Dataset on Substrate-Borne Vibrations of Constrictotermes cyphergaster (Blattodea: Isoptera) Termites

 , , ,
, , ,
Abstract
1. Summary
2. Data Descriptor
| time: | the moment, from the beginning of the recording session, when vibration was recorded |
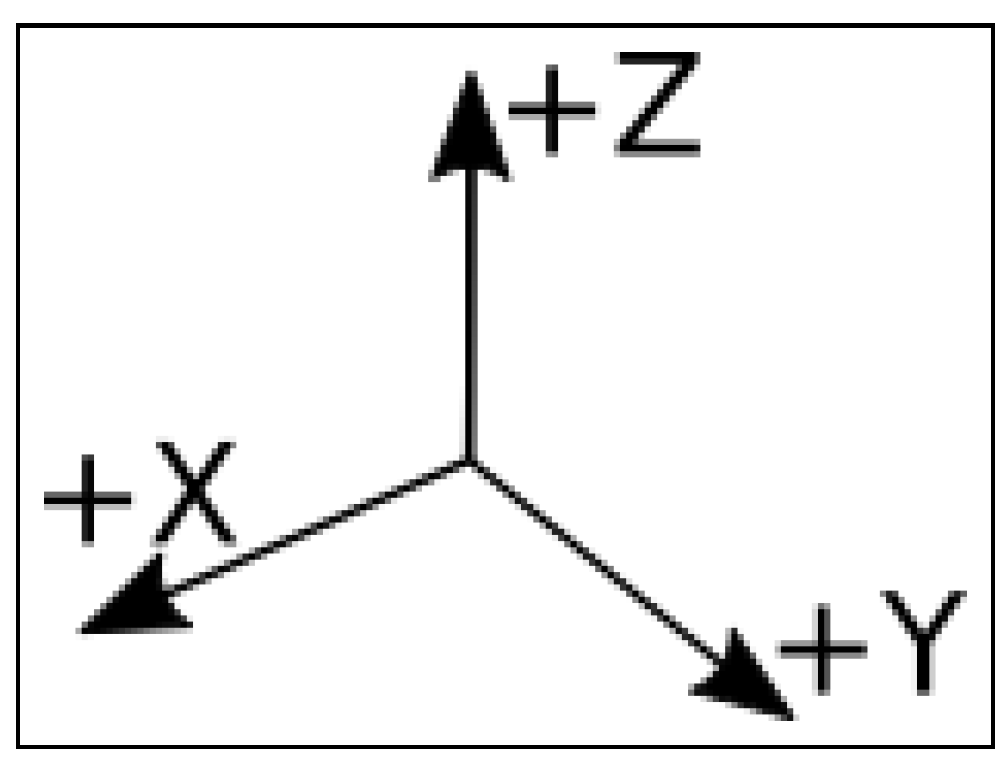
| Ax: | counts read by the accelerometer at the horizontal x axis |
| Ay: | counts read by the accelerometer at the horizontal y axis |
| Az: | counts read by the accelerometer at the vertical z axis |
| NestID: | the field identification of the nest from which the termite group was collected. Codes within NestID column are built as nnncccyyyy, where nnn = the nest sequential number used as a field label; ccc = the initials of the collector (name and surname); yyyy = the year in which the nest was taken from the field to the lab. |
| TypeStimulus: | the type of stimulus that was deposited within the injection chamber to be carried by the air injected into the arena to trigger termite vibrations. Codes within TypeStimulus are: air = only air; air_paper = air plus a piece of filter paper was deposited into the injection chamber and the air therein was injected into the arena; air_paper_hexane = a piece of filter paper onto which hexane was applied was deposited into the injection chamber and the air therein was injected into the arena; air_paper_extract = a piece of filter paper onto which a hexane extract from soldiers’ heads was applied was deposited into the injection chamber and the air therein was injected into the arena. |
| TermiteGroup: | the identification of the nest from which the termite group was collected and the stimulus they were exposed to. Codes within the TermiteGroup column are built as gnns, where: g = “group”; nn = the nest sequential number at which the termite group was collected; s = the stimulus that the termite group was exposed to. |
3. Methods
3.1. Ethical Statement
3.2. Termite Material
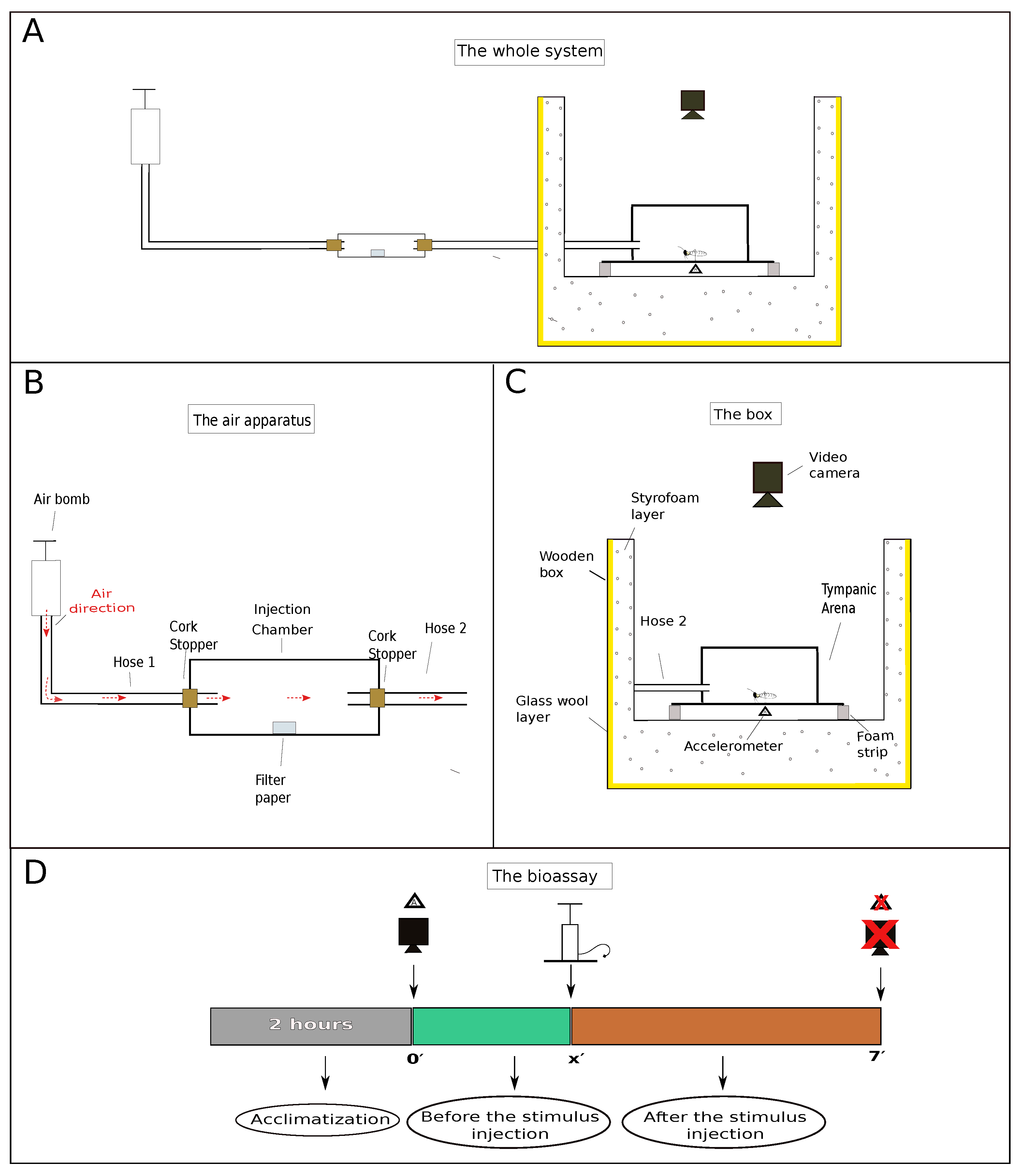
3.3. Experimental Procedures
- the air present in the injection chamber;
- the air present in the injection chamber after it had contact with a piece of filter paper (7 × 2 cm) previously deposited therein;
- the air present in the injection chamber after it had contact with a known amount of hexane that was loaded onto a piece of filter paper deposited in the injection chamber;
- the air present in the injection chamber after it had contact with hexane extracts of termite soldier heads. These extracts were loaded onto a piece of filter paper deposited in the injection chamber.
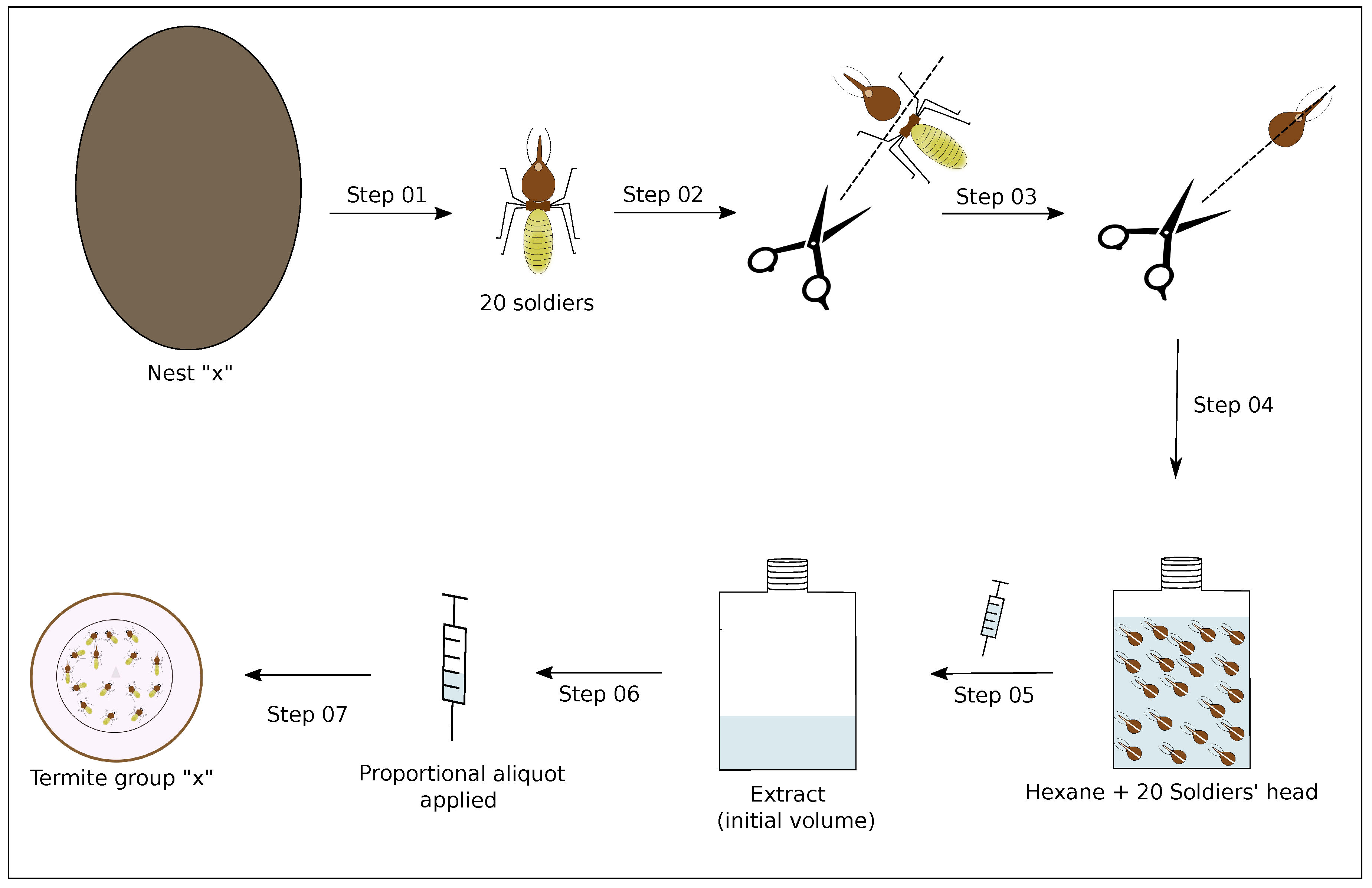
3.4. Extract Preparation
3.5. Behavioral Response and Parameters Measured
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hill, P.S. Vibration and animal communication: A review. Am. Zool. 2001, 41, 1135–1142. [Google Scholar] [CrossRef]
- Castellanos, I.; Barbosa, P. Evaluation of predation risk by a caterpillar using substrate-borne vibrations. Anim. Behav. 2006, 72, 461–469. [Google Scholar] [CrossRef]
- Fertin, A.; Casas, J. Orientation towards prey in antlions: Efficient use of wave propagation in sand. J. Exp. Biol. 2007, 210, 3337–3343. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Birch, M.C.; Wyatt, T.D. Mate location in the deathwatch beetle, Xestobium Rufovillosum De Geer (Anobiidae): Orientat. Substrate Vib. Anim. Behav. 1994, 47, 899–907. [Google Scholar] [CrossRef][Green Version]
- Schneider, S.S.; Stamps, J.A.; Gary, N.E. The vibration dance of the honey bee. I. Communication regulating foraging on two time scales. Anim. Behav. 1986, 34, 377–385. [Google Scholar] [CrossRef]
- Cristaldo, P.F.; Jandák, V.; Kutalová, K.; Rodrigues, V.B.; Brothánek, M.; Jiříček, O.; DeSouza, O.; Šobotník, J. The nature of alarm communication in Constrictotermes cyphergaster (Blattodea: Termitoidea: Termitidae): The integration of chemical and vibroacoustic signals. Biol. Open 2015, 4, 1649–1659. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.A.; Lai, J.C.; Toledano, E.; McDowall, L.; Rakotonarivo, S.; Lenz, M. Termites assess wood size by using vibration signals. Proc. Natl. Acad. Sci. USA 2005, 102, 3732–3737. [Google Scholar] [CrossRef] [PubMed]
- Oberst, S.; Bann, G.; Lai, J.; Evans, T.A. Cryptic termites avoid predatory ants by eavesdropping on vibrational cues from their footsteps. Ecol. Lett. 2017, 20, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Nunes, L.F.; Roxinol, J.A.; Cristaldo, P.F.; Marinho, R.; DeSouza, O. The use of tympanic arena as an alternative for behavioral vibroacoustic essays in termites (Blattodea: Isoptera). Sociobiology 2018, 65, 101–107. [Google Scholar] [CrossRef]
- Cunha, H.F.D.; Andrade Costa, D.; Espirito Santo Filho, K.D.; Silva, L.O.; Brandão, D. Relationship between Constrictotermes Cyphergaster Inquiline Termit. Cerrado (Isoptera: Termitidae). Sociobiology 2003, 42, 761–770. [Google Scholar]
- DeSouza, O.; Miramontes, O.; Santos, C.; Bernardo, D. Social facilitation affecting tolerance to poisoning in termites (Insecta, Isoptera). Insectes Sociaux 2001, 48, 21–24. [Google Scholar] [CrossRef]
- Šobotník, J.; Jirošová, A.; Hanus, R. Chemical warfare in termites. J. Insect Physiol. 2010, 56, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Costa-Leonardo, A.M.; Haifig, I. Pheromones and exocrine glands in Isoptera. Vitam. Horm. 2010, 83, 521–549. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Nest | Stimulus | Injection Time (s) | Aliquot Applied (μL) | Head Equivalence |
|---|---|---|---|---|
| N01YCC2017 | A | 120 | 0.00 | 0 |
| AP | 120 | 0.00 | 0 | |
| APH | 120 | 3.26 | 0 | |
| APE | 120 | 3.26 | 1 | |
| N02YCC2017 | A | 122 | 0.00 | 0 |
| AP | 120 | 0.00 | 0 | |
| APH | 120 | 6.69 | 0 | |
| APE | 122 | 6.69 | 3 | |
| N03YCC2017 | A | 120 | 0.00 | 0 |
| AP | 120 | 0.00 | 0 | |
| APH | 120 | 13.5 | 0 | |
| APE | 140 | 13.5 | 5 | |
| N04YCC2017 | A | 129 | 0.00 | 0 |
| AP | 120 | 0.00 | 0 | |
| APH | 125 | 24.68 | 0 | |
| APE | 123 | 24.68 | 7 | |
| N05YCC2017 | A | 123 | 0.00 | 0 |
| AP | – | – | – | |
| APH | 120 | 47.57 | 0 | |
| APE | 123 | 47.57 | 9 | |
| N06YCC2017 | A | 122 | 0.00 | 0 |
| AP | 120 | 0.00 | 0 | |
| APH | 120 | 4.61 | 0 | |
| APE | 120 | 4.61 | 1 | |
| N07YCC2017 | A | 122 | 0.00 | 0 |
| AP | 144 | 0.00 | 0 | |
| APH | 120 | 11.98 | 0 | |
| APE | 120 | 11.98 | 3 | |
| N08YCC2017 | A | 122 | 0.00 | 0 |
| AP | 120 | 0.00 | 0 | |
| APH | 120 | 28.9 | 0 | |
| APE | 180 | 28.9 | 5 | |
| N09YCC2017 | A | 120 | 0.00 | 0 |
| AP | 120 | 0.00 | 0 | |
| APH | 120 | 25.2 | 0 | |
| APE | 123 | 25.2 | 7 | |
| N10YCC2017 | A | 120 | 0.00 | 0 |
| AP | – | – | – | |
| APH | 126 | 49.32 | 0 | |
| APE | 122 | 49.32 | 9 | |
| N11YCC2017 | A | 120 | 0.00 | 0 |
| AP | 120 | 0.00 | 0 | |
| APH | 120 | 1.6 | 0 | |
| APE | 120 | 1.6 | 1 | |
| N12YCC2017 | A | 120 | 0.00 | 0 |
| AP | 120 | 0.00 | 0 | |
| APH | 122 | 15.54 | 0 | |
| APE | 120 | 15.54 | 3 | |
| N13YCC2017 | A | 120 | 0.00 | 0 |
| AP | 120 | 0.00 | 0 | |
| APH | 120 | 24.5 | 0 | |
| APE | 120 | 24.5 | 5 | |
| N14YCC2017 | A | 120 | 0.00 | 0 |
| AP | 120 | 0.00 | 0 | |
| APH | 113 | 20.79 | 0 | |
| APE | 141 | 20.79 | 7 | |
| N15YCC2017 | A | 120 | 0.00 | 0 |
| AP | 120 | 0.00 | 0 | |
| APH | 141 | 45.36 | 0 | |
| APE | 120 | 45.36 | 9 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fonseca Nunes, L.; Fellipe Cristaldo, P.; Sérgio Silva, P.; Bonato Felix, L.; Miranda Ribeiro, D.; DeSouza, O. Dataset on Substrate-Borne Vibrations of Constrictotermes cyphergaster (Blattodea: Isoptera) Termites. Data 2019, 4, 87. https://doi.org/10.3390/data4020087
Fonseca Nunes L, Fellipe Cristaldo P, Sérgio Silva P, Bonato Felix L, Miranda Ribeiro D, DeSouza O. Dataset on Substrate-Borne Vibrations of Constrictotermes cyphergaster (Blattodea: Isoptera) Termites. Data. 2019; 4(2):87. https://doi.org/10.3390/data4020087
Chicago/Turabian StyleFonseca Nunes, Lívia, Paulo Fellipe Cristaldo, Pedro Sérgio Silva, Leonardo Bonato Felix, Danilo Miranda Ribeiro, and Og DeSouza. 2019. "Dataset on Substrate-Borne Vibrations of Constrictotermes cyphergaster (Blattodea: Isoptera) Termites" Data 4, no. 2: 87. https://doi.org/10.3390/data4020087
APA StyleFonseca Nunes, L., Fellipe Cristaldo, P., Sérgio Silva, P., Bonato Felix, L., Miranda Ribeiro, D., & DeSouza, O. (2019). Dataset on Substrate-Borne Vibrations of Constrictotermes cyphergaster (Blattodea: Isoptera) Termites. Data, 4(2), 87. https://doi.org/10.3390/data4020087