Designing New Yeasts for Craft Brewing: When Natural Biodiversity Meets Biotechnology



Abstract

1. Introduction
2. Brewing Yeasts Through the Lens of Genomics
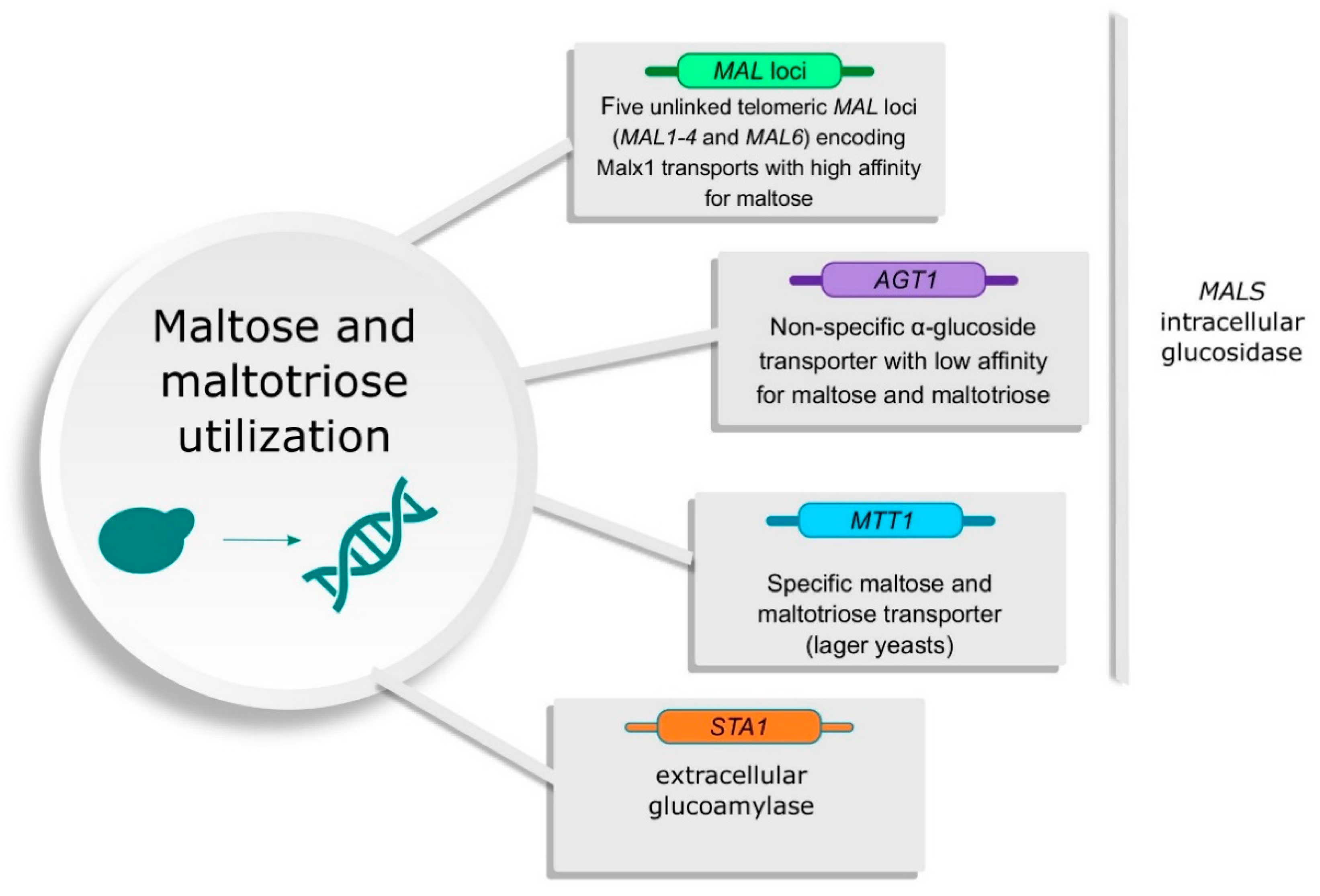
2.1. Saccharomyces cerevisiae Ale Yeasts
2.2. Saccharomyces Pastorianus
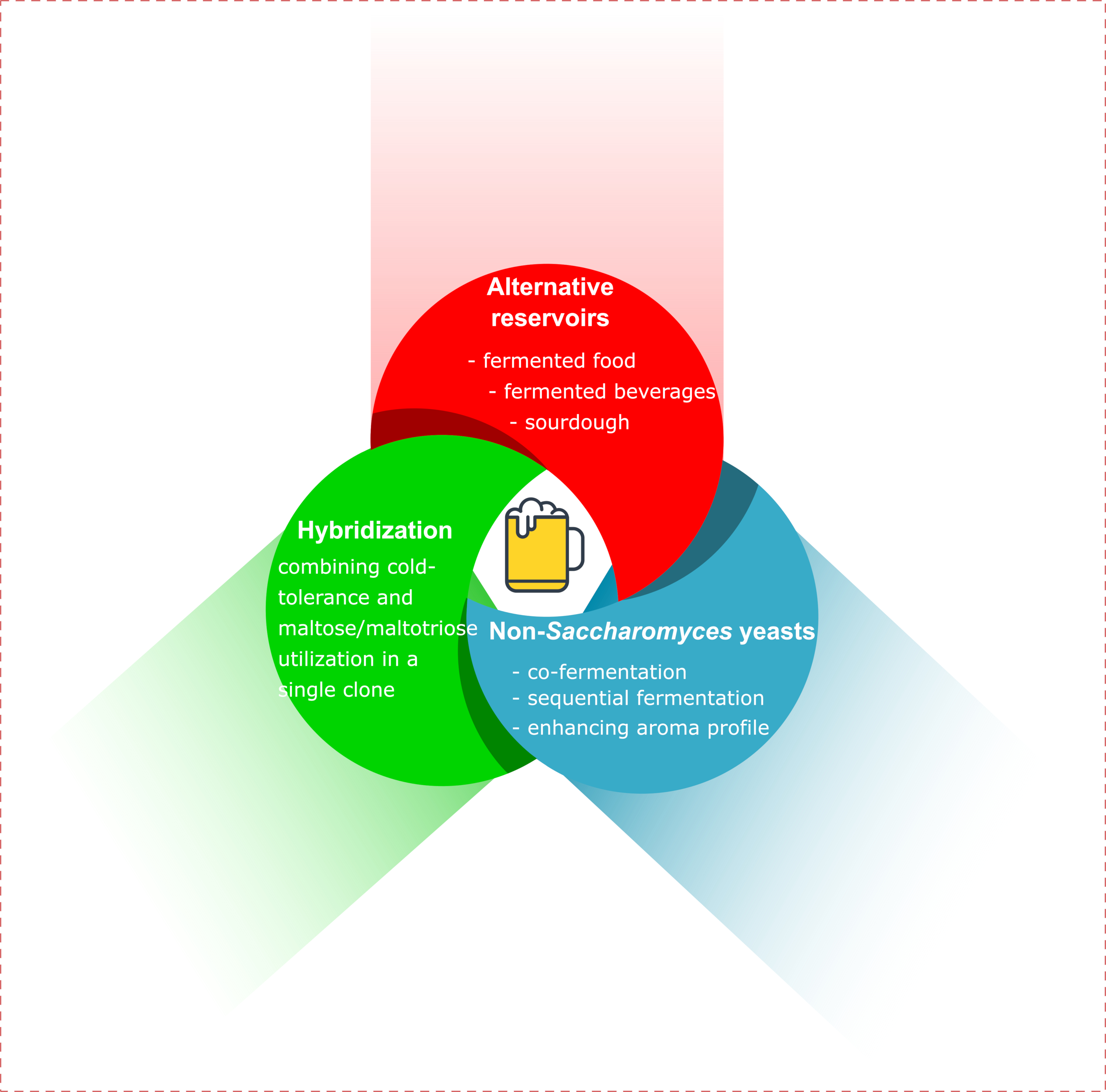
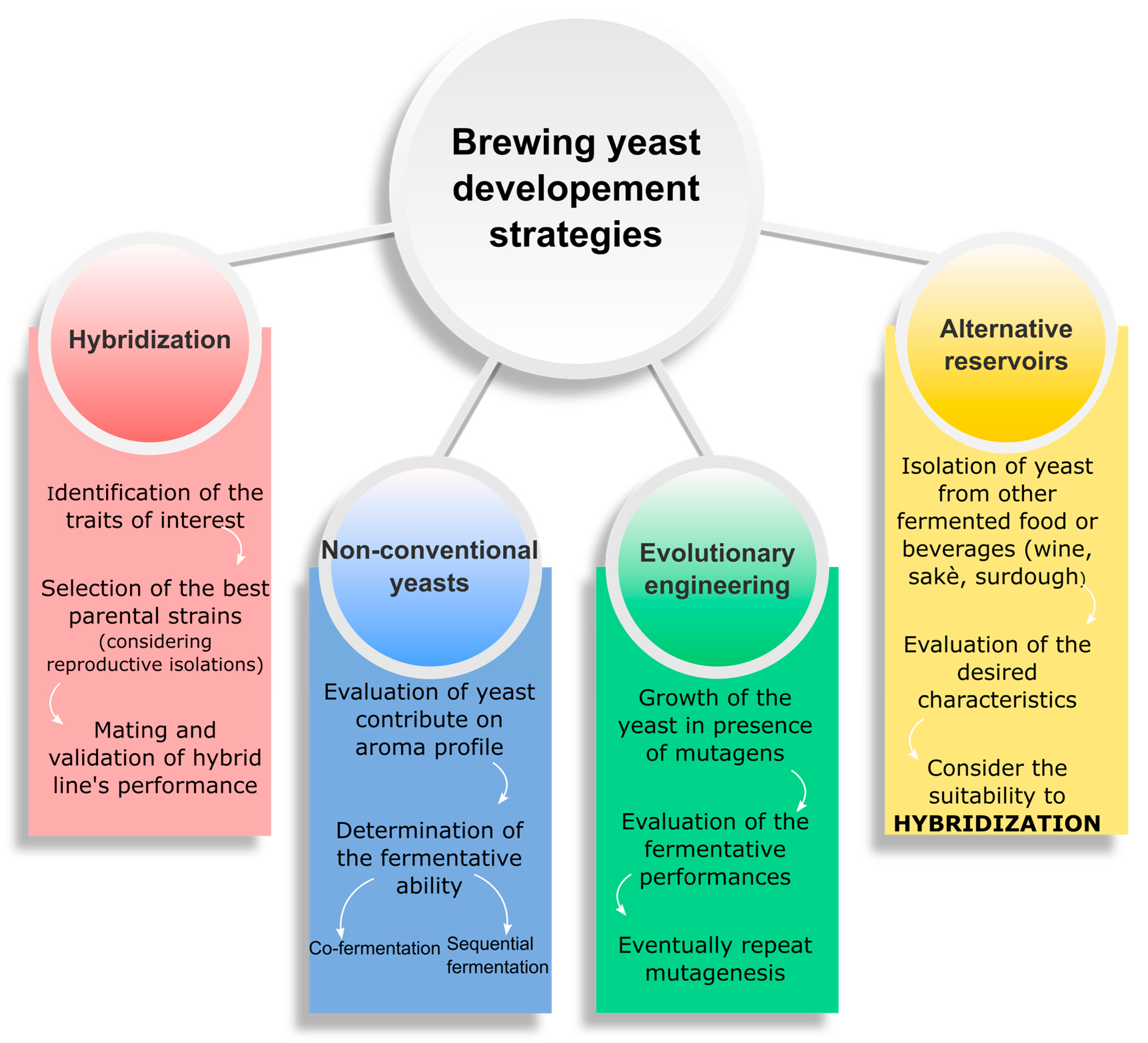
3. Mimic of Lager Yeasts by Artificial Hybridization
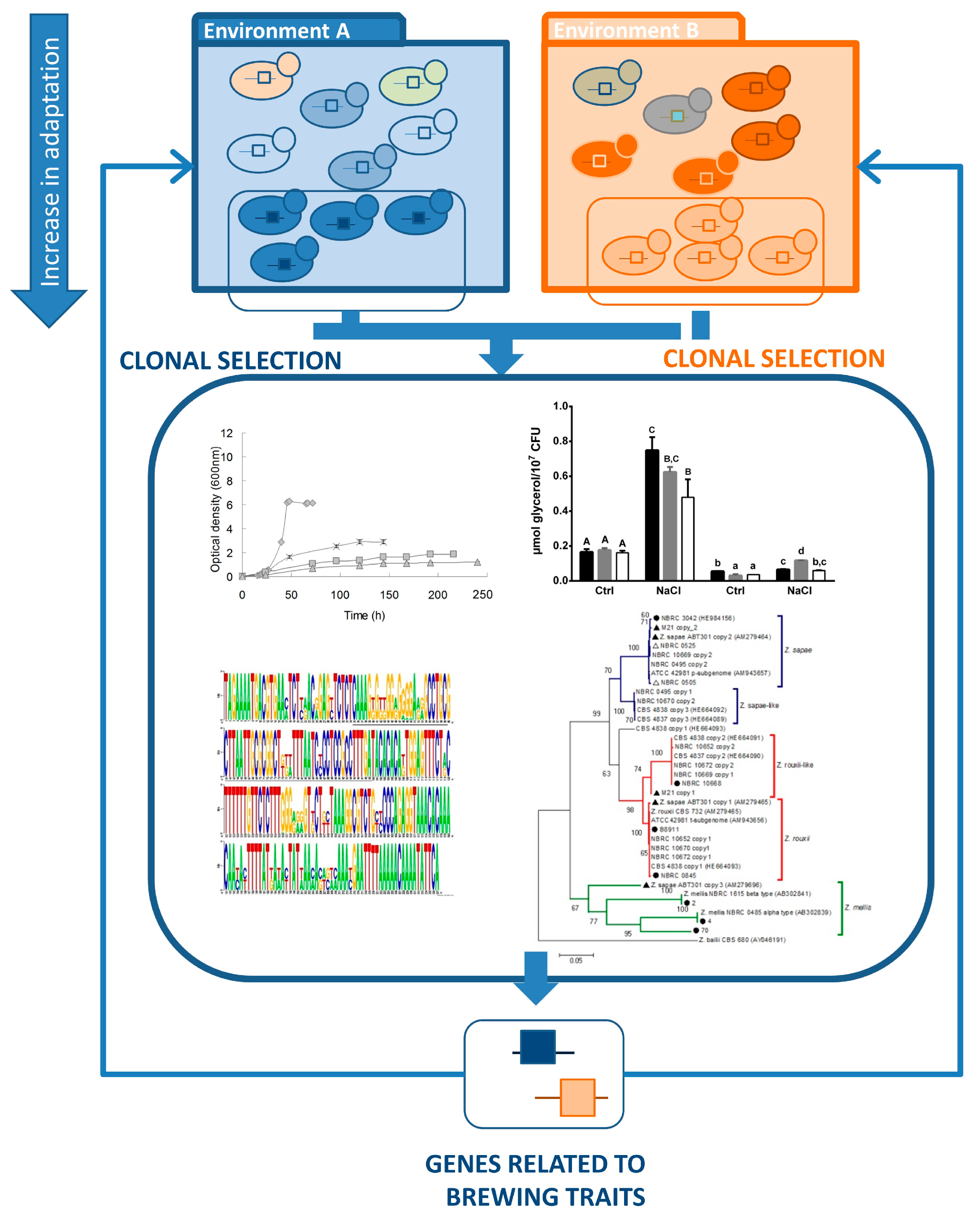
4. Evolutionary Engineering
5. Fermented Food as Reservoir of Novel S. cerevisiae Brewing Starters
6. Non-Saccharomyces Yeasts
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hornsey, I.S. A History of Beer and Brewing; Royal Society of Chemistry: London, UK, 2003. [Google Scholar]
- Nelson, M. The Barbarian’s Beverage: A History of Beer in Ancient Europe; Routledge Taylor & Francis: London, UK, 2005. [Google Scholar]
- Unger, R.W. Beer in the Middle Ages and the Renaissance; University of Pennsylvania Press: Philadelphia, PA, USA, 2004. [Google Scholar]
- Carlos, B.A.; Caballero, I.; Barrios, R.; Rojas, A. Innovations in the brewing industry: Light beer. Int. J. Food Sci. Nutr. 2014, 65, 655–660. [Google Scholar]
- Sicard, D.; Legras, J.L. Bread, beer and wine: Yeast domestication in the Saccharomyces sensu stricto complex. Comptes Rendus Biol. 2011, 334, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Aquilani, B.; Laureti, T.; Poponi, S.; Secondi, L. Beer choice and consumption determinants when craft beers are tasted: An exploratory study of consumer preferences. Food Qual. Prefer. 2015, 41, 214–224. [Google Scholar] [CrossRef]
- Stack, M.; Gartland, M.; Keane, T. Path dependency, behavioral lock-in and the international market for beer. In Brewing, Beer and Pubs; Cabras, I., Higgins, D., Preece, D., Eds.; Palgrave Macmillan: London, UK, 2016. [Google Scholar]
- Corbo, M.R.; Bevilacqua, A.; Petruzzi, L.; Casanova, F.P.; Sinigaglia, M. Functional beverages: The emerging side of functional foods. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1192–1206. [Google Scholar] [CrossRef]
- Wesson, T.; Nieva de Figueiredo, J. The importance of focus to market: A study of microbrewery performance. J. Bus. Ventur. 2001, 16, 377–403. [Google Scholar] [CrossRef]
- Gómez-Corona, C.; Lelievre-Desmas, M.; Buendía, H.B.E.; Chollet, S.; Valentin, D. Craft beer representation amongst men in two different cultures. Food Qual. Prefer. 2016, 53, 19–28. [Google Scholar] [CrossRef]
- Garavaglia, C.; Swinnen, J. Economic Perspectives on Craft Beer: A Revolution in the Global Beer Industry; Palgrave Macmillan: London, UK; Springer: Cham, Switzerland, 2018; pp. 3–51. [Google Scholar]
- Elzinga, K.G.; Tremblay, C.H.; Tremblay, V.J. Craft beer in the United States: History, numbers, and geography. J. Wine Econ. 2015, 10, 242–274. [Google Scholar] [CrossRef]
- Hagemann, M.N.; Bogner, K.; Marchioni, E.; Braun, S. Chances for dry-hopped non-alcoholic beverages? Part 1: Concept and market prospects. Brew. Sci. 2016, 69, 50–55. [Google Scholar]
- Brewers of Europe. Beer Statistics 2016. Available online: www.brewersofeurope.org (accessed on 23 September 2019).
- Clemons, E.K.; Gao, G.G.; Hitt, L.M. When online reviews meet hyperdifferentiation: A study of the craft beer industry. J. Manag. Inf. Syst. 2006, 23, 149–171. [Google Scholar] [CrossRef]
- Brewers Association. Analyzing 2017 Craft Brewery Growth. Available online: www.brewersassociation.org (accessed on 12 September 2019).
- Oliver, G. The Oxford Companion to Beer, 1st ed.; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Dzialo, M.C.; Park, R.; Steensels, J.; Lievens, B.; Verstrepen, K.J. Physiology, ecology and industrial applications of aroma formation in yeast. FEMS Microbiol. Rev. 2017, 41, S95–S128. [Google Scholar] [CrossRef]
- Pasteur, L. Études sur la bière, ses maladies, causes qui les provoquent, procédé pour la rendre inalterable. In Avec une Théorie Nouvelle de la Fermentation; Gauthier-Villars: Paris, France, 1876. [Google Scholar]
- Barnett, J.A.; Lichtenthaler, F.W. A history of research on yeasts 3: Emil Fischer, Eduard Buchner and their contemporaries, 1880–1900. Yeast 2001, 18, 363–388. [Google Scholar] [CrossRef]
- Gallone, B.; Steensels, J.; Baele, G.; Maere, S.; Verstrepen, K.J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; et al. Domestication and divergence of Saccharomyces cerevisiae beer yeasts. Cell 2016, 166, 1397.e16–1410.e16. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.; Pontes, A.; Almeida, P.; Barbosa, R.; Serra, M.; Libkind, D.; Hutzler, M.; Gonçalves, P.; Sampaio, J.P. Distinct domestication trajectories in top-fermenting beer yeasts and wine yeasts. Curr. Biol. 2016, 26, 2750–2761. [Google Scholar] [CrossRef] [PubMed]
- Peter, J.; De Chiara, M.; Friedrich, A.; Yue, J.-X.; Pflieger, D.; Bergström, A.; Sigwalt, A.; Barre, B.; Freel, K.; Llored, A.; et al. Genome evolution across 1,011 Saccharomyces cerevisiae isolates. Nature 2018, 556, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Selmecki, A.M.; Maruvka, Y.E.; Richmond, P.A.; Guillet, M.; Shoresh, N.; Sorenson, A.L.; De, S.; Kishony, R.; Michor, F.; Dowell, R.; et al. Polyploidy can drive rapid adaptation in yeast. Nature 2015, 519, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Yona, A.H.; Manor, Y.S.; Herbst, R.H.; Romano, G.H.; Mitchell, A.; Kupiec, M.; Pilpel, Y.; Dahan, O. Chromosomal duplication is a transient evolutionary solution to stress. Proc. Natl. Acad. Sci. USA 2012, 109, 21010–21015. [Google Scholar] [CrossRef]
- Horak, J. Regulations of sugar transporters: Insights from yeast. Curr. Genet. 2013, 59, 1–31. [Google Scholar] [CrossRef]
- Rautio, J.; Londesborough, J. Maltose transport by brewer’s yeast in brewer’s wort. J. Inst. Brew. 2003, 109, 251–261. [Google Scholar] [CrossRef]
- Dietvorst, J.; Londesborough, J.; Steensma, H.Y. Maltotriose utilization by lager yeast strains: MTT1 encodes a maltotriose transporter. Yeast 2005, 22, 775–788. [Google Scholar] [CrossRef]
- Stambuk, B.U.; Da Silva, M.A.; Panek, A.D.; De Araujo, P.S. Active α-glucoside transport in Saccharomyces cerevisiae. FEMS Microbiol. Lett. 1999, 170, 105–110. [Google Scholar] [CrossRef]
- Vidgren, V.; Ruohonen, L.; Londesborough, J. Characterization and functional analysis of the MAL and MPH Loci for maltose utilization in some ale and lager yeast strains. Appl. Environ. Microbiol. 2005, 71, 7846–7857. [Google Scholar] [CrossRef] [PubMed]
- Vidgren, V.; Multanen, J.P.; Ruohonen, L.; Londesborough, J. The temperature dependence of maltose transport in ale and lager strains of brewer’s yeast. FEMS Yeast Res. 2010, 10, 402–411. [Google Scholar] [CrossRef]
- Krogerus, K.; Magalhães, F.; Kuivanen, J.; Gibson, B. A deletion in the STA1 promoter determines maltotriose and starch utilization in STA1 Saccharomyces cerevisiae strains. Appl. Microbiol. Biotechnol. 2019, 103, 7597–7615. [Google Scholar] [CrossRef]
- Ogata, T.; Iwashita, Y.; Kawada, T. Construction of a brewing yeast expressing the glucoamylase gene STA1 by mating. J. Inst. Brew. 2017, 123, 66–69. [Google Scholar] [CrossRef]
- Mukai, N.; Masaki, K.; Fujii, T.; Kawamukai, M.; Iefuji, H. PAD1 and FDC1 are essential for the decarboxylation of phenylacrylic acids in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2010, 109, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Richard, P.; Viljanen, K.; Penttila, M. Overexpression of PAD1 and FDC1 results in significant cinnamic acid decarboxylase activity in Saccharomyces cerevisiae. AMB Express 2015, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Will, J.L.; Kim, H.S.; Clarke, J.; Painter, J.C.; Fay, J.C.; Gasch, A.P. Incipient balancing selection through adaptive loss of aquaporins in natural Saccharomyces cerevisiae populations. PLoS Genet. 2010, 6, e1000893. [Google Scholar] [CrossRef] [PubMed]
- Hansen, E.C. Recherches sur la physiologie et la morphologie des ferments alcooliques. XIII. Nouvelles etudes sur des levures de brasserie a fermentation basse. C. R. Trav. Lab. Carlsberg 1908, 7, 179–217. [Google Scholar]
- Rainieri, S.; Kodama, Y.; Kaneko, Y.; Mikata, K.; Nakao, Y.; Ashikari, T. Pure and mixed genetic lines of Saccharomyces bayanus and Saccharomyces pastorianus and their contribution to the lager brewing strain genome. Appl. Environ. Microbiol. 2006, 72, 3968–3974. [Google Scholar] [CrossRef]
- Nakao, Y.; Kanamori, T.; Itoh, T.; Kodama, Y.; Rainieri, S.; Nakamura, N.; Shimonaga, T.; Hattori, M.; Shimonaga, T.; Hattori, M.; et al. Genome sequence of the lager brewing yeast, an interspecies hybrid. DNA Res. 2009, 16, 115–129. [Google Scholar] [CrossRef]
- Libkind, D.; Hittinger, C.T.; Valério, E.; Gonçalves, C.; Dover, J.; Johnston, M.; Gonçalves, P.; Sampaio, J.P. Microbe domestication and the identification of the wild genetic stock of lager-brewing yeast. Proc. Natl. Acad. Sci. USA 2011, 108, 14539–14544. [Google Scholar] [CrossRef] [PubMed]
- Peris, D.; Sylvester, K.; Libkind, D.; Gonçalves, P.; Sampaio, J.P.; Alexander, W.G.; Hittinger, C.T. Population structure and reticulate evolution of Saccharomyces eubayanus and its lager-brewing hybrids. Mol. Ecol. 2014, 23, 2031–2045. [Google Scholar] [CrossRef] [PubMed]
- Peris, D.; Langdon, Q.K.; Moriarty, R.V.; Sylvester, K.; Bontrager, M.; Charron, G.; Leducq, J.B.; Landry, C.R.; Libkind, D.; Hittinger, C.T. Complex ancestries of lager-brewing hybrids were shaped by standing variation in the wild yeast Saccharomyces eubayanus. PLoS Genet. 2016, 12, e1006155. [Google Scholar] [CrossRef] [PubMed]
- Bing, J.; Han, P.-J.; Liu, W.Q.; Wang, Q.M.; Bai, F.Y. Evidence for a Far East Asian origin of lager beer yeast. Curr. Biol. 2014, 24, R380–R381. [Google Scholar] [CrossRef]
- Gayevskiy, V.; Goddard, M.R. Saccharomyces eubayanus and Saccharomyces arboricola reside in North Island native New Zealand forests. Environ. Microbiol. Rep. 2016, 18, 1137–1147. [Google Scholar] [CrossRef]
- Hebly, M.; Brickwedde, A.; Bolat, I.; Driessen, M.R.M.; De Hulster, E.A.F.; Van den Broek, M.; Pronk, J.T.; Geertman, J.M.; Daran, J.M.; Daran-Lapujade, P. S. cerevisiae × S. eubayanus interspecific hybrid, the best of both worlds and beyond. FEMS Yeast Res. 2015, 15, 1–14. [Google Scholar] [CrossRef]
- Fischer, G.; James, S.A.; Roberts, I.N.; Oliver, S.G.; Louis, E.J. Chromosomal evolution in Saccharomyces. Nature 2000, 405, 451–454. [Google Scholar] [CrossRef]
- Lee, H.Y.; Chou, J.Y.; Cheong, L.; Chang, N.H.; Yang, S.Y.; Leu, J.Y. Incompatibility of nuclear and mitochondrial genomes causes hybrid sterility between two yeast species. Cell 2008, 135, 1065–1073. [Google Scholar] [CrossRef]
- Bond, U.; Neal, C.; Donnelly, D.; James, T.C. Aneuploidy and copy number breakpoints in the genome of lager yeasts mapped by microarray hybridisation. Curr. Genet. 2004, 45, 360–370. [Google Scholar] [CrossRef]
- Van den Broek, M.; Bolat, I.; Nijkamp, J.F.; Ramos, E.; Luttik, M.A.; Koopman, F.; Geertman, J.M.; De Ridder, D.; Pronk, J.T.; Daran, J.M. Chromosomal copy number variation in Saccharomyces pastorianus is evidence for extensive genome dynamics in industrial lager brewing strains. Appl. Environ. Microbiol. 2015, 81, 6253–6267. [Google Scholar] [CrossRef]
- Rainieri, S.; Kodama, Y.; Nakao, Y.; Pulvirenti, A.; Giudici, P. The inheritance of mtDNA in lager brewing strains. FEMS Yeast Res. 2008, 8, 586–596. [Google Scholar] [CrossRef]
- Solieri, L. Mitochondrial inheritance in budding yeasts: Towards an integrated understanding. Trends Microbiol. 2010, 18, 521–530. [Google Scholar] [CrossRef]
- Baker, E.; Wang, B.; Bellora, N.; Peris, D.; Hulfachor, A.B.; Koshalek, J.A.; Adams, M.; Libkind, D.; Hittinger, C.T. The genome sequence of Saccharomyces eubayanus and the domestication of lager-brewing yeasts. Mol. Biol. Evol. 2015, 32, 2818–2831. [Google Scholar] [CrossRef]
- Okuno, M.; Kajitani, R.; Ryusui, R.; Morimoto, H.; Kodama, Y.; Itoh, T. Next-generation sequencing analysis of lager brewing yeast strains reveals the evolutionary history of interspecies hybridization. DNA Res. 2016, 23, 67–80. [Google Scholar] [CrossRef]
- Li, X.C.; Peris, D.; Hittinger, C.T.; Sia, E.A.; Fay, J.C. Mitochondria-encoded genes contribute to evolution of heat and cold tolerance in yeast. Sci. Adv. 2019, 5, eaav1848. [Google Scholar] [CrossRef]
- Baker, E.C.P.; Peris, D.; Moriarty, R.V.; Li, X.C.; Fay, J.C.; Hittinger, C.T. Mitochondrial DNA and temperature tolerance in lager yeasts. Sci. Adv. 2019, 5, eaav1869. [Google Scholar] [CrossRef]
- Liti, G.; Peruffo, A.; James, S.A.; Roberts, I.N.; Louis, E.J. Inferences of evolutionary relationships from a population survey of LTR-retrotransposons and telomeric-associated sequences in the Saccharomyces sensu stricto complex. Yeast 2005, 22, 177–192. [Google Scholar] [CrossRef]
- Dunn, B.; Sherlock, G. Reconstruction of the genome origins and evolution of the hybrid lager yeast Saccharomyces pastorianus. Genome Res. 2008, 18, 1610–1623. [Google Scholar] [CrossRef]
- Monerawela, C.; Bond, U. Brewing up a storm: The genomes of lager yeasts and how they evolved. Biotechnol. Adv. 2017, 35, 512–519. [Google Scholar] [CrossRef]
- Gibson, B.R.; Storgards, E.; Krogerus, K.; Vidgren, V. Comparative physiology and fermentation performance of Saaz and Frohberg lager yeast strains and the parental species Saccharomyces eubayanus. Yeast 2013, 30, 255–266. [Google Scholar] [CrossRef]
- Walther, A.; Hesselbart, A.; Wendland, J. Genome sequence of Saccharomyces carlsbergensis, the world’s first pure culture lager yeast. G3 2014, 4, 783–793. [Google Scholar] [CrossRef]
- Hewitt, S.K.; Donaldson, I.J.; Lovell, S.C.; Delneri, D. Sequencing and characterization of rearrangements in three S. pastorianus strains reveals the presence of chimeric genes and gives evidence of breakpoint reuse. PLoS ONE 2014, 9, e92203. [Google Scholar] [CrossRef]
- Tafer, H.; Sterflinger, K.; Lopandic, K. Draft genome sequence of the interspecies hybrid Saccharomyces pastorianus strain HA2560, isolated from a municipal wastewater treatment plant. Genome Announc. 2018, 6, e00341-18. [Google Scholar] [CrossRef]
- Monerawela, C.; James, T.C.; Wolfe, K.H.; Bond, U. Loss of lager specific genes and subtelomeric regions define two different Saccharomyces cerevisiae lineages for Saccharomyces pastorianus Group I and II strains. FEMS Yeast Res. 2015, 15, fou008. [Google Scholar] [CrossRef]
- Salazar, A.N.; De Vries, A.G.; Van den Broek, M.; Brouwers, N.; De la Torre Cortes, P.; Kuijpers, N.; Daran, J.M.G.; Abeel, T. Nanopore sequencing and comparative genome analysis confirm lager-brewing yeasts originated from a single hybridization. bioRxiv 2019, 603480. [Google Scholar] [CrossRef]
- Salema-Oom, M.; Pinto, V.V.; Goncalves, P.; Spencer-Martins, I. Maltotriose utilization by industrial Saccharomyces strains: Characterization of a new member of the α-glucoside transporter family. Appl. Environ. Microbiol. 2005, 71, 5044–5049. [Google Scholar] [CrossRef]
- Magalhães, F.; Vidgren, V.; Ruohonen, L.; Gibson, B. Maltose and maltotriose utilisation by group I strains of the hybrid lager yeast Saccharomyces pastorianus. FEMS Yeast Res. 2016, 16, fow053. [Google Scholar] [CrossRef]
- Baker, E.P.; Hittinger, C.T. Evolution of a novel chimeric maltotriose transporter in Saccharomyces eubayanus from parent proteins unable to perform this function. PLoS Genet. 2019, 15, e1007786. [Google Scholar] [CrossRef]
- Brouwers, N.; Gorter de Vries, A.R.; Van den Broek, M.; Weening, S.M.; Elink Schuurman, T.D.; Kuijpers, N.G.A.; Pronk, J.T.; Daran, J.G. In vivo recombination of Saccharomyces eubayanus maltose-transporter genes yields a chimeric transporter that enables maltotriose fermentation. PLoS Genet. 2019, 15, e1007853. [Google Scholar] [CrossRef]
- Brouwers, N.; Brickwedde, A.; Gorter de Vries, A.R.; Van den Broek, M.; Weening, S.M.; Van den Eijnden, L.; Diderich, J.A.; Bai, F.Y.; Pronk, J.T.; Daran, J.G. The genome sequences of Himalayan Saccharomyces eubayanus revealed genetic markers explaining heterotic maltotriose consumption by hybrid Saccharomyces pastorianus. Appl. Environ. Microbiol. 2019, AEM.01516. [Google Scholar] [CrossRef]
- Peris, D.; Lopes, C.A.; Belloch, C.; Querol, A.; Barrio, E. Comparative genomics among Saccharomyces cerevisiae × Saccharomyces kudriavzevii natural hybrid strains isolated from wine and beer reveals different origins. BMC Genom. 2012, 13, 407. [Google Scholar] [CrossRef]
- Nguyen, H.V.; Legras, J.L.; Neuvéglise, C.; Gaillardin, C. Deciphering the hybridisation history leading to the Lager lineage based on the mosaic genomes of Saccharomyces bayanus strains NBRC1948 and CBS380. PLoS ONE 2011, 6, e25821. [Google Scholar] [CrossRef]
- Giudici, P.; Solieri, L.; Pulvirenti, A.M.; Cassanelli, S. Strategies and perspectives for genetic improvement of wine yeasts. Appl. Microbiol. Biotechnol. 2005, 66, 622–628. [Google Scholar] [CrossRef]
- Steensels, J.; Snoek, T.; Meersman, E.; Picca Nicolino, M.; Voordeckers, K.; Verstrepen, K.J. Improving industrial yeast strains: Exploiting natural and artificial diversity. FEMS Microbiol. Rev. 2014, 38, 947–995. [Google Scholar] [CrossRef]
- Liti, G.; Barton, D.B.H.; Louis, E.J. Sequence diversity, reproductive isolation and species concepts in Saccharomyces. Genetics 2006, 174, 839–850. [Google Scholar] [CrossRef]
- Greig, D. Reproductive isolation in Saccharomyces. Heredity 2009, 102, 39–44. [Google Scholar] [CrossRef]
- Krogerus, K.; Magalhães, F.; Vidgren, V.; Gibson, B. New lager yeast strains generated by interspecific hybridization. J. Ind. Microbiol. Biotechnol. 2015, 42, 769–778. [Google Scholar] [CrossRef]
- Krogerus, K.; Arvas, M.; De Chiara, M.; Magalhães, F.; Mattinen, L.; Oja, M.; Vidgren, V.; Yue, J.X.; Liti, G.; Gibson, B. Ploidy influences the functional attributes of de novo lager yeast hybrids. Appl. Microbiol. Biotechnol. 2016, 100, 7203–7222. [Google Scholar] [CrossRef]
- Krogerus, K.; Seppänen-Laakso, T.; Castillo, S.; Gibson, B. Inheritance of brewing-relevant phenotypes in constructed Saccharomyces cerevisiae x Saccharomyces eubayanus hybrids. Microbiol. Cell Fact. 2017, 16, 66. [Google Scholar] [CrossRef]
- Krogerus, K.; Holmström, S.; Gibson, B. Enhanced wort fermentation with de novo lager hybrids adapted to high-ethanol environments. Appl. Environ. Microbiol. 2018, 84, e2302–e2317. [Google Scholar] [CrossRef]
- Mertens, S.; Steensels, J.; Saels, V.; De Rouck, G.; Aerts, G.; Verstrepen, K.J. A large set of newly created interspecific Saccharomyces hybrids increases aromatic diversity in lager beers. Appl. Environ. Microbiol. 2015, 81, 8202–8214. [Google Scholar] [CrossRef]
- Alexander, W.G.; Peris, D.; Pfannenstiel, B.T.; Opulente, D.A.; Kuang, M.; Hittinger, C.T. Efficient engineering of marker-free synthetic allotetraploids of Saccharomyces. Fungal Genet. Biol. 2016, 89, 10–17. [Google Scholar] [CrossRef]
- Diderich, J.A.; Weening, S.M.; Van den Broek, M.; Pronk, J.T.; Daran, J.G. Selection of Pof− Saccharomyces eubayanus variants for the construction of S. cerevisiae × S. eubayanus hybrids with reduced 4-vinyl guaiacol formation. Front. Microbiol. 2018, 9, 1640. [Google Scholar] [CrossRef]
- Mertens, S.; Gallone, B.; Steensels, J.; Herrera-Malaver, B.; Cortebeek, J.; Nolmans, R.; Saels, V.; Vyas, V.K.; Verstrepen, K.J. Reducing phenolic off-flavors through CRISPR-based gene editing of the FDC1 gene in Saccharomyces cerevisiae × Saccharomyces eubayanus hybrid lager beer yeasts. PLoS ONE 2019, 14, e0209124. [Google Scholar]
- Confédération Paysanne and Others v Premier Ministre and Ministre de l’Agriculture, de l’Agroalimentaire et de la Forêt; Judgment in Case C-528/16; Court of Justice of the European Union PRESS RELEASE No 111/18; Luxembourg. 2018. Available online: www./curia.europa.eu/juris/document/document.jsf?text=&docid=204387&pageIndex=0&doclang=EN&mode=req&dir=&occ=first&part=1&cid=193592 (accessed on 25 July 2018).
- Sato, M.; Kishimoto, M.; Watari, J.; Takashio, M. Breeding of brewer’s yeast by hybridization between a top-fermenting yeast Saccharomyces cerevisiae and a cryophilic yeast Saccharomyces bayanus. J. Biosci. Bioeng. 2002, 93, 509–511. [Google Scholar] [CrossRef]
- Nikulin, J.; Krogerus, K.; Gibson, B. Alternative Saccharomyces interspecies hybrid combinations and their potential for low-temperature wort fermentation. Yeast 2018, 35, 113–127. [Google Scholar] [CrossRef]
- Brickwedde, A.; Van den Broek, M.; Geertman, J.A.; Magalhães, F.; Kuijpers, N.G.A.; Gibson, B.; Pronk, J.T.; Daran, J.G. Evolutionary engineering in chemostat cultures for improved maltotriose fermentation kinetics in Saccharomyces pastorianus lager brewing yeast. Front. Microbiol. 2017, 8, 1690. [Google Scholar] [CrossRef]
- Gibson, B.; Vidgren, V.; Peddinti, G.; Krogerus, K. Diacetyl control during brewery fermentation via adaptive laboratory engineering of the lager yeast Saccharomyces pastorianus. J. Ind. Microbiol. Biotechnol. 2018, 45, 1103–1112. [Google Scholar] [CrossRef]
- Blieck, L.; Toye, G.; Dumortier, F.; Verstrepen, K.J.; Delvaux, F.R.; Thevelein, J.M.; Van Dijck, P. Isolation and characterization of brewer’s yeast variants with improved fermentation performance under high-gravity conditions. Appl. Environ. Microbiol. 2007, 73, 815–824. [Google Scholar] [CrossRef]
- Ekberg, J.; Rautio, J.; Mattinen, L.; Vidgren, V.; Londesborough, J.; Gibson, B.R. Adaptive evolution of the lager brewing yeast Saccharomyces pastorianus for improved growth under hyperosmotic conditions and its influence on fermentation performance. FEMS Yeast Res. 2013, 13, 335–349. [Google Scholar] [CrossRef]
- Huuskonen, A.; Markkula, T.; Vidgren, V.; Lima, L.; Mulder, L.; Geurts, W.; Walsh, M.; Londesborough, J. Selection from industrial lager yeast strains of variants with improved fermentation performance in very-high-gravity worts. Appl. Environ. Microbiol. 2010, 76, 1563–1573. [Google Scholar] [CrossRef]
- Yu, Z.; Zhao, H.; Li, H.; Zhang, Q.; Lei, H.; Zhao, M. Selection of Saccharomyces pastorianus variants with improved fermentation performance under very high-gravity wort conditions. Biotechnol. Lett. 2012, 34, 367–370. [Google Scholar] [CrossRef]
- Lee, S.; Villa, K.; Patino, H. Yeast strain development for enhanced production of desirable alcohols/esters in beer. J. Am. Soc. Brew. Chem. 1995, 53, 13–156. [Google Scholar] [CrossRef]
- Watanabe, M.; Tanaka, N.; Mishima, M.; Takemura, S. Isolation of sake yeast mutants resistant to isoamyl monofluoroacetate to improve isoamyl acetate productivity. J. Ferment. Bioeng. 1993, 76, 229–231. [Google Scholar] [CrossRef]
- Hirooka, K.; Yamamoto, Y.; Tsutsui, N.; Tanaka, T. Improved production of isoamyl acetate by a sake yeast mutant resistant to an isoprenoid analog and its dependence on alcohol acetyltransferase activity, but not on isoamyl alcohol production. J. Biosci. Bioeng. 2005, 99, 125–129. [Google Scholar] [CrossRef]
- Ichikawa, E.; Hosokawa, N.; Hata, Y.; Abe, Y.; Suginami, K.; Imayasu, S. Breeding of a sake yeast with improved ethyl caproate productivity. Agric. Biol. Chem. 1991, 55, 2153–2154. [Google Scholar]
- Fukuda, K.; Watanabe, M.; Asano, K.; Ouchi, K.; Takasawa, S. Isolation and genetic study of p-fluoro-dl-phenylalanine-resistant mutants overproducing β-phenethyl-alcohol in Saccharomyces cerevisiae. Curr. Genet. 1991, 20, 449–452. [Google Scholar] [CrossRef]
- Araújo, T.M.; Souza, M.T.; Diniz, R.H.S.; Yamakawa, C.K.; Soares, L.B.; Lenczak, J.L.; De Castro Oliveira, J.V.; Goldman, G.H.; Barbosa, E.A.; Campos, A.C.S.; et al. Cachaça yeast strains: Alternative starters to produce beer and bioethanol. Antonie van Leeuwenhoek 2018, 111, 1749–1766. [Google Scholar] [CrossRef]
- Tokpohozin, S.E.; Fischer, S.; Becker, T. Selection of a new Saccharomyces yeast to enhance relevant sorghum beer aroma components, higher alcohols and esters. Food Microbiol. 2019, 83, 181–186. [Google Scholar] [CrossRef]
- Rossi, S.; Turchetti, B.; Sileoni, V.; Marconi, O.; Perretti, G. Evaluation of Saccharomyces cerevisiae strains isolated from non-brewing environments in beer production. J. Inst. Brew. 2018, 124, 381–388. [Google Scholar] [CrossRef]
- Canonico, L.; Comitini, F.; Ciani, M. Dominance and influence of selected Saccharomyces cerevisiae strains on the analytical profile of craft beer refermentation. J. Inst. Brew. 2014, 120, 262–267. [Google Scholar] [CrossRef]
- Cubillos, F.A.; Gibson, B.; Grijalva-Vallejos, N.; Krogerus, K.; Nikulin, J. Bioprospecting for brewers: Exploiting natural diversity for naturally diverse beers. Yeast 2019, 36, 383–398. [Google Scholar] [CrossRef]
- Dlusskaya, E.; Jänsch, A.; Schwab, C.; Gänzle, M.G. Microbial and chemical analysis of a kvass fermentation. Eur. Food Res. Technol. 2008, 227, 261–266. [Google Scholar] [CrossRef]
- Ekberg, J.; Gibson, B.; Joensuu, J.J.; Krogerus, K.; Magalhães, F.; Mikkelson, A.; Seppänen-Laakso, T.; Wilpola, A. Physicochemical characterization of sahti, an ‘ancient’ beer style indigenous to Finland. J. Inst. Brew. 2015, 121, 464–473. [Google Scholar]
- Londesborough, J. Fermentation of maltortiose by brewer’s and baker’s yeast. Biotechnol. Lett. 2001, 23, 1995–2000. [Google Scholar] [CrossRef]
- Marongiu, A.; Zara, G.; Legras, J.-L.; Del Caro, A.; Mascia, I.; Fadda, C.; Budroni, M. Novel starters for old processes: Use of Saccharomyces cerevisiae strains isolated from artisanal sourdough for craft beer production at a brewery scale. J. Ind. Microbiol. Biotechnol. 2015, 42, 85–92. [Google Scholar] [CrossRef]
- Mascia, I.; Fadda, C.; Kerabín, M.; Dostálek, P.; Del Caro, A. Aging of craft durum wheat beer fermented with sourdough yeasts. LWT Food Sci. Technol. 2016, 65, 487–494. [Google Scholar] [CrossRef]
- Ripari, V.; Tomassetti, M.; Cecchi, T.; Berardi, E. Recipe, volatiles profile, sensory analysis, physico-chemical and microbial characterization of acidic beers from both sourdough yeasts and lactic acid bacteria. Eur. Food Res. Technol. 2018, 244, 2027–2040. [Google Scholar] [CrossRef]
- Vidgren, V.; Londesborough, J. 125th anniversary review: Yeast flocculation and sedimentation in brewing. J. Inst. Brew. 2011, 117, 475–487. [Google Scholar] [CrossRef]
- Steensels, J.; Daenen, L.; Malcorps, P.; Derdelinckx, G.; Verachtert, H.; Verstrepen, K.J. Brettanomyces yeasts—From spoilage organisms to valuable contributors to industrial fermentations. Int. J. Food Microbiol. 2015, 206, 24–38. [Google Scholar] [CrossRef]
- Varela, C. The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl. Microbiol. Biotechnol. 2016, 100, 9861–9874. [Google Scholar] [CrossRef] [PubMed]
- Gamero, A.; Quintilla, R.; Groenewal, M.; Alkema, W.; Boekhout, T.; Hazelwood, L. High-throughput screening of a large collection of non-conventional yeasts reveals their potential for aroma formation in food fermentation. Food Microbiol. 2016, 60, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Van Rijswijck, I.M.H.; Wolkers-Rooijackers, J.C.M.; Abee, T.; Smid, E.J. Performance of non-conventional yeasts in co-culture with brewers’ yeast for steering ethanol and aroma production. Microbiol. Biotechnol. 2017, 10, 1591–1602. [Google Scholar] [CrossRef]
- Toh, D.W.K.; Chua, J.Y.; Liu, S.Q. Impact of simultaneous fermentation with Saccharomyces cerevisiae and Torulaspora delbrueckii on volatile and non-volatile constituents in beer. LWT-Food Sci. Technol. 2018, 91, 26–33. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled mixed culture fermentation: A new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F. Non-Saccharomyces wine yeasts have a promising role in biotechnological approaches to winemaking. Ann. Microbiol. 2010, 61, 25–32. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef]
- Holt, S.; Mukherjee, V.; Lievens, B.; Verstrepen, K.J.; Thevelein, J.M. Bioflavoring by non-conventional yeasts in sequential beer fermentations. Food Microbiol. 2017, 72, 55–66. [Google Scholar] [CrossRef]
- Canonico, L.; Galli, E.; Ciani, E.; Comitini, F.; Ciani, M. Exploitation of three non-conventional yeast species in the brewing process. Microorganisms 2019, 7, 11. [Google Scholar] [CrossRef]
- De Francesco, G.; Turchetti, B.; Sileoni, V.; Marconi, O.; Perretti, G. Screening of new strains of Saccharomycodes ludwigii and Zygosaccharomyces rouxii to produce low-alcohol beer. J. Inst. Brew. 2015, 121, 113–121. [Google Scholar]
- Meier-Dörnberg, T.; Hutzler, M.; Jacob, F.; Schneiderbanger, H. Geschmacklich ansprechend. Brauindustrie 2015, 7, 12–15. [Google Scholar]
- Saerens, S.; Swiegers, J.H. Production of Low-Alcohol or Alcohol-Free Beer with Pichia kluyveri Yeast Strains. Patent No. WO2014135673A2, 12 September 2014. [Google Scholar]
- Petruzzi, L.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Brewer’s yeast in controlled and uncontrolled fermentations, with a focus on novel, nonconventional, and superior strains. Food Rev. Int. 2016, 32, 341–363. [Google Scholar] [CrossRef]
- Haslbeck, K.; Jerebic, S.; Zarnkow, M. Characterization of the unfertilized and fertilized hop varieties progress and hallertauer tradition—Analysis of free and glycosidic-bound flavor compounds and β-glucosidase activity. Brew. Sci. 2017, 70, 148–158. [Google Scholar]
- Vervoort, Y.; Herrera-Malaver, B.; Mertens, S.; Guadalupe Medina, V.; Duitama, J.; Michiels, L.; Derdelinck, G.; Voordeckers, K.; Verstrepen, K.J. Characterization of the recombinant Brettanomyces anomalus β-glucosidase and its potential for bioflavouring. J. Appl. Microbiol. 2016, 121, 721–733. [Google Scholar] [CrossRef]
- Michel, M.; Kopecká, J.; Meier-Dörnberg, T.; Zarnkow, M.; Jacob, F.; Hutzler, M. Screening for new brewing yeasts in the non-Saccharomyces sector with Torulaspora delbrueckii as model. Yeast 2016, 33, 129–144. [Google Scholar] [CrossRef]
- Basso, R.F.; Alcarde, A.R.; Portugal, C.B. Could non-Saccharomyces yeasts contribute on innovative brewing fermentations? Food Res. Int. 2016, 86, 112–120. [Google Scholar] [CrossRef]
- Callejo, M.J.; García Navas, J.J.; Alba, R.; Escott, C.; Loira, I.; González, M.C.; Morata, A. Wort fermentation and beer conditioning with selected non-Saccharomyces yeasts in craft beers. Eur. Food Res. Technol. 2019, 245, 1229–1238. [Google Scholar] [CrossRef]
- Domizio, P.; House, J.F.; Joseph, C.M.L.; Bisson, L.F.; Bamforth, C.W. Lachancea thermotolerans as an alternative yeast for the production of beer. J. Inst. Brew. 2016, 122, 599–604. [Google Scholar]
- Ravasio, D.; Carlin, S.; Boekhout, T.; Groenewald, M.; Vrhovsek, U.; Walther, A.; Wendland, J. Adding flavor to beverages with non-conventional yeasts. Fermentation 2018, 4, 15. [Google Scholar] [CrossRef]
- Canonico, L.; Agarbati, A.; Comitini, F.; Ciani, M. Torulaspora delbrueckii in the brewing process: A new approach to enhance bioflavour and to reduce ethanol content. Food Microbiol. 2016, 56, 45–51. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Secondary Metabolites | Flavor Impact | References |
|---|---|---|---|
| 5,5,5-trifluoro dl-leucine | Increase in 3-methylbutyl acetate | banana/pear aroma | [93] |
| isoamyl monofluoroacetate | Increase in 3-methylbutyl acetate | banana/pear aroma | [94] |
| 1-farnesylpyridinium | Increase in 3-methylbutyl acetate | banana/pear aroma | [95] |
| chlorsulfuron | decrease in diacetyl | buttery aroma | [88] |
| cerulenin | Increase in ethyl caproate | apple aroma | [96] |
| fluoro-dl-phenylalanine | Increase in phenylethyl acetate | rose aroma | [97] |
| Yeast | Strain | Fermentation Conditions | Reference |
|---|---|---|---|
| Blastobotrys mokoenaii | X9113 | pure | [118] |
| Brettanomyces anomalus | X9073 | pure/sequentially inoculated with Ale 514 brewing yeast | [118] |
| Brettanomyces bruxellensis | CBS 3025, AWRI1499 | pure/sequentially inoculated with Ale 514 brewing yeast | [118] |
| Brettanomyces naardenensis | NRRL Y-5740 | pure/sequentially inoculated with Ale 514 brewing yeast | [118] |
| Candida stellata | X9023 | pure | [118] |
| Citeromyces matritensis | ST1312/081 | pure | [118] |
| Debaryomyces hansenii | x38 | pure | [118] |
| Kodamaea ohmeri | x22 | pure | [118] |
| Lachancea thermotolerans | DiSVA 322 | pure/co-culture with S. cerevisiae starter strain US-05 | [131] |
| Lachancea thermotolerans | x9005 | pure | [118] |
| Metschnikowia reukaufi | Y6.3K/FT11 B | pure | [118] |
| Pichia anomala | x9015, x10 | pure/sequentially inoculated with Ale 514 brewing yeast | [118] |
| Pichia kluyverii | x21, x36 | pure/sequentially inoculated with Ale 514 brewing yeast | [118] |
| Pichia kudriavzevii | x12, X9035 | pure/sequentially inoculated with Ale 514 brewing yeast | [118] |
| Saccharomycodes ludwigii | DBVPG 3010, DBVPG 3304, DBVPG 3398, DBVPG 3931, DBVPG 4116, DBVPG 6721 | pure | [120] |
| Starmerella bacillaris | X9029 | pure | [118] |
| Starmerella bombicola | V10.2Y A1 | pure | [118] |
| Torulaspora delbrueckii | DiSVA 254 | pure/co-culture with S. cerevisiae starter strain US-05 | [131] |
| Torulaspora delbrueckii | ST1312/167 | pure/sequentially inoculated with Ale 514 brewing yeast | [118] |
| Wickerhamomyces anomalus | DiSVA 2 | pure/co-culture with S. cerevisiae starter strain US-05 | [119] |
| Zygosaccharomyces rouxii | DBVPG 4084, DBVPG 6187, DBVPG 6424, DBVPG 6463, DBVPG 6921 | pure | [120] |
| Zygotorulaspora florentina | DiSVA 263 | pure/co-culture with S. cerevisiae starter strain US-05 | [119] |
| Zygotorulaspora florentina | X9022 | pure/sequentially inoculated with Ale 514 brewing yeast | [118] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iattici, F.; Catallo, M.; Solieri, L. Designing New Yeasts for Craft Brewing: When Natural Biodiversity Meets Biotechnology. Beverages 2020, 6, 3. https://doi.org/10.3390/beverages6010003
Iattici F, Catallo M, Solieri L. Designing New Yeasts for Craft Brewing: When Natural Biodiversity Meets Biotechnology. Beverages. 2020; 6(1):3. https://doi.org/10.3390/beverages6010003
Chicago/Turabian StyleIattici, Fabrizio, Martina Catallo, and Lisa Solieri. 2020. "Designing New Yeasts for Craft Brewing: When Natural Biodiversity Meets Biotechnology" Beverages 6, no. 1: 3. https://doi.org/10.3390/beverages6010003
APA StyleIattici, F., Catallo, M., & Solieri, L. (2020). Designing New Yeasts for Craft Brewing: When Natural Biodiversity Meets Biotechnology. Beverages, 6(1), 3. https://doi.org/10.3390/beverages6010003