


Attenuation of Hypothyroidism-Induced Cognitive Impairment by Modulating Serotonin Mediation

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Ethics Statement
2.2. Drug-Induced Hypothyroidism
2.3. Open Field Test (OFT)
2.4. T-Maze with Spontaneous Alternation
2.5. Novel Object Recognition Test (NOR)
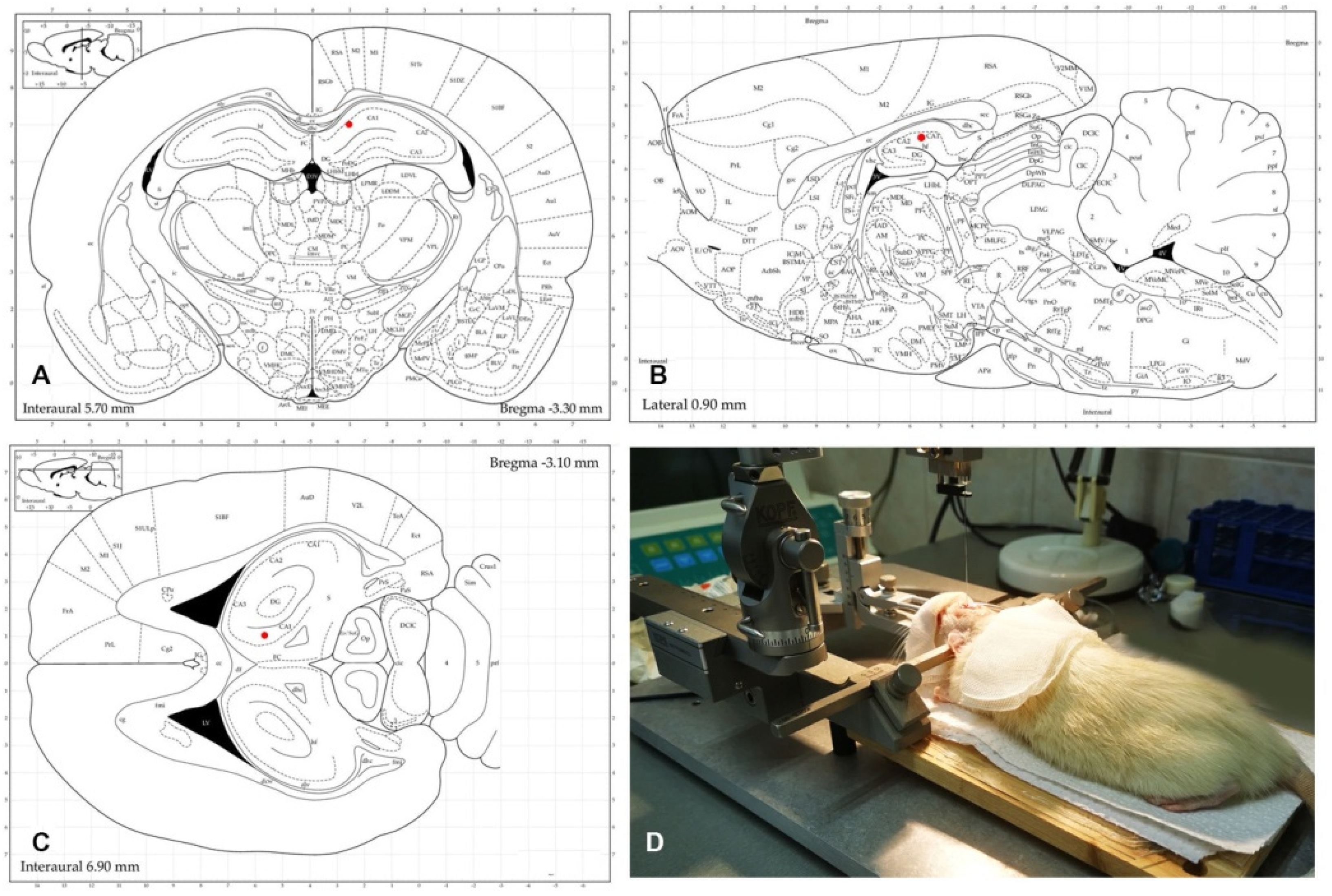
2.6. Stereotactic Injection
2.7. Statistics
3. Results
3.1. fT4
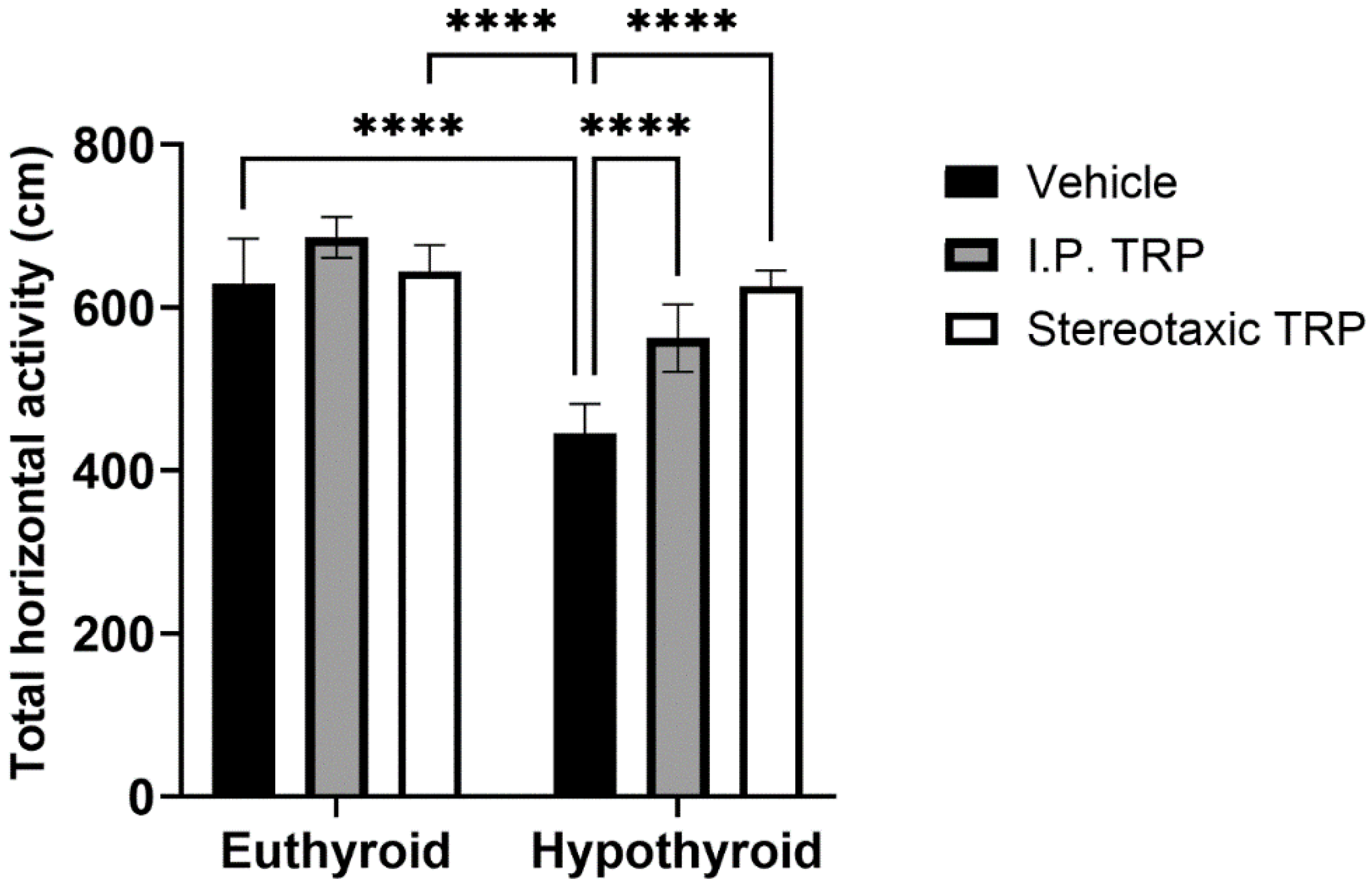
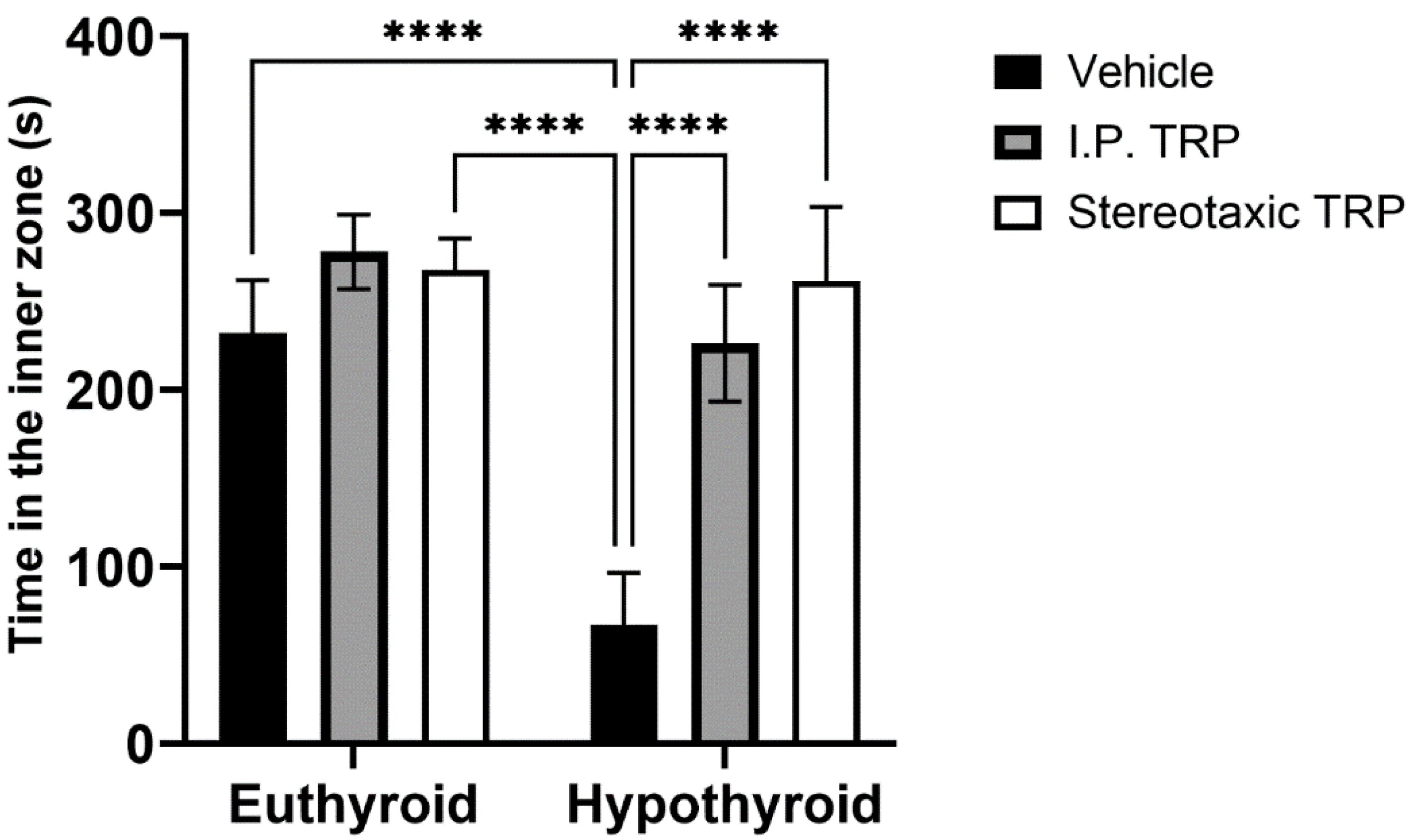
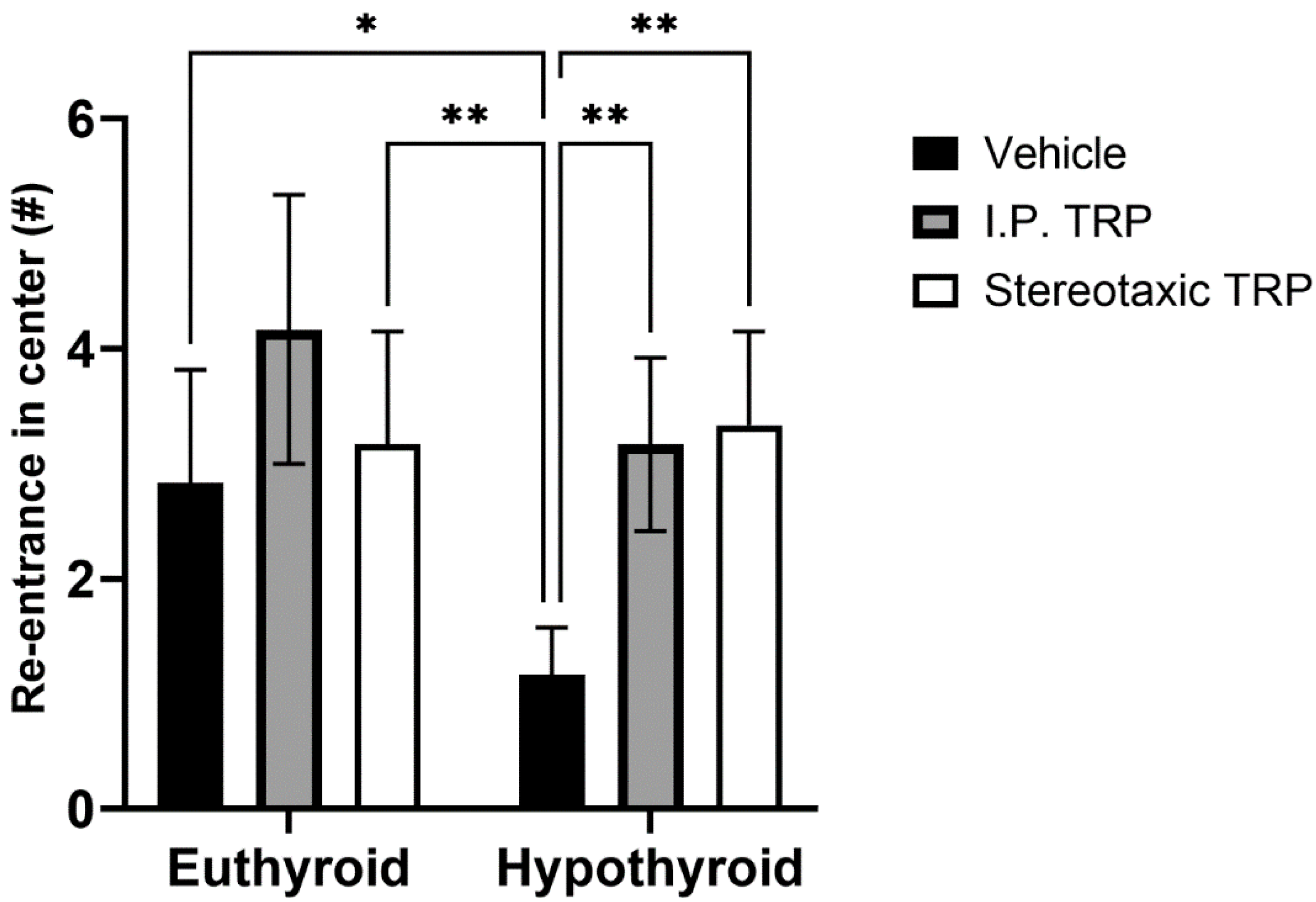
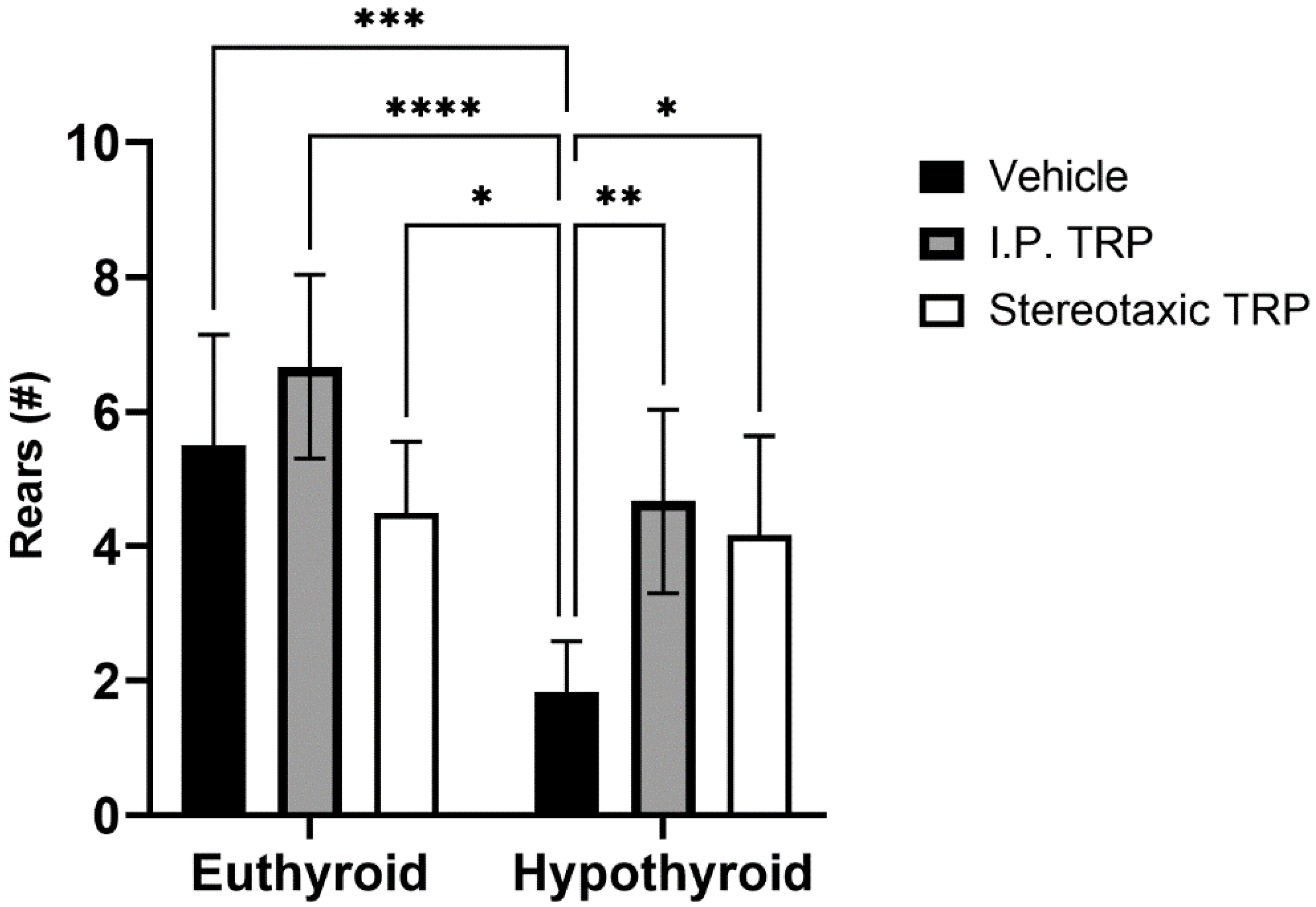
3.2. Open Field Test
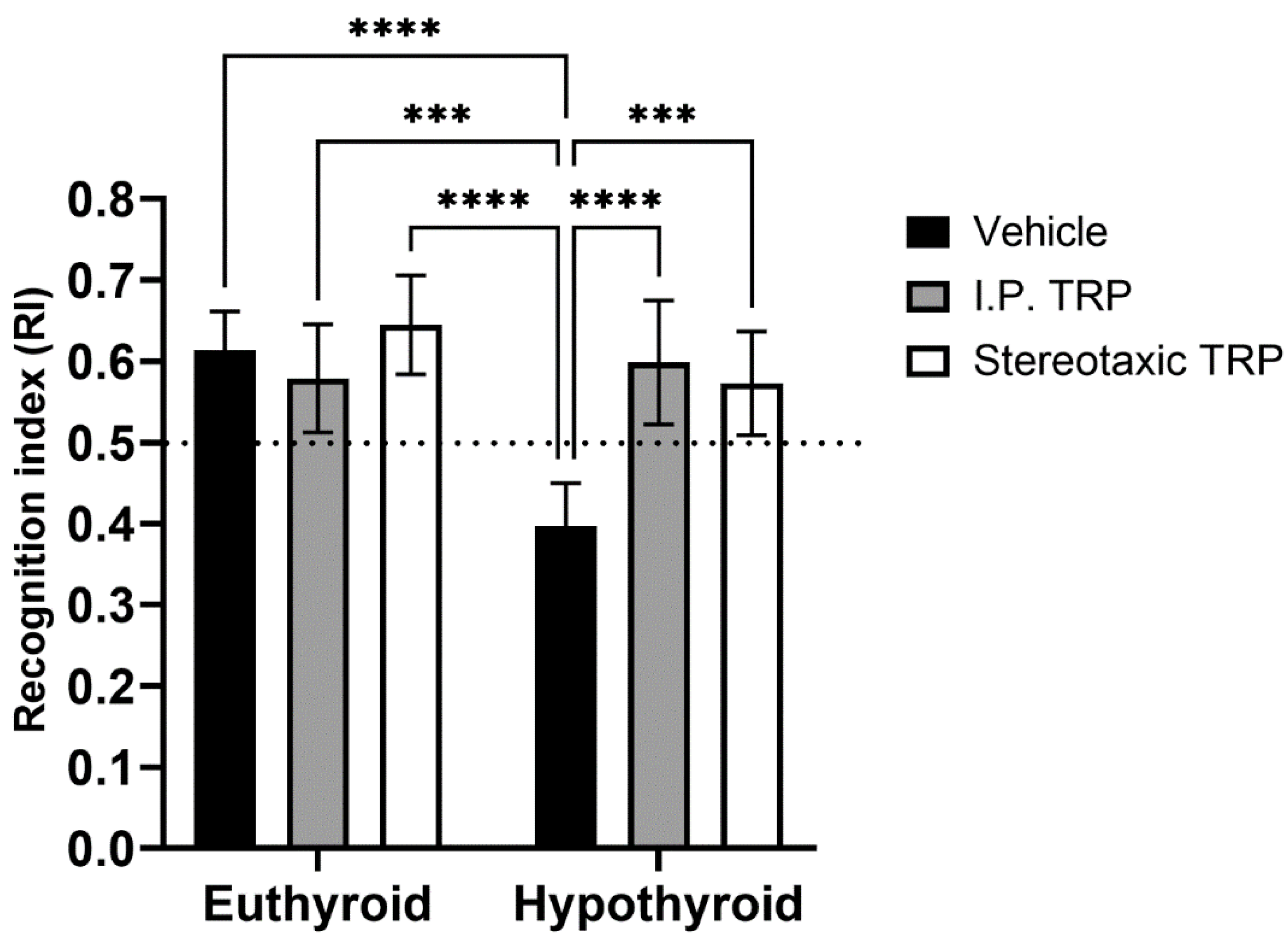
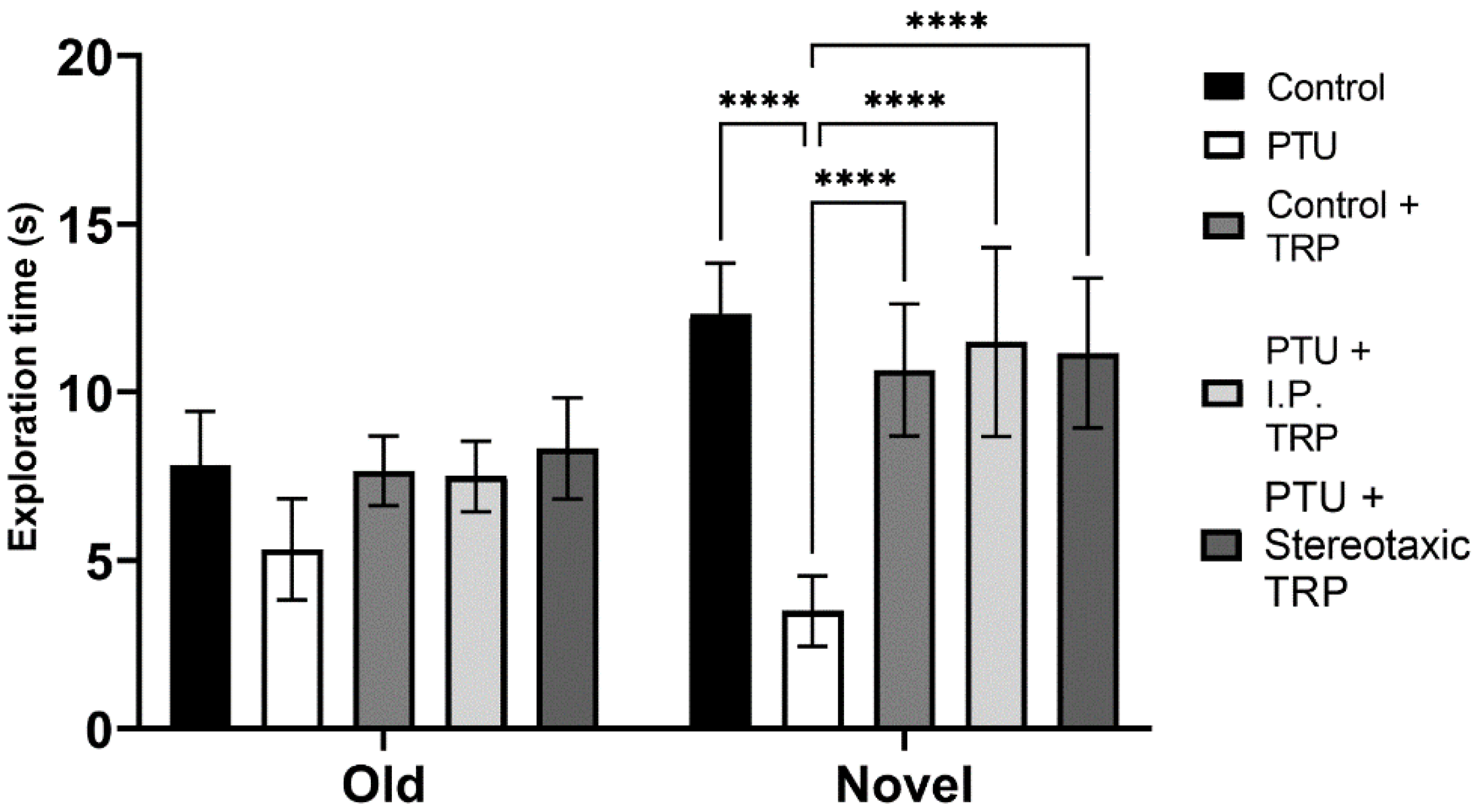
3.3. Novel Object Recognition Test
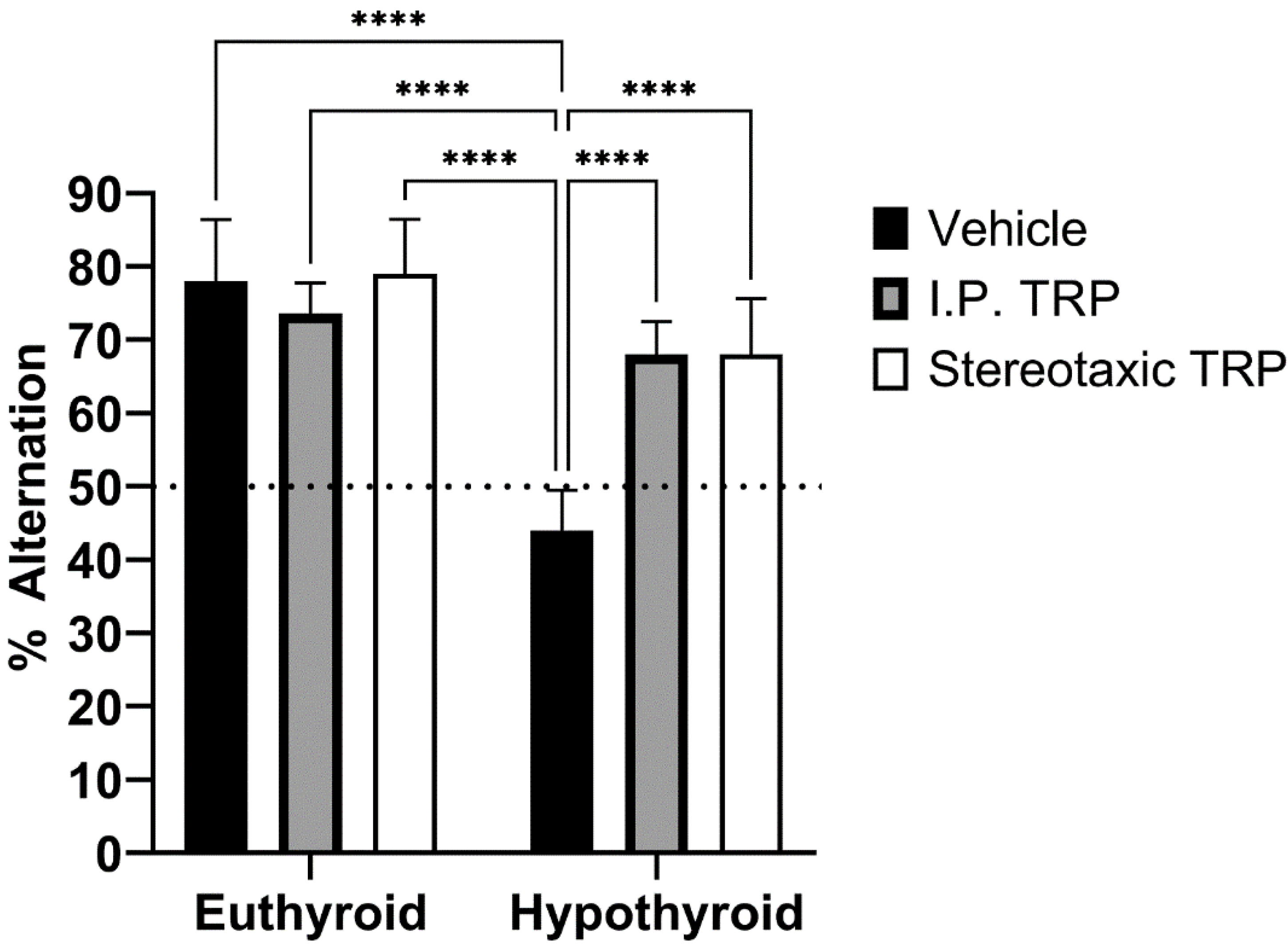
3.4. T-Maze Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bocheva, G.; Valcheva-Traykova, M.; Landzhov, B. Does hypothyroidism augment sun-induced skin damage? Redox Rep. 2018, 23, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Valcheva-Traykova, M.L.; Bocheva, G. Effect of ultraviolet radiation on the free radicals formation in hypothyroid rat’s liver. Bulg. Chem. Commun. 2016, 48, 384–388. [Google Scholar]
- Gottwald-Hostalek, U.; Schulte, B. Low awareness and under-diagnosis of hypothyroidism. Curr. Med. Res. Opin. 2022, 38, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Alzoubi, K.H.; Gerges, N.Z.; Aleisa, A.M.; Alkadhi, K.A. Levothyroxin restores hypothyroidism-induced impairment of hippocampus-dependent learning and memory: Behavioral, electrophysiological, and molecular studies. Hippocampus 2009, 19, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Rovet, J.F. The role of thyroid hormones for brain development and cognitive function. Endocr. Dev. 2014, 26, 26–43. [Google Scholar] [PubMed]
- Hage, M.P.; Azar, S.T. The Link between Thyroid Function and Depression. J. Thyroid Res. 2012, 2012, 590648. [Google Scholar] [CrossRef] [PubMed]
- Lieber, I.; Ott, M.; Öhlund, L.; Lundqvist, R.; Eliasson, M.; Sandlund, M.; Werneke, U. Patterns of Thyroid Hormone Prescription in Patients with Bipolar or Schizoaffective Disorder: Findings from the LiSIE Retrospective Cohort Study. J. Clin. Med. 2021, 10, 5062. [Google Scholar] [CrossRef]
- Stern, M.; Finch, A.; Haskard-Zolnierek, K.B.; Howard, K.; Deason, R.G. Cognitive decline in mid-life: Changes in memory and cognition related to hypothyroidism. J. Health Psychol. 2022, 13591053221107745. [Google Scholar] [CrossRef]
- Churilov, L.P.; Sobolevskaia, P.A.; Stroev, Y.I. Thyroid gland and brain: Enigma of Hashimoto’s encephalopathy. Best Pract. Res. Clin. Endocrinol. Metab. 2019, 33, 101364. [Google Scholar] [CrossRef]
- Waliszewska-Prosół, M.; Ejma, M. Hashimoto Encephalopathy-Still More Questions than Answers. Cells 2022, 11, 2873. [Google Scholar] [CrossRef]
- Hughes, J.H.; Gallagher, P.; Young, A.H. Effects of acute tryptophan depletion on cognitive function in euthymic bipolar patients. Eur. Neuropsychopharmacol. 2002, 12, 123–128. [Google Scholar] [CrossRef]
- Švob Štrac, D.; Pivac, N.; Mück-Šeler, D. The serotonergic system and cognitive function. Transl. Neurosci. 2016, 7, 35–49. [Google Scholar] [CrossRef]
- Głombik, K.; Detka, J.; Kurek, A.; Budziszewska, B. Impaired Brain Energy Metabolism: Involvement in Depression and Hypothyroidism. Front. Neurosci. 2020, 14, 586939. [Google Scholar] [CrossRef]
- Haider, S.; Khaliq, S.; Ahmed, S.P.; Haleem, D.J. Long-term tryptophan administration enhances cognitive performance and increases 5HT metabolism in the hippocampus of female rats. Amino Acids 2006, 31, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.; Saleem, S.; Shameem, S.; Ahmed, S.P.; Parveen, T.; Haleem, D.J. Is anorexia in thioacetamide-induced cirrhosis related to an altered brain serotonin concentration? Pol. J. Pharmacol. 2004, 56, 73–78. [Google Scholar] [PubMed]
- Richter-Levin, G.; Segal, M. Serotonin, aging and cognitive functions of the hippocampus. Rev. Neurosci. 1996, 7, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Ólafsdóttir, H.F.; Bush, D.; Barry, C. The Role of Hippocampal Replay in Memory and Planning. Curr. Biol. 2018, 28, R37–R50. [Google Scholar] [CrossRef]
- Jans, L.A.; Korte-Bouws, G.A.; Korte, S.M.; Blokland, A. The effects of acute tryptophan depletion on affective behaviour and cognition in Brown Norway and Sprague Dawley rats. J. Psychopharmacol. 2010, 24, 605–614. [Google Scholar] [CrossRef]
- Tousson, E.; Ibrahim, W.; Arafa, N.; Akela, M.A. Monoamine concentrations changes in the PTU-induced hypothyroid rat brain and the ameliorating role of folic acid. Hum. Exp. Toxicol. 2012, 31, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Bortolotto, V.C.; Pinheiro, F.C.; Araujo, S.M.; Poetini, M.R.; Bertolazi, B.S.; de Paula, M.T.; Meichtry, L.B.; de Almeida, F.P.; de Freitas Couto, S.; Jesse, C.R.; et al. Chrysin reverses the depressive-like behavior induced by hypothyroidism in female mice by regulating hippocampal serotonin and dopamine. Eur. J. Pharmacol. 2018, 822, 78–84. [Google Scholar] [CrossRef]
- Bocheva, G.; Landzhov, B.; Bozhilova-Pastirova, A.; Boyadjieva, N.; Krushkov, I. Effect of hypothyroidism on TSH-receptor expression in rats and its possible role in the pathogenesis of thyroid-associated dermatopathy. C. R. Acad. Bulg. Sci. 2007, 60, 805–808. [Google Scholar]
- Bocheva, G.; Varadinova, M.; Boyadjieva, N. Propylthiouracil causes behavior changes in rats: The role of serotonin. Ann. Gen. Psychiatry 2007, 7, 351. [Google Scholar] [CrossRef]
- Jones, M.L.; Kimbrough, T.D.; Weekley, L.B. Disturbances of Tryptophan Metabolism in Mice Acutely Deprived of Tryptophan. Ann. Nutr. Metab. 1985, 29, 209–215. [Google Scholar] [CrossRef]
- Hilaire, G.; Morin, D.; Lajard, A.M.; Monteau, R. Changes in serotonin metabolism may elicit obstructive apnoea in the newborn rat. J. Physiol. 1993, 466, 367–381. [Google Scholar]
- Rodriguez, A.; Zhang, H.; Klaminder, J.; Brodin, T.; Andersson, P.L.; Andersson, M. ToxTrac: A fast and robust software for tracking organisms. Methods Ecol. Evol. 2018, 9, 460–464. [Google Scholar] [CrossRef]
- Redei, E.E.; Solberg, L.C.; Kluczynski, J.M.; Pare, W.P. Paradoxical hormonal and behavioral responses to hypothyroid and hyperthyroid states in the Wistar-Kyoto rat. Neuropsychopharmacology 2001, 24, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Rial, R.V.; Tur, J.A.; Palmer, A.M.; Tur, J. Altered responsiveness to ambiental stimuli in altered thyroidal states. Physiol. Behav. 1987, 41, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Wilcoxon, J.S.; Nadolski, G.J.; Samarut, J.; Chassande, O.; Redei, E.E. Behavioral inhibition and impaired spatial learning and memory in hypothyroid mice lacking thyroid hormone receptor alpha. Behav. Brain Res. 2007, 177, 109–116. [Google Scholar] [CrossRef]
- Sapronov, N.S.; Fedotova, Y.O. The effect of L-tryptophan on conditioned reflex learning and behavior in rats with experimental pathology of the thyroid gland. Neurosci. Behav. Physiol. 2002, 32, 237–241. [Google Scholar] [CrossRef]
- Gerges, N.Z.; Alzoubi, K.H.; Park, C.R.; Diamond, D.M.; Alkadhi, K.A. Adverse effect of the combination of hypothyroidism and chronic psychosocial stress on hippocampus-dependent memory in rats. Behav. Brain Res. 2004, 155, 77–84. [Google Scholar] [CrossRef]
- Artis, A.S.; Bitiktas, S.; Taskin, E.; Dolu, N.; Liman, N.; Suer, C. Experimental hypothyroidism delays field excitatory post-synaptic potentials and disrupts hippocampal long-term potentiation in the dentate gyrus of hippocampal formation and Y-maze performance in adult rats. J. Neuroendocrinol. 2012, 24, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Koromilas, C.; Liapi, C.; Schulpis, K.H.; Kalafatakis, K.; Zarros, A.; Tsakiris, S. Structural and functional alterations in the hippocampus due to hypothyroidism. Metab. Brain Dis. 2010, 25, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Chaalal, A.; Poirier, R.; Blum, D.; Gillet, B.; Le Blanc, P.; Basquin, M.; Buée, L.; Laroche, S.; Enderlin, V. PTU-induced hypothyroidism in rats leads to several early neuropathological signs of Alzheimer’s disease in the hippocampus and spatial memory impairments. Hippocampus 2014, 24, 1381–1393. [Google Scholar] [CrossRef] [PubMed]
- Ghenimi, N.; Alfos, S.; Redonnet, A.; Higueret, P.; Pallet, V.; Enderlin, V. Adult-onset hypothyroidism induces the amyloidogenic pathway of amyloid precursor protein processing in the rat hippocampus. J. Neuroendocrinol. 2010, 22, 951–959. [Google Scholar] [PubMed]
- Yu, J.; Tang, Y.Y.; Feng, H.B.; Cheng, X.X. A behavioral and micro positron emission tomography imaging study in a rat model of hypothyroidism. Behav. Brain Res. 2014, 271, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, R.; Gourion, D.; Llorca, P.M. L’anhédonie dans la dépression [Anhedonia in depression]. Encephale 2013, 39, 296–305. [Google Scholar] [CrossRef]
- Antunes, M.; Biala, G. The novel object recognition memory: Neurobiology, test procedure, and its modifications. Cogn. Process. 2012, 13, 93–110. [Google Scholar] [CrossRef]
- de Lima, M.N.; Luft, T.; Roesler, R.; Schröder, N. Temporary inactivation reveals an essential role of the dorsal hippocampus in consolidation of object recognition memory. Neurosci. Lett. 2006, 405, 142–146. [Google Scholar] [CrossRef]
- Rastogi, R.B.; Singhal, R.L. The effect of thyroid hormone on serotonergic neurones: Depletion of serotonin in discrete brain areas of developing hypothyroid rats. Naunyn Schmiedebergs Arch. Pharmacol. 1978, 304, 9–13. [Google Scholar] [CrossRef]
- Meaney. M.J.; Diorio, J.; Francis, D.; Weaver, S.; Yau, J.; Chapman, K.; Seckl, J.R. Postnatal Handling Increases the Expression of cAMP-Inducible Transcription Factors in the Rat Hippocampus: The Effects of Thyroid Hormones and Serotonin. J. Neurosci. 2000, 20, 3926–3935. [Google Scholar] [CrossRef]
- Savard, P.; Mérand, Y.; Di Paolo, T.; Dupont, A. Effect of neonatal hypothyroidism on the serotonin system of the rat brain. Brain Res. 1984, 292, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.B.; Singhal, R.L. Influence of neonatal and adult hyperthyroidism on behavior and biosynthetic capacity for norepinephrine, dopamine and 5-hydroxytryptamine in rat brain. J. Pharmacol. Exp. Ther. 1976, 198, 609–618. [Google Scholar] [PubMed]
- Henley, W.N.; Chen, X.; Klettner, C.; Bellush, L.L.; Notestine, M.A. Hypothyroidism increases serotonin turnover and sympathetic activity in the adult rat. Can. J. Physiol. Pharmacol. 1991, 69, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, S.; Bakalov, D.; Bocheva, G. Pathophysiology and Management Possibilities of Thyroid-Associated Depression. Acta Med. Bulg. 2022, 49, 68–72. [Google Scholar] [CrossRef]
- Ogren, S.O.; Eriksson, T.M.; Elvander-Tottie, E.; D’Addario, C.; Ekström, J.C.; Svenningsson, P.; Meister, B.; Kehr, J.; Stiedl, O. The role of 5-HT(1A) receptors in learning and memory. Behav. Brain Res. 2008, 195, 54–77. [Google Scholar] [CrossRef]
- Elvander-Tottie, E.; Eriksson, T.M.; Sandin, J.; Ogren, S.O. 5-HT(1A) and NMDA receptors interact in the rat medial septum and modulate hippocampal-dependent spatial learning. Hippocampus 2009, 19, 1187–1198. [Google Scholar] [CrossRef]
- Sumiyoshi, T.; Bubenikova-Valesova, V.; Horacek, J.; Bert, B. Serotonin1A receptors in the pathophysiology of schizophrenia: Development of novel cognition-enhancing therapeutics. Adv. Ther. 2008, 25, 1037–1056. [Google Scholar] [CrossRef]
- Friedman, M. Analysis, Nutrition, and Health Benefits of Tryptophan. Int. J. Tryptophan Res. 2018, 11, 1178646918802282. [Google Scholar] [CrossRef]
- Haider, S.; Khaliq, S.; Haleem, D.J. Enhanced serotonergic neurotransmission in the hippocampus following tryptophan administration improves learning acquisition and memory consolidation in rats. Pharmacol. Rep. 2007, 59, 53–57. [Google Scholar]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakalov, D.; Iliev, P.; Sabit, Z.; Tafradjiiska-Hadjiolova, R.; Bocheva, G. Attenuation of Hypothyroidism-Induced Cognitive Impairment by Modulating Serotonin Mediation. Vet. Sci. 2023, 10, 122. https://doi.org/10.3390/vetsci10020122
Bakalov D, Iliev P, Sabit Z, Tafradjiiska-Hadjiolova R, Bocheva G. Attenuation of Hypothyroidism-Induced Cognitive Impairment by Modulating Serotonin Mediation. Veterinary Sciences. 2023; 10(2):122. https://doi.org/10.3390/vetsci10020122
Chicago/Turabian StyleBakalov, Dimitar, Petar Iliev, Zafer Sabit, Radka Tafradjiiska-Hadjiolova, and Georgeta Bocheva. 2023. "Attenuation of Hypothyroidism-Induced Cognitive Impairment by Modulating Serotonin Mediation" Veterinary Sciences 10, no. 2: 122. https://doi.org/10.3390/vetsci10020122
APA StyleBakalov, D., Iliev, P., Sabit, Z., Tafradjiiska-Hadjiolova, R., & Bocheva, G. (2023). Attenuation of Hypothyroidism-Induced Cognitive Impairment by Modulating Serotonin Mediation. Veterinary Sciences, 10(2), 122. https://doi.org/10.3390/vetsci10020122