1. Blending the Art of Bach and the Craft of Bacchus
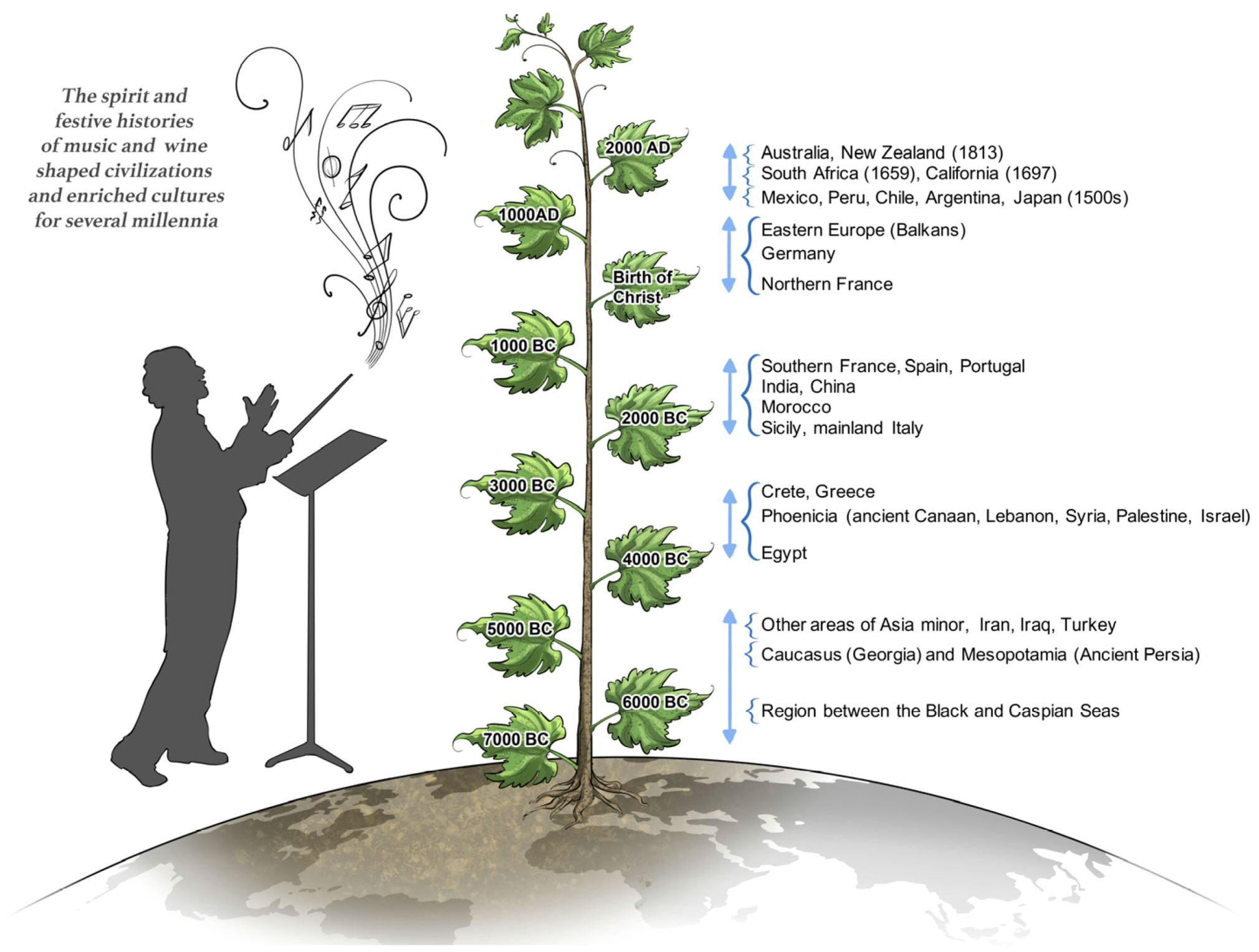
Music and wine are found in every known culture—past and present—varying markedly between places and times, and shaping civilizations in very significant ways the world over. Music might have been present in the ancestral population prior to the dispersal of humans around the world 50,000 years ago, while the history of winemaking stretches back to almost 7000 BC (
Figure 1). Over the centuries, the histories and spirit of music and wine merged to meld art and craft, and greatly enrich the way we live today.
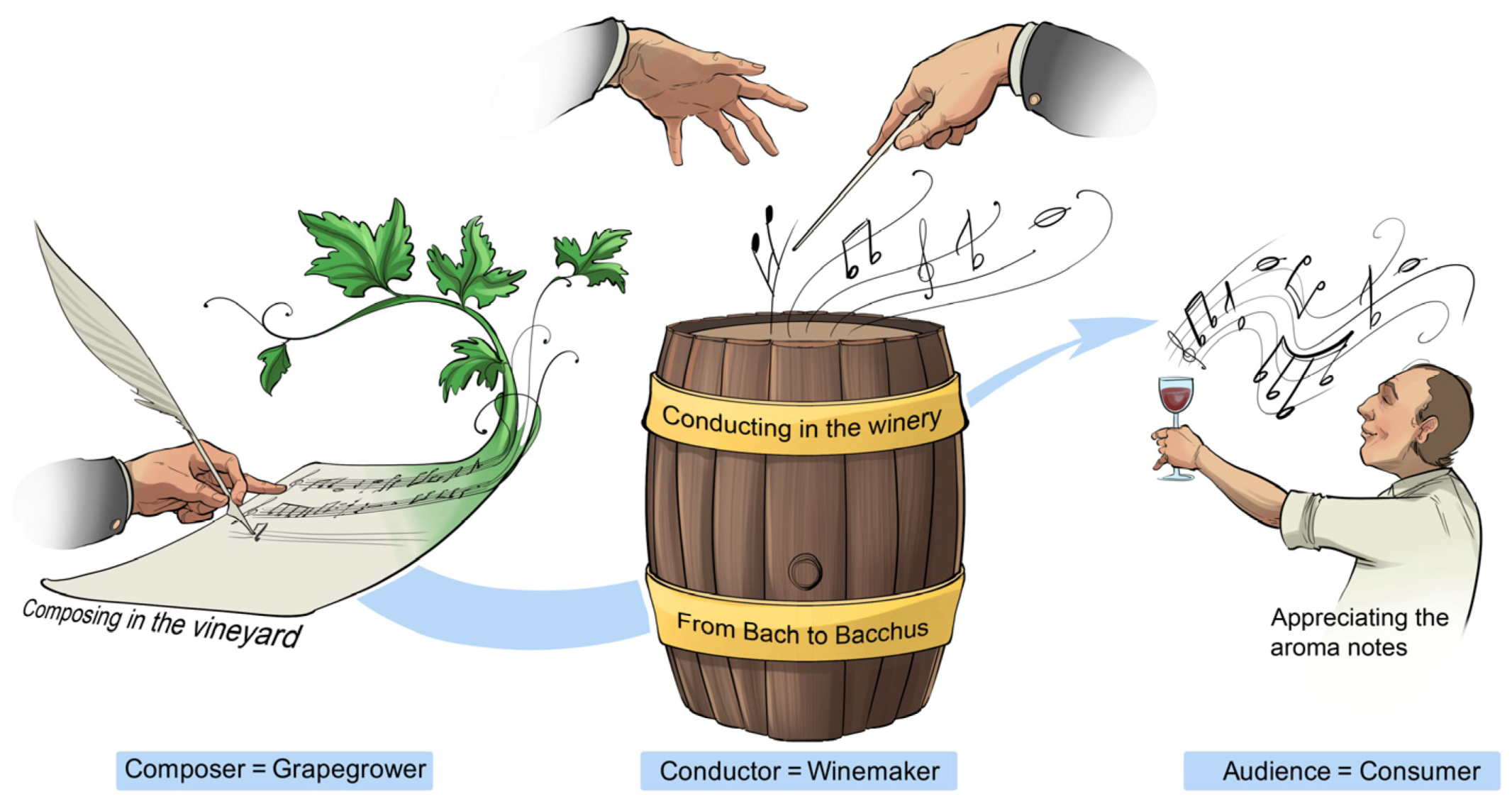
The art of composing and conducting music, and the craftsmanship of grapegrowing and winemaking, are both considered expressions of human creativity and imagination—drawing metaphoric parallels between music and wine is natural (
Figure 2). Beethoven once said “music is the wine which inspires one to new generative processes”, and went on to describe himself as “Bacchus who presses out glorious wine for humankind and makes them spiritually drunken”. We will never know whether Beethoven would have agreed with a view that, for example, Bach’s artform was more of a musical expression and application of his imagination, feelings, thoughts and ideas while Bacchus’ craft relied more heavily on a learned skill and experience in the creation of the tangible product of wine. What we do know, however, is that both the
sound of music and the
taste of wine provides us pleasure in a crossmodal sensory fashion. Scientific evidence is now emerging that shows that regular wine drinkers are able to reliably match certain wines with particular pieces of music [
2,
3,
4].
Akin to a composer who intertwines elements to create an original piece of music—a melody, underlying harmony or chord progression, and sometimes lyrics—a grapegrower composes the notes of a wine in the vineyard. By applying their craft to what nature and the land bestow on their vineyards, viticulturists nurture their vines to produce grapes as sheet music for various types and styles of wines. While honouring what the composer has written, the conductor has the liberty to creatively orchestrate the original composition to his instruments of choice and to rearrange the tempo to satiate the musical appetite of his audience. In the winery, the winemaker honours the score arranged by the grapegrower in the vineyard, and uniquely interprets grape compositions by conducting different arrangements of selected pre-fermentation practices, yeast strains, fermentation conditions, post-fermentation technologies, packaging materials, as well as marketing and retailing strategies to achieve their own vision. In smaller operational settings, the interconnected choices made at each step in the from-grape-to-glass value chain might be simpler because maestro vintners can be both the composer in the vineyard and the conductor in the winery.
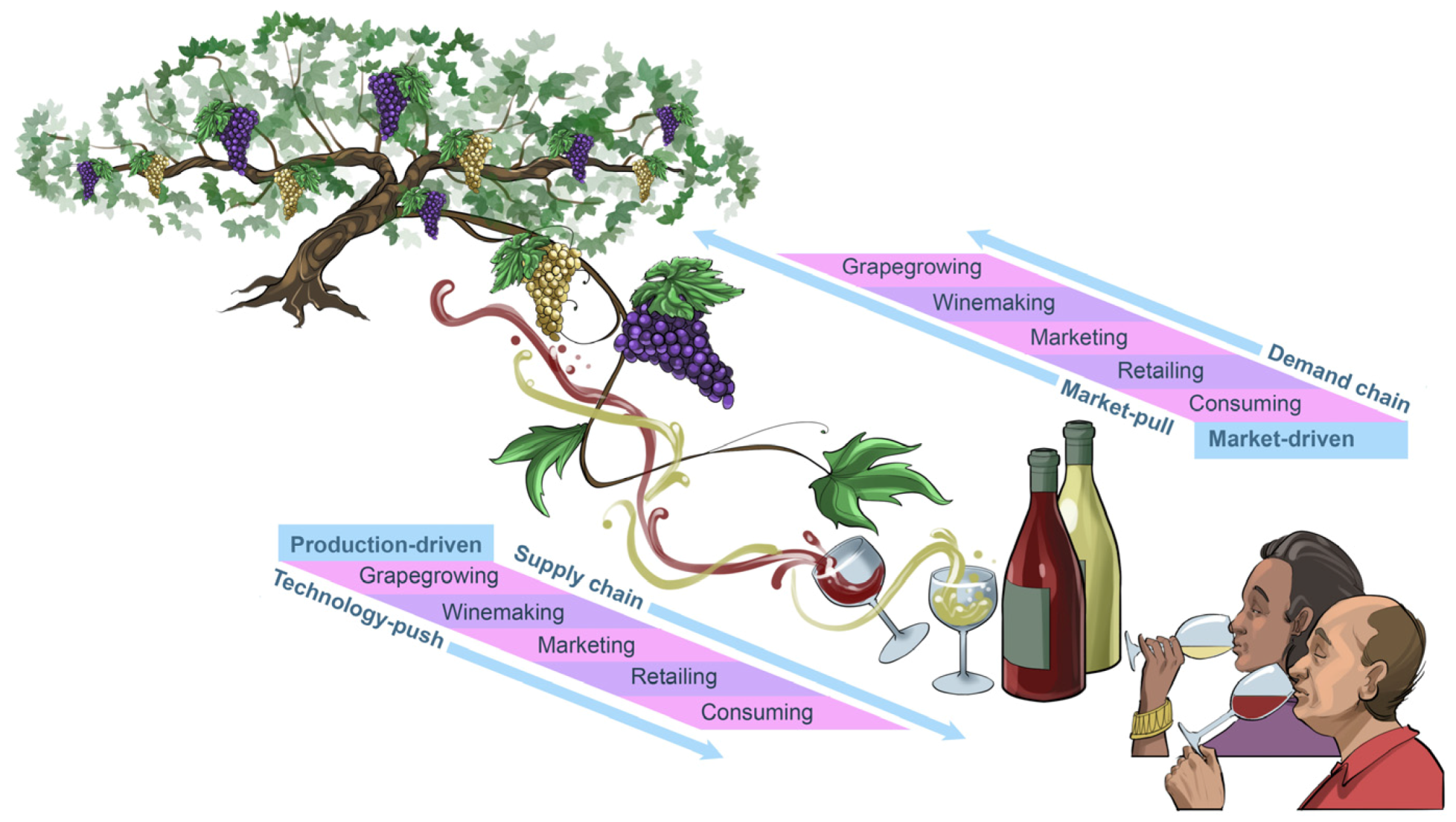
Good music and good wine please different senses in similar ways. Pulling the cork from a bottle of wine in a pleasant social setting evokes a similar experience and appreciation as attending a concert and listening to a familiar piece of music, even when there is all the uncertainty of no two performances ever being quite the same (
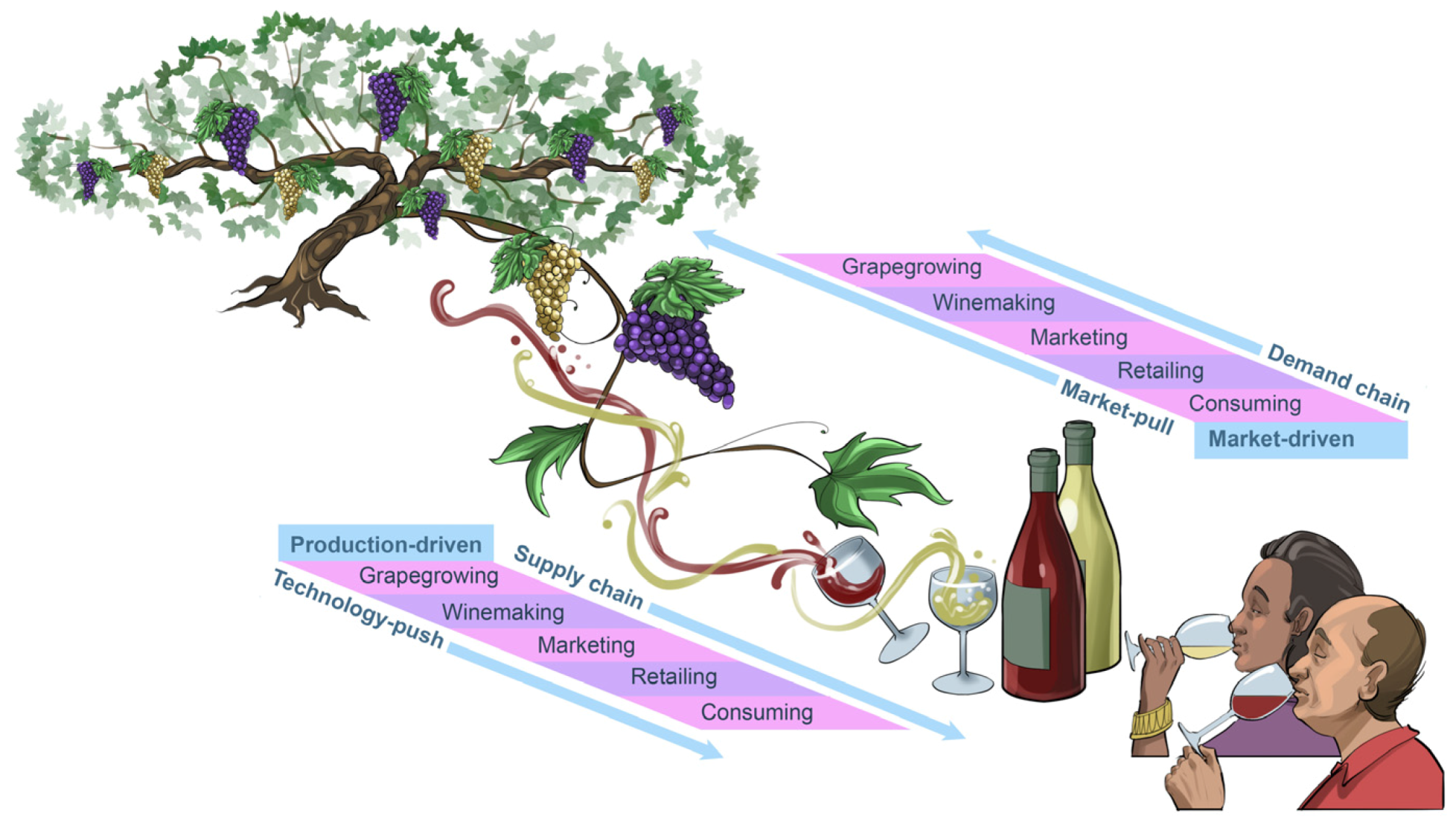
Figure 3). Our individual preferences for certain music genres and wine styles often evolve over time. Musicians and winemakers know all too well that, to keep their audiences and consumers engaged and entertained, their art and craft also need to evolve with time. Some of these changes will be due to
market-pull forces and others will be catlysed by the forces of
technology-push (
Figure 4). The result of these two sets of dynamic, trendsetting forces often convert today’s
innovation into tomorrow’s
tradition. However, sophisticated audiences and consumers usually embrace the excitement of new
trends without losing their appreciation for the charm of the timeless
classics.
Great musicians and vintners are able to accurately interpret the dynamics of these interconnected factors in a timely fashion, and to correctly anticipate what the future preferences of their target audiences and consumer markets might be. With wine, accurate market anticipation and fleet-footed agility require masterful orchestration of a harmonious blend of an artist’s intuition, a craftsman’s skill and a scientist’s logic—and that makes for crowd-thrilling entertainment, standing ovations and encore sales in the global marketplace.
In this context, this article seeks to shine the spotlight on the fermentation performance of classic and trendy yeasts in the winery, and to trumpet the sound of future music from the research laboratory.
2. Translating Grape Compositions into Enchanting and Distinctive Wine Genres
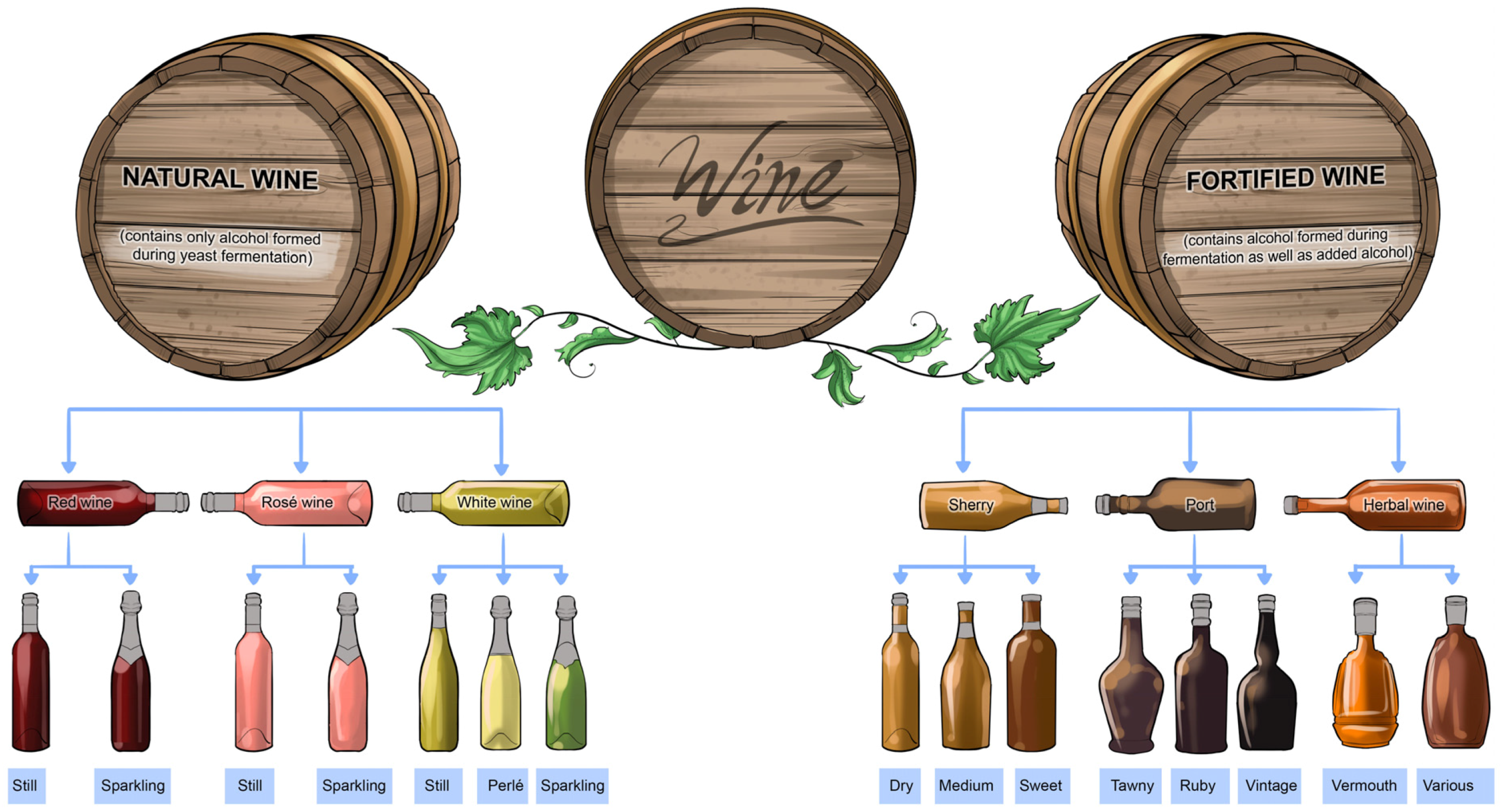
Listening to a philharmonic orchestra’s performance, an audience values the quality and harmony of a multitude of pleasing sounds, as well as feeling inspired by the whole experience. This is not dissimilar to the expectations and preferences of wine drinkers who often use the terms
quality and
value when they refer to a particular wine (
Figure 5). To the consumer who compares different wines for purchase, the term
quality relates to the ‘intrinsic’ quality of the wine, meaning how the wine gratifies on appearance, the nose and the palate, as well as the perceived value [
7]. When consumers use the term
value, they usually refer to both the
intrinsic value and the
image of a wine in relation to the price. The image of a product depends on how a wine is marketed, the origin and regionality of a wine, how environmentally-sound the winery’s practices are, how many medals have been awarded, how high sommeliers and other influencers rate a wine, and price. Consumers will consider a wine high in value if the product is sensorially pleasing and recognisable, and perceived as high in image at a competitive price [
7].
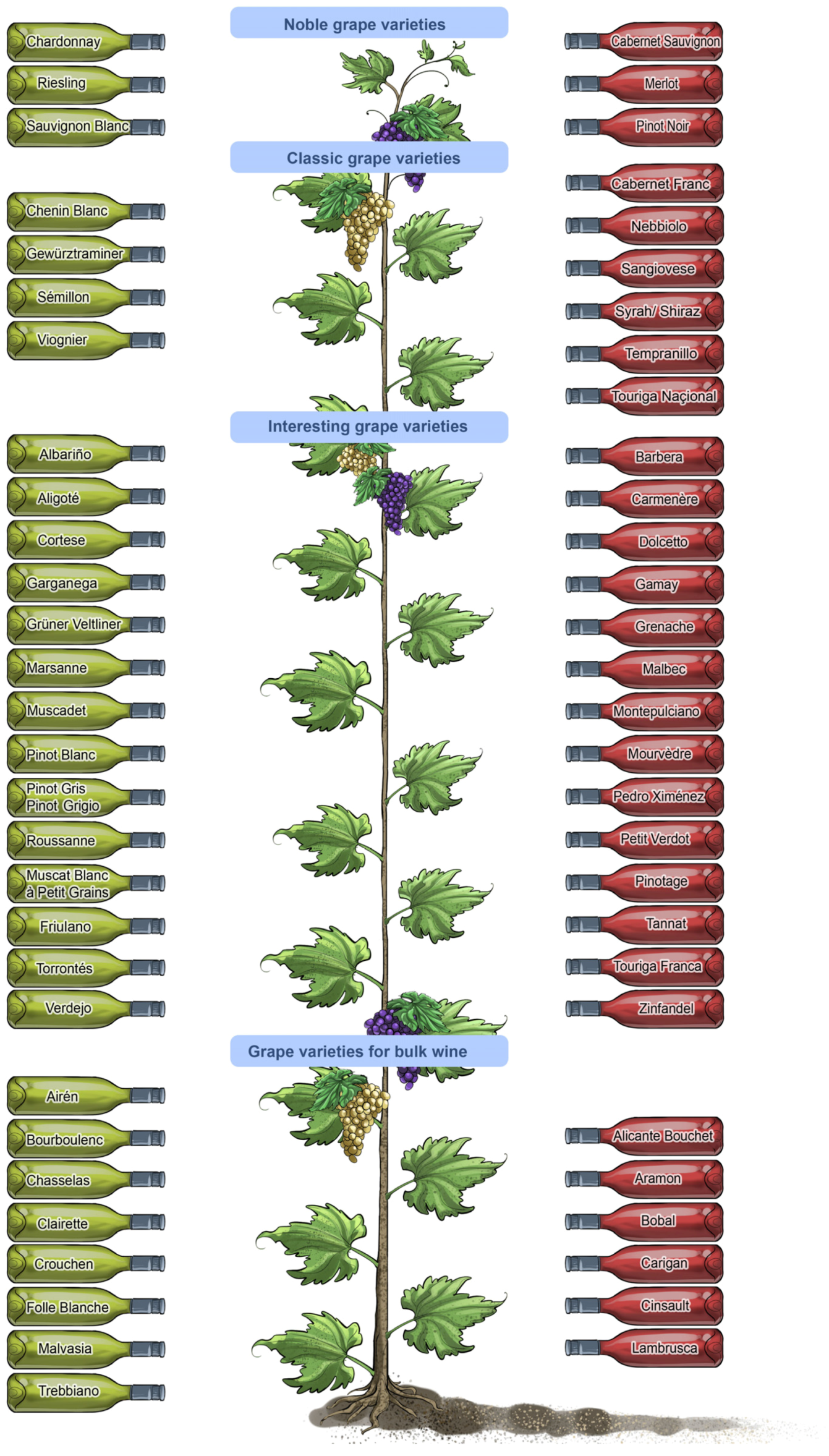
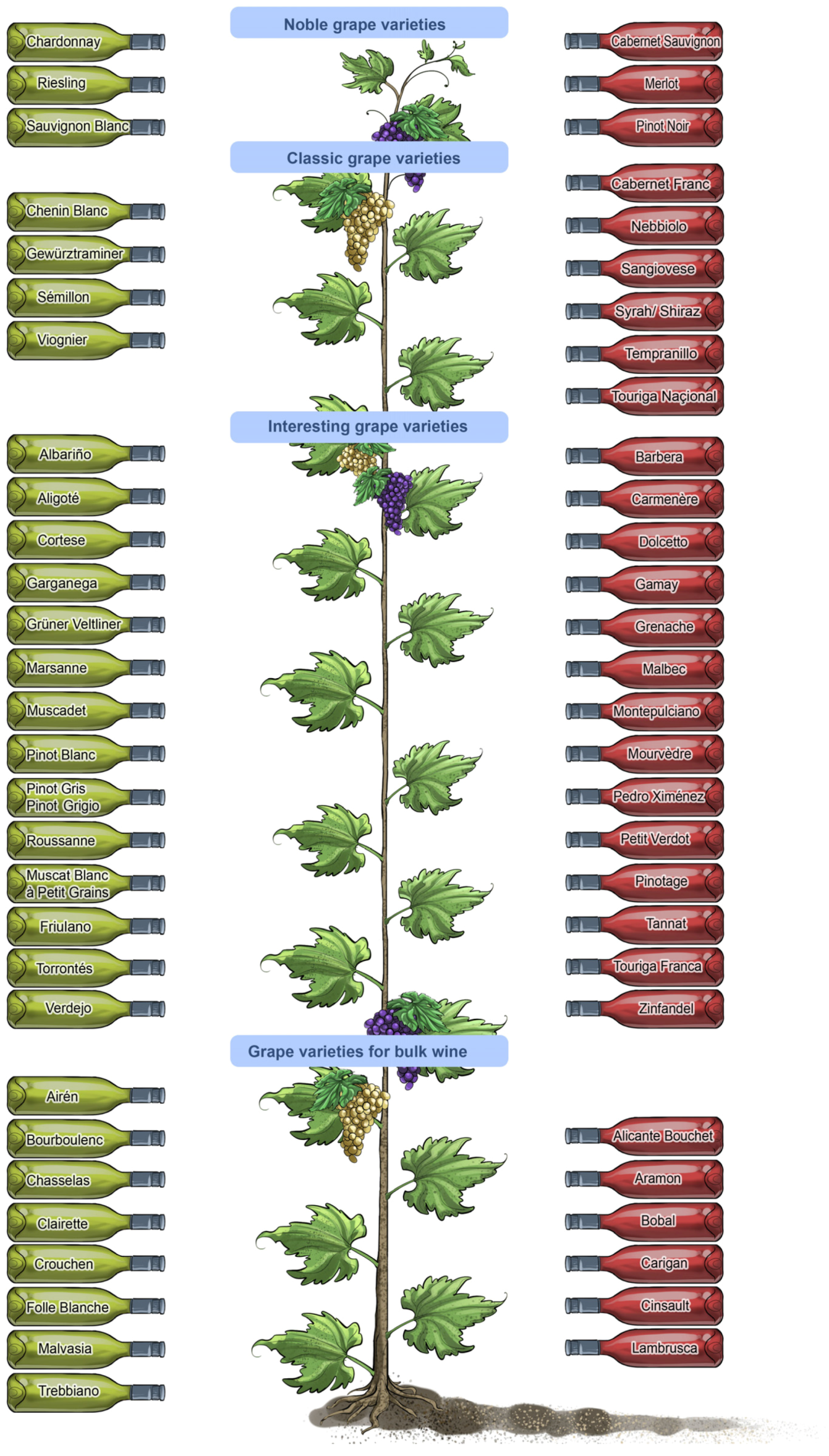
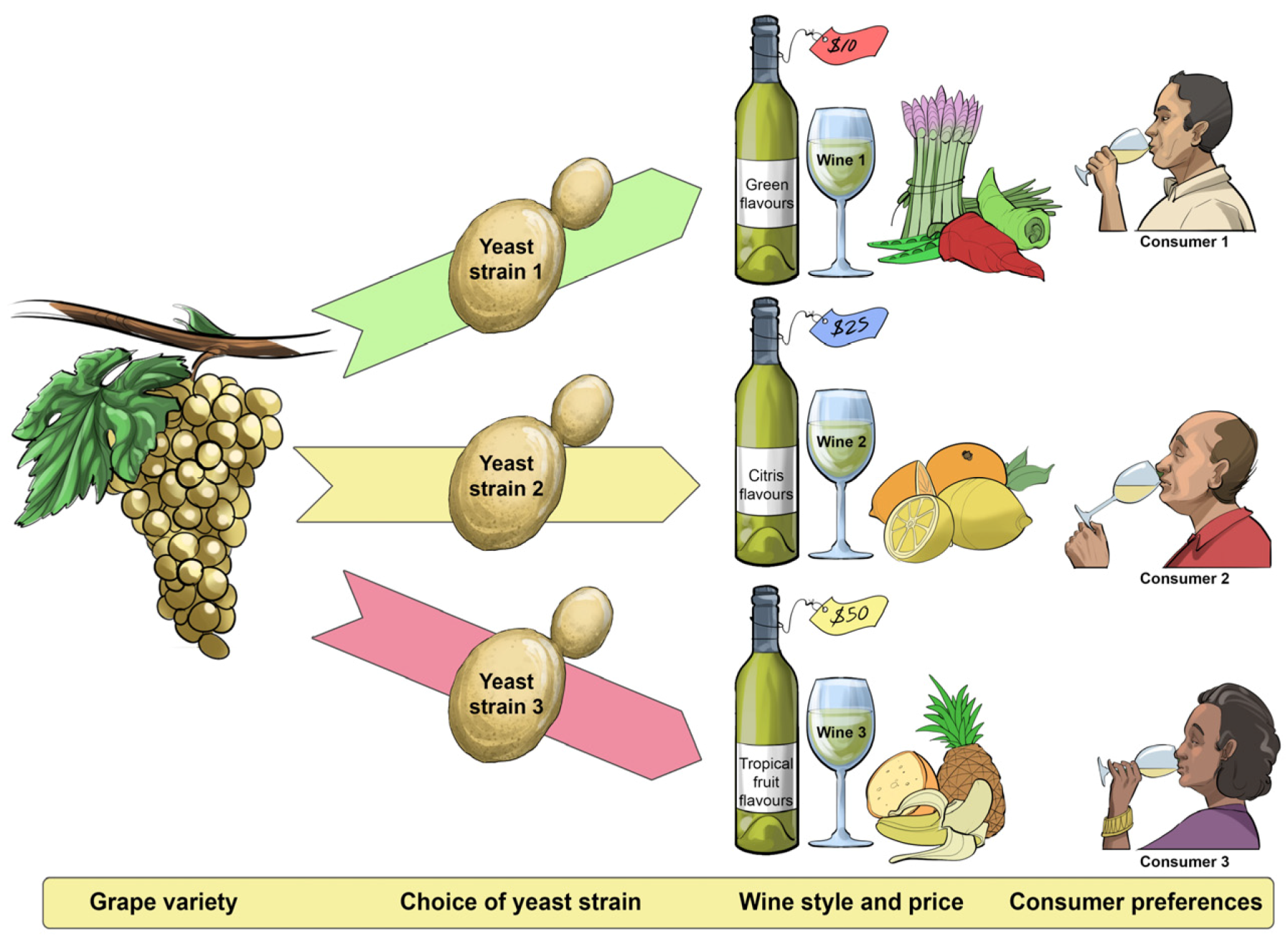
Similar to music enthusiasts—young and old, male and female—who vary widely in their preferences for different music genres, wine consumers’ senses are not uniformly sensitive or attuned to the subtle assortment of changing sensations evoked by different wine types and styles (
Figure 6). Some of the diversity in sensory perception and preferences for different wine products among individuals and populations is cultural—some genetic and some learned—and some preferences are also influenced by gender, age and fashion [
5].
For the past 35 years, the global wine industry has been producing approximately 15% in excess of annual consumption and struggles to hold on to its 11% share of the global market for alcoholic drinks [
8]. By volume beer occupies about 80% of the alcoholic drinks market with spirits making up the other 9%. The production value of the ~32 billion bottles (or equivalents) of wine sold every year is roughly US$100 billion, representing about a quarter of the total value of the global market for alcoholic drinks [
8]. The widening gap between wine production and consumption poses a real threat to the profitability of many wine businesses in all wine producing countries. Unsurprisingly, this market pressure drives vintners to strive for a deeper understanding of, and insights into, the expectations and preferences of individuals and populations in their targeted markets so that they can deliver desirable and distinctive wines accordingly.
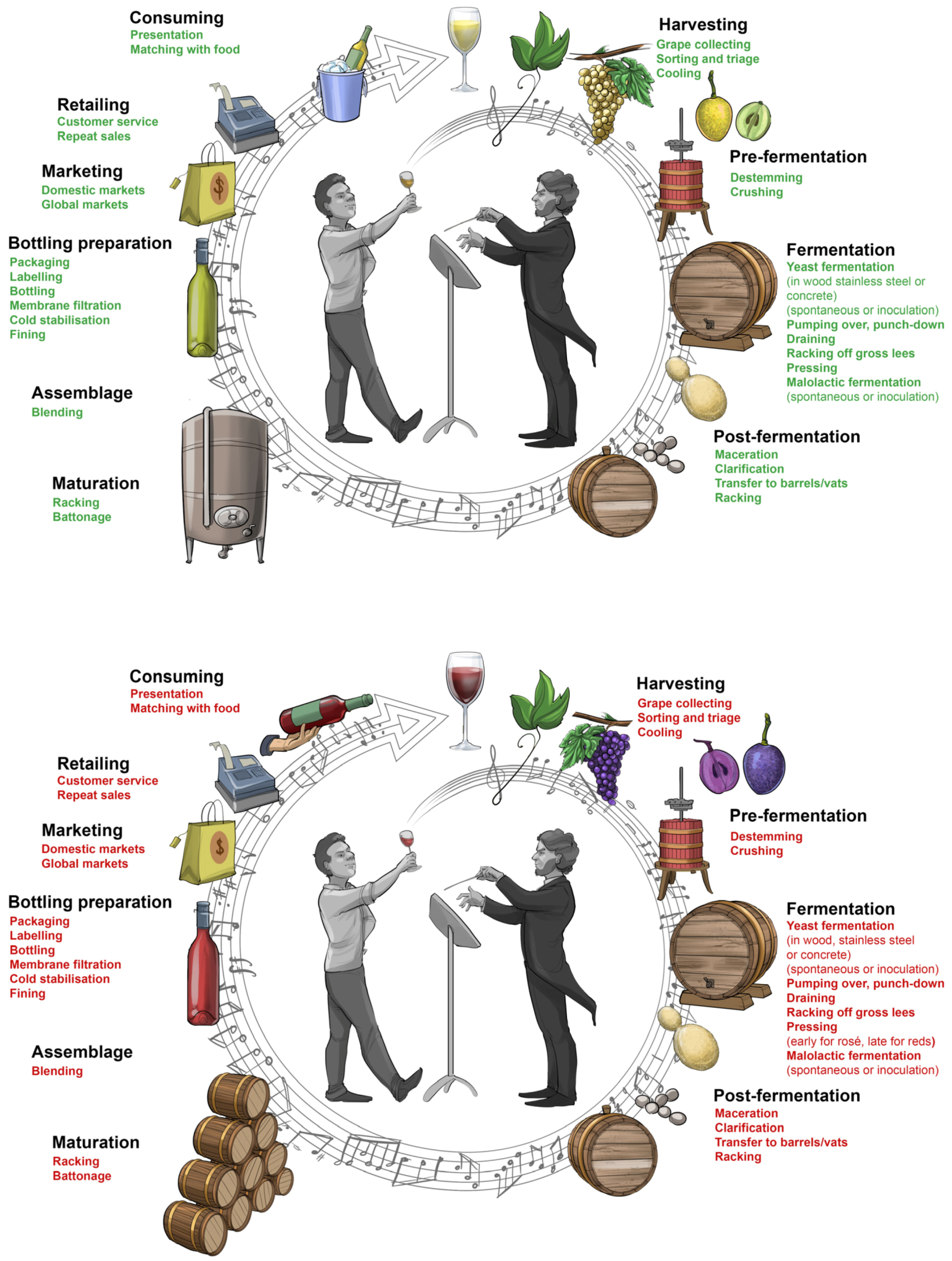
It is impossible to orchestrate appealing music from a badly composed piece—similarly, it is impossible to make good wine from badly grown grapes. So, the process of winemaking really starts in the vineyard with optimally ripened grapes on the vine. The grapes, as they are harvested, contain the potential of the wine but the winemaker has a critical role in realising that potential in full.
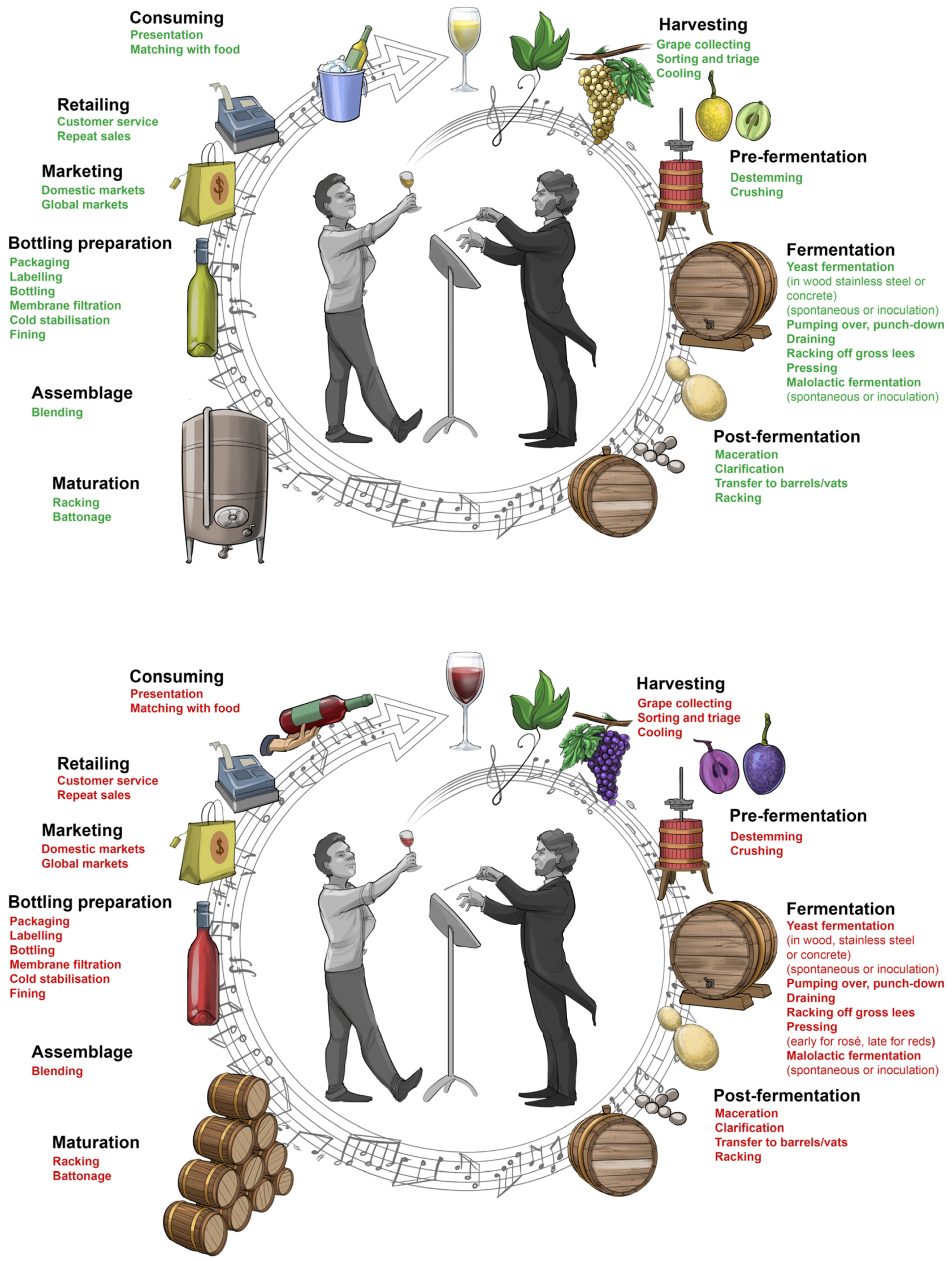
At each step of the winemaking process, the winemaker is required to make well-informed, insightful decisions as to what technique, technology and practice will best guide the process to fulfill the potential of the grapes and achieve a predetermined outcome. There are five basic stages: (i) harvesting, crushing and pressing; (ii) fermentation; (iii) clarification, fining and stabilisation; (iv) maturation and ageing; and (v) bottling, labelling and packaging (
Figure 7). However, there are variations and deviations along the way that enable winemakers to put their own individual stamp on the production chain, thereby making their wine unique (for further reading see [
10,
11,
12]).
Grape harvesting. Determining when to harvest the grapes is the first step and decision-making point in the winemaking process [
12]. This decision is heavily influenced by the weather and it requires a touch of science (i.e., chemical analysis) and old-fashioned tasting of grapes. The minute grapes are picked by hand or mechanically harvested, will largely determine the acidity, sweetness, alcoholic strength, tannin texture, astringency and flavour of the wine [
10]. Once the grapes are delivered at the winery, they are sorted into bunches, and rotten or under-ripe grapes are removed.
Grape crushing and wine pressing. The next steps involve destemming, crushing and pressing of the sorted grapes [
12]. The traditional stomping of grapes by workers’ feet has mostly been replaced by mechanical crushing and pressing of grapes into ‘must’. White wine production requires grape skins, seeds, and stems to be removed immediately after crushing to prevent unwanted colour and tannins from leaching into the wine. Whereas the production of red wine requires that, following crushing, the grape skins remain in the juice during the vinification (fermentation) process. During maceration—the time period during which the skins and grape juice are allowed to interact—polyphenolic material consisting of anthocyanins (water-soluble pigments), tannins and flavour-active compounds are leached from the grape skins, seeds and stems into the must, giving red wine its colour, structure and aroma [
11]. With longer maceration, the onset of spontaneous fermentation is delayed by cooling the must—a process called ‘cold soak’ or ‘cold maceration’. Short periods (usually no more than a couple of days) of maceration and cold-soaking are also done for a limited range of white wine styles, but these practices are mostly applied in red wine making. Depending on the grape variety and intended style of red wine to be made, the length of maceration can be extended for a period of several weeks and is one of the most influential controls available to the winemaker to shape the style of the end-product. In addition, the way the wine is pressed, can also significantly influence the style and quality of the end-product. The practice of wine pressing has evolved from the traditional foot-stomping of grapes to basket-presses and to modern-day bladder-presses (batch or continuous mechanical presses) by which an airbag fills up with air and gently squeezes the grapes against the inner wall of a perforated cylinder. Generally, the wine will first be ‘racked’ so that clear ‘free-run wine’ separates from the solids before the pomace (grape skins, pulp, seeds and stems) is pressed. ‘Press wine’ is usually inferior in quality to ‘free-run wine’ and much more astringent [
11]. At the crushing or pressing stage, the winemaker might make a number of additions to the juice (for white wine) or to the must (for red wine) to adjust its composition prior to fermentation. These additions might include acid (‘acidification’ in hot climate regions), sugar (‘chaptalisation’ in cold climate regions), sulfur dioxide (as an anti-microbial and anti-oxidisation agent) and ascorbic acid (as an anti-oxidant in white winemaking).
Alcoholic and malolactic fermentation. The alcoholic fermentation is conducted by ambient yeast (‘spontaneous’ fermentation) or added yeast (‘inoculated’ fermentation). During this process, the main grape sugars, glucose and fructose, are assimilated and converted to ethyl alcohol (ethanol) and carbon dioxide (CO
2) by the anaerobic metabolism (glycolysis) of yeast cells. One glucose molecule generates two ethanol and two carbon dioxide molecules, i.e., C
6H
12O
6→2C
2H
5OH + 2CO
2. The yeast-driven alcoholic fermentation is similar for white and red wine except for the fact that fermentation of red wine occurs in the presence of grape solids and at a slightly higher temperature (18–30 °C). The temperature maintained in the fermentation mass is the main factor that determines the duration of fermentation—even more so than the initial sugar concentration, yeast type, aeration of the must and the quantity of micro-nutrients in the juice or must [
10]. Typically, red wine fermentations are complete within a week whereas white wines, which are often fermented at much lower temperatures (12–18 °C), might take several weeks to ferment to dryness (that is, when all the sugars have been consumed). Depending on what style of wine a winemaker is striving to achieve, the alcoholic fermentation is usually undertaken in oak barrels, large oak vats, open-top concrete vessels and stainless steel tanks [
8]. Irrespective of the type of fermentation vessel, the large amount of grape skins, stems and seeds in red must, interacting with effervescing carbon dioxide from the fermentation, produces a solid mass (known as the ‘cap’) which floats to the top. To maintain contact between the red grape solids and the juice, the ‘cap’ is regularly mixed through the juice. ‘Cap management’ can be done manually (‘punch-down’), mechanically (using rotary tanks) or by pump-over methods (pumping the liquid from the bottom of the vessel over the ‘cap’).
Malolactic bacteria drive the secondary fermentation known as malolactic fermentation. During this post-alcoholic fermentation process, malic acid is converted into lactic acid (which has lower acidity) and carbon dioxide: COOH-CHOH-CH2-COOH→COOH-CHOH-CH3 + CO2. Malolactic fermentation is uniformly done for all red wine except for the lightest-bodied red wine. By contrast, malolactic fermentation is uncommon in white wine making, except for the production of certain styles of Chardonnay and sparkling wines. In addition to its role in the deacidification of wine, malolactic fermentation can also add to the aroma, flavour and complexity of wine, as well as rendering the wine microbiologically stable (lowering the risk of gas production in bottled wine).
Wine clarification and stabilisation. Once the fermentation stage is complete, clarification begins by removing suspended and insoluble material (referred to as ‘lees’) from young wines [
10]. The ‘lees’ include excess tartrates, pectins and gums, some proteins and small numbers of microbial cells (including dead yeast cells). Clarification and stabilisation of wine can be achieved by simply holding the wine in storage until the larger particles settle (a process known as ‘settling’ or ‘débourbage’). After the initial ‘settling’, the fine ‘lees’ remaining in the vessel of the unfinished wine can be gently stirred (a process known as ‘batonnage’) before the final ‘débourbage’, followed by siphoning or ‘racking’ of the clear upper layer from the compact layer of solids at the bottom of the vessel [
8]. This process can be accelerated by physical means, such as cold filtration, centrifugation and flotation, or by the much cheaper chemical process known as ‘fining’. Refrigeration and cold filtration will reduce the concentration of tartrates in young wines supersaturated in tartrates to levels below concentrations that would subsequently form crystals in the bottle [
11]. The purpose of adding fining agent preparations (e.g., clarifying enzyme preparations and the volcanic clay bentonite) to wine is to soften or reduce its astringency and/or bitterness; remove proteins capable of haze formation; or reduce colour by the adsorption and precipitation of polymeric phenols and tannins. The fining agent reacts with wine components either chemically or physically, to form a new complex that can separate from the wine. The degree to how thoroughly the wine is clarified and stabilised will depend on the style target of the winemaker: ‘everyday’ wines are usually more thoroughly clarified and stabilised whereas there is a market for ‘unfiltered’ wines—some consumers consider these to be of better quality as they accept that a sediment can slowly form in bottles and they believe that the less stable constituents contribute positively to the ageing process [
11].
Wine maturation and ageing. The period of time during which wine is handled and stored in the winery is commonly referred to as ‘maturation’ (sometimes in contact with wood, e.g., in oak barrels for wooded wine styles, and sometimes in stainless steel tanks for zesty white wines), while the period of time that wine is stored in the bottle is known as ‘ageing’ [
8,
11]. Before blending and bottling, most wines are usually stored for several months in tanks or wooden cooperage. During oak barrel maturation (which is more commonly used for red wine than white wine), wine undergoes a series of complex transformations, including controlled oxidisation which can result in smoother and rounder wines because of the polymerisation of tannins. Oak-induced transformations can significantly change the aroma, composition and quality of the end-product by imparting a range of flavours including vanilla, oakiness and toasty smokiness [
7]. By contrast, ‘ageing’ of bottled wine only involves a subtle evolution of the wine. Contrary to popular opinion, age in a wine is not necessarily a virtue—only a relatively small subset of wine types benefit from extended bottle ageing [
11]. The majority of wines produced today are meant to be drunk within a couple of years of bottling, whether sealed under cork closures, screw caps or glass stoppers.
Growing the grapes, making the wine, packaging and labelling of the end-product are only half of the story—there is still many a slip between the cup and the lip. If the marketing, retailing or selling of wine is not done well, that is potentially akin to being on centre stage with no audience to applaud the performance. Moreover, experienced vintners and marketeers of wine also know that even with perfectly executed well-considered strategies to promote and sell wine, things can still be turned upside down by unexpected and sudden changes in the marketplace. For example, on 17 November 1991, when American television host, Morley Safer, of the popular news program, Sixty Minutes, held up a glass of red wine, looked into the camera and declared that “the answer to France’s low rate of heart disease [despite their unusually high consumption of cheese and high-fat, high-cholesterol meats] may lie in this inviting glass” the so-called French Paradox was born. Within one year, consumption of red wine increased more than 40% in the US alone. Another example is the 2004 film titled Sideways. When the main character, Miles (played by Paul Giamatti) was asked by an attractive waitress, Maya (played by Virginia Madsen) “Why are you so into Pinot?”, his 60-s response about how Pinot Noir vines only grow in “specific tucked-away corners of the world” and how dependent they are on the patient nurturing of a select group of growers to fully express the grapes’ flavours, which are “the most haunting and brilliant and thrilling and subtle and ancient on this planet”, American wine consumers fell in love with this grape cultivar. Before this movie, Pinot Noir was considered a hard-to-grow grape and a tough-to-sell wine. The Sideways Effect had an instantaneous impact on the sales of Pinot Noir. This resulted in a massive and rapid expansion of Pinot Noir plantings, and a corresponding decline in Merlot sales and value also as a result of this character who disliked Merlot wine.
The unforeseen impact of these two examples, the French Paradox and the Sideways Effect, provides valuable lessons to vintners and marketeers alike—always expect the unexpected in the marketplace and be prepared to face the music of shifting consumer preferences.
3. Playing Microbiological Musical Chairs in the Fermentation Vat
The recruitment of an optimal arrangement of talented musicians playing different instruments in an orchestra is a complex task, which demands a great deal of skill. The fermentation of grape must into wine is an equally complex ecological and biochemical process involving the sequential development of microbial species, as affected by prevailing conditions in vineyards and wineries [
13,
14,
15]. Grapes and grape musts naturally contain a mixture of fungi, yeasts, bacteria, mycoviruses and bacteriophages (
Figure 8). Grape must is relatively complete in nutrient content; however, it can support the growth of only a limited number of microbial species. The type and numerical presence of indigenous microbes depend on factors such as the method of grape harvest (hand-picked or mechanical), grape temperature, transport from vineyard to winery (distance/time, initial grape temperature, air temperature, sulfite addition), condition of grapes (time, temperature, sulfite addition) and must pre-treatment (cellar hygiene, aeration, enzyme treatment, sulfite addition, clarification method, temperature, inoculation with yeast starter cultures) [
9].
Depending on the abundance and robustness of the microbial species in grape must, these organisms might or might not survive and/or play an active role in fermentation under the particular oenological conditions applied by the winemaker. Similar to a conductor choosing different instrumentalists required to perform different roles in the orchestra, the winemaker chooses fermentation conditions and practices that determine which yeasts and bacteria will be allowed to help shape the wine for a targetted style. In essence, the objective of a winemaker is to apply [in addition to the natural selective pressures that exist in grape must (e.g., osmotic pressure exerted by high sugar concentration)] fermentation conditions (e.g., low pH) and practices (e.g., sulfite additions) that would eliminate spoilage bacteria (e.g., acetic acid bacteria and certain spoilage species of lactic acid bacteria) and yeasts (e.g., Brettanomyces, also known as Dekkera).
The selectivity of fermenting must is further bolstered once anaerobic conditions are established in the vat or tank; certain nutrients become depleted and the increasing concentration of ethanol starts to eliminate alcohol-sensitive microbial species. Such conditions invariably favour yeast species with the most efficient fermentative metabolism that are able to withstand low-pH, high-sulfite and high-ethanol environments.
With its Crabtree-positive carbon metabolism,
Saccharomyces cerevisiae is the yeast best equipped to take advantage of these restrictive winemaking conditions—it is able to generate energy under fermentative conditions and limit the growth of competing microorganisms by producing inhibiting metabolites such as ethanol and carbon dioxide. The prevalence of
S. cerevisiae (indigenous or inoculated) in wine ferments is therefore expected and desired [
13]. Unfortunately, an undesirable yeast,
Brettanomyces bruxellensis, can also survive the harsh conditions prevailing during wine fermentation as well as during wine maturation, particularly in oak barrels for extended periods [
16]. If present, this spoilage yeast converts hydroxycinnamic acids into odourous ethylphenols, producing ‘metallic’, ‘medicinal’ and ‘barnyard’ aromas in wine.
The most practical way for winemakers to gain the upper hand against
Brettanomyces contamination is to: employ sound cellar hygiene practices; inoculate their grape musts with fast-fermenting
S. cerevisiae strains; and maintain an optimal concentration of free sulfite in wine that would, on one hand, eliminate
Brettanomyces contaminants while on the other, allow
S. cerevisiae to rapidly complete the alcoholic fermentation [
16]. And, if malolactic fermentation is desired, this would also permit
Oenococcus oeni to convert malic acid into lactic acid. Inoculating grape must with a robust
S. cerevisiae strain and applying an optimal combination of low pH and a high concentration of free sulfite (SO
2) would not only restrict
Brettanomyces contaminants but would also eliminate undesirable acetic and lactic bacteria. Such winemaking practices would ensure rapid, reliable and trouble-free (non-‘sluggish’ and non-‘stuck’) fermentations as well as consistent, predictable and desirable wine flavours and end-product quality (e.g., no volatile acidity or presence of other unwanted metabolites).
The only down-side of the widely-used practice of inoculating grape must with a strong-fermenting
S. cerevisiae strain is that desirable non-
Saccharomyces yeasts originating from the grapes have limited opportunity to positively contribute to the complexity of wine [
9]. Some winemakers prefer to conduct spontaneous fermentations (comprising mixed and sequential dominance of non-
Saccharomyces and
Saccharomyces yeasts). They are prepared to take the risk of contamination of their grape must by spoilage yeasts because they consider indigenous non-
Saccharomyces yeasts as essential to the authenticity of their wines by imparting desired and distinct superior regional characteristics [
13]. In this context, the phrase ‘non-
Saccharomyces yeasts’ has emerged as a loose colloquial term now widely used in the wine industry for all yeast species found in wine other than
S. cerevisiae, with the proviso that this only includes yeasts with a positive role in the production of wine.
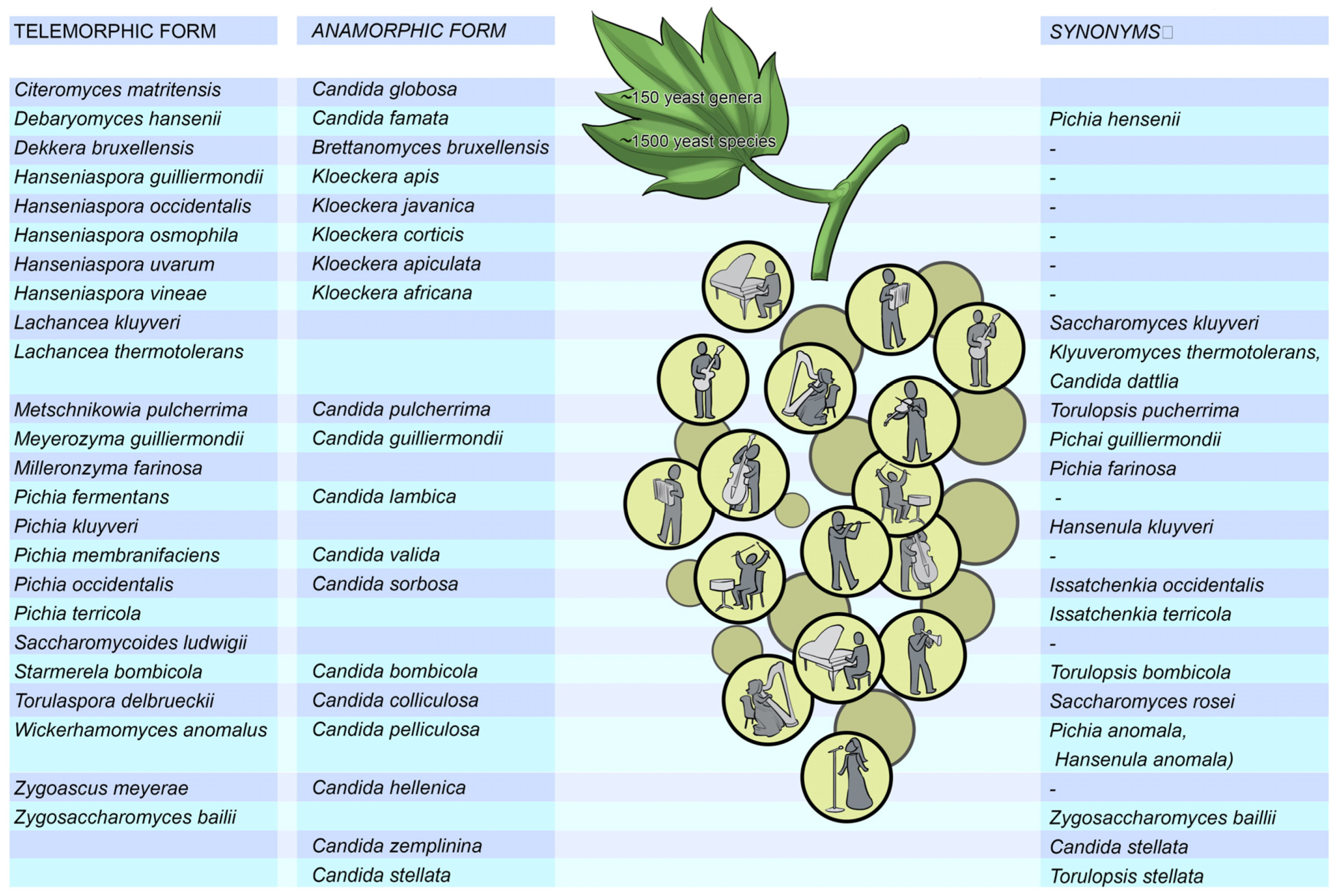
During the initial phases of fermentation, non-
Saccharomyces yeasts are both present and active in all ferments whether inoculated or not (
Figure 9). These non-
Saccharomyces yeasts can be classed into three groups: (i) yeasts that are mostly aerobic, such as species of
Candida,
Cryptococcus,
Debaryomyces,
Pichia and
Rhodoturula; (ii) apiculate yeasts with low fermentative activity, such as
Hanseniaspora uvarum (
Kloeckera apiculata),
Hanseniaspora guilliermondii (
Kloeckera apis) and
Hanseniaspora occidentalis (
Kloeckera javanica); and (iii) yeasts with a fermentative metabolism, for example
Kluyveromyces marxianus (
Candida kefyr),
Metschnikowia pulcherrima (
Candida pulcherrima),
Torulaspora delbrueckii (
Candida colliculosa), and
Zygosaccharomyces bailii [
13].
During fermentation, some of these yeasts are metabolically active until an ethanol concentration of 3% to 4% is reached while others remain present for longer until
S. cerevisiae finally exert absolute dominance to successfully complete the alcoholic fermentation. Apart from their effect on the growth of bacteria and
Saccharomyces yeasts, some of the flavour-active non-
Saccharomyces yeasts can have a significant impact on the absolute and relative concentration of aroma compounds and the quality of the end-product [
13].
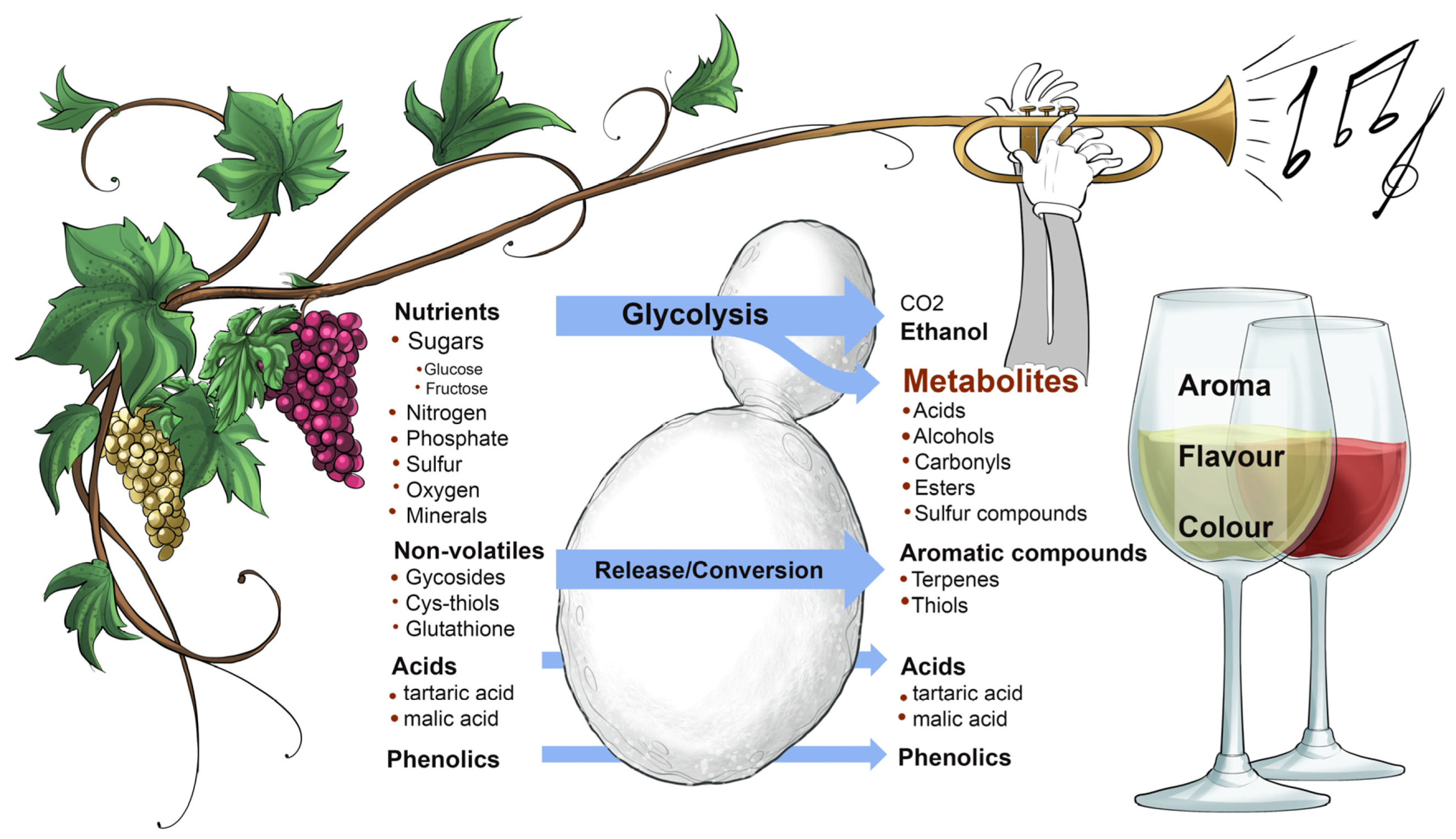
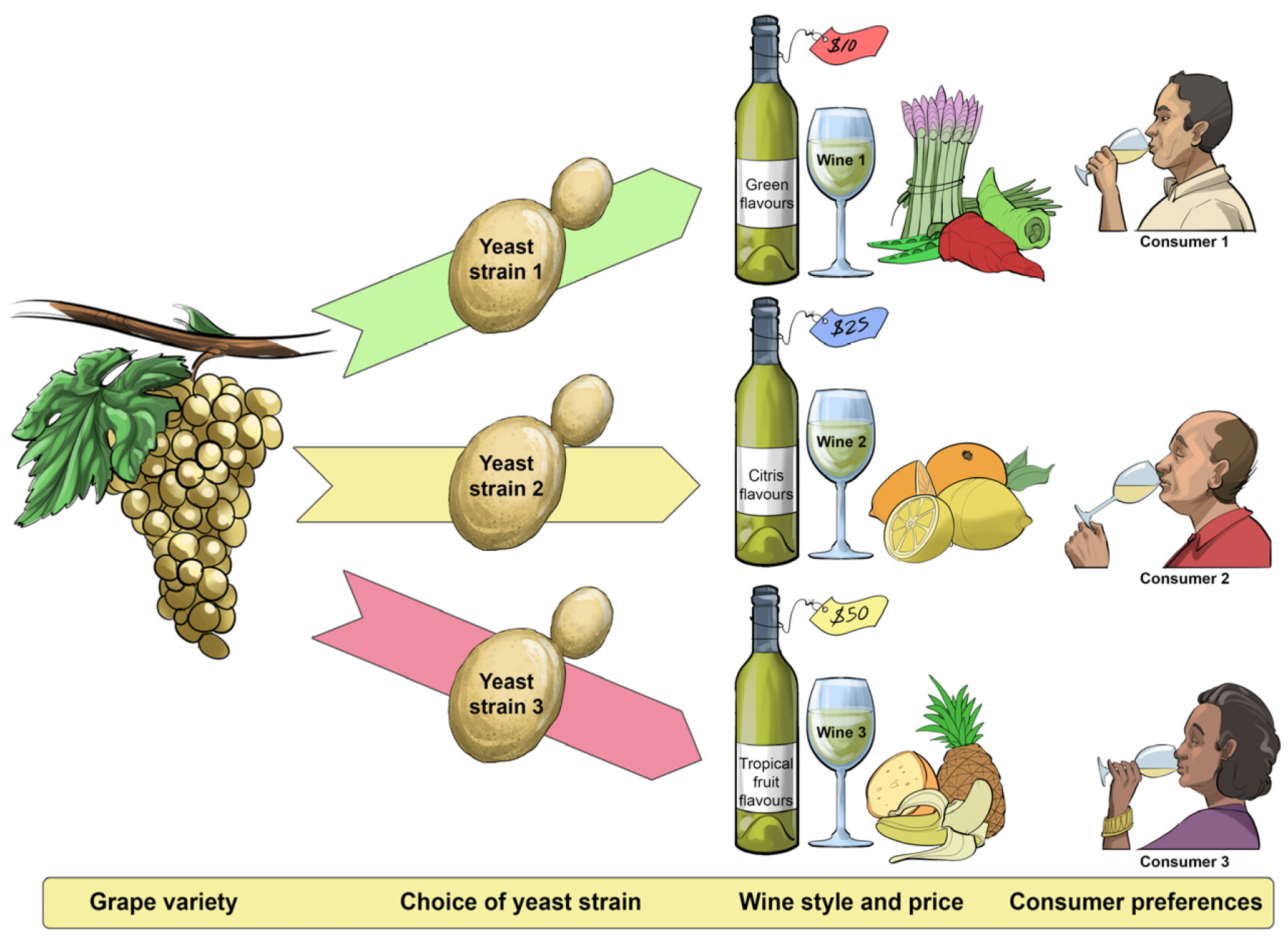
5. Fine-Tuning Saccharomyces cerevisiae Strains for Different Wine Performances
In an orchestra, every musician must know their role in performing a particular music composition and they are responsible for fine-tuning their own instruments before the conductor steps up to the lectern to commence the performance. In inoculated ferments, winemakers know that the primary role of
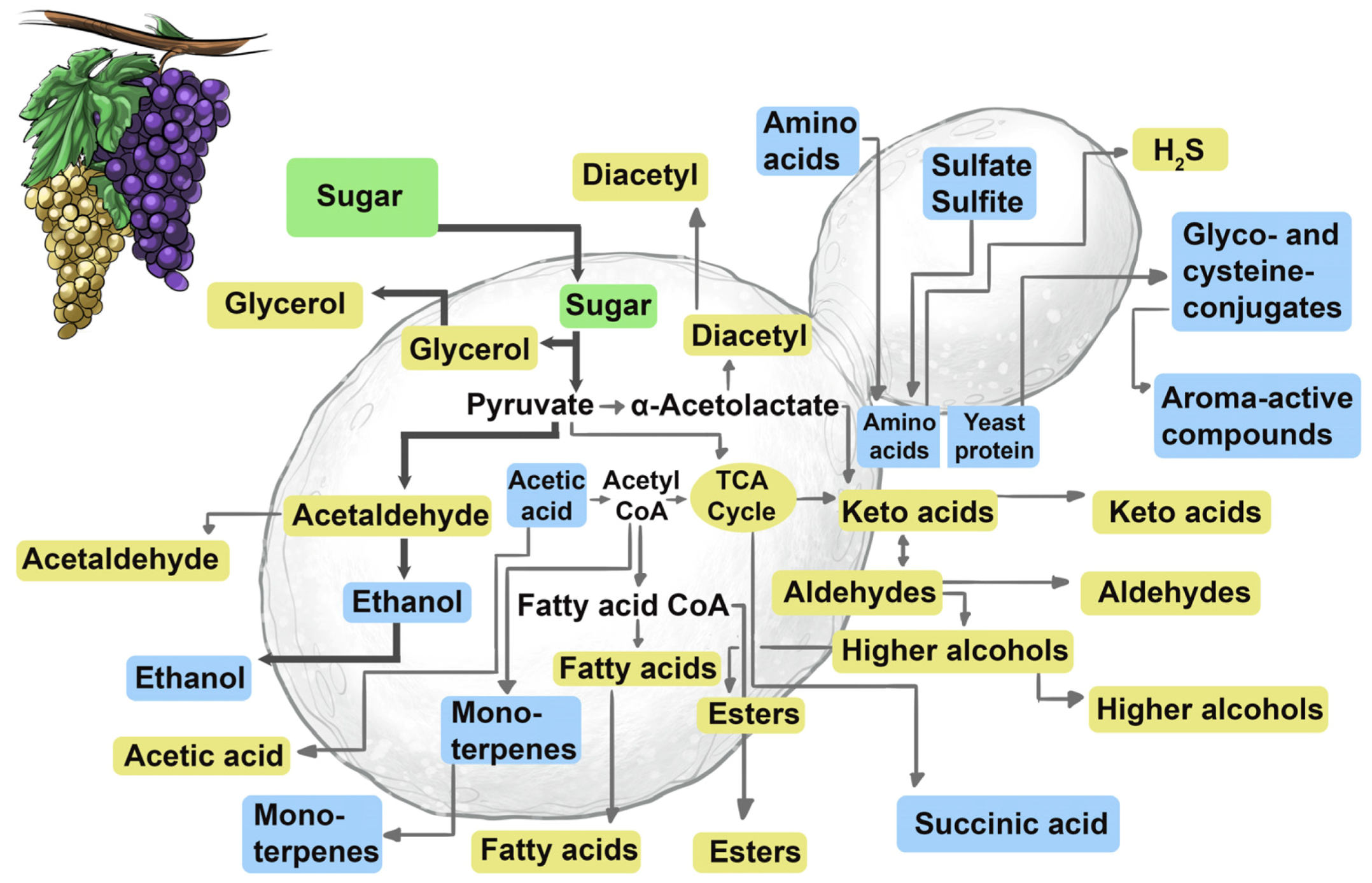
S. cerevisiae is to catalyse the rapid, complete and efficient conversion of grape sugars (mainly glucose and fructose) to ethanol, carbon dioxide and other desirable wine aroma-shaping metabolites (e.g., acids, alcohols, carbonyls, esters, terpenes, thiols) without the development of off-flavours (e.g., hydrogen sulfide) (
Figure 11 and
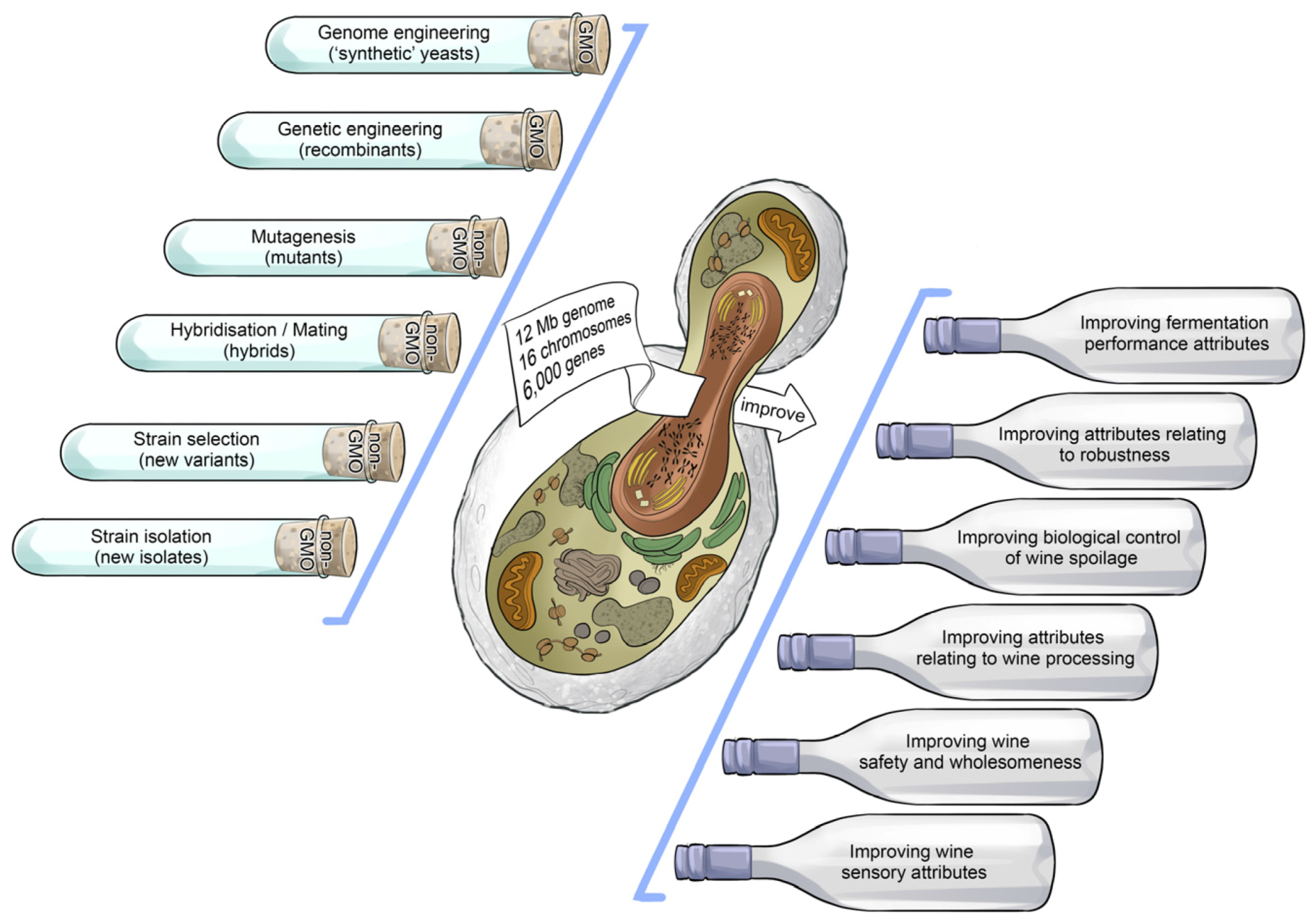
Figure 12). The most exquisite performance comes only from a precisely-tuned instrument, and so it is also with inoculated yeast. Strain development approaches usually start with the isolation and selection of variants according to their robustness, fermentation performance and sensory attributes (
Figure 13). Once the best candidate strains have been identified, yeast researchers employ hybridisation, mutagenesis, genetic engineering or genome engineering techniques to genetically improve the oenological properties of selected strains (
Figure 14). Improved mutants and hybrids are routinely used in commercial winemaking whereas genetically modified (GM) strains generated through genetic and genome engineering are largely reserved for research purposes at this stage [
6,
9,
19,
20,
21,
22,
23,
24]. Researchers are investigating non-GM strategies, including hybridisation, mutagenesis and adaptive laboratory evolution, to circumvent the current anti-GM constraints from consumers facing comestible food products generated through bioengineering strategies [
25].
5.1. Toning Down Disharmonious Off-Flavours in Wine
Under-prepared musicians, faulty instruments and discordant notes can spoil the entire performance of an orchestra and cause long-term reputational damage with audiences. Likewise, unbalanced wines or wines containing unwanted flavours can damage a brand and deter consumers from repeat-sales.
Just like not all violinists are Yehudi Menuhin, not all strains of
S. cerevisiae are superstar performers. Most commercially-available wine strains of
S. cerevisiae are able to catalyse the rapid, complete and efficient conversion of grape sugars to ethanol and carbon dioxide but not all of these strains produce optimal concentrations of desirable secondary metabolites, and in some instances even produce off-flavours under certain fermentation conditions. The genetic variation among wine strains of
S. cerevisiae is well documented [
26,
27,
28,
29,
30,
31,
32].
Breeding of new strains has been quite effective in providing winemakers with better options to improve fermentation performance and the profile of aroma compounds of wines [
33,
34,
35]. There are several examples of commercialised hybrid strains (e.g., NT116, VIN13, VL3, etc.) originating from mating and cross-breeding [
36]. Similarly, interspecific hybridisation between commercial
S. cerevisiae strains and closely-related species of the
Saccharomyces sensu stricto complex (
S. arboricolus,
S. cariocanus,
S. eubayanus,
S. kudriavzevii,
S. mikatae,
S. paradoxus,
S. uvarum and
Naumovozyma castellii) has also been used to introduce aroma and flavour diversity to wines as a means of improving style and increasing product differentiation [
32,
33,
34,
35].
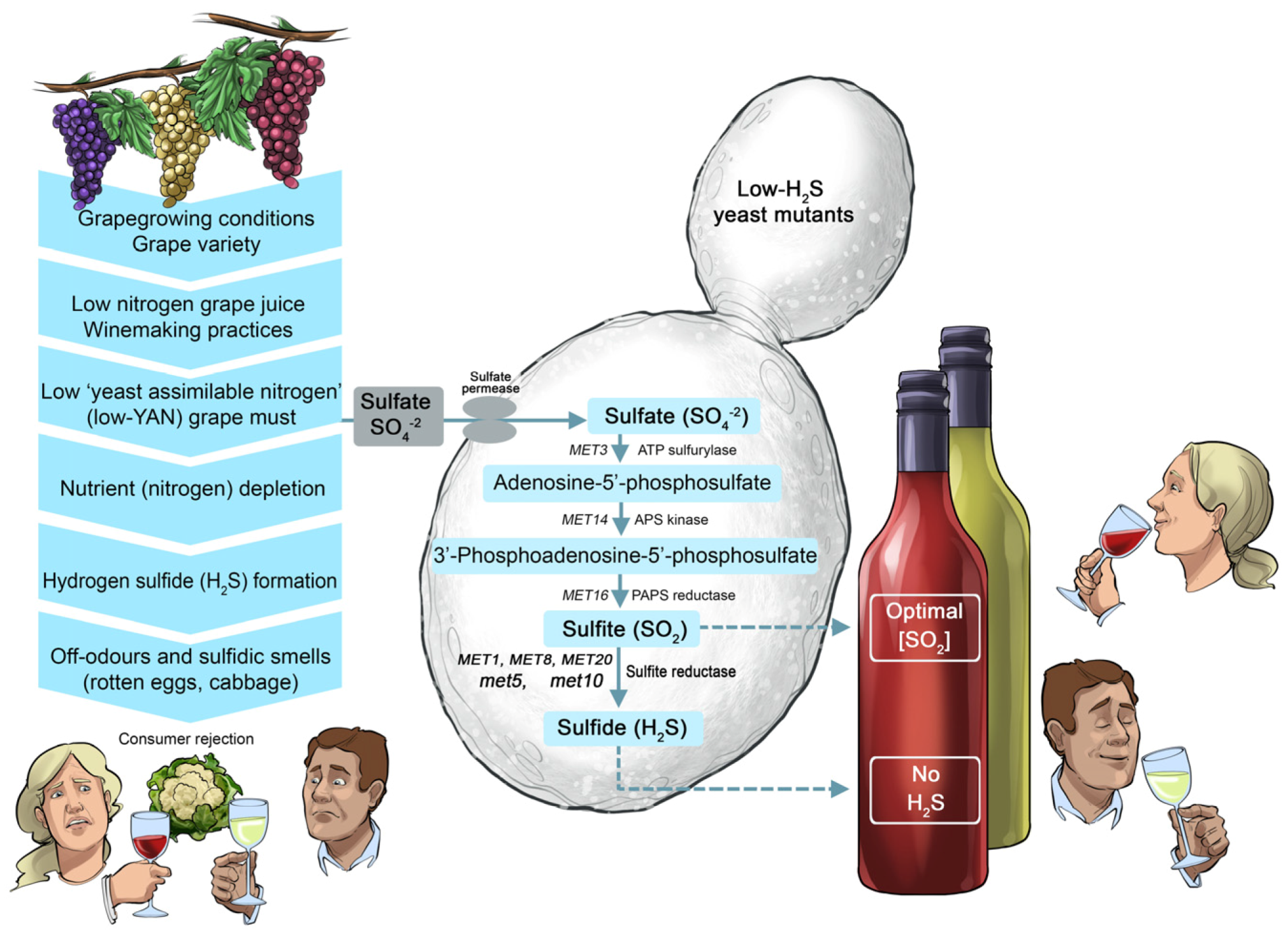
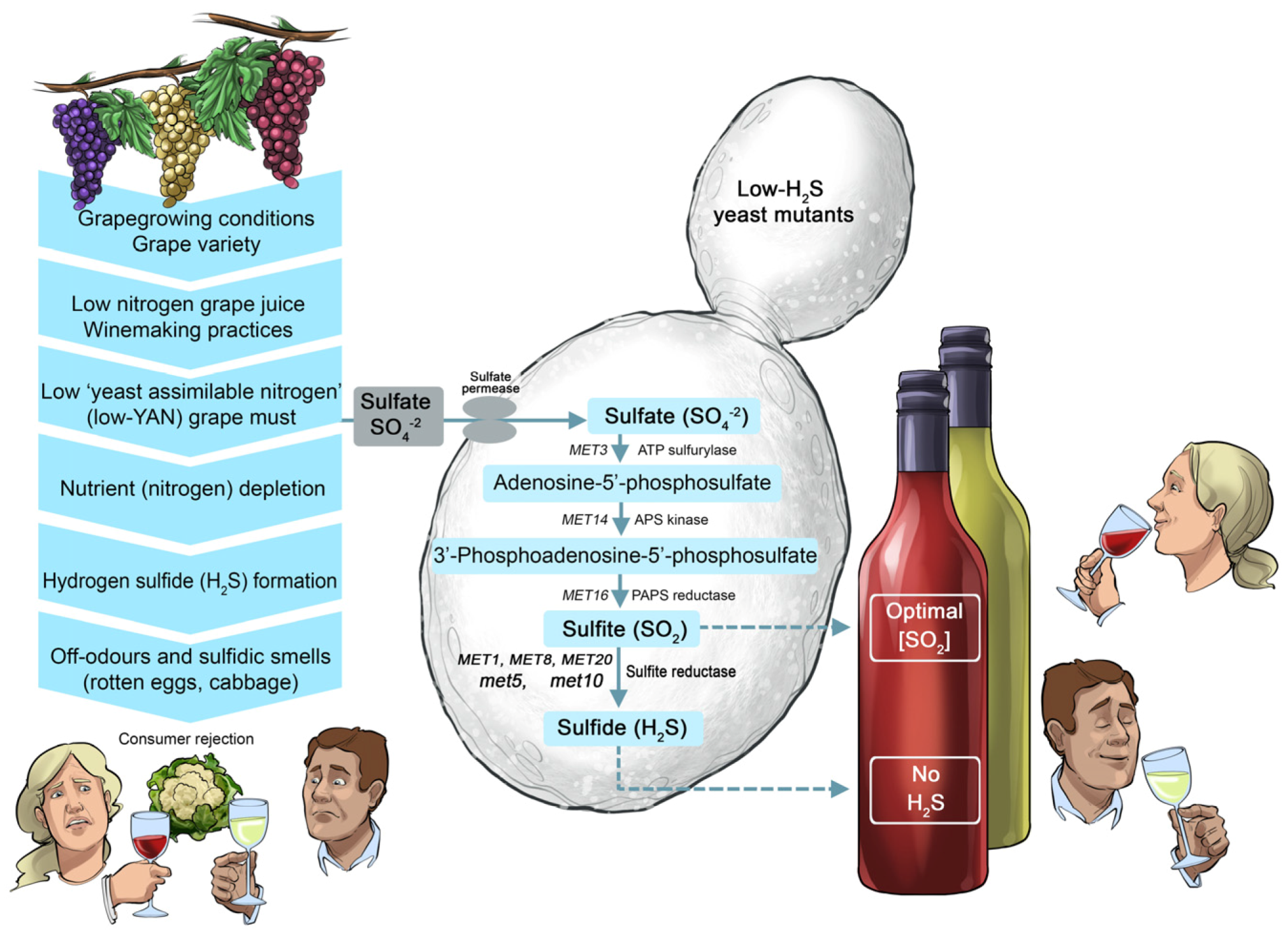
There are also examples where mutagenesis was effectively applied to improve existing commercial wine strains so that no longer can they form off-flavours (
Figure 15). The development of low-H
2S and low-volatile acidity (VA) mutants of the Maurivin PDM strain is a case in point [
37,
38,
39]. The low-H
2S variants of PDM were generated by mutating the
MET10 and
MET5 genes, which encode the catalytic α- and β-subunits of the sulfite reductase enzyme. These mutations significantly decreased or eliminated the capacity to produce unwanted ‘reductive’ characters (typified by cabbage-like aromas) during the fermentation of grape musts low in nitrogen [
37,
38]. Further improvement of these strains was achieved by generating mutants that produced considerably less acetic acid (the main component of volatile acidity and colloquially referred to as VA by winemakers) [
39,
40]. When present in concentrations above the sensory threshold, acetic acid imparts an undesirable vinegary character. These low-VA strains had either mutations in the
YAP1 (involved in oxidative stress) or
AAF1 (involved in fermentation stress response) gene.
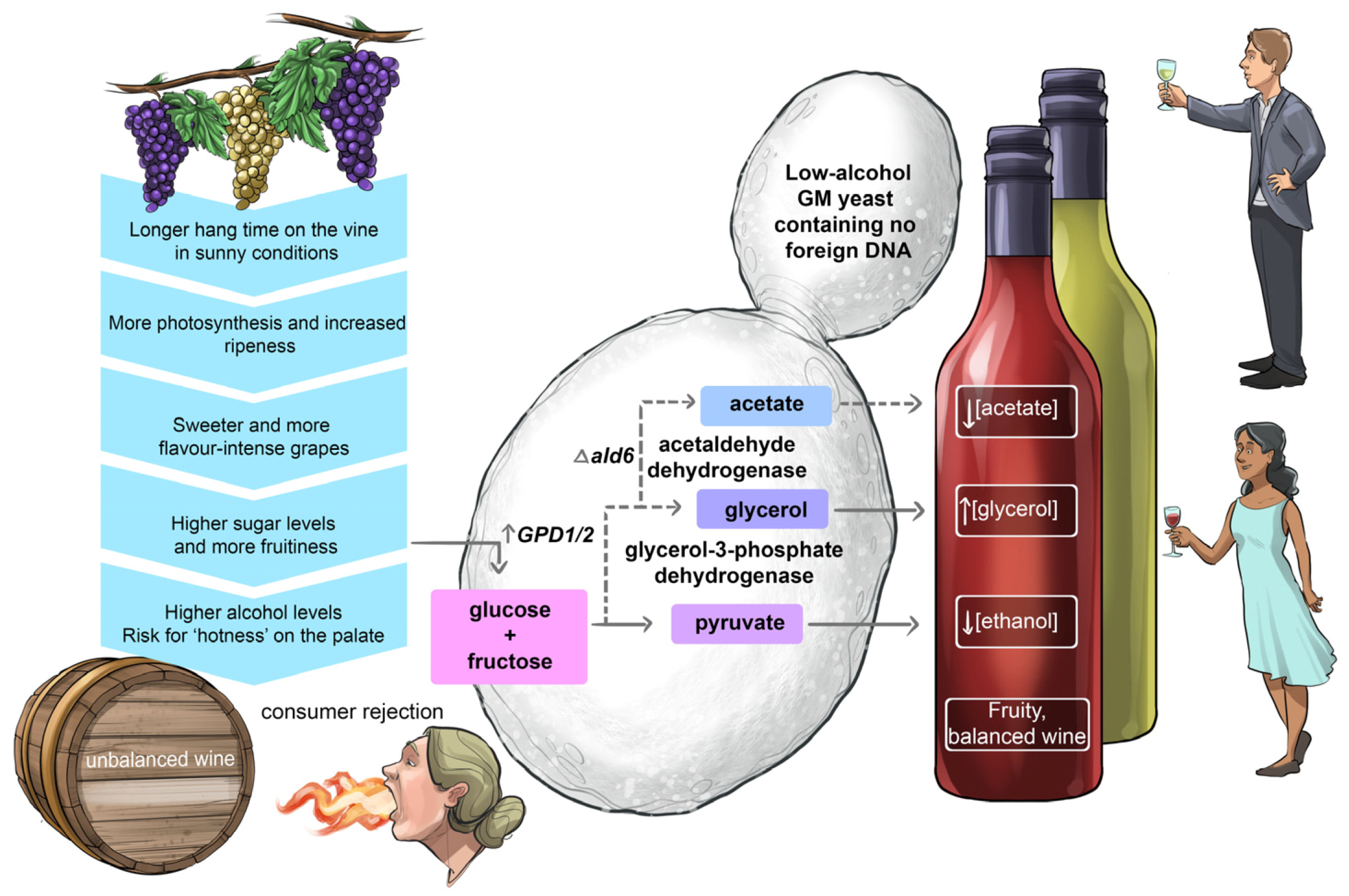
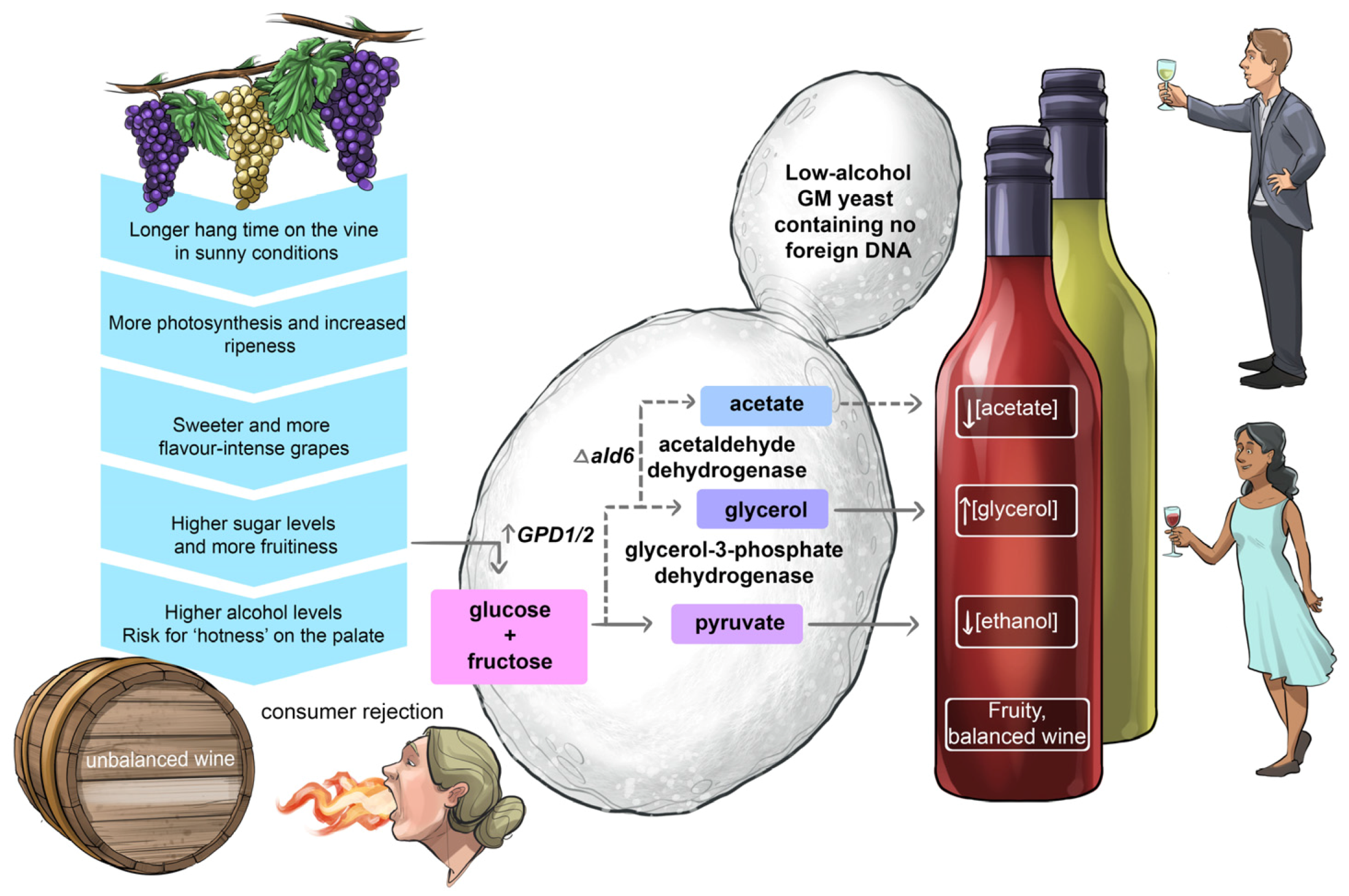
5.2. Striking the Right Chord between Alcohol Strength and Fruitiness in Wine
Like a conductor striving to harmonise all the sounds from all instruments playing in an orchestra, a winemaker’s goal is to produce well-balanced wines where the alcoholic strength, acidity, sweetness, fruitiness and tannin structure complement each other so that no single component dominates on the palate. To strike the right chemical balance in wine is no small feat. For example, many consumers prefer rich and fruitier styles of wine made from mature, ripe grapes. Such wines are free from undesirable unripe, vegetable flavours but they also contain much higher levels of alcohol than wines made from early-harvested grapes. Depending on the particular style, too high a concentration of ethanol can be perceived as ‘hotness’ on the palate, making overly-alcoholic wines appear unbalanced [
41,
42,
43].
In an attempt to generate non-GM low-ethanol wine strains for the production of consumer demand-driven fruity wines with reduced alcoholic strength, an adaptive laboratory evolution approach was used [
44,
45,
46]. This strategy was reasonably successful in that the resulting mutants produced significantly higher concentrations of glycerol at the expense of ethanol biosynthesis.
Over the past two decades, several genetic engineering strategies have also been explored to generate wine strains capable of partially diverting carbon flow away from ethanol (
Figure 16). The most promising results were achieved by over-expression of either one of the two paralogue genes,
GPD1 and
GPD2, encoding glycerol-3-phosphate dehydrogenase isozymes [
47,
48,
49,
50,
51,
52,
53]. For example, multiple copies of
GPD1 gene constructs were successfully integrated into the genome of two wine strains and these bioengineered strains were able to lower the ethanol content from 15.6% (
v/
v) to 13.2% (
v/
v) and 15.6% (
v/
v) to 12% (
v/
v) in Chardonnay and Cabernet Sauvignon wines, respectively [
53]. Unfortunately, both strains produced unacceptable concentrations of acetaldehyde and acetoin, imparting negative ‘bruised apple’ aromas to the resulting wine. To redirect the metabolic flow away from acetaldehyde, acetoin and acetic acid towards the sensorially neutral metabolite 2,3-butanediol, the
BDH1 gene—encoding butanediol dehydrogenase—was overexpressed in these high-glycerol/low-ethanol GM stains. This resulted in a significant decrease in the formation of acetaldehyde, acetic acid and acetoin; however, unexpectedly the overexpression of
BDH1 also interfered with the formation of glycerol and ethanol, most likely driven by alterations in the redox balance of the yeast’s metabolism [
53].
The desire to tone down the decibels of ethanol in fruity wines made from well-ripened grapes remains high on the priority list of vintners; however, the development of an ‘ideal’ low-alcohol wine yeast that can achieve an optimal balance between alcoholic strength and fruitiness can at best be described as a ‘work in progress’.
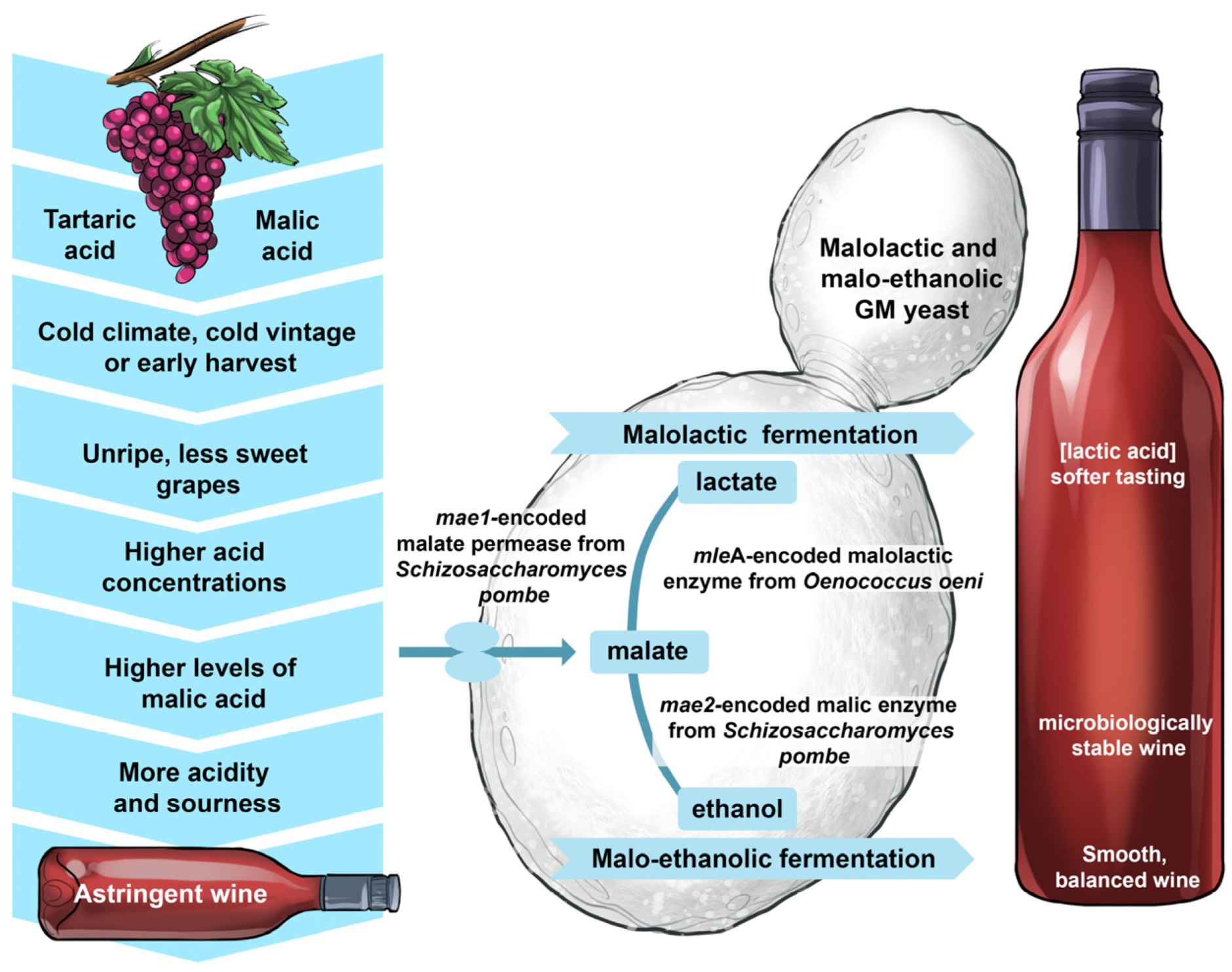
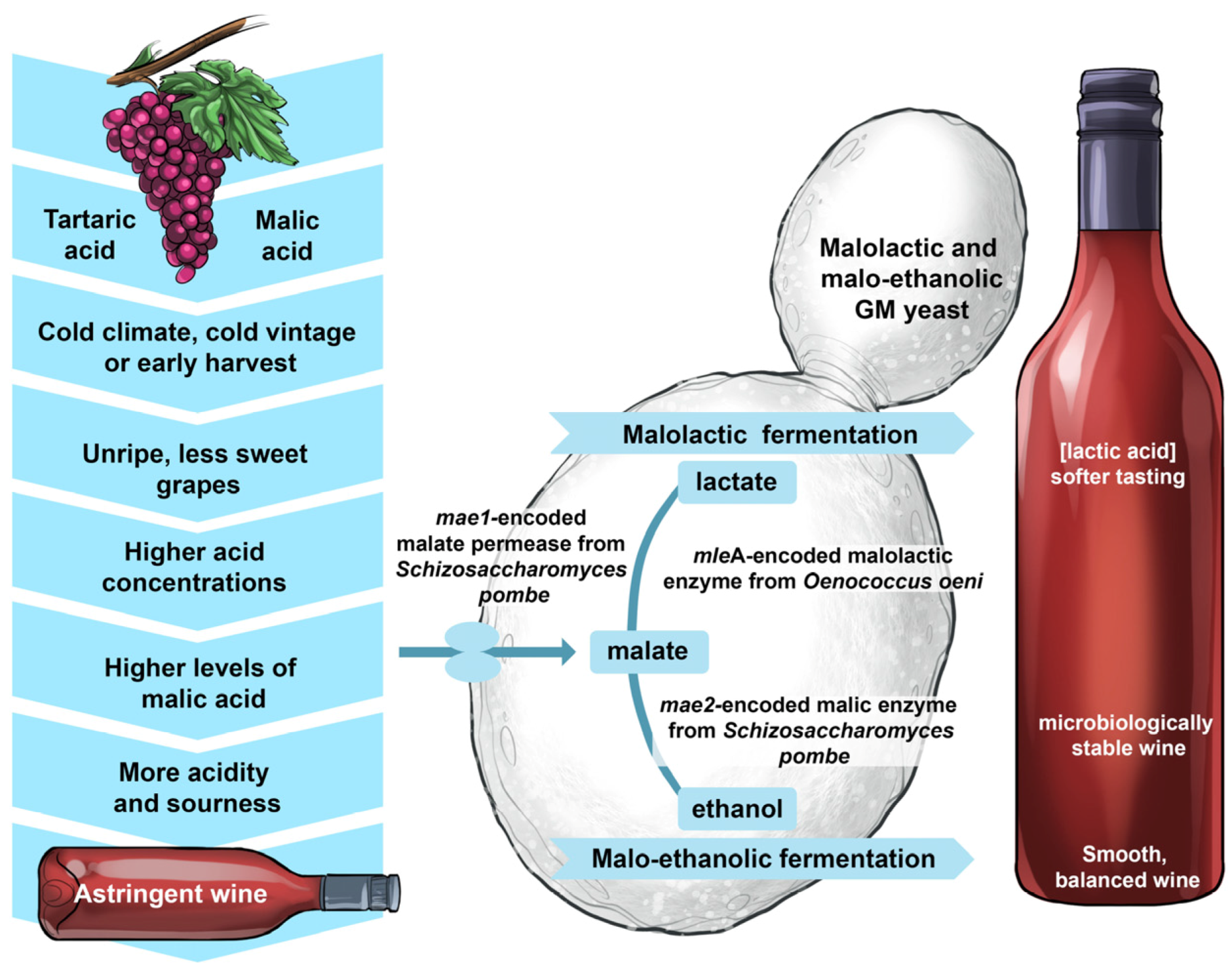
5.3. Performing Effortlessly in Solo Malolactic and Malo-Ethanolic Fermentations
There is no doubt that it would demand much more effort to direct a large philharmonic orchestra than, say, a string quartet—the less participants, the less effort is required. This intuitive assumption can be extended to winemaking—the less microbes involved, the less complex the process. This was the objective when researchers developed yeast strains capable of conducting both the alcoholic and malolactic fermentation [
54,
55].
Normally, malolactic fermentation is driven by fastidious lactic acid bacteria, such as O. oeni, following alcoholic fermentation conducted by S. cerevisiae yeast. During malolactic fermentation, grape-derived malic acid is deacidified (decarboxylated) to lactic acid. It often happens that malolactic fermentation is ‘sluggish’ or gets ‘stuck’ because of inhibitory conditions ranging from low pH, high alcohol content and poor nutrient availability. There is also the potential for some bacteria to produce biogenic amines that can impose health risks to consumers of fermented products. A wine yeast strain that is able to convert grape sugars into ethanol, carbon dioxide and flavour-active metabolites as well as sharp-tasting malic acid into soft-tasting lactic acid would therefore simplify and improve the reliability of the winemaking process.
A widely-used wine strain of
S. cerevisiae, equipped with the
Schizosaccharomyces pombe malate transporter gene (
mae1) and the
O. oeni malolactic enzyme gene (
mleA), was engineered, thoroughly tested, cleared by the American and Canadian regulatory authorities and commercialised under the name ML01 [
54,
55]. These two genes were placed under the control of the strong constitutive regulatory sequences of the
S. cerevisiae PGK1 promoter and terminator and integrated into the genome of the host strain. ML01 is capable of performing malolactic fermentation in parallel with alcoholic fermentation (
Figure 17). In fermentation trials, it was shown that 5 g/L of malic acid was decarboxylated to lactic acid within five days, without negative impacts on the sensory aspects on wine. Further analyses of the phenotype, genotype, transcriptome and proteome revealed that ML01 was ‘substantially equivalent’ to its parental industrial wine yeast.
A second variant also became available with the integration of the S. pombe malate transporter gene (mae1) and malic enzyme gene (mae2) into the genome of the engineered strain. With this strain, malate is decarboxylated to pyruvate, which is then converted to ethanol.
In 2005, the ML01 malolactic strain became the first commercialised GM wine yeast released to the market. However, the prevailing anti-GM sentiments from consumers have prevented the widespread use of this strain in commercial wine production.
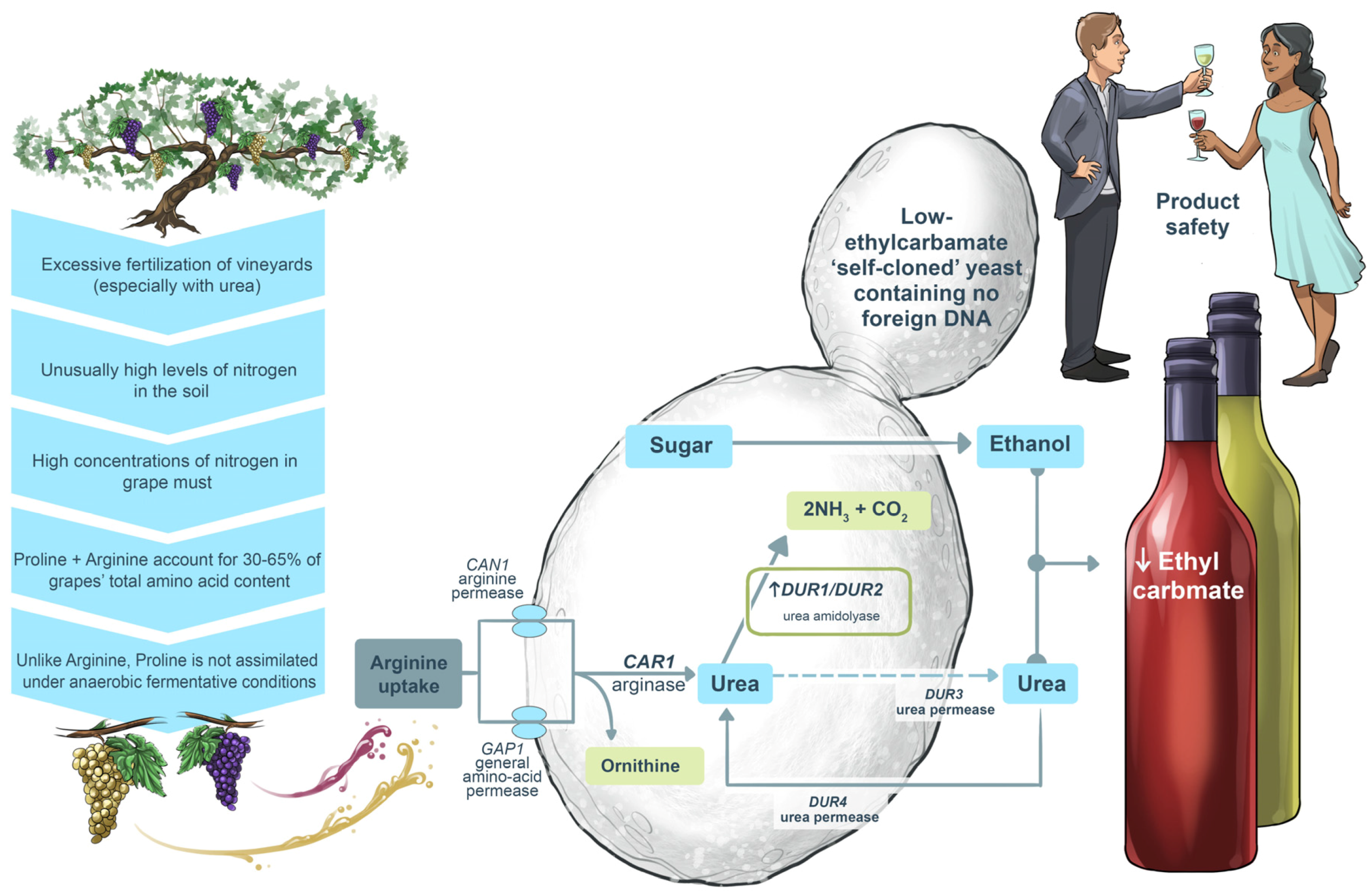
5.4. Safeguarding against Unhealthy Notes of Ethyl Carbamate in Wine Production
A second GM wine yeast, ECMo01, obtained clearance from the American and Canadian regulatory bodies in 2006 [
56]. ECMo01 was constructed to reduce the risk of ethyl carbamate production during fermentation. This potentially carcinogenic compound is the product of urea reacting with ethanol. Fortunately, ethyl carbamate is ordinarily produced at such low levels (if at all) in winemaking that it is normally not a concern. Nonetheless, it is present in some fortified wines from certain wine-producing regions.
ECMo01 has an extra copy of the
S. cerevisiae DUR1,2 gene under the control of the yeast
PGK1 regulatory sequences (
Figure 18). This gene codes for urea amidolyase, which converts urea into ammonia and carbon dioxide, thereby removing substrate for ethyl carbamate production. The ammonia formed by this reaction is consumed as a preferred nitrogen source by the yeast cells. ECMo01 has been reported to reduce ethyl carbamate in Chardonnay wine by almost 90%, and analyses of ECMo01’s phenotype and transcriptome also revealed that the ECMo01 yeast is ‘substantially equivalent’ to its parental strain [
56]. However, although this ECMo01 is a
cis (or ‘self’) cloned yeast (it carries no foreign DNA and, therefore, is not transgenic), it suffered the same fate as ML01in the market place in terms of reluctance to use wine yeasts in commercial wine production that are considered to be genetically-modified organisms (GMOs).
5.5. Directing a Symphony of Aromas and Flavoursome Tastes in a Wine Glass
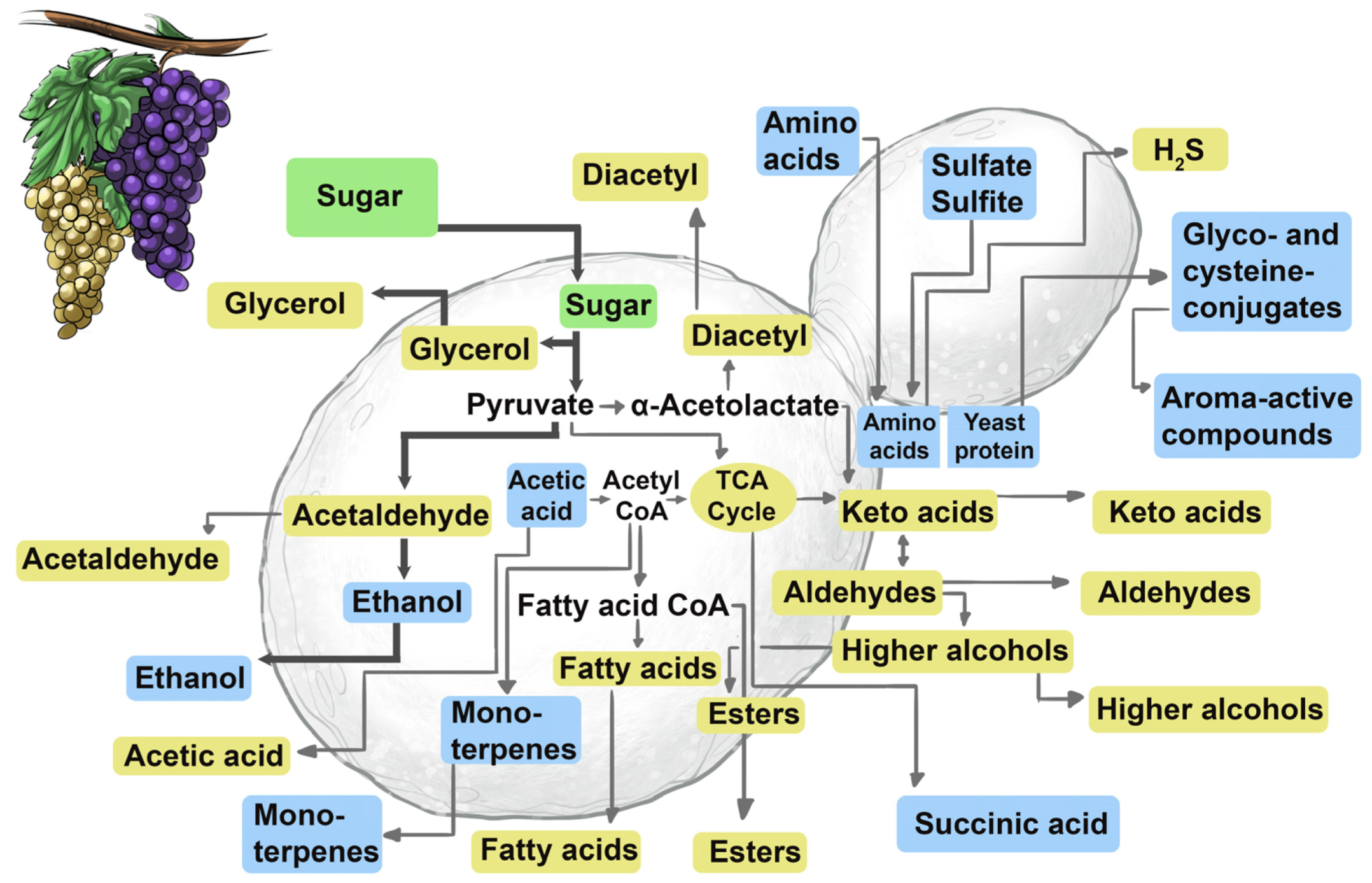
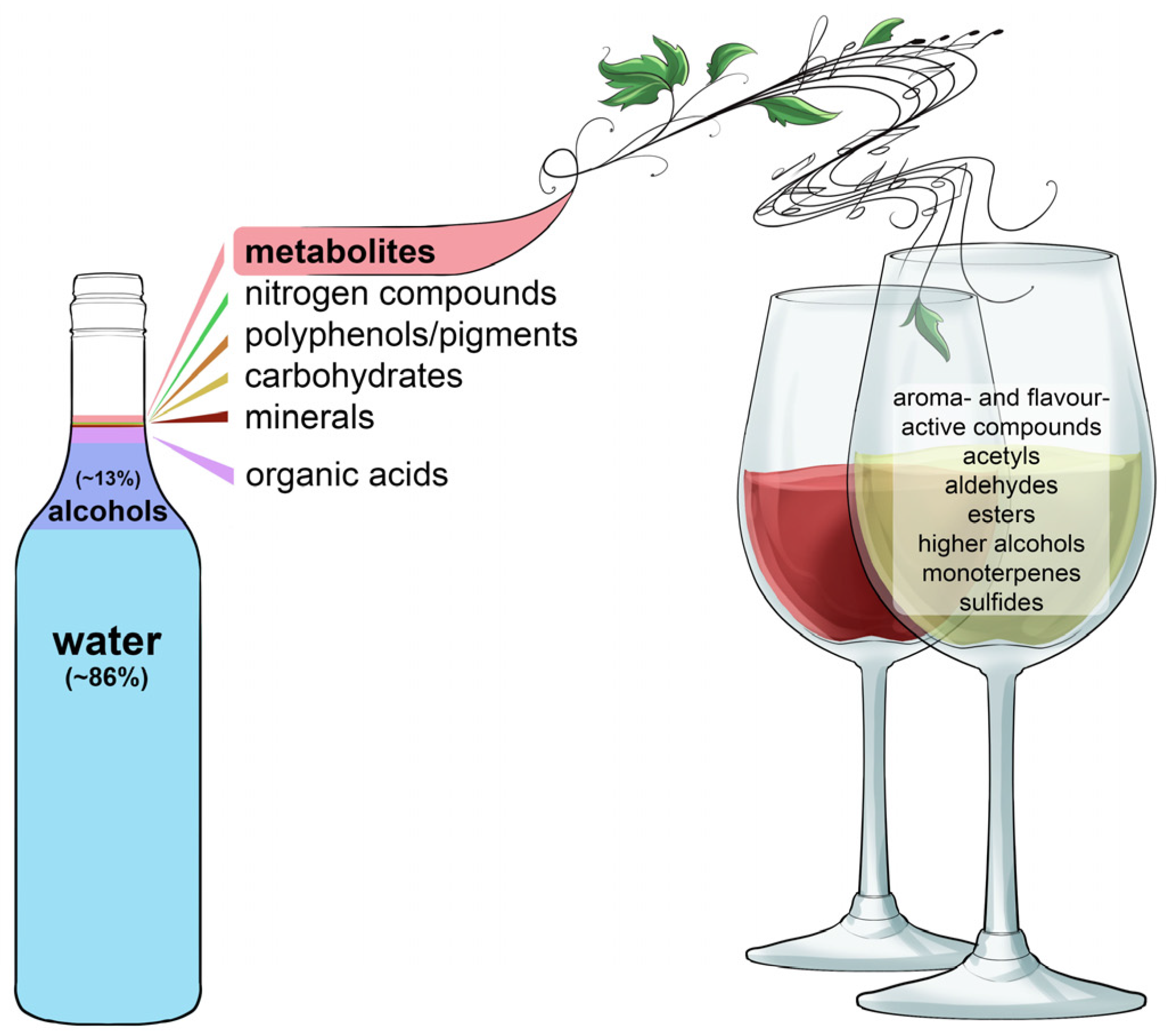
The combination and harmony of complex notes produced by a symphony orchestra determine the quality of their performance. This can also be said for the complex mixture of chemical compounds that define a wine’s appearance, aroma, flavour and mouth-feel properties (
Figure 19). The sensorially-active chemical compounds originate from the grapes, microbes and oak (when used).
The grape-derived compounds give wines their basic structure and provide their varietal distinction (
Figure 20). Examples of flavour-active grape-derived compounds are aliphatics, benzene-derivatives, esters (
cis-rose oxide, ethyl acetate, ethyl butyrate, ethyl formate, ethyl hexanoate, ethyl propionate, furaneol, isoamyl acetate, isobutyl acetate, methyl cinnamate), lactones (sotolon), norisprenoids (e.g., β-demascenone, β-ionone, 1,1,6-trimetyl-1,2-dihydronaphtalene), phenols, pyrazines (e.g., 3-isobutyl-2metoxypyrazine), sesquiterpenes (e.g., rotundone), monoterpenes (e.g., linalool, geraniol, hotrienol, nerol, citronellol,
cis-rose oxide and α-terpineol) and thiols (e.g., 4-mercapto-4-metyl-pentan-2-one, 3-mercaptohexan-ol, 3-mercaptohexyl acetate, etc.) [
7]. Each of these compounds impart, individually or in combination with others, specific characters to wines [
8]. For instance, the ‘bell pepper’ character of Cabernet Sauvignon stems from 3-isobutyl-2-methoxypyrazine while the strawberry-like characters of Pinot Noir are driven by a combination of several esters, including ethyl acetate, ethyl butyrate, ethyl formate, ethyl hexanoate, furaneol and methyl cinnamate [
8]. However, some esters can also act in pairs or individually as aroma-defining compounds. For example, the fig-like character of Semillon is the result of two esters, ethyl propanate and isobutyl acetate [
8]. An example of an ester that acts individually is
cis-rose oxide that determines the well-known lychee-like flavour of Gewürztraminer. When Gamay grapes undergo carbonic maceration, isoamyl acetate imparts pronounced banana-like aromas in the resulting wine [
8]. Terpenes are another class of compounds that determine characteristic grape flavours and aromas; for example, the sesquiterpene rotundone is responsible for the spicy and peppery notes in Shiraz [
57,
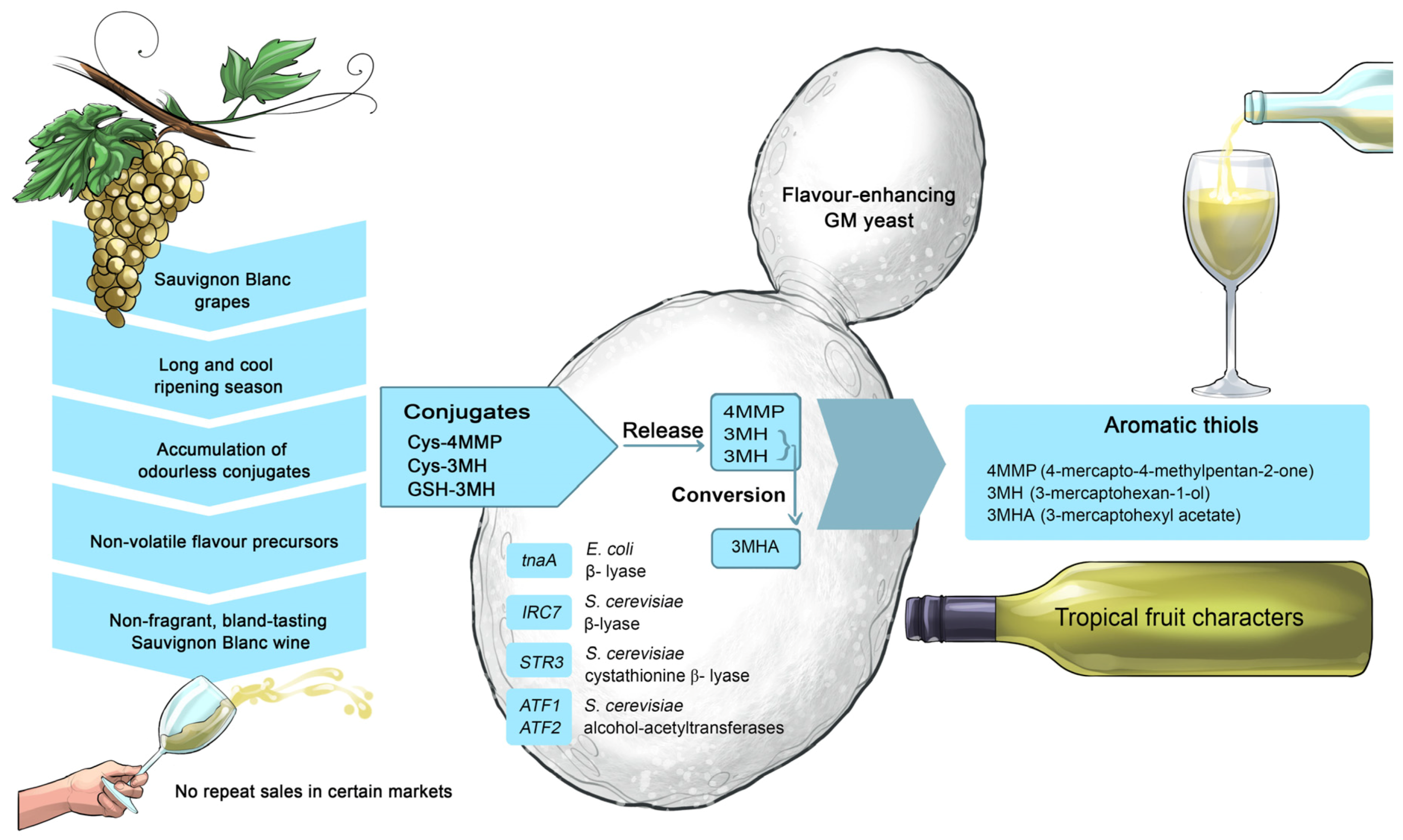
58] and the monoterpene geraniol defines the ‘grapey’ flavour of Muscat grapes. The lactone, sotolon, is responsible for the honey-like characters in botrytised wines and 1,6 trimethyl-1,2-dihydronaphthalene drives the kerosene-like characters in Riesling. Thiols play a major role in the fragrance of Sauvignon Blanc: 4-mercapto-4-methylpentan-2-one (4MMP) imparts grassy and box-tree flavours while 3-mercaptohexan-ol (3MH) and 3-mercaptohexyl acetate (3MHA) are responsible for tropical fruit flavours similar to those typifying the smell of gooseberries and passion-fruit [
59,
60,
61,
62,
63,
64,
65,
66,
67,
68,
69].
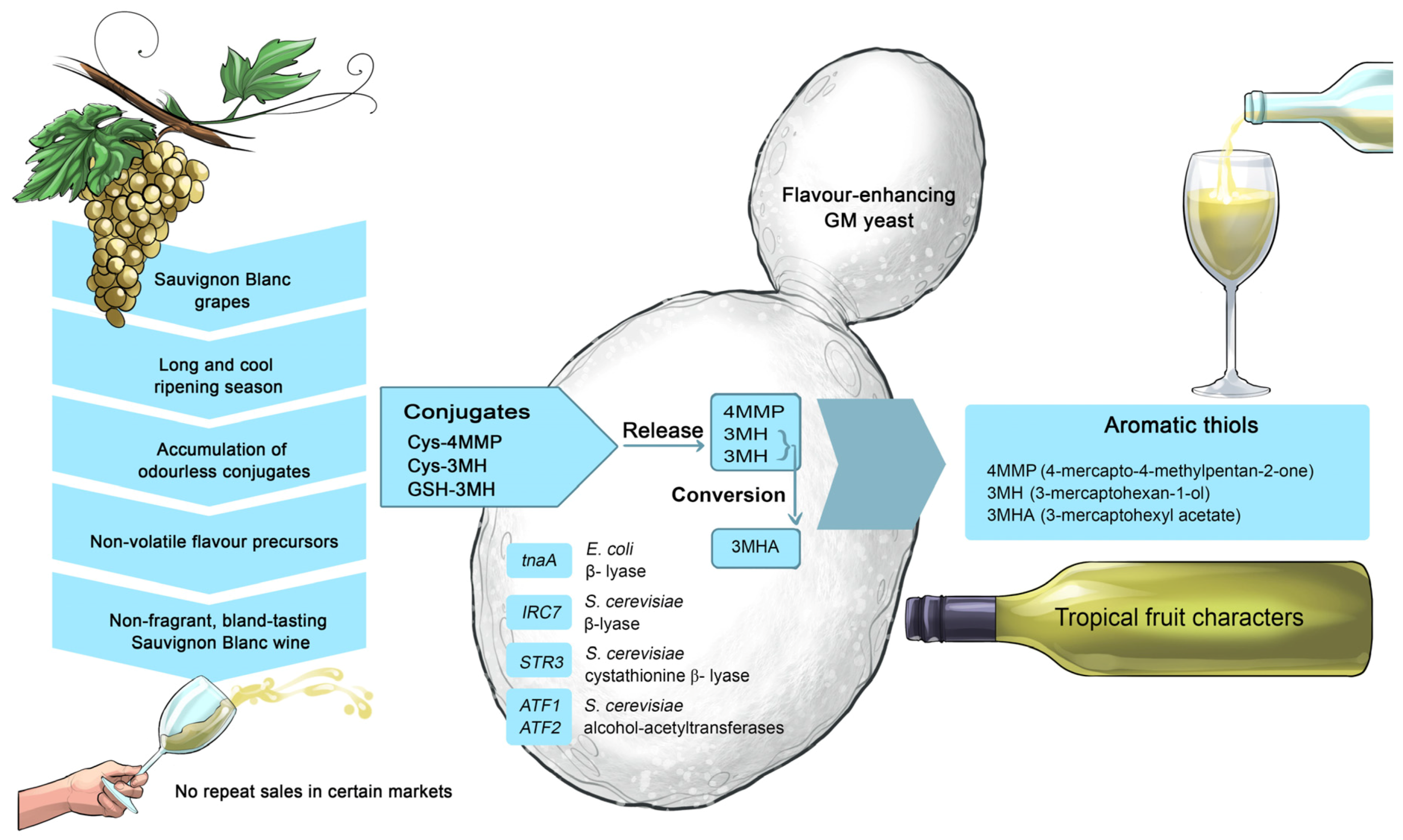
The interaction of yeasts with these grape-derived compounds and the de novo synthesis of its own secondary metabolites (esters, fusel alcohols, carbonyls, volatile fatty acids, sulfur compounds, etc.) give wine its ‘vinous’ character [
65,
70,
71,
72]. During the alcoholic fermentation, several grape-derived compounds are biotransformed and released from grape-derived precursors. For example, non-volatile cysteinylated grape thiols (Cys-4MMP and Cys-3MH) can be released from cysteine (4MMP and 3MH) and converted into the more potent volatile compound, 3MHA [
58,
59,
60,
61,
62,
63,
64,
65,
66,
67,
68]. Wine yeast strains can be engineered to enhance their capacity and capability to modulate these grape-derived thiol precursors [
65] (
Figure 21). During malolactic fermentation, malolactic bacteria deacidifies wine by converting malic acid into lactic acid, and produces additional metabolites, such as diacetyl which imparts a buttery flavour to wine [
7]. In addition, when wine comes in contact with oak during fermentation and/or maturation, a series of complex chemical transformations can result in significant changes in the aroma and flavour profile of the end-product [
73]. Toasted oak staves are often used in oak barrel manufacturing and the related potent aroma compounds (vanillin and vanillin-derivates, volatile phenols and lactones) can strongly influence the aroma and style of wine. When yeast fermentation is conducted in oak barrels, some of these oak-derived compounds migrate from the wood to the fermenting grape must, which in turn, can be transformed into highly aromatic compounds [
73]. For instance, vanillin can be reduced by yeast to vanillic alcohol, which has a much lower sensory threshold than vanillin. Similarly, furfuryl alcohol and furfuryl mercaptan (2-furanmethanethiol) can be formed from oak-derived furfural. The accumulation of oak-derived compounds in wine during barrel storage increase the intensity of sensory descriptors such as coconut, spice and vanilla.
Vintners use a large number of variations of viticultural and oenological practices to obtain the optimal absolute and relative concentration of these grape-, oak- and microbial-derived chemical compounds to shape the aroma and flavour profile of their wines according to the preferences of their targeted consumer markets—a harmonious symphony in a wine glass.
6. Orchestrating Future Music in the Research Laboratory
No-one knows where music came from but we do know that the earliest human ‘music-makers’ used whatever was available to create musical instruments with which they could create rhythms for telling their stories. Their traditional poetry with rhythm eventually evolved into songs as they invented and improved their musical instruments. The invention of these first musical instruments came about accidentally, much like the early inventions in winemaking. As civilisations became more sophisticated, inventions became more deliberate until innovations became continuous.
The evolution of the earliest forms of, say, the harpsichord, clavichord, piano and organ to today’s electronic keyboards, digital pianos and synthesisers is not dissimilar from the progression of yeast strain development—from the passive use of ambient yeasts in spontaneous fermentations to the application of hybrids, mutants and bioengineered strains with specialised winemaking properties. It is tempting to speculate that the invention of the first electronic music synthesiser might have been perceived by musicians and music enthusiasts as much of a threat to the future of, for example, classical music played by a philharmonic orchestra, as a yeast with ‘synthetic’ DNA would be perceived by some vintners and consumers to pose to the future of winemaking.
Over the years, those in the music industry have come to realise that composers, conductors and musicians are benefitting from the availability of synthesisers in writing their music, preparing for their performances and practicing their beloved art. They know that technology, in general, and digital music synthesisers, more specifically, are not a threat but a great help in their effort to improve their art form. Over the past decades, the wine industry benefitted enormously from a wide range of technological inventions; however, while the prospect of ‘synthetically-tailored’ yeast strains might be music to the ears of some avant-garde scientists, such news might send shivers down the spine of many winemakers and wine drinkers.
It is for this reason that the first notes of future music emanating from trailblazing Synthetic Biology laboratories about ‘synthetic’ yeasts be crystal clear and dealt with sensitively. Those who research in the emerging field of synthetic genomics have to work relentlessly to engage all stakeholders, including fellow scientists, industry practitioners, policymakers, regulators, commentators and consumers [
24]. It is important for researchers to explain that the development of ‘synthetic’ yeast models in the laboratory would help yeast researchers to further expand their understanding of the biological fundamentals, limitations and potential of existing wine yeasts—much like what the physicist Richard Feynman meant when he famously said “What I cannot create, I do not understand”. This ‘philosophy’ is what inspired a consortium of a dozen laboratories from five countries—the US, UK, Australia, China and Singapore—to embark on a collaborative project aimed at the synthesis of the entire genome of a laboratory
S. cerevisiae strain (S288c) by early 2018. The goals of this large international project—the Synthetic Yeast Genome Project (referred to as the
Yeast 2.0 or
Sc2.0 project)—include answering a broad spectrum of “profound questions about fundamental properties of chromosomes, genome organisation, gene content, function of RNA splicing, the extent to which small RNAs play a role in yeast biology, the distinction between prokaryotes and eukaryotes, and questions relating to genome structure and evolution” while recognising that “the eventual ‘synthetic yeast’ being designed and refined could ultimately play an important practical role” (
www.syntheticyeast.org). The early learnings from the
Yeast 2.0 project are already starting to forge new frontiers for wine yeast development [
24].
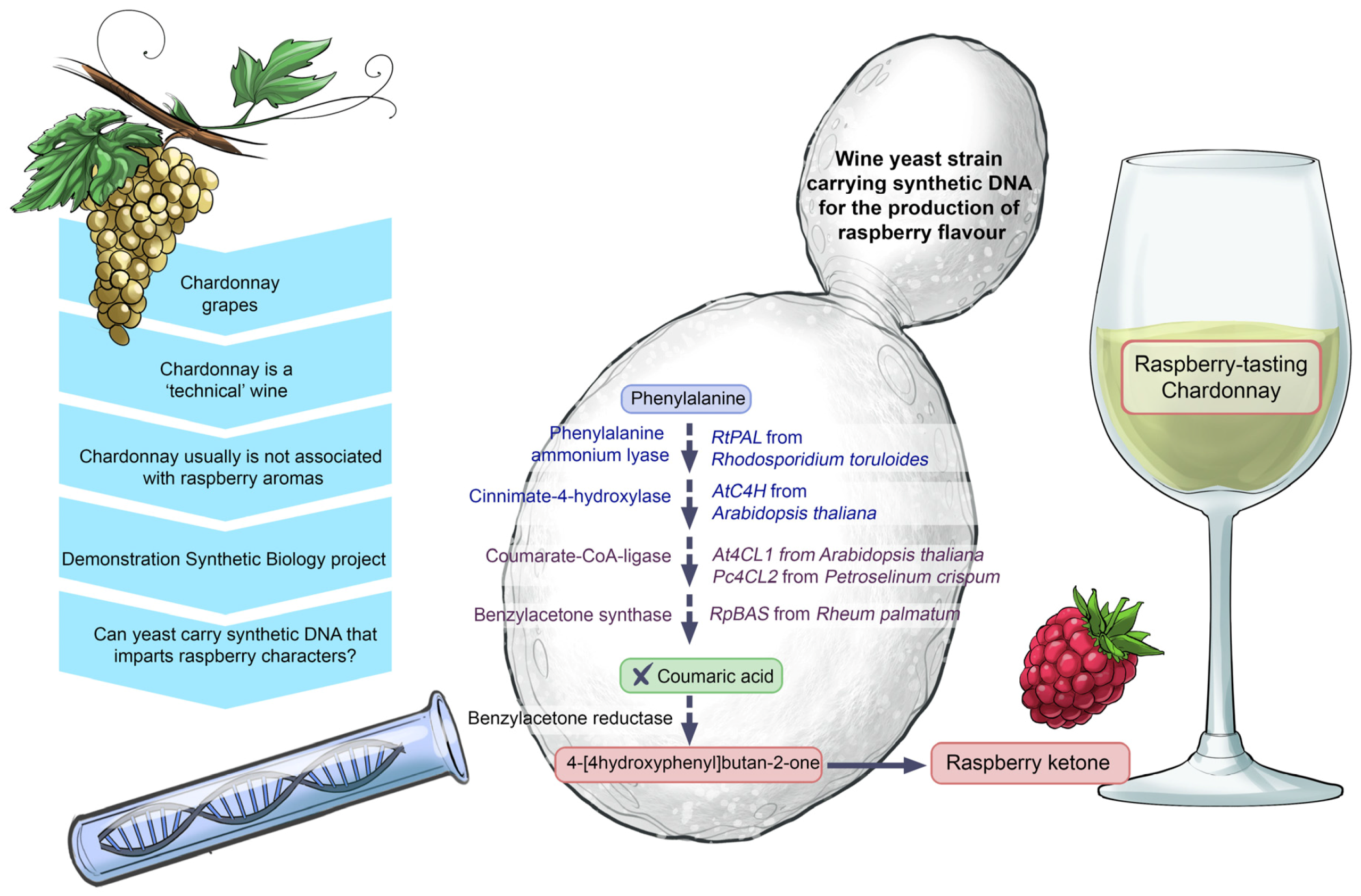
The technology used in the
Yeast 2.0 project on a laboratory yeast strain has indeed spilled over into wine yeast research with the development of a synthetic raspberry wine yeast model—the first wine yeast containing a ‘synthetic DNA circuit’. This wine strain of
S. cerevisiae was developed as an instrument for the unravelling of metabolic flavour pathways relevant to winemaking [
74]. The primary objective of this project was not to apply the ‘synthetic’ raspberry yeast in commercial winemaking; rather, the goal was to use the ‘synthetic’ yeast as a study model to uncover the biochemical pathways leading to the formation of the raspberry ketone, 4-[4-hydroxyphenyl]butane-2-one. A ‘DNA circuit’ consisting of four chemically-synthesised, codon-optimised genes were expressed in a
S. cerevisiae wine strain, AWRI1631. The synthetic copies were derived from the following genes:
RtPAL from an oleaginous yeast,
Rhodosporidium toruloides;
AtC4H from the well-studied model plant,
Arabidopsis thaliana;
Pc4CL2 from parsley,
Petroselinum crispum; and
RpBAS from rhubarb,
Rheum palmatum. The wine yeast containing the synthetic version of these genes was used to produce experimental Chardonnay, which smelled and tasted like raspberries (
Figure 22). This ‘synthetic’ wine yeast offers prospects for the development of non-GM wine yeast starter strains with a capability to intensify the raspberry notes in both red and white wine. If successful, this could assist winemakers in their effort to consistently produce wine to definable market specifications and styles.
The prospect of ‘synthetically-tailored’ Yeast 2.0 strains is much more real than most stakeholders in the wine industry realise—the Yeast 2.0 project is already 75% complete and on track to equip the first yeast cell with 16 chemically-synthesised chromosomes (~12 Mb carrying ~6000 genes). This yeast will become the first eukaryotic cell to cross the horizon of in silico design of complex cells through de novo synthesis, reshuffling and editing of yeast genomes. As with most disruptive innovations, the ‘techno’ sound of Yeast 2.0 future music is bound to unsettle and cause discordant notes within the global wine industry.
There is no question that the future music of scientific research and breakthrough discoveries in leading-edge yeast laboratories will continue to play on; however, the question is whether there will be receptive and appreciative audiences to applaud and call for encore performances.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}