Pregnancy Complications and Feto-Maternal Monitoring in Rabbits


 ,
, 
Simple Summary
Abstract
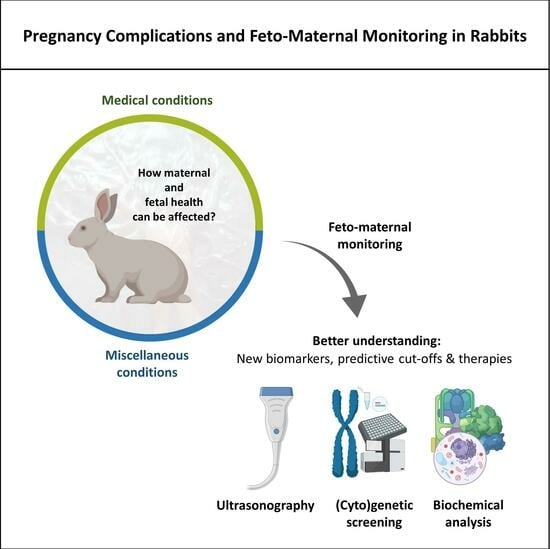
1. Introduction
2. Medical Conditions during Pregnancy: How Do They Affect Its Course?
2.1. Chronic Maternal Hypertension
2.2. Gestational Diabetes
2.3. Ectopic Gestation
2.4. Maternal Uterine Ischemia and Fetal Hypoxia
2.5. Intrauterine Growth Restriction
2.6. Superfetation
3. Other Factors That Can Influence Fetal Development and Offspring Health and Behavior
3.1. Maternal Age
3.2. Maternal Stress
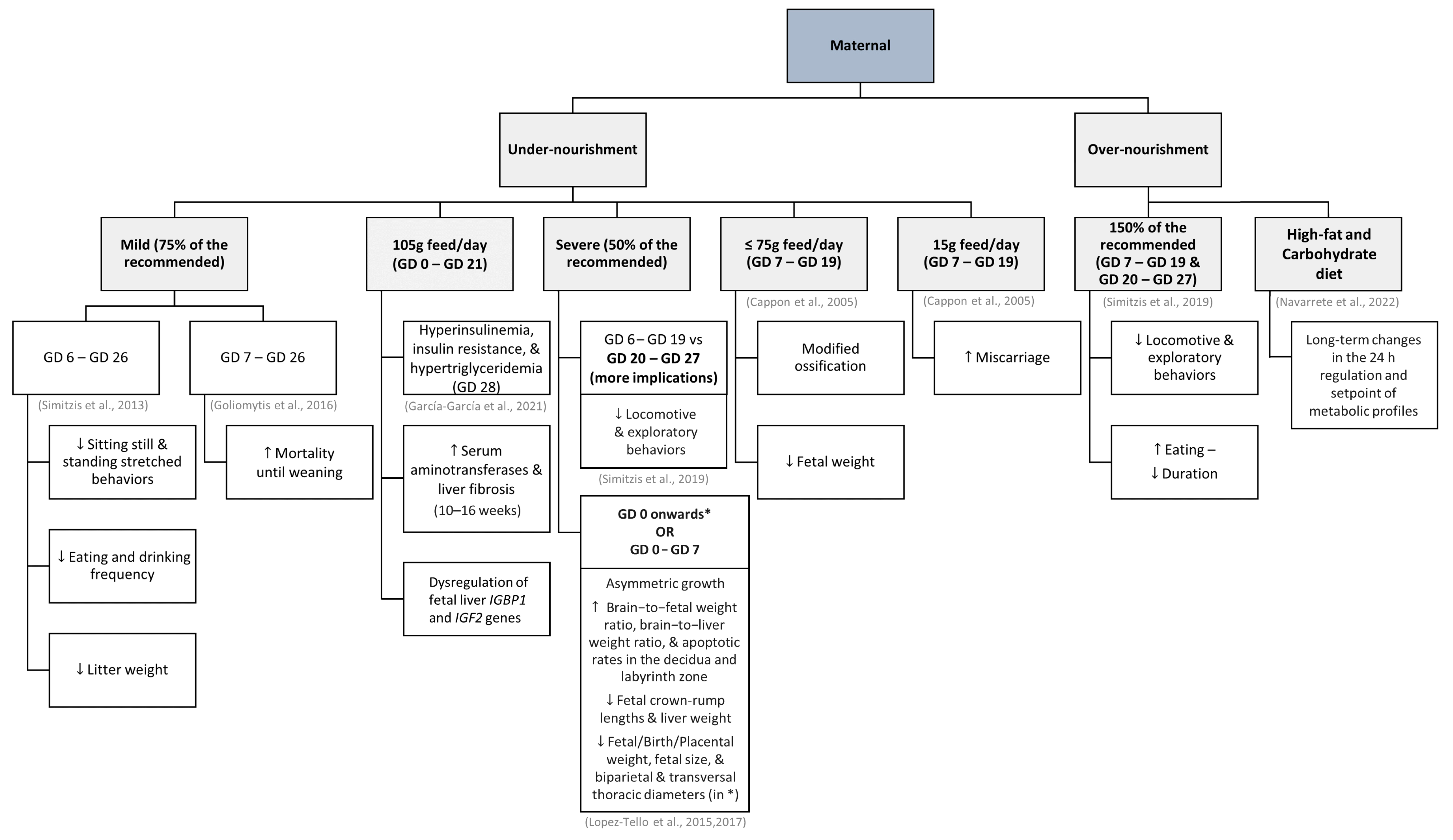
3.3. Maternal Nutritional Status
3.4. Maternal Physical Condition
3.5. Maternal and Embryonic Genotype
3.6. Intrauterine Location
4. Monitoring Techniques
4.1. Ultrasonography
4.2. Precautions before and during the Examination
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Commission. Commercial Rabbit Farming in the European Union; European Commission: Luxembourg, 2017. [Google Scholar]
- Zamaratskaia, G.; Havrysh, O.; Korzeniowska, M.; Getya, A. Potential and Limitations of Rabbit Meat in Maintaining Food Security in Ukraine. Meat Sci. 2023, 204, 109293. [Google Scholar] [CrossRef]
- Food and Agriculture Organization UNdata|Record View|Meat, Rabbit. Available online: http://data.un.org/Data.aspx?q=meat+datamart%5bFAO%5d&d=FAO&f=itemCode%3a1141 (accessed on 5 October 2023).
- Szendrő, K.; Szabó-Szentgróti, E.; Szigeti, O. Consumers’ Attitude to Consumption of Rabbit Meat in Eight Countries Depending on the Production Method and Its Purchase Form. Foods 2020, 9, 654. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Gerini, F.; Ikram, A.; Saeed, F.; Feng, X.; Chen, Y. Rabbit Meat—Production, Consumption and Consumers’ Attitudes and Behavior. Sustainability 2023, 15, 2008. [Google Scholar] [CrossRef]
- Chavatte-Palmer, P.; Laigre, P.; Simonoff, E.; Chesné, P.; Challah-Jacques, M.; Renard, J.P. In Utero Characterisation of Fetal Growth by Ultrasound Scanning in the Rabbit. Theriogenology 2008, 69, 859–869. [Google Scholar] [CrossRef]
- Sakowicz, A.; Bralewska, M.; Kamola, P.; Pietrucha, T. Reliability of Rodent and Rabbit Models in Preeclampsia Research. Int. J. Mol. Sci. 2022, 23, 14344. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Tello, J.; Arias-Alvarez, M.; Gonzalez-Bulnes, A.; Sferuzzi-Perri, A.N. Models of Intrauterine Growth Restriction and Fetal Programming in Rabbits. Mol. Reprod. Dev. 2019, 86, 1781–1809. [Google Scholar] [CrossRef] [PubMed]
- Akkuş, T.; Erdoğan, G. Ultrasonographic Evaluation of Feto-Placental Tissues at Different Intrauterine Locations in Rabbit. Theriogenology 2019, 138, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Coombs, P.; Walton, S.L.; Maduwegedera, D.; Flower, R.L.; Denton, K.M. Fetal Growth and Well-being in a Study of Maternal Hypertension in Rabbits. Anat. Rec. 2020, 303, 2646–2656. [Google Scholar] [CrossRef]
- Evci, E.C.; Aslan, S.; Schäfer-Somi, S.; Ergene, O.; Sayıner, S.; Darbaz, I.; Seyrek-İntaş, K.; Wehrend, A. Monitoring of Canine Pregnancy by Considering Anti-Mullerian Hormone, C-Reactive Protein, Progesterone and Complete Blood Count in Pregnant and Non-Pregnant Dogs. Theriogenology 2023, 195, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Mantziaras, G.; Luvoni, G.C. Advanced Ultrasound Techniques in Small Animal Reproduction Imaging. Reprod. Domest. Anim. 2020, 55, 17–25. [Google Scholar] [CrossRef]
- Von Borell, E.; Langbein, J.; Després, G.; Hansen, S.; Leterrier, C.; Marchant, J.; Marchant-Forde, R.; Minero, M.; Mohr, E.; Prunier, A. Heart Rate Variability as a Measure of Autonomic Regulation of Cardiac Activity for Assessing Stress and Welfare in Farm Animals—A Review. Physiol. Behav. 2007, 92, 293–316. [Google Scholar] [CrossRef] [PubMed]
- Nowland, M.H.; Brammer, D.W.; Garcia, A.; Rush, H.G. Biology and Diseases of Rabbits. In Laboratory Animal Medicine; Elsevier: Amsterdam, The Netherlands, 2015; pp. 411–461. [Google Scholar]
- Harcourt-Brown, F. Infectious Diseases of Domestic Rabbits. In Textbook of Rabbit Medicine; Elsevier: Amsterdam, The Netherlands, 2002; p. 361. [Google Scholar]
- Marai, I.F.M.; Habeeb, A.A.M.; Gad, A.E. Rabbits’ Productive, Reproductive and Physiological Performance Traits as Affected by Heat Stress: A Review. Livest. Prod. Sci. 2002, 78, 71–90. [Google Scholar] [CrossRef]
- Dorning, J.; Stephen, H. The Welfare of Farmed Rabbits in Commercial Production Systems. Tech. Rep. 2017, 279–292. [Google Scholar] [CrossRef]
- Quesenberry, K.; Carpenter, J. Ferrets, Rabbits, and Rodents: Clinical Medicine and Surgery, 3rd ed.; Quesenberry, K., Carpenter, J., Eds.; Elsevier Inc.: St. Louis, MO, USA, 2012; ISBN 9781416066217. [Google Scholar]
- Menchetti, L.; Canali, C.; Castellini, C.; Boiti, C.; Brecchia, G. The Different Effects of Linseed and Fish Oil Supplemented Diets on Insulin Sensitivity of Rabbit Does during Pregnancy. Res. Vet. Sci. 2018, 118, 126–133. [Google Scholar] [CrossRef]
- Tena-Betancourt, E.; Tena-Betancourt, C.A.; Zúniga-Muñoz, A.M.; Hernández-Godínez, B.; Ibáñez-Contreras, A.; Graullera-Rivera, V. Multiple Extrauterine Pregnancy with Early and near Full-Term Mummified Fetuses in a New Zealand White Rabbit (Oryctolagus cuniculus). J. Am. Assoc. Lab. Anim. Sci. 2014, 53, 204–207. [Google Scholar]
- Tena, E.; Hernández, B.; Ibáñez, A.; Reyes, S.; Zúñiga, A.; Graullera, V. Posible Superfetación En Una Coneja Raza NZB (Oryctolagus cuniculus) Concomitante a Retraso Del Desarrollo Intrauterino (RDIU). Spei Domus 2013, 9, 49–56. [Google Scholar] [CrossRef]
- Cappon, G.D.; Fleeman, T.L.; Chapin, R.E.; Hurtt, M.E. Effects of Feed Restriction during Organogenesis on Embryo-Fetal Development in Rabbit. Birth Defects Res. B Dev. Reprod. Toxicol. 2005, 74, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, R. The Systolic Blood Pressure of the Normal Rabbit Measured by a Slightly Modified van Leersum Method. J. Exp. Med. 1927, 46, 443–461. [Google Scholar] [CrossRef]
- McArdle, A.M.; Maduwegedera, D.; Moritz, K.; Flower, R.L.; Denton, K.M.; Roberts, C.T. Chronic Maternal Hypertension Affects Placental Gene Expression and Differentiation in Rabbits. J. Hypertens. 2010, 28, 959–968. [Google Scholar] [CrossRef] [PubMed]
- McArdle, A.M.; Roberts, C.T.; Maduwegedera, D.; Flower, R.L.; Denton, K.M. Chronic Maternal Hypertension Characterized by Renal Dysfunction Is Associated with Reduced Placental Blood Flow during Late Gestation in Rabbits. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R1043–R1049. [Google Scholar] [CrossRef]
- Denton, K.M.; Flower, R.L.; Stevenson, K.M.; Anderson, W.P. Adult Rabbit Offspring of Mothers with Secondary Hypertension Have Increased Blood Pressure. Hypertension 2003, 41, 634–639. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hall, J. MSD Manual|Eclampsia in Small Animals. Available online: https://www.msdvetmanual.com/metabolic-disorders/disorders-of-calcium-metabolism/eclampsia-in-small-animals#:~:text=Eclampsia%20(periparturient%20hypocalcemia)%20is%20characterized,2%E2%80%933%20weeks%20after%20whelping (accessed on 6 October 2023).
- Abitbol, M.M.; Gallo, G.R.; Pirani, C.L.; Ober, W.B. Production of Experimental Toxemia in the Pregnant Rabbit. Am. J. Obstet. Gynecol. 1976, 124, 460–470. [Google Scholar] [CrossRef]
- Harcourt-Brown, F.M.; Harcourt-Brown, S.F. Clinical Value of Blood Glucose Measurement in Pet Rabbits. Vet. Rec. 2012, 170, 674. [Google Scholar] [CrossRef]
- Hewitt, C.D.; Innes, D.J.; Savory, J.; Wills, M.R. Normal Biochemical and Hematological Values in New Zealand White Rabbits. Clin. Chem. 1989, 35, 1777–1779. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.; Masamune, T.; Ino, H.; Ueda, K.; Ishiyama, T.; Sessler, D.I.; Matsukawa, T. Severe Hypoglycemia Reduces the Shivering Threshold in Rabbits. BMC Anesthesiol. 2019, 19, 126. [Google Scholar] [CrossRef]
- Conaway, H.H.; Faas, F.H.; Smith, S.D.; Sanders, L.L. Spontaneous Diabetes Mellitus in the New Zealand White Rabbit: Physiologic Characteristics. Metabolism 1981, 30, 50–56. [Google Scholar] [CrossRef]
- Thieme, R.; Ramin, N.; Fischer, S.; Püschel, B.; Fischer, B.; Navarrete Santos, A. Gastrulation in Rabbit Blastocysts Depends on Insulin and Insulin-like-Growth-Factor 1. Mol. Cell. Endocrinol. 2012, 348, 112–119. [Google Scholar] [CrossRef]
- Rousseau-Ralliard, D.; Couturier-Tarrade, A.; Thieme, R.; Brat, R.; Rolland, A.; Boileau, P.; Aubrière, M.C.; Daniel, N.; Dahirel, M.; Derisoud, E.; et al. A Short Periconceptional Exposure to Maternal Type-1 Diabetes Is Sufficient to Disrupt the Feto-Placental Phenotype in a Rabbit Model. Mol. Cell. Endocrinol. 2019, 480, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.Q.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 Is an Omega-3 Fatty Acid Receptor Mediating Potent Anti-Inflammatory and Insulin-Sensitizing Effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. A Defect in the Activity of Δ6 and Δ5 Desaturases May Be a Factor Predisposing to the Development of Insulin Resistance Syndrome. Prostaglandins Leukot. Essent. Fat. Acids 2005, 72, 343–350. [Google Scholar] [CrossRef]
- Hughes, K. Abdominal Ectopic Pregnancy and Impaired Postnatal Mammary Gland Development, Consistent with Physiologic Agalactia, in a Wild European Rabbit, Oryctolagus cuniculus. Front. Vet. Sci. 2019, 6, 254. [Google Scholar] [CrossRef]
- Marco-Jiménez, F.; García-Domínguez, X.; Valdes-Hernández, J.; Vicente, J.S. Extra-Uterine (Abdominal) Full Term Foetus in a 15-Day Pregnant Rabbit. BMC Vet. Res. 2017, 13, 307. [Google Scholar] [CrossRef]
- Segura Gil, P.; Peris Palau, B.; Martínez Martínez, J.; Ortega Porcel, J.; Corpa Arenas, J.M. Abdominal Pregnancies in Farm Rabbits. Theriogenology 2004, 62, 642–651. [Google Scholar] [CrossRef]
- Jacobson, H.A.; Kibbe, D.P.; Kirkpatrick, R.L. Ectopic Fetuses in Two Cottontail Rabbits. J. Wildl. Dis. 1975, 11, 540–542. [Google Scholar] [CrossRef][Green Version]
- Stott, P.; Wight, N. Female Reproductive Tract Abnormalities in European Hares (Lepus europaeus) in Australia. J. Wildl. Dis. 2004, 40, 696–703. [Google Scholar] [CrossRef][Green Version]
- Tan, S.; Zhou, F.; Nielsen, V.G.; Wang, Z.; Gladson, C.L.; Parks, D.A. Increased Injury Following Intermittent Fetal Hypoxia-Reoxygenation Is Associated with Increased Free Radical Production in Fetal Rabbit Brain. J. Neuropathol. Exp. Neurol. 1999, 58, 972–981. [Google Scholar] [CrossRef]
- Millicovsky, G.; DeSesso, J.M. Differential Embryonic Cardiovascular Responses to Acute Maternal Uterine Ischemia: An in Vivo Microscopic Study of Rabbit Embryos with Either Intact or Clamped Umbilical Cords. Teratology 1980, 22, 335–343. [Google Scholar] [CrossRef]
- Tan, S.; Bose, R.; Derrick, M. Hypoxia-Ischemia in Fetal Rabbit Brain Increases Reactive Nitrogen Species Production: Quantitative Estimation of Nitrotyrosine. Free Radic. Biol. Med. 2001, 30, 1045–1051. [Google Scholar] [CrossRef]
- Tan, S.; Zhou, F.; Nielsen, V.G.; Wang, Z.; Gladson, C.L.; Parks, D.A. Sustained Hypoxia-Ischemia Results in Reactive Nitrogen and Oxygen Species Production and Injury in the Premature Fetal Rabbit Brain. J. Neuropathol. Exp. Neurol. 1998, 57, 544–553. [Google Scholar] [CrossRef]
- Tan, S.; Liu, Y.-Y.; Nielsen, V.G.; Skinner, K.; Kirk, K.A.; Baldwin, S.T.; Parks, D.A. Maternal Infusion of Antioxidants (Trolox and Ascorbic Acid) Protects the Fetal Heart in Rabbit Fetal Hypoxia. Pediatr. Res. 1996, 39, 499–503. [Google Scholar] [CrossRef]
- Derrick, M.; Luo, N.L.; Bregman, J.C.; Jilling, T.; Ji, X.; Fisher, K.; Gladson, C.L.; Beardsley, D.J.; Murdoch, G.; Back, S.A.; et al. Preterm Fetal Hypoxia-Ischemia Causes Hypertonia and Motor Deficits in the Neonatal Rabbit: A Model for Human Cerebral Palsy? J. Neurosci. 2004, 24, 24–34. [Google Scholar] [CrossRef]
- Sase, M.; Lee, J.J.; Ross, M.G.; Buchmiller-Crair, T.L. Effect of Hypoxia on Fetal Rabbit Gastrointestinal Motility. J. Surg. Res. 2001, 99, 347–351. [Google Scholar] [CrossRef]
- Gumus, H.G.; Illa, M.; Pla, L.; Zamora, M.; Crispi, F.; Gratacos, E. Nutritional Intra-Amniotic Therapy Increases Survival in a Rabbit Model of Fetal Growth Restriction. PLoS ONE 2018, 13, e0193240. [Google Scholar] [CrossRef]
- Figueroa, H.; Lozano, M.; Suazo, C.; Eixarch, E.; Illanes, S.E.; Carreño, J.E.; Villanueva, S.; Hernández-Andrade, E.; Gratacós, E.; Irarrazabal, C.E. Intrauterine Growth Restriction Modifies the Normal Gene Expression in Kidney from Rabbit Fetuses. Early Hum. Dev. 2012, 88, 899–904. [Google Scholar] [CrossRef]
- López-Tello, J.; Arias-Álvarez, M.; Jiménez-Martínez, M.Á.; Barbero-Fernández, A.; García-García, R.M.; Rodríguez, M.; Lorenzo, P.L.; Torres-Rovira, L.; Astiz, S.; González-Bulnes, A.; et al. The Effects of Sildenafil Citrate on Feto-Placental Development and Haemodynamics in a Rabbit Model of Intrauterine Growth Restriction. Reprod. Fertil. Dev. 2017, 29, 1239–1248. [Google Scholar] [CrossRef]
- Pla, L.; Illa, M.; Loreiro, C.; Lopez, M.C.; Vázquez-Aristizabal, P.; Kühne, B.A.; Barenys, M.; Eixarch, E.; Gratacós, E. Structural Brain Changes during the Neonatal Period in a Rabbit Model of Intrauterine Growth Restriction. Dev. Neurosci. 2021, 42, 217–229. [Google Scholar] [CrossRef]
- Figueroa, H.; Cifuentes, J.; Lozano, M.; Alvarado, C.; Cabezas, C.; Eixarch, E.; Fernández, E.; Contreras, L.; Illanes, S.E.; Hernández-Andrade, E.; et al. Nitric Oxide Synthase and Changes in Oxidative Stress Levels in Embryonic Kidney Observed in a Rabbit Model of Intrauterine Growth Restriction. Prenat. Diagn. 2016, 36, 628–635. [Google Scholar] [CrossRef]
- Van Vliet, E.; Eixarch, E.; Illa, M.; Arbat-Plana, A.; González-Tendero, A.; Hogberg, H.T.; Zhao, L.; Hartung, T.; Gratacos, E. Metabolomics Reveals Metabolic Alterations by Intrauterine Growth Restriction in the Fetal Rabbit Brain. PLoS ONE 2013, 8, e64545. [Google Scholar] [CrossRef]
- Simões, R.V.; Muñoz-Moreno, E.; Carbajo, R.J.; González-Tendero, A.; Illa, M.; Sanz-Cortés, M.; Pineda-Lucena, A.; Gratacós, E. In Vivo Detection of Perinatal Brain Metabolite Changes in a Rabbit Model of Intrauterine Growth Restriction (IUGR). PLoS ONE 2015, 10, e0131310. [Google Scholar] [CrossRef]
- Illa, M.; Eixarch, E.; Muñoz-Moreno, E.; Batalle, D.; Leal-Campanario, R.; Gruart, A.; Delgado-García, J.M.; Figueras, F.; Gratacós, E. Neurodevelopmental Effects of Undernutrition and Placental Underperfusion in Fetal Growth Restriction Rabbit Models. Fetal Diagn. Ther. 2017, 42, 189–197. [Google Scholar] [CrossRef]
- Eixarch, E.; Batalle, D.; Illa, M.; Muñoz-Moreno, E.; Arbat-Plana, A.; Amat-Roldan, I.; Figueras, F.; Gratacos, E. Neonatal Neurobehavior and Diffusion MRI Changes in Brain Reorganization Due to Intrauterine Growth Restriction in a Rabbit Model. PLoS ONE 2012, 7, e31497. [Google Scholar] [CrossRef] [PubMed]
- Schipke, J.; Gonzalez-Tendero, A.; Cornejo, L.; Willführ, A.; Bijnens, B.; Crispi, F.; Mühlfeld, C.; Gratacós, E. Experimentally Induced Intrauterine Growth Restriction in Rabbits Leads to Differential Remodelling of Left versus Right Ventricular Myocardial Microstructure. Histochem. Cell Biol. 2017, 148, 557–567. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, M.A.; Saleh, S.A.-A.; Maher, M.A.; Khidre, A.M. Utero-Placental Perfusion Doppler Indices in Growth Restricted Fetuses: Effect of Sildenafil Citrate. J. Matern.-Fetal Neonatal Med. 2018, 31, 1045–1050. [Google Scholar] [CrossRef] [PubMed]
- López-Tello, J.; Barbero, A.; González-Bulnes, A.; Astiz, S.; Rodríguez, M.; Formoso-Rafferty, N.; Arias-Álvarez, M.; Rebollar, P.G. Characterization of Early Changes in Fetoplacental Hemodynamics in a Diet-Induced Rabbit Model of IUGR. J. Dev. Orig. Health Dis. 2015, 6, 454–461. [Google Scholar] [CrossRef]
- Roellig, K.; Menzies, B.R.; Hildebrandt, T.B.; Goeritz, F. The Concept of Superfetation: A Critical Review on a ‘Myth’ in Mammalian Reproduction. Biol. Rev. 2011, 86, 77–95. [Google Scholar] [CrossRef]
- Pickard, J.N. Cases of Prolonged Gestation, Superfœtation and Abnormal Pseudo-Pregnancy in the Rabbit. Vet. J. (1900) 1928, 84, 471–473. [Google Scholar] [CrossRef]
- Stieve, H. von Zur Fortpflanzungsbiologie Des Europäischen Feldhasen (Lepus europaeus Pallas). Zool. Anz. 1952, 148, 101–114. [Google Scholar]
- Maurer, R.R.; Foote, R.H. Maternal Ageing and Embryonic Mortality in the Rabbit. I. Repeated Superovulation, Embryo Culture and Transfer. J. Reprod. Fertil. 1971, 25, 329–341. [Google Scholar] [CrossRef]
- Rosell, J.M.; de la Fuente, L.F.; Carbajo, M.T.; Fernández, X.M. Reproductive Diseases in Farmed Rabbit Does. Animals 2020, 10, 1873. [Google Scholar] [CrossRef]
- Peppa, M.; Uribarri, J.; Vlassara, H. Aging and Glycoxidant Stress. Hormones 2008, 7, 123–132. [Google Scholar] [CrossRef] [PubMed]
- De Nivelle, J.; Thoma, J.; Toto Nienguesso, A.; Seeling, T.; Jung, J.-S.; Navarrete Santos, A.; Schindler, M. Rabbit as an Aging Model in Reproduction: Advanced Maternal Age Alters Glo1 Expression the Endometrium at the Time of Implantation. Appl. Sci. 2020, 10, 7732. [Google Scholar] [CrossRef]
- Toufexis, D.; Rivarola, M.A.; Lara, H.; Viau, V. Stress and the Reproductive Axis. J. Neuroendocrinol. 2014, 26, 573–586. [Google Scholar] [CrossRef]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the Hypothalamic-Pituitary-Adrenocortical Stress Response. Compr. Physiol. 2016, 6, 603. [Google Scholar]
- Fitzgerald, E.; Hor, K.; Drake, A.J. Maternal Influences on Fetal Brain Development: The Role of Nutrition, Infection and Stress, and the Potential for Intergenerational Consequences. Early Hum. Dev. 2020, 150, 105190. [Google Scholar] [CrossRef]
- Coussons-Read, M.E. Effects of Prenatal Stress on Pregnancy and Human Development: Mechanisms and Pathways. Obstet. Med. 2013, 6, 52–57. [Google Scholar] [CrossRef]
- Bal’magiya, T.A.; Surovtseva, Z.F. Growth Pattern of Rabbit Fetuses during Normal Pregnancy and with Inhibition of the “Gestation Dominant”. Bull. Exp. Biol. Med. 1974, 77, 384–387. [Google Scholar] [CrossRef]
- Brouček, J. Effect of Noise on Performance, Stres, and Behaviour of Animals. Slovak J. Anim. Sci. 2014, 47, 111–123. [Google Scholar]
- Peveler, J.L.; Hickman, D.L. Effects of Music Enrichment on Individually Housed Male New Zealand White Rabbits. J. Am. Assoc. Lab. Anim. Sci. 2018, 57, 695–697. [Google Scholar] [CrossRef] [PubMed]
- Benedek, I.; Altbäcker, V.; Molnár, T. Stress Reactivity near Birth Affects Nest Building Timing and Offspring Number and Survival in the European Rabbit (Oryctolagus cuniculus). PLoS ONE 2021, 16, e0246258. [Google Scholar] [CrossRef] [PubMed]
- NC3Rs The 3Rs. Available online: https://www.nc3rs.org.uk/the-3rs (accessed on 25 August 2020).
- Von Holst, D.; Hutzelmeyer, H.; Kaetzke, P.; Khaschei, M.; Schönheiter, R. Social Rank, Stress, Fitness, and Life Expectancy in Wild Rabbits. Naturwissenschaften 1999, 86, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Stephens, D.B.; Adams, C.E. Observations on the Effects of Vibration Stress and Sound on Pregnancy, Parturition and Respiration in the Rabbit. Lab. Anim. 1982, 16, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Tomruk, A.; Ozgur-Buyukatalay, E.; Ozturk, G.G.; Ulusu, N.N. Short-Term Exposure to Radiofrequency Radiation and Metabolic Enzymes’ Activities during Pregnancy and Prenatal Development. Electromagn. Biol. Med. 2022, 41, 370–378. [Google Scholar] [CrossRef]
- Gregg, X.T.; Prchal, J.T. Red Blood Cell Enzymopathies. In Hematology; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Suc, N.Q.; Van Binh, D.; Le Thi Thu Ha, P.T.R. Effect of Housing System (Cage versus Underground Shelter) on Performance of Rabbits on Farms. Livest. Res. Rural Dev. 1996, 8, 34. [Google Scholar]
- Verga, M. Some Characteristics of Rabbit Behavior and Their Relationship with Husbandry Systems. J. Appl. Rabbit. Res. 1993, 15, 55. [Google Scholar]
- Fayez, I.; Marai, M.; Rashwan, A.A. Rabbits Behavioural Response to Climatic and Managerial Conditions—A Review. Arch. Tierz. Dummerstorf 2004, 47, 469–482. [Google Scholar]
- Bagliacca, M.; Camillo, F.; Paci, G. Temperatura e Performances Di Conigli Maschi Riproduttori. Riv. Coniglicoltura 1987, 10, 61–65. [Google Scholar]
- Nielsen, S.; Alvarez, J.; Bicout, D.J.; Calistri, P.; Depner, K.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; Gortázar Schmidt, C.; Michel, V.; et al. Health and Welfare of Rabbits Farmed in Different Production Systems. EFSA J. 2020, 18, e05944. [Google Scholar] [CrossRef]
- Quintela, L.; Peña, A.; Barrio, M.; Vega, M.; Diaz, R.; Maseda, F.; Garcia, P. Reproductive Performance of Multiparous Rabbit Lactating Does: Effect of Lighting Programs and PMSG Use. Reprod. Nutr. Dev. 2001, 41, 247–257. [Google Scholar] [CrossRef][Green Version]
- Symeon, G.K.; Goliomytis, M.; Bizelis, I.; Papadomichelakis, G.; Pagonopoulou, O.; Abas, Z.; Deligeorgis, S.G.; Chadio, S.E. Effects of Gestational Maternal Undernutrition on Growth, Carcass Composition and Meat Quality of Rabbit Offspring. PLoS ONE 2015, 10, e0118259. [Google Scholar] [CrossRef] [PubMed]
- Simitzis, P.E.; Skoupa, E.P.; Goliomytis, M.; Kominakis, A.; Deligeorgis, S.G. The Effects of Mild Maternal Undernutrition on the Post-Weaning Behaviour of Rabbits. Appl. Anim. Behav. Sci. 2013, 148, 157–163. [Google Scholar] [CrossRef]
- García-García, R.M.; Arias-álvarez, M.; Millán, P.; Rodríguez, M.; Sánchez-Rodríguez, A.; Lorenzo, P.L.; Rebollar, P.G. Gestation Food Restriction and Refeeding Compensate Maternal Energy Status and Alleviate Metabolic Consequences in Juvenile Offspring in a Rabbit Model. Nutrients 2021, 13, 310. [Google Scholar] [CrossRef] [PubMed]
- García-García, R.M.; Arias-Álvarez, M.; Rodríguez, M.; Sánchez-Rodríguez, A.; Formoso-Rafferty, N.; Lorenzo, P.L.; Rebollar, P.G. Effects of Feed Restriction during Pregnancy on Maternal Reproductive Outcome, Foetal Hepatic IGF Gene Expression and Offspring Performance in the Rabbit. Animal 2021, 15, 100382. [Google Scholar] [CrossRef] [PubMed]
- Goliomytis, M.; Skoupa, E.-P.; Konga, A.; Symeon, G.K.; Charismiadou, M.A.; Deligeorgis, S.G. Influence of Gestational Maternal Feed Restriction on Growth Performance and Meat Quality of Rabbit Offsprings. Animal 2016, 10, 157–162. [Google Scholar] [CrossRef]
- Simitzis, P.; Kiriakopoulos, A.; Symeon, G.; Goliomytis, M.; Kominakis, A.; Bizelis, I.; Chadio, S.; Pagonopoulou, O.; Deligeorgis, S. Effect of Prenatal Overfeeding on the Growth Performance and Postweaning Behavioral Parameters in Rabbits. J. Vet. Behav. 2019, 33, 38–45. [Google Scholar] [CrossRef]
- Lopez-Tello, J.; Arias-Alvarez, M.; Jimenez-Martinez, M.A.; Garcia-Garcia, R.M.; Rodriguez, M.; Lorenzo Gonzalez, P.L.; Bermejo-Poza, R.; Gonzalez-Bulnes, A.; Garcia Rebollar, P. Competition for Materno-Fetal Resource Partitioning in a Rabbit Model of Undernourished Pregnancy. PLoS ONE 2017, 12, e0169194. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, E.; Díaz, G.; Salazar, A.M.; Montúfar-Chaveznava, R.; Caldelas, I. Long-Term Changes in the Diurnal Temporal Regulation and Set Points of Metabolic Parameters Associated with Chronic Maternal Overnutrition in Rabbits. Am. J. Physiol. Endocrinol. Metab. 2022, 323, E503–E516. [Google Scholar] [CrossRef] [PubMed]
- Kofman, O. The Role of Prenatal Stress in the Etiology of Developmental Behavioural Disorders. Neurosci. Biobehav. Rev. 2002, 26, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Dupont, C.; Ralliard-Rousseau, D.; Tarrade, A.; Faure, C.; Dahirel, M.; Sion, B.; Brugnon, F.; Levy, R.; Chavatte-Palmer, P. Impact of Maternal Hyperlipidic Hypercholesterolaemic Diet on Male Reproductive Organs and Testosterone Concentration in Rabbits. J. Dev. Orig. Health Dis. 2014, 5, 183–188. [Google Scholar] [CrossRef]
- Szendro, Z.; Szendro, K.; Zotte, A.D. Management of Reproduction on Small, Medium and Large Rabbit Farms: A Review. Asian-Australas. J. Anim. Sci. 2012, 25, 738–748. [Google Scholar] [CrossRef]
- Theau-clément, M.; Guardia, S.; Davoust, C.; Galliot, P.; Souchet, C.; Bignon, L.; Fortun-lamothe, L. Performance and Sustainability of Two Alternative Rabbit Breeding Systems. World Rabbit Sci. 2016, 24, 253–265. [Google Scholar] [CrossRef]
- Hoffman, L.H.; Olson, G.E.; Carson, D.D.; Chilton, B.S. Progesterone and Implanting Blastocysts Regulate Muc1 Expression in Rabbit Uterine Epithelium*. Endocrinology 1998, 139, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Mocé, M.L.; Santacreu, M.A.; Climent, A.; Blasco, A. The Effect of Divergent Selection for Uterine Capacity on Prenatal Survival in Rabbits: Maternal and Embryonic Genetic Effects1. J. Anim. Sci. 2004, 82, 68–73. [Google Scholar] [CrossRef][Green Version]
- Vicente, J.; Llobat, M.; Jiménez-Trigos, E.; Lavara, R.; Marco-Jiménez, F. Effect of Embryonic and Maternal Genotype on Embryo and Foetal Survival in Rabbit. Reprod. Domest. Anim. 2013, 48, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Naturil-Alfonso, C.; Marco-Jiménez, F.; Jiménez-Trigos, E.; Saenz-de-Juano, M.; Viudes-de-Castro, M.; Lavara, R.; Vicente, J. Role of Embryonic and Maternal Genotype on Prenatal Survival and Foetal Growth in Rabbit. Reprod. Domest. Anim. 2015, 50, 312–320. [Google Scholar] [CrossRef]
- Turna, B.; Erdoğan, G. Assessment of Fetal Growth by B-Mode and Doppler Ultrasonography in Rabbit during Pregnancy. Sağlık Bilim. Vet. Derg. Fırat Üniversitesi 2016, 30, 89–93. [Google Scholar]
- El-Gayar, M.; Khalil, H.; Hanafy, A.; Yaseen, M.; Hegaze, E.; Marthold, D.; Gauly, M.; Holtz, W. Pregnancy Detection in Rabbits by Ultrasonography as Compared to Manual Palpation. Egypt J. Anim. Prod. 2014, 51, 196–199. [Google Scholar]
- Yuan, H.; Liu, C.; Wang, H.; Wang, L.; Dai, L. Early Pregnancy Diagnosis of Rabbits: A Non-Invasive Approach Using Vis-NIR Spatially Resolved Spectroscopy. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2021, 264, 120251. [Google Scholar] [CrossRef] [PubMed]
- Limmanont, C.; Ponglowhapan, S.; Tanhan, P.; Sirinarumitr, T.; Sirinarumitr, K. Fetal Head Diameter in Dogs and Cats Measured by Radiography and Ultrasonography. Thai J. Vet. Med. 2019, 49, 175–182. [Google Scholar]
- Biswal, S.S.; Satapathy, S.; Panda, S.K.; Sahoo, A.K.; Kanungo, S.; Joshi, S.K.; Sahoo, B.; Reddy, S.; Sethy, R.K.; Narayan, U.V.S. Successful Management of a Secondary Abdominal Pregnancy with Partial Fetal Mummification in a Doe. Communications 2022, 46, 711–717. [Google Scholar]
- Prestes, N.C.; Chalhoub, M.C.L.; Lopes, M.D.; Takahira, R.K. Amniocentesis and Biochemical Evaluation of Amniotic Fluid in Ewes at 70, 100 and 145 Days of Pregnancy. Small Rumin. Res. 2001, 39, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Crociati, M.; Sylla, L.; De Vincenzi, A.; Stradaioli, G.; Monaci, M. How to Predict Parturition in Cattle? A Literature Review of Automatic Devices and Technologies for Remote Monitoring and Calving Prediction. Animals 2022, 12, 405. [Google Scholar] [CrossRef] [PubMed]
- Shung, K.K. High Frequency Ultrasonic Imaging. J. Med. Ultrasound 2009, 17, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Winter, N.; Dohrn, M.F.; Wittlinger, J.; Loizides, A.; Gruber, H.; Grimm, A. Role of High-Resolution Ultrasound in Detection and Monitoring of Peripheral Nerve Tumor Burden in Neurofibromatosis in Children. Childs Nerv. Syst. 2020, 36, 2427–2432. [Google Scholar] [CrossRef]
- Dekoninck, P.; Endo, M.; Sandaite, I.; Richter, J.; De Catte, L.; Van Calster, B.; Toelen, J.; Himmelreich, U.; Claus, F.; Deprest, J. A Pictorial Essay on Fetal Rabbit Anatomy Using Micro-Ultrasound and Magnetic Resonance Imaging. Prenat. Diagn. 2014, 34, 84–89. [Google Scholar] [CrossRef]
- Jakubovic, R.; Ramjist, J.; Gupta, S.; Guha, D.; Sahgal, A.; Foster, F.S.; Yang, V.X.D. High-Frequency Micro-Ultrasound Imaging and Optical Topographic Imaging for Spinal Surgery: Initial Experiences. Ultrasound Med. Biol. 2018, 44, 2379–2387. [Google Scholar] [CrossRef]
- Mazandarani, M.; Barati, F.; Farahmand, K.; Hoseini, F.; Eghbalsaied, S. Ultrasound Monitoring of Pregnancy in Mini-Lop Rabbits. Theriogenology 2021, 174, 47–52. [Google Scholar] [CrossRef]
- Idris, Y.; Audu, A.; Lawal, M.; Ibinaiye, O.; Fadason, F.; Muazu, N.; Echekwu, W. Sonographic Diagnosis of Pregnancy and Study of Gestational Changes in Rabbit Does. Nig. Vet. J. 2016, 37, 133–139. [Google Scholar]
- Animal Ultrasound Association, A. Safe Pregnancy Scanning in Very Small Animals: Rabbits and Guinea Pigs—Animal Ultrasound Association. Available online: https://www.animalultrasoundassociation.org/ultrasound-rabbits-guinea-pigs/ (accessed on 31 August 2021).
- Ypsilantis, P.; Saratsis, P.H. Early Pregnancy Diagnosis in the Rabbit by Real Time Ultrasonography. World Rabbit Sci. 1999, 7, 95–99. [Google Scholar] [CrossRef]
- LBNMedical Ultrasound Transducer Types and How to Select the Right Transducer. Available online: https://lbnmedical.com/ultrasound-transducer-types/ (accessed on 6 September 2021).
- Amran, U.N.; Zaiki, F.W.A.; Dom, S.M. A Review of the Thermal Effects During Pregnancy by Using Ultrasound: Doppler Mode. Pertanika J. Sci. Technol. 2019, 27, 357–370. [Google Scholar]
- Dom, S.M.; Razak, H.R.A.; Zaiki, F.W.A.; Saat, N.H.; Abd Manan, K.; Isa, I.N.C.; Hashim, U.F. Ultrasound Exposure during Pregnancy Affects Rabbit Foetal Parathyroid Hormone (PTH) Level. Quant. Imaging Med. Surg. 2013, 3, 49. [Google Scholar]


{kind=link}
{kind=link}
| Imaging Tool | Technical Information | Animal Preparation | Sedation | Parameters Evaluated | Authors | |
|---|---|---|---|---|---|---|
| High- Resolution ultrasound | <1 mm, using a high-resolution 12–5 MHz linear transducer | Anterior abdominal region shaved; supine position | No | Biometric data (fetal number and location; crown-rump length; femur length; renal length and circumference; biparietal diameter; abdominal circumference) and morphology (head, neck, face, thorax, lung, diaphragm, cardiac, abdomen, spine, extremities, genitalia, and fetal environment) | [10] | |
| Doppler | Spectral | N/A | Anterior abdominal region shaved; supine position | No | Umbilical arteries: resistive index | [10] |
| Ventral area shaved; left paralumbar fossa region examined | Uterine and umbilical arteries: pulsatility index and resistance index | [9] | ||||
| With B-mode (8.0 MHz; Gain: 97%; Deepness: 6 cm): Biparietal diameter; trunk diameter; and echotexture of placentas | ||||||
| Vivid q ultrasound (1.4–2.5 MHz phased array probe); 70 Hz high pass filter; Angle of insonation: <30º | Midline laparotomy and exteriorization of both uterine horns | Yes | Ductus venosus pulsatility index; aortic isthmus flow; left and right ventricular sphericity indices calculated as base-to-apex length/basal ventricular diameter; left ejection fraction cardiac output normalized by body weight | [58] | ||
| With M-mode: longitudinal axis motion and annular peak velocities at the septal mitral and lateral tricuspid annulus (apical 4-chamber view); septal wall thickness (transverse 4-chamber view) | ||||||
| Color | Linear 7.5 MHz probe | Abdomen clipped; placed on their backs and prevented from moving in a custom-made case | No | Vesicle, placental, fetal length, and head size; umbilicus, kidneys, aortic diameter, limbs, among others made occasionally | [6] | |
| Pulsed | N/A | N/A | N/A | Blood flow in the ductus venosus (pulsatility index), umbilical vein, and umbilical artery | [112] | |
| Micro-ultrasound | Linear 20 or 30 MHz probe; 30 µm resolution | Exteriorization of the uterine horn with the fetus of interest after a midline laparotomy; performed directly on the uterus constantly irrigated with warmed saline | Yes | Abdominal, cardiac, and thoracic circumferences; humerus and femur length; umbilical vein and artery diameter; deepest vertical pocket of amniotic fluid; and placental thickness | [112] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinto-Pinho, P.; Pinto, M.d.L.; Monteiro, J.; Fardilha, M.; Pinto-Leite, R.; Colaço, B. Pregnancy Complications and Feto-Maternal Monitoring in Rabbits. Vet. Sci. 2023, 10, 622. https://doi.org/10.3390/vetsci10100622
Pinto-Pinho P, Pinto MdL, Monteiro J, Fardilha M, Pinto-Leite R, Colaço B. Pregnancy Complications and Feto-Maternal Monitoring in Rabbits. Veterinary Sciences. 2023; 10(10):622. https://doi.org/10.3390/vetsci10100622
Chicago/Turabian StylePinto-Pinho, Patrícia, Maria de Lurdes Pinto, José Monteiro, Margarida Fardilha, Rosário Pinto-Leite, and Bruno Colaço. 2023. "Pregnancy Complications and Feto-Maternal Monitoring in Rabbits" Veterinary Sciences 10, no. 10: 622. https://doi.org/10.3390/vetsci10100622
APA StylePinto-Pinho, P., Pinto, M. d. L., Monteiro, J., Fardilha, M., Pinto-Leite, R., & Colaço, B. (2023). Pregnancy Complications and Feto-Maternal Monitoring in Rabbits. Veterinary Sciences, 10(10), 622. https://doi.org/10.3390/vetsci10100622