
Effects of Low Protein Diet on Production Performance and Intestinal Microbial Composition in Pigs

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design, Animals, and Diets
2.2. Animal Performance Indicators
2.3. Intestinal Microbial Composition Analysis
2.4. Data Processing and Analysis
3. Results
3.1. Effect of Dietary Protein Level on the Growth Performance of Hexi Pigs
3.2. Effects of Dietary Protein Levels on Production Performance and Meat Quality of Hexi Pigs
3.3. 16S rRNA Sequencing of the Jejunum and Cecum of Fattening Pigs under Different Dietary Protein Levels
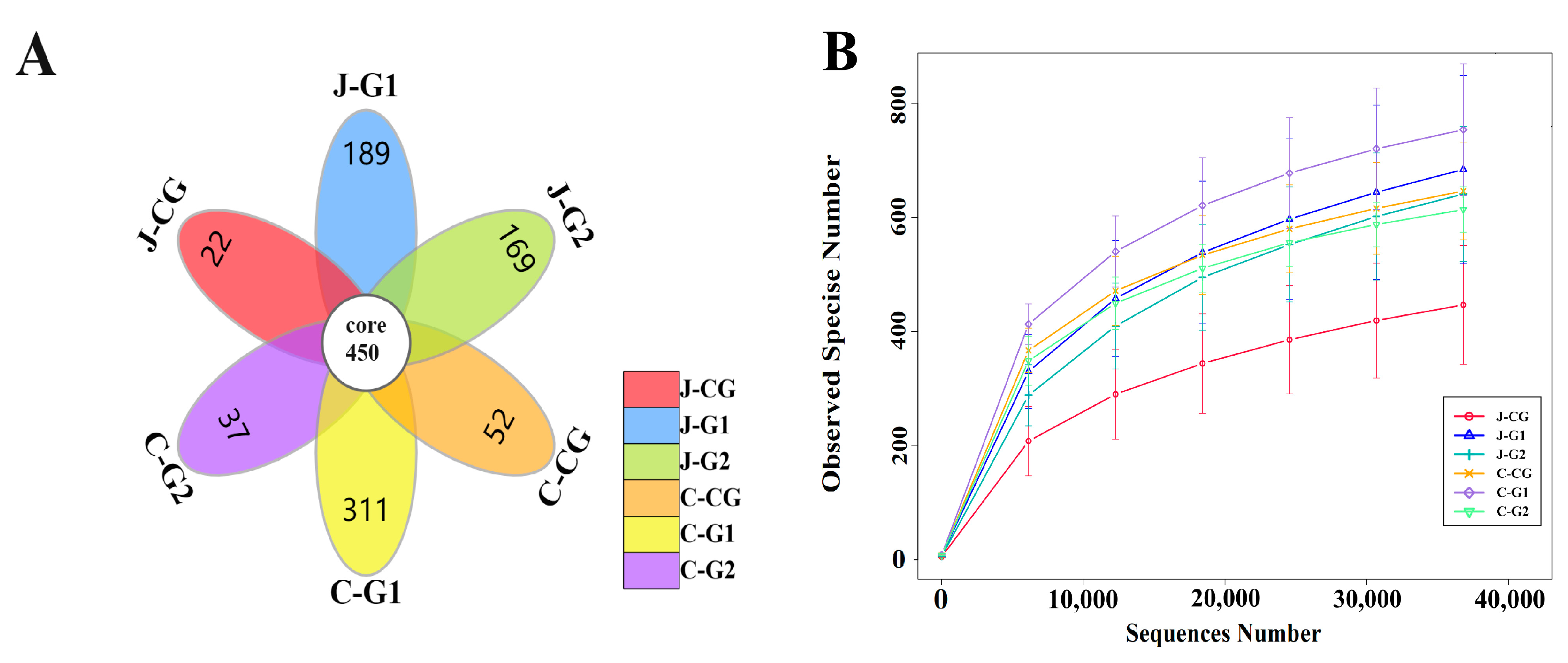
3.4. Analysis of the Microbial Diversity in the Jejunum and Cecum of Fattening Pigs under Different Dietary Protein Levels
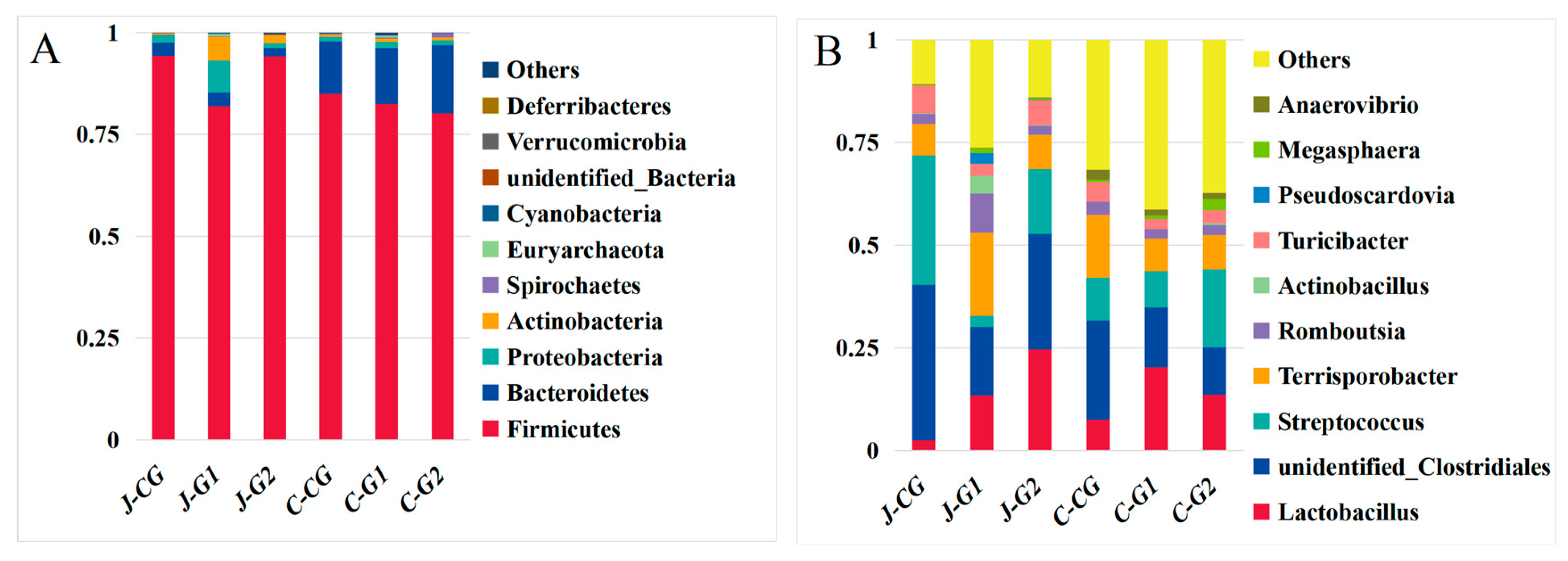
3.5. Analysis of the Microbial Composition of the Jejunum and Cecum of Fattening Pigs under Different Dietary Protein Levels
3.6. Gene Function Prediction
4. Discussion
4.1. Effect of the Dietary Protein Level on the Growth Performance of Hexi Pigs
4.2. The Effect of the Dietary Protein Level on the Production Performance and Meat Quality of Hexi Pigs
4.3. The Effect of the Dietary Protein Level on the Diversity and Abundance of Intestinal Flora in Hexi Pigs
4.4. The Effect of the Dietary Protein Level on the Composition of Intestinal Flora and KEGG Function of Hexi Pigs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oliveira, C.H.; Bernardes, R.D.; Dias, K.M.M.; Ribeiro, A.M.; Rodrigueiro, R.J.B.; Koo, B.K.; Tak, J.; Park, C.; Calderano, A.A.; Albino, L.F.T. Research Note: The influence of different isoleucine: Lysine ratios on the growth performance of broiler chickens fed low-protein diets. Poult. Sci. 2023, 102, 102270. [Google Scholar] [CrossRef] [PubMed]
- Rocha, G.C.; Duarte, M.E.; Kim, S.W. Advances, Implications, and Limitations of Low-Crude-Protein Diets in Pig Production. Animals 2022, 12, 3478. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Han, S.; Zhou, J.; Li, P.; Wang, G.; Yu, H.; Cai, S.; Zeng, X.; Johnston, L.J.; Levesque, C.L.; et al. Effects of dietary crude protein level and N-carbamylglutamate supplementation on nutrient digestibility and digestive enzyme activity of jejunum in growing pigs. J. Anim. Sci. 2020, 98, skaa088. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.; Liang, C.; Chen, D.; Tian, G.; Zheng, P.; He, J.; Yu, J.; Mao, X.; Gu, Z.; Yang, W.; et al. Effects of low-energy diet supplemented with betaine on growth performance, nutrient digestibility, and serum metabolomic profiles in growing pigs. J. Anim. Sci. 2023, 101, skad080. [Google Scholar] [CrossRef]
- Gloaguen, M.; Le, F.N.; Corrent, E.; Primot, Y.; Milgen, J. The use of free amino acids allows formulating very low crude protein diets for piglets. J. Anim. Sci. 2014, 92, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Mi, M.M.; Zhang, Q.Y.; Bao, N.; Pan, L.; Zhao, Y.; Qin, G.X. Relationship between the amino acid release kinetics of feed proteins and nitrogen balance in finishing pigs. Animal 2021, 15, 100359. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, H.; Wan, K.; Zhou, K.; Wang, Y.; Li, J.; Tang, Z.; Sun, W.; Wu, L.; An, R.; et al. Effects of supplementing low-protein diets with sodium dichloroacetate and glucose on growth performance, carcass traits, and meat quality of growing-finishing pigs. J. Anim. Sci. 2022, 100, skab359. [Google Scholar] [CrossRef]
- Shriver, J.A.; Carter, S.D.; Sutton, A.L.; Richert, B.T.; Senne, B.W.; Pettey, L.A. Effects of adding fiber sources to reduced-crude protein, amino acid-supplemented diets on nitrogen excretion, growth performance, and carcass traits of finishing pigs. J. Anim. Sci. 2003, 81, 492–502. [Google Scholar] [CrossRef]
- Isaacson, R.; Kim, H.B. The intestinal microbiome of the pig. Anim. Health Res. Rev. 2012, 13, 100–109. [Google Scholar] [CrossRef]
- Li, F.; Li, C.; Chen, Y.; Liu, J.; Zhang, C.; Irving, B.; Fitzsimmons, C.; Plastow, G.; Guan, L.L. Host genetics influence the rumen microbiota and heritable rumen microbial features associate with feed efficiency in cattle. Microbiome 2019, 7, 92. [Google Scholar] [CrossRef]
- Ma, N.; Tian, Y.; Wu, Y.; Ma, X. Contributions of the Interaction Between Dietary Protein and Gut Microbiota to Intestinal Health. Curr. Protein Pept. Sci. 2017, 18, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Bikker, P.; Dirkzwager, A.; Fledderus, J.; Trevisi, P.; le Huërou-Luron, I.; Lallès, J.P.; Awati, A. The effect of dietary protein and fermentable carbohydrates levels on growth performance and intestinal characteristics in newly weaned piglets. J. Anim. Sci. 2006, 84, 3337–3345. [Google Scholar] [CrossRef]
- Abdallah, A.; Elemba, E.; Zhong, Q.; Sun, Z. Gastrointestinal Interaction between Dietary Amino Acids and Gut Microbiota: With Special Emphasis on Host Nutrition. Curr. Protein Pept. Sci. 2020, 21, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Davila, A.M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.H.; Sanz, Y.; Tomé, D. Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host. Pharmacol. Res. 2013, 68, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Bishu, S. Sensing of nutrients and microbes in the gut. Curr. Opin. Gastroenterol. 2016, 32, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Y.; Tian, Y.; Huang, C.; Li, D.; Zhong, Q.; Ma, X. Interaction between Microbes and Host Intestinal Health: Modulation by Dietary Nutrients and Gut-Brain-Endocrine-Immune Axis. Curr. Protein Pept. Sci. 2015, 16, 592–603. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, J.; Wang, G.; Cai, S.; Zeng, X.; Qiao, S. Advances in low-protein diets for swine. J. Anim. Sci. Biotechnol. 2018, 9, 60. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, X.; Tang, Z.; Li, Y.; Li, T.; Xu, Q.; Zhen, J.; Huang, F.; Yang, J.; Chen, C.; et al. Low-Protein Diets Decrease Porcine Nitrogen Excretion but with Restrictive Effects on Amino Acid Utilization. J. Agric. Food Chem. 2018, 66, 8262–8271. [Google Scholar] [CrossRef]
- Yu, D.; Zhu, W.; Hang, S. Effects of low-protein diet on the intestinal morphology, digestive enzyme activity, blood urea nitrogen, and gut microbiota and metabolites in weaned pigs. Arch Anim. Nutr. 2019, 73, 287–305. [Google Scholar] [CrossRef]
- Luo, Z.; Cheng, Y.; Zhu, W. The effect of low-protein diet on the cecal metabolites and flora of fattening pigs. Anim. Husb. Vet. Med. 2015, 47, 5–9. Available online: https://kns.cnki.net/kcms2/article/abstract?v=5DzVwdTmeh8X5Z3Wr5qTUaWSZU-0mM3B0JOP6xpV4aoF6ZLol5jtUecwIC0K8OLjYeFf02zMDGt8Fz7f0gkkqIiLT9dXxycdTJevhnW-2ZYK6-rfLUMxUvDUCHg6nuIYESxuYUsUPuY=&uniplatform=NZKPT&flag=copy (accessed on 5 May 2015). (In Chinese).
- Wang, H.; Shen, J.; Pi, Y.; Gao, K.; Zhu, W. Low-protein diets supplemented with casein hydrolysate favor the microbiota and enhance the mucosal humoral immunity in the colon of pigs. J. Anim. Sci. Biotechnol. 2019, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, G.; Song, L.; Chai, M.; Wang, Y.; Shui, S.; Zhang, H.; Sha, Y.; Yao, Y. Effects of Dietary Protein Levels on Bamei Pig Intestinal Colony Compositional Traits. Biomed. Res. Int. 2020, 2020, 2610431. [Google Scholar] [CrossRef] [PubMed]
- Local Pig Breed—Hexi Pig. Rural. New Technol. 2015, 5, 41. Available online: https://kns.cnki.net/kcms2/article/abstract?v=3uoqIhG8C44YLTlOAiTRKibYlV5Vjs7ir5D84hng_y4D11vwp0rrtZAu8Dj24nQqEwdKu6W35Mlj9Q8E-1IazVSgVLx9C2eF&uniplatform=NZKPT (accessed on 5 May 2015).
- Wang, D.; Chen, G.; Chai, M.; Shi, C.; Geng, Y.; Che, Y.; Li, Y.; Liu, S.; Gao, Y.; Hou, H. Effects of dietary protein levels on production performance, meat quality and flavor of fattening pigs. Front. Nutr. 2022, 9, 910519. [Google Scholar] [CrossRef]
- GB/T 19479-2019; Good Manufacturing Practice for Livestock and Poultry Slaughtering—Pig. State Administration for Market Regulation, Standardization administration: Beijing, China, 2019.
- Wu, L.Y. The Effect of Microecological Preparations and Achyranthesbidentata Polysaccharides on Pig Growth Performance and Meat Quality. Master’s Thesis, Hunan Agricultural University, Changsha, China, 2011. Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=CMFD2012&filename=1011204036.nh (accessed on 5 May 2011).
- Li, Y.H.; Li, F.N.; Duan, Y.H.; Guo, Q.P.; Wen, C.Y.; Wang, W.L.; Huang, X.G.; Yin, Y.L. Low-protein diet improves meat quality of growing and finishing pigs through changing lipid metabolism, fiber characteristics, and free amino acid profile of the muscle. J. Anim. Sci. 2018, 96, 3221–3232. [Google Scholar] [CrossRef]
- Skrlep, M.; Poklukar, K.; Kress, K.; Vrecl, M.; Fazarinc, G.; Batorek Lukac, N.; Weiler, U.; Stefanski, V.; Candek-Potokar, M. Effect of immunocastration and housing conditions on pig carcass and meat quality traits. Transl. Anim. Sci. 2020, 4, txaa055. [Google Scholar] [CrossRef] [PubMed]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Werner, J.J.; Zhou, D.; Caporaso, J.G.; Knight, R.; Angenent, L.T. Comparison of Illumina paired-end and single-direction sequencing for microbial 16S rRNA gene amplicon surveys. ISME J. 2012, 6, 1273–1276. [Google Scholar] [CrossRef]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Tuitoek, K.; Young, L.G.; de Lange, C.F.; Kerr, B.J. The effect of reducing excess dietary amino acids on growing-finishing pig performance: An elevation of the ideal protein concept. J. Anim. Sci. 1997, 75, 1575–1583. [Google Scholar] [CrossRef] [PubMed]
- Prandini, A.; Sigolo, S.; Morlacchini, M.; Grilli, E.; Fiorentini, L. Microencapsulated lysine and low-protein diets: Effects on performance, carcass characteristics and nitrogen excretion in heavy growing-finishing pigs. J. Anim. Sci. 2013, 91, 4226–4234. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, L.; Zhou, J.; Zeng, X.; Qiao, S. Effects of using cassava as an amylopectin source in low protein diets on growth performance, nitrogen efficiency, and postprandial changes in plasma glucose and related hormones concentrations of growing pigs. J. Anim. Sci. 2021, 99, skab332. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.M.; Bai, G.D.; Xu, X.; He, W.; Yang, Z.; Gao, F. Effects of adding tyrosine to low protein diets on the growth performance and nitrogen metabolism of fattening pigs. J. Northeast. Agric. Univ. 2019, 50, 58–66. [Google Scholar] [CrossRef]
- Xie, C.; Zhang, S.; Zhang, G.; Zhang, F.; Chu, L.; Qiao, S. Estimation of the optimal ratio of standardized ileal digestible threonine to lysine for finishing barrows fed low crude protein diets. Asian-Australas J. Anim. Sci. 2013, 26, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Zhang, L.; Li, J.; Luo, Y.; Zhang, B.; Xing, S.; Zhu, Y.; Sun, H.; Gao, F.; Zhou, G. Effects of Dietary Crude Protein Levels and Cysteamine Supplementation on Protein Synthetic and Degradative Signaling in Skeletal Muscle of Finishing Pigs. PLoS ONE 2015, 10, e0139393. [Google Scholar] [CrossRef]
- Norgaard, J.V.; Hansen, M.J.; Soumeh, E.A.; Adamsen, A.P.S.; Poulsen, H.D. Effect of protein level on performance, nitrogen utilisation and carcass composition in finisher pigs. Acta Agric. Scand. A Anim. Sci. 2014, 64, 123–129. [Google Scholar] [CrossRef]
- Qin, C.; Huang, P.; Qiu, K.; Sun, W.; Xu, L.; Zhang, X.; Yin, J. Influences of dietary protein sources and crude protein levels on intracellular free amino acid profile in the longissimus dorsi muscle of finishing gilts. J. Anim. Sci. Biotechnol. 2015, 6, 52. [Google Scholar] [CrossRef]
- Liu, S.; Xie, J.; Fan, Z.; Ma, X.; Yin, Y. Effects of low protein diet with a balanced amino acid pattern on growth performance, meat quality and cecal microflora of finishing pigs. J. Sci. Food Agric. 2023, 103, 957–967. [Google Scholar] [CrossRef]
- Zhu, Y.P.; Zhou, P.; Li, J.L.; Zhang, L.; Gao, F.; Zhou, G.H. Effects of Low Protein Level Diets Supplemented with Essential Amino Acids and Cysteamine on Meat Quality and Related Genes Expression of Growing Pigs. Acta Vet. Et Zootech. Sin. 2017, 48, 660–668. [Google Scholar] [CrossRef]
- Zhang, S.; Chu, L.; Qiao, S.; Mao, X.; Zeng, X. Effects of dietary leucine supplementation in low crude protein diets on performance, nitrogen balance, whole-body protein turnover, carcass characteristics and meat quality of finishing pigs. Anim. Sci. J. 2016, 87, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Bidner, B.S.; Ellis, M.; Witte, D.P.; Carr, S.N.; McKeith, F.K. Influence of dietary lysine level, pre-slaughter fasting, and rendement napole genotype on fresh pork quality. Meat Sci. 2004, 68, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Goerl, K.F.; Eilert, S.J.; Mandigo, R.W.; Chen, H.Y.; Miller, P.S. Pork characteristics as affected by two populations of swine and six crude protein levels. J. Anim. Sci. 1995, 73, 3621–3626. [Google Scholar] [CrossRef]
- Ruusunen, M.; Partanen, K.; Pösö, R.; Puolannea, E. The effect of dietary protein supply on carcass composition, size of organs, muscle properties and meat quality of pigs. Livest. Sci. 2006, 107, 170–181. [Google Scholar] [CrossRef]
- Li, N.; Xie, C.Y.; Zeng, X.F.; Wang, D.H.; Qiao, S.Y. Effects of dietary crude protein level and amino acid balance on growth performance, carcass traits and meat quality of finishing pigs. Chin. J. Anim. Nutr. 2018, 30, 498–506. [Google Scholar] [CrossRef]
- Guo, X.; Xia, X.; Tang, R.; Zhou, J.; Zhao, H.; Wang, K. Development of a real-time PCR method for Firmicutes and Bacteroidetes in faeces and its application to quantify intestinal population of obese and lean pigs. Lett. Appl. Microbiol. 2008, 47, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef]
- Matsui, T.; Tanaka, J.; Namihira, T.; Shinzato, N. Antibiotics production by an actinomycete isolated from the termite gut. J. Basic Microbiol. 2012, 52, 731–735. [Google Scholar] [CrossRef]
- Htoo, J.K.; Araiza, B.A.; Sauer, W.C.; Rademacher, M.; Zhang, Y.; Cervantes, M.; Zijlstra, R.T. Effect of dietary protein content on ileal amino acid digestibility, growth performance, and formation of microbial metabolites in ileal and cecal digesta of early-weaned pigs. J. Anim. Sci. 2007, 85, 3303–3312. [Google Scholar] [CrossRef]
- Konstantinov, S.R.; Favier, C.F.; Zhu, W.Y.; Williams, B.A.; Klüß, J.; Souffrant, W.B.; de Vos, W.M.; Akkermans, A.D.; Smidt, H. Microbial diversity studies of the porcine gastrointestinal ecosystem during weaning transition. Anim. Res. 2005, 53, 317–324. [Google Scholar] [CrossRef]
- Fan, P.; Liu, P.; Song, P.; Chen, X.; Ma, X. Moderate dietary protein restriction alters the composition of gut microbiota and improves ileal barrier function in adult pig model. Sci. Rep. 2017, 7, 43412. [Google Scholar] [CrossRef] [PubMed]
- Rist, V.T.; Weiss, E.; Eklund, M.; Mosenthin, R. Impact of dietary protein on microbiota composition and activity in the gastrointestinal tract of piglets in relation to gut health: A review. Animal 2013, 7, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.B. Possible ways of modifying type and amount of products from microbial fermentation in the gut. In Gut Environment of Pigs; Piva, A., Knudsen, K.E.B., Lindberg, J.E., Eds.; Nottingham University Press: Nottingham, UK, 2001; pp. 181–200. [Google Scholar]
- Zhao, J.; Zhang, X.; Liu, H.; Brown, M.A.; Qiao, S. Dietary Protein and Gut Microbiota Composition and Function. Curr. Protein Pept. Sci. 2019, 20, 145–154. [Google Scholar] [CrossRef]
- Greenhill, C. Gut microbiota: Firmicutes and Bacteroidetes involved in insulin resistance by mediating levels of glucagon-like peptide 1. Nat. Rev. Endocrinol. 2015, 11, 254. [Google Scholar] [CrossRef]
- Shoaie, S.; Karlsson, F.; Mardinoglu, A.; Nookaew, I.; Bordel, S.; Nielsen, J. Understanding the interactions between bacteria in the human gut through metabolic modeling. Sci. Rep. 2013, 3, 2532. [Google Scholar] [CrossRef]
- Opapeju, F.O.; Krause, D.O.; Payne, R.L.; Rademacher, M.; Nyachoti, C.M. Effect of dietary protein level on growth performance, indicators of enteric health, and gastrointestinal microbial ecology of weaned pigs induced with postweaning colibacillosis. J. Anim. Sci. 2009, 87, 2635–2643. [Google Scholar] [CrossRef]
- Fan, P. The Effect of Low-Protein Diets on the Gut Microflora of Weaned Piglets and Fattening Pigs. Master’s Thesis, China Agricultural University, Beijing, China, 2016. Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?filename=1016085516.nh&dbname=CMFDTEMP (accessed on 1 May 2016).
- Xia, Y.; Zhang, Y.; Xu, J.; Guo, S.; Ding, B. Effects of Lactobacillus fermentum and Bacillus coagulans on growth performance and intestinal health of broilers infected by Clostridium perfringens. China Anim. Husb. Vet. Med. 2019, 46, 2927–2936. [Google Scholar] [CrossRef]
- Wang, J.; Wang, S.; Liu, H.; Zhang, W.; Zhang, D.; Wang, Y.; Ji, H. Effects of low protein level diets on the growth performance and intestinal flora of growing-finishing pigs. J. Anim. Nutr 2018, 30, 4864–4873. [Google Scholar] [CrossRef]
- Devillard, E.; McIntosh, F.M.; Duncan, S.H.; Wallace, R.J. Metabolism of linoleic acid by human gut bacteria: Different routes for biosynthesis of conjugated linoleic acid. J. Bacteriol. 2007, 189, 2566–2570. [Google Scholar] [CrossRef]
- Fan, P.; Li, L.; Rezaei, A.; Eslamfam, S.; Che, D.; Ma, X. Metabolites of Dietary Protein and Peptides by Intestinal Microbes and their Impacts on Gut. Curr. Protein Pept. Sci. 2015, 16, 646–654. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Basic Diet Composition (%) | |||
|---|---|---|---|
| Raw Material | CG | G1 | G2 |
| Corn | 63.50 | 68.00 | 72.00 |
| Soybean meal | 18.30 | 12.90 | 7.20 |
| Wheat bran | 5.00 | 5.00 | 5.00 |
| Alfalfa meal | 5.00 | 6.50 | 9.00 |
| Bentonite | 4.00 | 2.90 | 1.60 |
| Soybean oil | 1.50 | 1.70 | 2.00 |
| Compound Enzyme preparation | 0.10 | 0.10 | 0.10 |
| Lys | 0.09 | 0.23 | 0.37 |
| Met | - | 0.03 | 0.05 |
| Thr | - | 0.08 | 0.15 |
| Try | - | 0.03 | 0.05 |
| CaCO3 | 0.51 | 0.46 | 0.30 |
| CaHPO4 | 1.15 | 1.22 | 1.33 |
| 0.5% fattening Pig core feed ① | 0.50 | 0.50 | 0.50 |
| NaCl | 0.35 | 0.35 | 0.35 |
| Total | 100.00 | 100.00 | 100.00 |
| Nutrient levels ② | |||
| DE/(MJ/kg) | 13.14 | 13.13 | 13.11 |
| CP | 15.94 (16.00% ③) | 14.00 (14.00% ③) | 12.05 (12.00% ③) |
| CF | 3.22 | 3.50 | 4.02 |
| Ca | 0.60 | 0.61 | 0.60 |
| TP | 0.55 | 0.55 | 0.55 |
| Na | 0.16 | 0.16 | 0.17 |
| Cl | 0.27 | 0.27 | 0.28 |
| SID Lys | 0.86 | 0.86 | 0.86 |
| SID Met | 0.26 | 0.26 | 0.26 |
| SID Thr | 0.59 | 0.59 | 0.59 |
| SID Try | 0.19 | 0.19 | 0.19 |
| Items | CG | G1 | G2 | p-Value |
|---|---|---|---|---|
| Initial body weight/kg (IW) | 61.68 ± 2.16 | 61.45 ± 1.95 | 61.58 ± 2.19 | 0.788 |
| Final body weight/kg (FW) | 111.48 ± 3.26 | 113.05 ± 2.81 | 110.78 ± 3.22 | 0.498 |
| Average daily feed intake/kg·day−1 (ADFI) | 3.02 ± 0.15 | 2.97 ± 0.16 | 3.20 ± 0.13 | 0.387 |
| Average daily gain/kg·day−1 (ADG) | 0.83 ± 0.03 | 0.86 ± 0.05 | 0.82 ± 0.06 | 0.169 |
| Ratio of feed to gain(F/G) | 3.64 ± 0.06 | 3.45 ± 0.11 | 3.90 ± 0.15 | 0.125 |
| Items | CG | G1 | G2 | p-Value |
|---|---|---|---|---|
| Slaughter rate/% | 67.99 ± 0.02 | 69.68 ± 0.33 | 72.19 ± 2.96 | 0.055 |
| Backfat thickness/mm | 36.85 ± 5.72 ab | 23.80 ± 4.31 b | 31.83 ± 11.13 a | 0.006 |
| Carcass length/cm | 115.83 ± 4.44 | 119.13 ± 9.50 | 118.00 ± 4.58 | 0.225 |
| Loin eye area/cm2 | 58.53 ± 7.56 a | 44.08 ± 6.20 ab | 30.75 ± 9.25 b | 0.013 |
| Drip loss/% | 5.45 ± 1.15 b | 7.53 ± 0.69 a | 4.82 ± 1.74 b | 0.005 |
| Cooking loss/% | 35.46 ± 4.07 a | 26.94 ± 7.53 b | 27.98 ± 8.33 ab | 0.005 |
| Cooking percentage/% | 64.54 ± 2.97 | 73.06 ± 8.56 | 72.02 ± 9.29 | 0.402 |
| Filtration rate/% | 6.34 ± 1.07 | 7.11 ± 1.38 | 7.21 ± 7.12 | 0.126 |
| Shear force/N | 44.50 ± 7.14 | 43.42 ± 6.69 | 37.57 ± 5.27 | 0.614 |
| pH45min | 6.23 ± 0.23 a | 5.88 ± 0.16 b | 6.03 ± 0.33 ab | 0.023 |
| pH24h | 5.30 ± 0.06 b | 5.34 ± 0.04 b | 5.66 ± 0.31 a | <0.01 |
| pH48h | 5.21 ± 0.09 b | 5.22 ± 0.15 b | 5.61 ± 0.26 a | 0.002 |
| L45min | 34.42 ± 2.80 | 39.00 ± 5.95 | 40.62 ± 5.09 | 0.097 |
| a45min | 13.73 ± 2.48 | 13.20 ± 1.93 | 12.42 ± 1.66 | 0.522 |
| b45min | 7.12 ± 1.32 | 7.67 ± 1.73 | 8.05 ± 1.16 | 0.508 |
| Items | CG | G1 | G2 | p-Value |
|---|---|---|---|---|
| Jejunum | ||||
| ACE | 605.44 ± 148.78 | 899.95 ± 280.22 | 870.83 ± 214.62 | 0.275 |
| Chao1 | 584.91 ± 141.34 | 910.85 ± 295.45 | 949.92 ± 376.38 | 0.306 |
| Simpson | 0.76 ± 0.03 | 0.90 ± 0.06 | 0.80 ± 0.07 | 0.058 |
| Shannon | 3.15 ± 0.56 b | 4.75 ± 0.46 a | 3.77 ± 0.65 ab | 0.035 |
| Cecum | ||||
| ACE | 795.33 ± 120.47 | 900.93 ± 173.35 | 731.85 ± 58.40 | 0.324 |
| Chao1 | 798.85 ± 120.28 | 879.10 ± 147.68 | 723.18 ± 42.19 | 0.309 |
| Simpson | 0.91 ± 0.01 | 0.94 ± 0.02 | 0.93 ± 0.03 | 0.430 |
| Shannon | 5.25 ± 0.11 | 5.70 ± 0.40 | 5.46 ± 0.59 | 0.459 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Chen, G.; Li, W.; Chai, M.; Zhang, H.; Su, Y. Effects of Low Protein Diet on Production Performance and Intestinal Microbial Composition in Pigs. Vet. Sci. 2023, 10, 655. https://doi.org/10.3390/vetsci10110655
Wang D, Chen G, Li W, Chai M, Zhang H, Su Y. Effects of Low Protein Diet on Production Performance and Intestinal Microbial Composition in Pigs. Veterinary Sciences. 2023; 10(11):655. https://doi.org/10.3390/vetsci10110655
Chicago/Turabian StyleWang, Dong, Guoshun Chen, Wenzhong Li, Mingjie Chai, Hua Zhang, and Yingyu Su. 2023. "Effects of Low Protein Diet on Production Performance and Intestinal Microbial Composition in Pigs" Veterinary Sciences 10, no. 11: 655. https://doi.org/10.3390/vetsci10110655
APA StyleWang, D., Chen, G., Li, W., Chai, M., Zhang, H., & Su, Y. (2023). Effects of Low Protein Diet on Production Performance and Intestinal Microbial Composition in Pigs. Veterinary Sciences, 10(11), 655. https://doi.org/10.3390/vetsci10110655