Responses of Germination to Light and to Far-Red Radiation—Can they be Predicted from Diaspores Size?


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
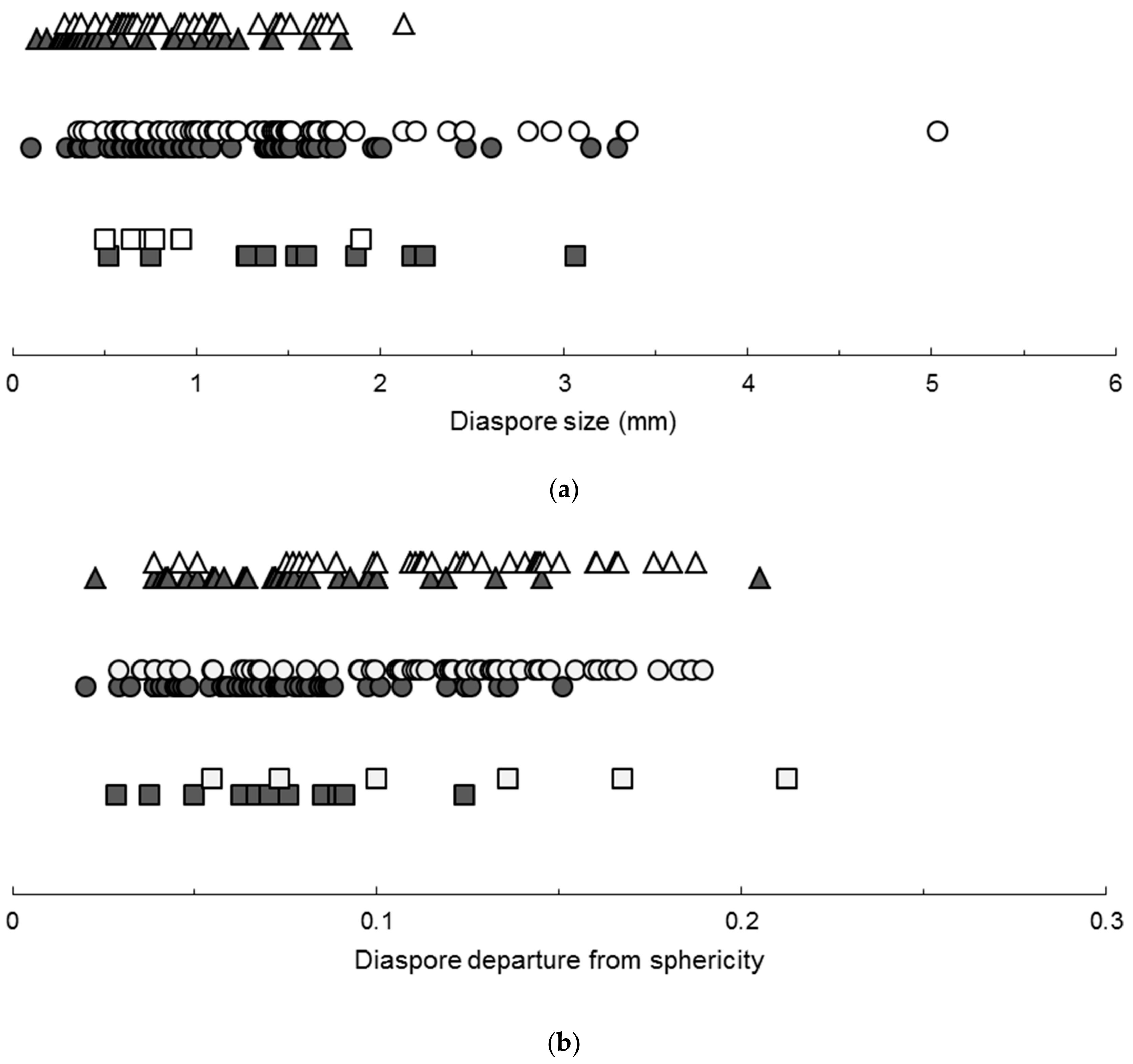
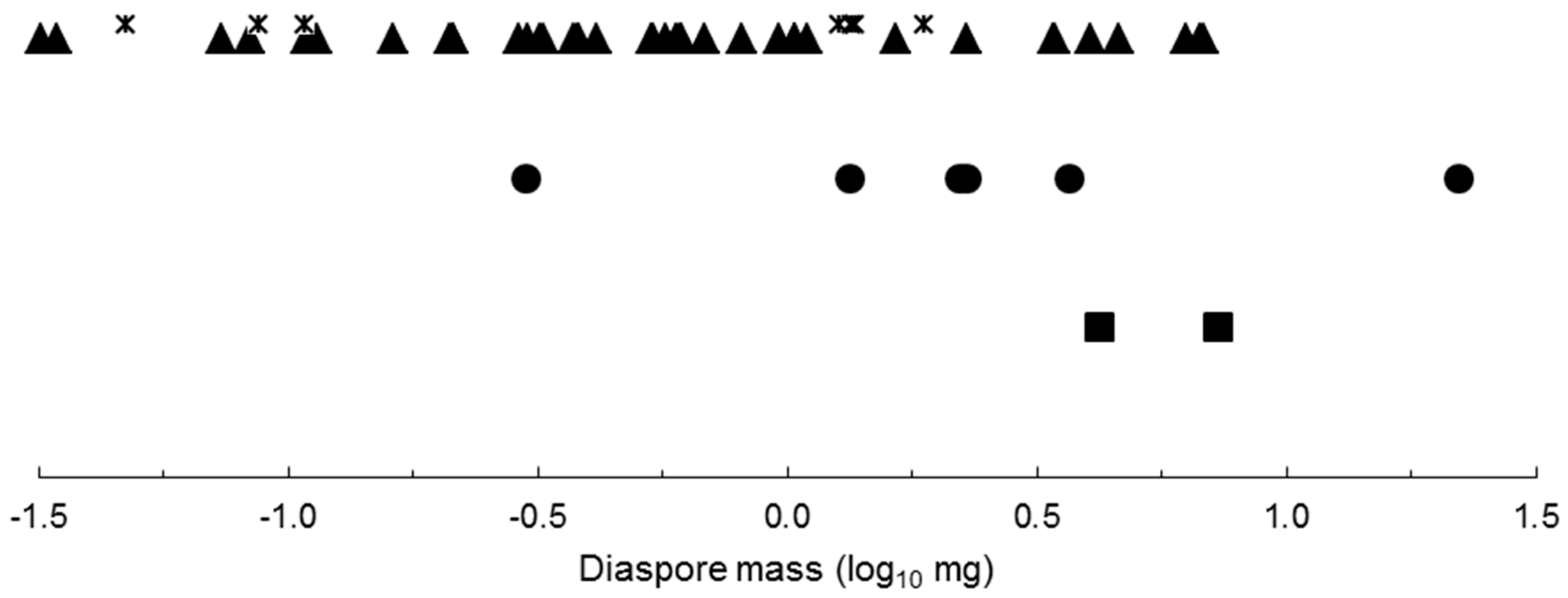
2.1. Photoblastic Responses
| 1. Diaspore thickness ≤ 0.292 mm | Positive photoblasty |
| Diaspore thickness > 0.292 mm | Indifferent photoblasty |
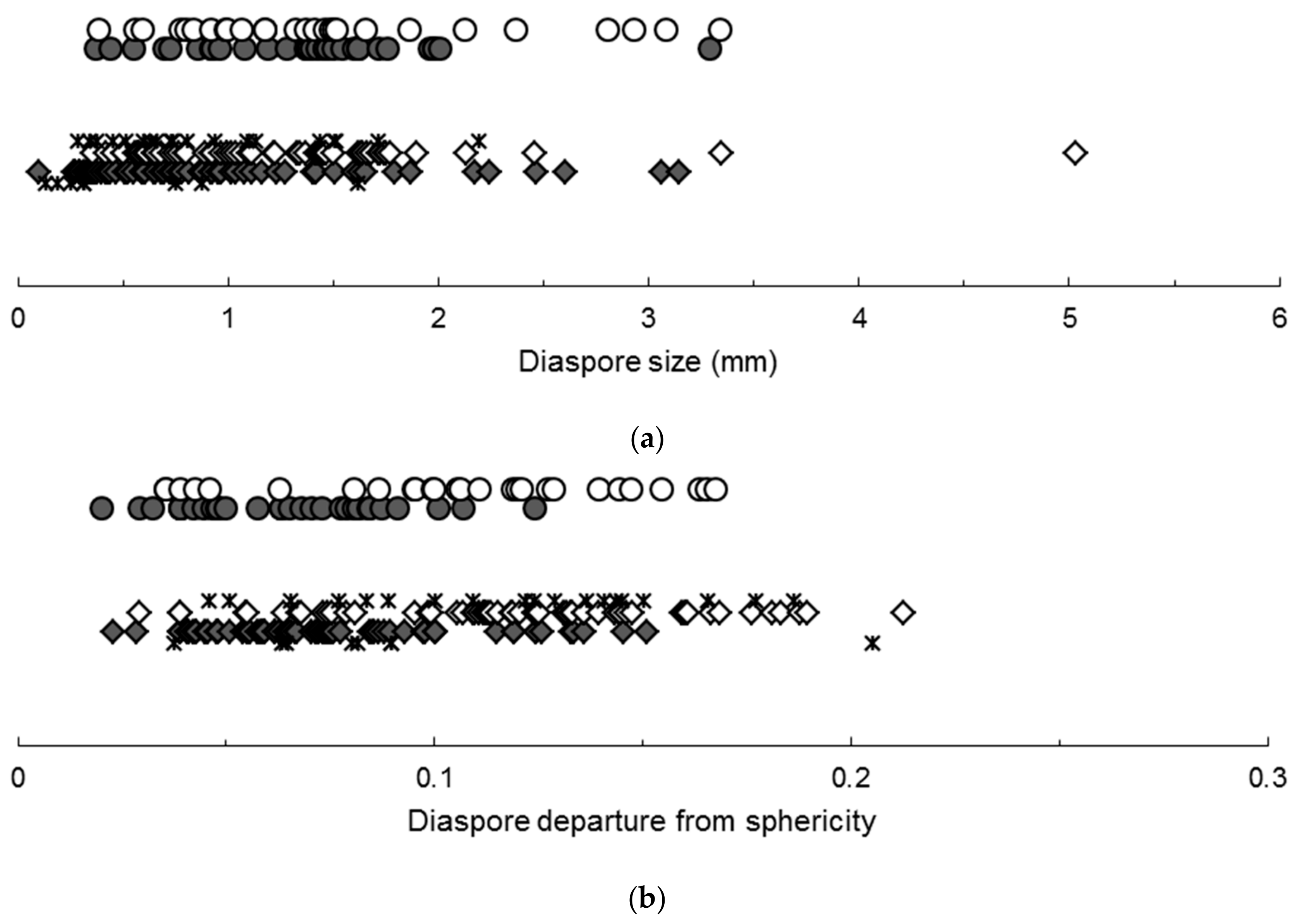
2.2. Effects of Far-Red Radiation
3. Discussion
3.1. Photoblastic Responses
3.2. Effects of Far-Red Radiation
4. Materials and Methods
4.1. Sources of Data
4.1.1. Seed Volume Dataset
4.1.2. Germination Dataset
4.1.3. Combining Seed Volume and Germination Datasets
4.2. Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B

References
- Ganhão, E.; Dias, L.S. Seed volume dataset – an ongoing inventory of seed size expressed by volume. Data 2019, 4, 61. [Google Scholar] [CrossRef]
- Milberg, P.; Andersson, L.; Thompson, K. Large-seeded species are less dependent on light for germination than small-seeded ones. Seed Sci. Res. 2000, 10, 99–104. [Google Scholar] [CrossRef]
- Górski, T. Germination of seeds in the shadow of plants. Physiol. Plant. 1975, 34, 342–346. [Google Scholar] [CrossRef]
- Górski, T.; Górska, K.; Nowicki, J. Germination of seeds of various herbaceous species under leaf canopy. Flora 1977, 166, 249–259. [Google Scholar] [CrossRef]
- Górski, T.; Górska, K.; Rybicki, J. Studies on the germination of seeds under leaf canopy. Flora 1978, 167, 289–299. [Google Scholar] [CrossRef]
- Pons, T.L. Seed responses to light. In Seeds. The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI Publishing: Wallingford, UK, 2000; pp. 237–260. [Google Scholar]
- Kitajima, K.; Fenner, M. Ecology of seedling regeneration. In Seeds. The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI Publishing: Wallingford, UK, 2000; pp. 331–359. [Google Scholar]
- Dias, L.S.; Dias, A.S. The relationship between shape and size of diaspores depends on being seeds or fruits. Horticulturae 2019, 5, 65. [Google Scholar] [CrossRef]
- Harper, J.L.; Lovell, P.H.; Moore, K.G. The shapes and sizes of seeds. Annu. Rev.Ecol. Syst. 1970, 1, 327–356. [Google Scholar] [CrossRef]
- Thompson, K.; Band, S.R.; Hodgson, J.G. Seed size and shape predict persistence in soil. Funct. Ecol. 1993, 7, 236–241. [Google Scholar] [CrossRef]
- Leishman, M.R.; Westoby, M. Seed size and shape are not related to persistence in soil in Australia in the same way as in Britain. Funct. Ecol. 1998, 12, 480–485. [Google Scholar] [CrossRef]
- Funes, G.; Basconcelo, S.; Díaz, S.; Cabido, M. Seed size and shape are good predictors of seed persistence in soil in temperate mountain grasslands of Argentina. Seed Sci. Res. 1999, 9, 341–345. [Google Scholar] [CrossRef]
- Moles, A.T.; Hodson, D.W.; Webb, C.J. Seed size and shape and persistence in the soil in the New Zealand flora. Oikos 2000, 89, 541–545. [Google Scholar] [CrossRef]
- Thompson, K.; Jalili, A.; Hodgson, J.G.; Hamzeh’ee, B.; Shaw, S.; Shirvany, A.; Yazdani, S.; Khoshnevis, M.; Zarrinkamar, F.; Ghahramani, M.-A.; et al. Seed size, shape and persistence in the soil in an Iranian flora. Seed Sci. Res. 2001, 11, 345–355. [Google Scholar] [CrossRef]
- Cerabolini, B.; Cerlani, R.M.; Caccianiga, M.; Andreis, R.; Raimondi, B. Seed size, shape and persistence in soil: a test on Italian flora from Alps to Mediterranean coasts. Seed Sci. Res. 2003, 13, 75–85. [Google Scholar] [CrossRef]
- Peco, B.; Traba, J.; Levassor, C.; Sánchez, A.M.; Azcárate, F.M. Seed size, shape and persistence in dry Mediterranean grass and scrublands. Seed Sci. Res. 2003, 13, 87–95. [Google Scholar] [CrossRef]
- Yu, S.; Sternberg, M.; Kutiel, P.; Chen, H. Seed mass, shape, and persistence in the soil seed bank of Israeli coastal sand dune flora. Evol. Ecol. Res. 2007, 9, 325–340. [Google Scholar]
- Cervantes, E.; Martín, J.J.; Saadaoui, E. Updated methods for seed shape. Scientifica 2016, 2106, 5691825. [Google Scholar] [CrossRef]
- Cervantes, E.; Gómez, J.J.M. Seed shape description and quantification by comparison with geometric models. Horticulturae 2019, 5, 60. [Google Scholar] [CrossRef]
- Cervantes, E.; Gómez, J.J.M.; Pozo, D.G.; Dias, L.S. An Angiosperm species dataset reveals relationships between seed size and two-dimensional shape. Horticulturae 2019, 5, 71. [Google Scholar] [CrossRef]
- Salisbury, E.J. The Reproductive Capacity of Plants; G. Bell and Sons: London, UK, 1942. [Google Scholar]
- Seed Volume Dataset. Available online: http://home.uevora.pt/~lsdias/SVDS_03plus.csv (accessed on 21 May 2020).
- Casco, H.; Dias, L.S. Estimating seed mass and volume from linear dimensions of seeds. Seed Sci. Technol. 2008, 36, 230–236. [Google Scholar] [CrossRef]
- Dias, L.S.; Ganhão, E. Extending the range for accurate estimation of seed volume from incomplete linear dimension data. Seed Sci. Technol. 2012, 40, 129–133. [Google Scholar] [CrossRef]
- Bakker, J.P.; Poschlod, P.; Strykstra, R.J.; Bekker, R.M.; Thompson, K. Seed banks and seed dispersal: important topics in restoration ecology. Acta Bot. Neerl. 1996, 45, 461–490. [Google Scholar] [CrossRef]
- World Flora Online. Available online: http://www.worldfloraonline.org/ (accessed on 19 February 2020).
- Breiman, L.; Friedman, J.H.; Olshen, R.A.; Stone, C.J. Classification and Regression Trees; Chapman & Hall/CRC: Boca Raton, FL, USA, 1984. [Google Scholar]
- Gueguen, A.; Nakache, J.P. Méthode de discrimination basée sur la construction d’un arbre de décision binaire. Rev. Stat. Appl. 1988, 36, 19–38. [Google Scholar]
- Lebart, L.; Morineau, A.; Piron, M. Statistique Exploratoire Multidimensionelle, 3rd ed.; Dunod: Paris, France, 2000. [Google Scholar]
- Ury, H.K. A comparison of four procedures for multiple comparisons among means (pairwise contrasts) for arbitrary sample sizes. Technometrics 1976, 18, 89–97. [Google Scholar] [CrossRef]
- Gardner, W.A. Effect of light on germination of light-sensitive species. Bot. Gaz. 1921, 71, 249–288. [Google Scholar] [CrossRef]
- Probert, R.J. The role of temperature in the regulation of seed dormancy and germination. In Seeds. The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI Publishing: Wallingford, UK, 2000; pp. 261–292. [Google Scholar]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva Dias, L.; Ganhão, E.; Soveral Dias, A. Responses of Germination to Light and to Far-Red Radiation—Can they be Predicted from Diaspores Size? Data 2020, 5, 49. https://doi.org/10.3390/data5020049
Silva Dias L, Ganhão E, Soveral Dias A. Responses of Germination to Light and to Far-Red Radiation—Can they be Predicted from Diaspores Size? Data. 2020; 5(2):49. https://doi.org/10.3390/data5020049
Chicago/Turabian StyleSilva Dias, Luís, Elsa Ganhão, and Alexandra Soveral Dias. 2020. "Responses of Germination to Light and to Far-Red Radiation—Can they be Predicted from Diaspores Size?" Data 5, no. 2: 49. https://doi.org/10.3390/data5020049
APA StyleSilva Dias, L., Ganhão, E., & Soveral Dias, A. (2020). Responses of Germination to Light and to Far-Red Radiation—Can they be Predicted from Diaspores Size? Data, 5(2), 49. https://doi.org/10.3390/data5020049