.; Elia, G.; Greco, G.; Corrente, M.; et al. Norovirus in Captive Lion Cub (Panthera Leo). Emerg. Infect. Dis. 2007, 13, 1071–1073. [Google Scholar] [CrossRef]
- Li, Y.; Huang, H.; Lan, T.; Wang, W.; Zhang, J.; Zheng, M.; Cao, L.; Sun, W.; Lu, H. First Detection and Complete Genome Analysis of the Lyon IARC Polyomavirus in China from Samples of Diarrheic Cats. Virus Genes 2021, 57, 284–288. [Google Scholar] [CrossRef]
- Mira, F.; Canuti, M.; Purpari, G.; Cannella, V.; Di Bella, S.; Occhiogrosso, L.; Schirò, G.; Chiaramonte, G.; Barreca, S.; Pisano, P.; et al. Molecular Characterization and Evolutionary Analyses of Carnivore Protoparvovirus 1 NS1 Gene. Viruses 2019, 11, 308. [Google Scholar] [CrossRef]
- Lu, G.; Huang, M.; Chen, X.; Sun, Y.; Huang, J.; Hu, R.; Li, S. Identification and Genome Characterization of a Novel Feline Picornavirus Proposed in the Hunnivirus Genus. Infect. Genet. Evol. 2019, 71, 47–50. [Google Scholar] [CrossRef]
- Okamoto, H.; Takahashi, M.; Nishizawa, T.; Tawara, A.; Fukai, K.; Muramatsu, U.; Naito, Y.; Yoshikawa, A. Genomic characterization of TT viruses (TTVs) in pigs, cats and dogs and their relatedness with species-specific TTVs in primates and tupaias. J. Gen. Virol. 2002, 83, 1291–1297. [Google Scholar] [CrossRef]
- Biagini, P.; Uch, R.; Belhouchet, M.; Attoui, H.; Cantaloube, J.F.; Brisbarre, N.; de Micco, P. Circular genomes related to anelloviruses identified in human and animal samples by using a combined rolling-circle amplification/sequence-independent single primer amplification approach. J. Gen. Virol. 2007, 88, 2696–2701. [Google Scholar] [CrossRef]
- Zhu, C.X.; Shan, T.L.; Cui, L.; Luo, X.N.; Liu, Z.J.; Tang, S.D.; Liu, Z.W.; Yuan, C.L.; Lan, D.L.; Zhao, W.; et al. Molecular detection and sequence analysis of feline Torque teno virus (TTV) in China. Virus Res. 2011, 156, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, H.; Wang, Y.; Liu, Z.; Li, J.; Guo, L.; Yang, S.; Shen, Q.; Zhao, X.; Cui, L.; et al. Identification and genomic characterization of a novel species of feline anellovirus. Virol. J. 2016, 13, 146. [Google Scholar] [CrossRef] [PubMed]
- Jarošová, V.; Hrazdilová, K.; Filipejová, Z.; Schánilec, P.; Celer, V. Whole genome sequencing and phylogenetic analysis of feline anelloviruses. Infect. Genet. Evol. 2015, 32, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Kraberger, S.; Serieys, L.E.; Richet, C.; Fountain-Jones, N.M.; Baele, G.; Bishop, J.M.; Nehring, M.; Ivan, J.S.; Newkirk, E.S.; Squires, J.R.; et al. Complex Evolutionary History of Felid Anelloviruses. Virology 2021, 562, 176–189. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, L.; Tu, Z.; Sun, S.; Sun, Y.; Yi, L.; Tu, C.; He, B. Virome Profiling of an Amur leopard cat Reveals Multiple Anelloviruses and a Bocaparvovirus. Vet. Sci. 2022, 9, 640. [Google Scholar] [CrossRef]
- Munday, J.S.; Dunowska, M.; Hills, S.F.; Laurie, R.E. Genomic Characterization of Felis Catus Papillomavirus-3: A Novel Papillomavirus Detected in a Feline Bowenoid in Situ Carcinoma. Vet. Microbiol. 2013, 165, 319–325. [Google Scholar] [CrossRef]
- Chu, S.; Wylie, T.N.; Wylie, K.M.; Johnson, G.C.; Skidmore, Z.L.; Fleer, M.; Griffith, O.L.; Bryan, J.N. A Virome Sequencing Approach to Feline Oral Squamous Cell Carcinoma to Evaluate Viral Causative Factors. Vet. Microbiol. 2020, 240, 108491. [Google Scholar] [CrossRef]
- Donato, G.; Masucci, M.; De Luca, E.; Alibrandi, A.; De Majo, M.; Berjaoui, S.; Martino, C.; Mangano, C.; Lorusso, A.; Pennisi, M.G. Feline Morbillivirus in Southern Italy: Epidemiology, Clinico-Pathological Features and Phylogenetic Analysis in Cats. Viruses 2021, 13, 1449. [Google Scholar] [CrossRef]
- Park, E.-S.; Suzuki, M.; Kimura, M.; Mizutani, H.; Saito, R.; Kubota, N.; Hasuike, Y.; Okajima, J.; Kasai, H.; Sato, Y.; et al. Epidemiological and Pathological Study of Feline Morbillivirus Infection in Domestic Cats in Japan. BMC Vet. Res. 2016, 12, 228. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Nakagawa, S.; Yoshikawa, R.; Kuwahara, C.; Hagiwara, H.; Asai, K.-I.; Kawakami, K.; Yamamoto, Y.; Ogawa, M.; Miyazawa, T. Genetic Diversity of Feline Morbilliviruses Isolated in Japan. J. Gen. Virol. 2014, 95, 1464–1468. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Boyle, J.F.; Floyd, K.; Fudge, A.; Barker, J. An enteric coronavirus infection of cats and its relationship to feline infectious peritonitis. Am. J. Vet. Res. 1981, 42, 368–377. [Google Scholar] [PubMed]
- Addie, D.D.; Jarrett, O. A study of naturally occurring feline coronavirus infections in kittens. Vet. Rec. 1992, 130, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Addie, D.D.; Toth, S.; Murray, G.D.; Jarrett, O. Risk of feline infectious peritonitis in cats naturally infected with feline coronavirus. Am. J. Vet. Res. 1995, 56, 429–434. [Google Scholar]
- Pedersen, N.C. An overview of feline enteric coronavirus and infectious peritonitis virus infections. Feline Pract. 1995, 23, 7–20. [Google Scholar]
- Pedersen, N.C.; Allen, C.E.; Lyons, L.A. Pathogenesis of feline enteric coronavirus infection. J. Feline Med. Surg. 2008, 10, 529–541. [Google Scholar] [CrossRef]
- Hayashi, T.; Watabe, Y.; Nakayama, H.; Fujiwara, K. Enteritis due to feline infectious peritonitis virus. Nihon Juigaku Zasshi 1982, 44, 97–106. [Google Scholar] [CrossRef]
- Dea, S.; Roy, R.S.; Elazhary, M.A. Coronavirus-like Particles in the Feces of a Cat with Diarrhea. Can. Vet. J. 1982, 23, 153–155. [Google Scholar]
- Felten, S.; Klein-Richers, U.; Unterer, S.; Bergmann, M.; Leutenegger, C.M.; Pantchev, N.; Balzer, J.; Zablotski, Y.; Hofmann-Lehmann, R.; Hartmann, K. Role of Feline Coronavirus as Contributor to Diarrhea in Cats from Breeding Catteries. Viruses 2022, 14, 858. [Google Scholar] [CrossRef]
- Klein-Richers, U.; Hartmann, K.; Hofmann-Lehmann, R.; Unterer, S.; Bergmann, M.; Rieger, A.; Leutenegger, C.; Pantchev, N.; Balzer, J.; Felten, S. Prevalence of Feline Coronavirus Shedding in German Catteries and Associated Risk Factors. Viruses 2020, 12, 1000. [Google Scholar] [CrossRef]
- Povey, C.; Ingersoll, J. Cross-protection among feline caliciviruses. Infect. Immun. 1975, 11, 877–885. [Google Scholar] [CrossRef]
- Fastier, L.B. A new feline virus isolated in tissue culture. Am. J. Vet. Res. 1957, 18, 382–389. [Google Scholar] [PubMed]
- Marshall, J.A.; Kennett, M.L.; Rodger, S.M.; Studdert, M.J.; Thompson, W.L.; Gust, I.D. Virus and virus-like particles in the faeces of cats with and without diarrhoea. Aust. Vet. J. 1987, 64, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, M. Different stabilities to bile among feline calicivirus strains of respiratory and enteric origin. Vet. Microbiol. 1992, 31, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Soma, T.; Nakagomi, O.; Nakagomi, T.; Mochizuki, M. Detection of Norovirus and Sapovirus from diarrheic dogs and cats in Japan. Microbiol. Immunol. 2015, 59, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, L.; Deng, X.; Kapusinszky, B.; Pesavento, P.A.; Delwart, E. Faecal virome of cats in an animal shelter. J. Gen. Virol. 2014, 95, 2553–2564. [Google Scholar] [CrossRef]
- Lu, G.; Zhang, X.; Luo, J.; Sun, Y.; Xu, H.; Huang, J.; Ou, J.; Li, S. First report and genetic characterization of feline kobuvirus in diarrhoeic cats in China. Transbound. Emerg. Dis. 2018, 65, 1357–1363. [Google Scholar] [CrossRef]
- Niu, T.J.; Yi, S.S.; Wang, X.; Wang, L.H.; Guo, B.Y.; Zhao, L.Y.; Zhang, S.; Dong, H.; Wang, K.; Hu, X.G. Detection and genetic characterization of kobuvirus in cats: The first molecular evidence from Northeast China. Infect. Genet. Evol. 2019, 68, 58–67. [Google Scholar] [CrossRef]
- Reuter, G.; Pankovics, P.; Knowles, N.J.; Boros, Á. Two closely related novel picornaviruses in cattle and sheep in Hungary from 2008 to 2009, proposed as members of a new genus in the family Picornaviridae. J. Virol. 2012, 86, 13295–132302. [Google Scholar] [CrossRef]
- Du, J.; Lu, L.; Liu, F.; Su, H.; Dong, J.; Sun, L.; Zhu, Y.; Ren, X.; Yang, F.; Guo, F.; et al. Distribution and characteristics of rodent picornaviruses in China. Sci. Rep. 2016, 6, 34381. [Google Scholar] [CrossRef]
- Firth, C.; Bhat, M.; Firth, M.A.; Williams, S.H.; Frye, M.J.; Simmonds, P.; Conte, J.M.; Ng, J.; Garcia, J.; Bhuva, N.P.; et al. Detection of zoonotic pathogens and characterization of novel viruses carried by commensal Rattus norvegicus in New York City. mBio 2014, 5, e01933-14. [Google Scholar] [CrossRef]
- Zhang, M.; Li, Q.; Wu, F.; Ou, Z.; Li, Y.; You, F.; Chen, Q. Epidemiology, Genetic Characterization, and Evolution of Hunnivirus Carried by Rattus norvegicus and Rattus tanezumi: The First Epidemiological Evidence from Southern China. Pathogens 2021, 10, 661. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Yu, J.; Song, X.; Zhang, J.; Zhang, B. The discovery of novel papillomaviruses in cats in Southwest China. Virus Genes 2023, 28, 1–5. [Google Scholar] [CrossRef]
- Yamashita-Kawanishi, N.; Sawanobori, R.; Matsumiya, K.; Uema, A.; Chambers, J.K.; Uchida, K.; Shimakura, H.; Tsuzuki, M.; Chang, C.Y.; Chang, H.W.; et al. Detection of felis catus papillomavirus type 3 and 4 DNA from squamous cell carcinoma cases of cats in Japan. J. Vet. Med. Sci. 2018, 80, 1236–1240. [Google Scholar] [CrossRef]
- Munday, J.S.; Thomson, N.A.; Henderson, G.; Fairley, R.; Orbell, G.M. Identification of Felis catus papillomavirus 3 in skin neoplasms from four cats. J. Vet. Diagn. Investig. 2018, 30, 324–328. [Google Scholar] [CrossRef] [PubMed]
- De Luca, E.; Sautto, G.A.; Crisi, P.E.; Lorusso, A. Feline Morbillivirus Infection in Domestic Cats: What Have We Learned So Far? Viruses 2021, 13, 683. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.; Lau, S.K.; Wong, B.H.; Fan, R.Y.; Wong, A.Y.; Zhang, A.J.; Wu, Y.; Choi, G.K.; Li, K.S.; Hui, J.; et al. Feline morbillivirus, a previously undescribed paramyxovirus associated with tubulointerstitial nephritis in domestic cats. Proc. Natl. Acad. Sci. USA 2012, 109, 5435–5440. [Google Scholar] [CrossRef]
- Sato, H.; Yoneda, M.; Honda, T.; Kai, C. Morbillivirus receptors and tropism: Multiple pathways for infection. Front. Microbiol. 2012, 3, 75. [Google Scholar] [CrossRef]
- Karlsson, O.E.; Belák, S.; Granberg, F. The effect of preprocessing by sequence-independent, single-primer amplification (SISPA) on metagenomic detection of viruses. Biosecur. Bioterror. 2013, 11, S227–S234. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Virus | Primer | Sequence (5′ to 3′) | Sense | Length (bp) | Tm °C | Target Gene | References |
|---|---|---|---|---|---|---|---|
| FPV | 555Fw | CAGGAAGATATCCAGAAGGA | + | 590 | 50 | VP | [20] |
| 555ReV | GGTGCTAGTTGATATGTAATAAACA | − | |||||
| BoVs | BoVF | GCCAGCACNGGNAARACMAA | + | 141 | 50 | NS1 | [4] |
| BoVR | CATNAGNCAYTCYTCCCACCA | − | |||||
| FBuV | 165F | CTGGTTTAATCCAGCAGACT | + | 207 | 53 | VP2 | [21] |
| 371R | TGAAGACCA AGGTAGTAG | − | |||||
| FeChPV | FechaF1 | GGTGCGACGACGGAAGATAT | + | 332 | 58 | NS1 | [8] |
| FechaR1 | CAACACCACCATCTCCTGCT | − | |||||
| FechaF2 | GCTGCAGTTCAGGTAGCTCA | + | 310 | ||||
| PyVs | PyVF | CAWGCTGTRTITAGTAATA | + | 240 | 48 | LT-Ag | [22] |
| PyVR | RWTTATTMACHCCITTAC | - | |||||
| FCoV | FCoVF | CTGCATGTCAAACTATTG | + | 800 | 50 | S | [24] |
| FCoVR1 | CTTGTGCATCAGCACTC | − | |||||
| FCoVR2 | TGCTATTATGGGACGG | − | 217 | ||||
| FCV | Cali1 | AACCTGCGCTAACGTGCTTA | + | 926 | 57 | RdRp | [25] |
| CalcR | CAGTGACAATACACCCAGAAG | − | |||||
| CaliINT1 | TGGTGATGATGAATGGGCATC | + | 324 | 58 | |||
| CaliINT2 | ACACCAGAGCCAGAGATAGA | − | |||||
| NoVs | JV12Y | ATACCACCTATGATGCAGAYTA | + | 273 | 37 | RdRp | [26] |
| JV13I | TCATCATCACCATAGAAGAG | − | |||||
| AstVs | panAstV-F1 | GARTTYGATTGGRCKCGKTAYGA | + | 50 | RdRp | [27] | |
| panAstV-F2 | GARTTYGATTGGRCKAGGTAYGA | + | |||||
| panAstV-R1 | GGYTTKACCCACATNCCRAA | − | |||||
| panAstVn-F1 | CGKTAYGATGGKACKATHCC | + | 420 | 50 | |||
| panAstVn-F2 | AGGTAYGATGGKACKATHCC | + | |||||
| FeKoV | FeKoVF | CATGCTCCTCGGTGGTCTCA | + | 631 | 55 | 3DRdRp | [28] |
| FeKoVR | GTCCGGGTCCATCACAGGGT | − | |||||
| SaKoVs | SaKoV-for | GGTAGCGCGGTCGGTTGCGACCC | + | 688 | 55 | 3DRdRp | [29] |
| SaKoV-rev | CCCAGGACTGGTAGTTGTTAG | − |
| Virus | Positive Samples % (Positive/Total) | Diarrhoeic % (Positive/Total) | Non-Diarrhoeic % (Positive/Total) |
|---|---|---|---|
| FPV | 35.5 (22/62) | 72.4 (21/29) | 3.0 (1/33) |
| FcoV | 32.2 (20/62) | 24.1 (7/29) | 39.4 (13/33) |
| FeChPV | 12.9 (8/62) | 13.8 (4/29) | 12.1 (4/33) |
| FCV | 6.4 (4/62) | 13.8 (4/29) | 0.0 (0/33) |
| FeKoV | 1.6 (1/62) | 3.4 (1/29) | 0.0 (0/33) |
| NoV | 1.6 (1/62) | 3.4 (1/29) | 0.0 (0/33) |
| PyV | 1.6 (1/62) | 3.4 (1/29) | 0.0 (0/33) |
| SaKoV | 1.6 (1/62) | 3.4 (1/29) | 0.0 (0/33) |
| BoV | 0.0 (0/62) | 0.0 (0/29) | 0.0 (0/33) |
| FbuV | 0.0 (0/62) | 0.0 (0/29) | 0.0 (0/33) |
| AstV | 0.0 (0/62) | 0.0 (0/29) | 0.0 (0/33) |
| Total positive cats | 66.1 (41/62) | 86.2 (25/29) | 48.4 (16/33) |
| Viruses | Diarrhoeic % (Positive/Total) | Non-Diarrhoeic % (Positive/Total) | Total % (Positive/Total) |
|---|---|---|---|
| FPV + FCoV | 24.1 (7/29) | 0.0 (0/33) | 11.3 (7/62) |
| FPV + FCV | 6.9 (2/29) | 0.0 (0/33) | 3.2 (2/62) |
| FPV + FeChPV | 6.9 (2/29) | 0.0 (0/33) | 3.2 (2/62) |
| FCoV + FeChPV | 0.0 (0/29) | 6.1 (2/33) | 3.2 (2/62) |
| FPV + NoV +FCoV | 3.4 (1/29) | 0.0 (0/33) | 1.6 (1/62) |
| FeChPV+ PyV + SaKoV | 3.4 (1/29) | 0.0 (0/33) | 1.6 (1/62) |
| Total co-infected cats | 44.8 (13/29) | 6.1 (2/33) | 24.2 (15/62) |
| Sample ID | #168G/2020 | #185G/2020 | #188G/2020 | #207G/2020 | #212G/2020 | #221G/2021 | #224G/2021 | #229G/2021 | Contigs | |
|---|---|---|---|---|---|---|---|---|---|---|
| Viral Family | Species | |||||||||
| Parvoviridae | Feline parvovirus | 1 | 2 | 2 | 1 | 1 | 7 | |||
| Feline chaphamaparvovirus | 1 | 2 | 2 | 5 | ||||||
| Polyomaviridae | Lyon IARC polyomaviruses | 1 | 1 | |||||||
| Anelloviridae | Torque teno felis virus | 1 | 2 | 1 | 1 | 1 | 6 | |||
| Papillomaviridae | Felis catus papillomavirus | 4 | 4 | 8 | ||||||
| Caliciviridae | Feline calicivirus | 1 | 1 | 2 | 1 | 5 | ||||
| Picornaviridae | Feline sakobuvirus | 1 | 1 | |||||||
| Feline hunnivirus | 1 | 1 | ||||||||
| Paramyxoviridae | Feline morbillivirus | 7 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Profio, F.; Sarchese, V.; Fruci, P.; Aste, G.; Martella, V.; Palombieri, A.; Di Martino, B. Exploring the Enteric Virome of Cats with Acute Gastroenteritis. Vet. Sci. 2023, 10, 362. https://doi.org/10.3390/vetsci10050362
Di Profio F, Sarchese V, Fruci P, Aste G, Martella V, Palombieri A, Di Martino B. Exploring the Enteric Virome of Cats with Acute Gastroenteritis. Veterinary Sciences. 2023; 10(5):362. https://doi.org/10.3390/vetsci10050362
Chicago/Turabian StyleDi Profio, Federica, Vittorio Sarchese, Paola Fruci, Giovanni Aste, Vito Martella, Andrea Palombieri, and Barbara Di Martino. 2023. "Exploring the Enteric Virome of Cats with Acute Gastroenteritis" Veterinary Sciences 10, no. 5: 362. https://doi.org/10.3390/vetsci10050362
APA StyleDi Profio, F., Sarchese, V., Fruci, P., Aste, G., Martella, V., Palombieri, A., & Di Martino, B. (2023). Exploring the Enteric Virome of Cats with Acute Gastroenteritis. Veterinary Sciences, 10(5), 362. https://doi.org/10.3390/vetsci10050362