
Research Progress on Cordycepin Synthesis and Methods for Enhancement of Cordycepin Production in Cordyceps militaris


Abstract
:1. Introduction
2. Chemical Synthesis of Cordycepin (COR)
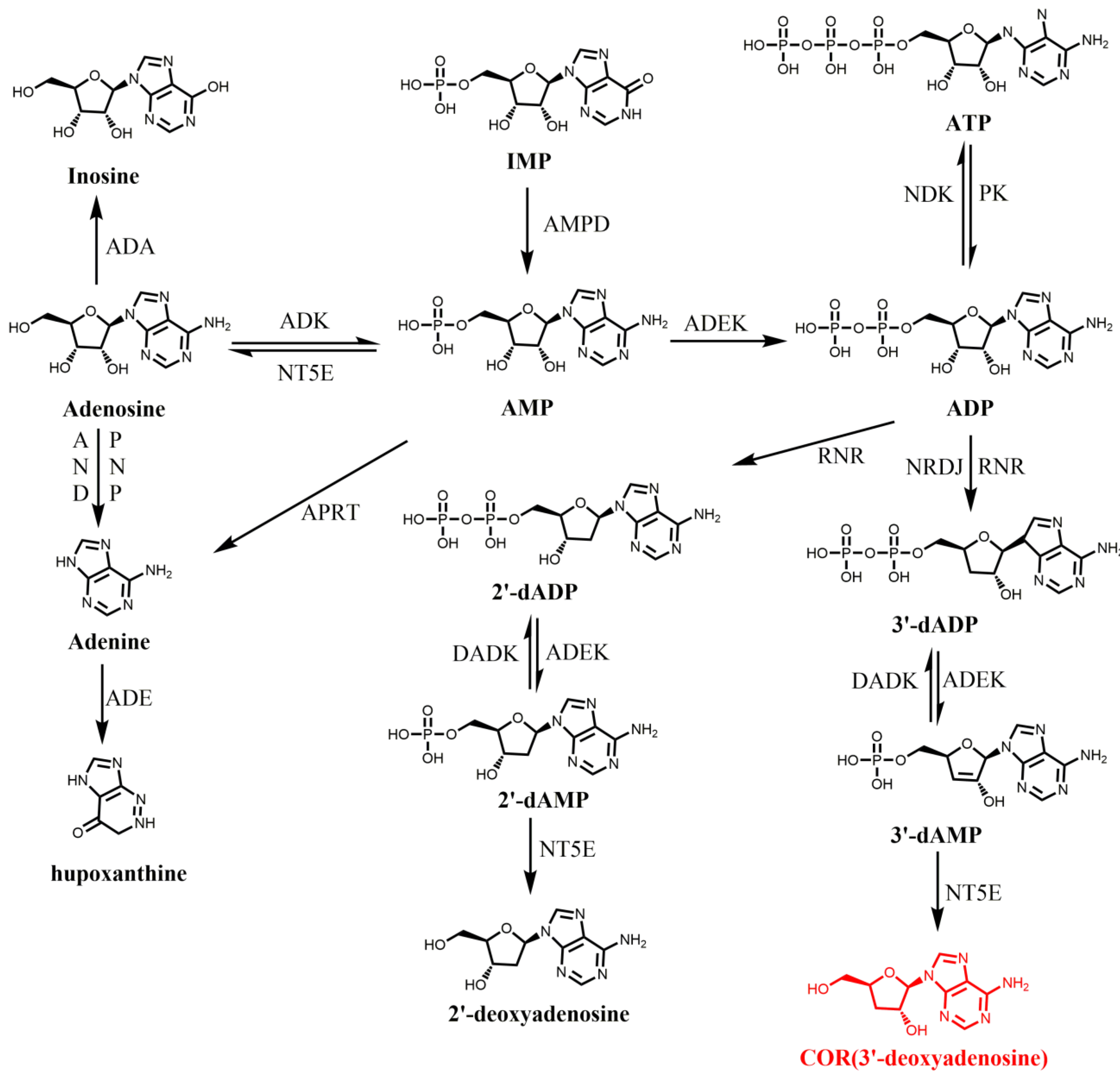
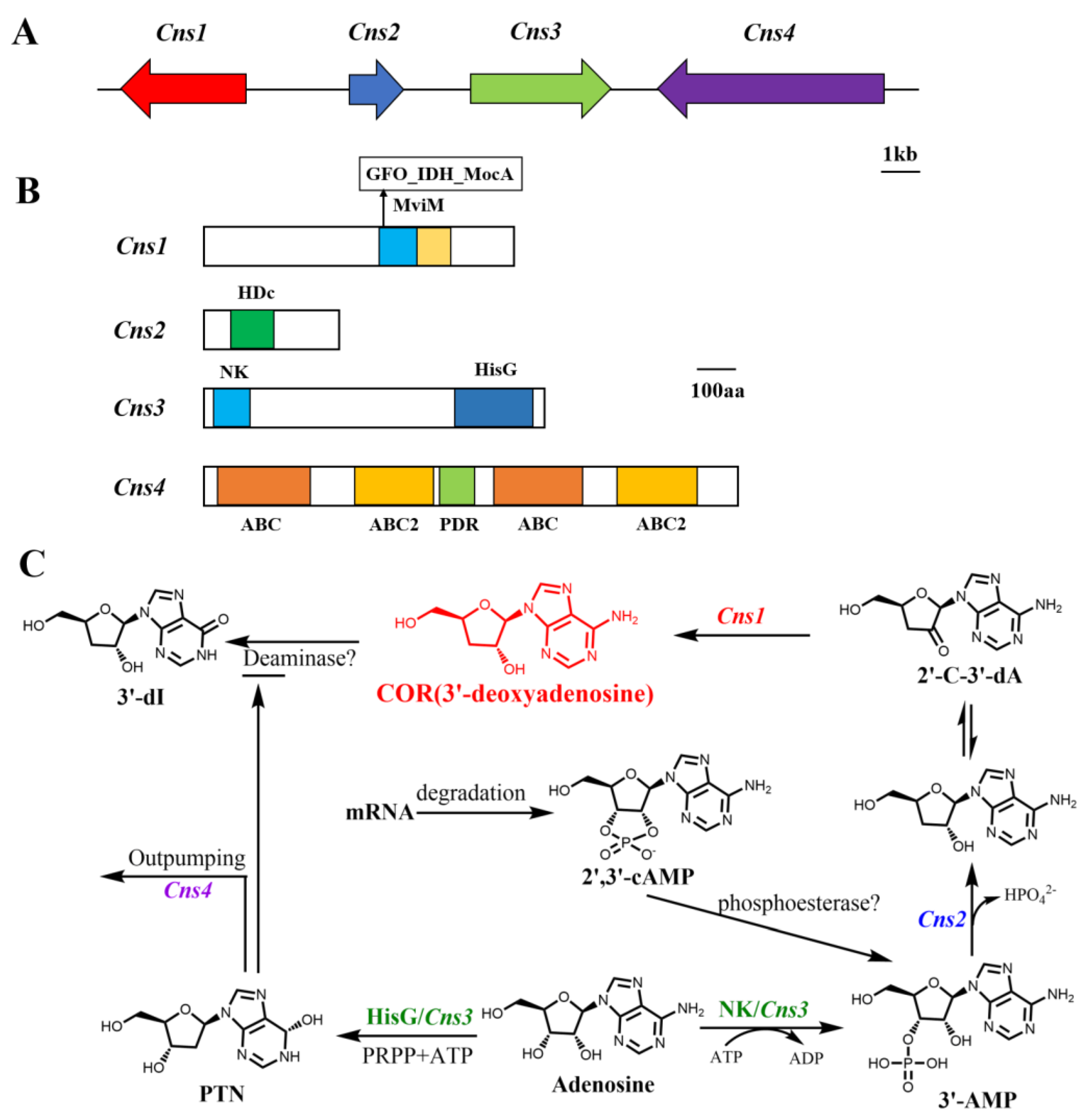
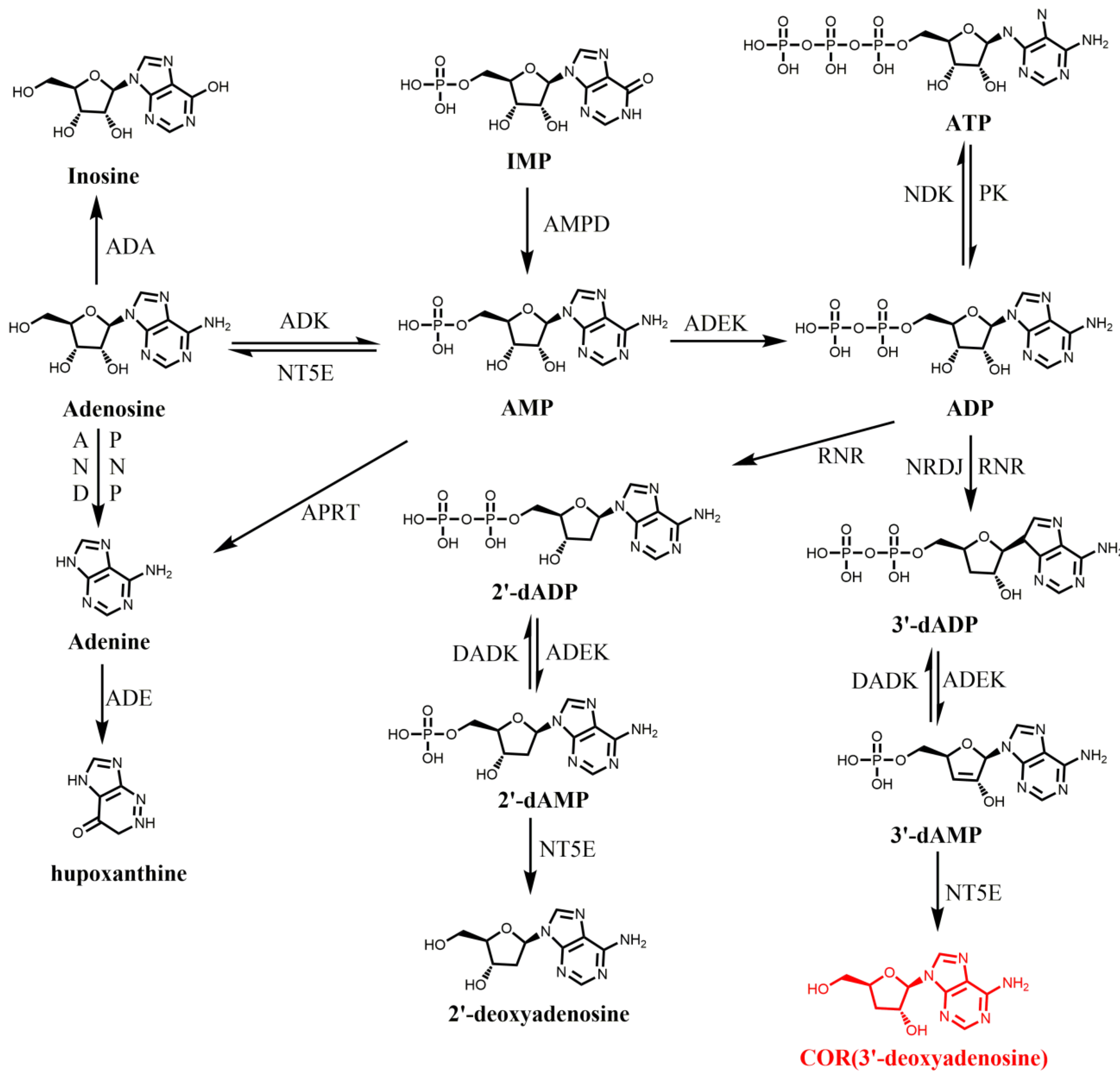
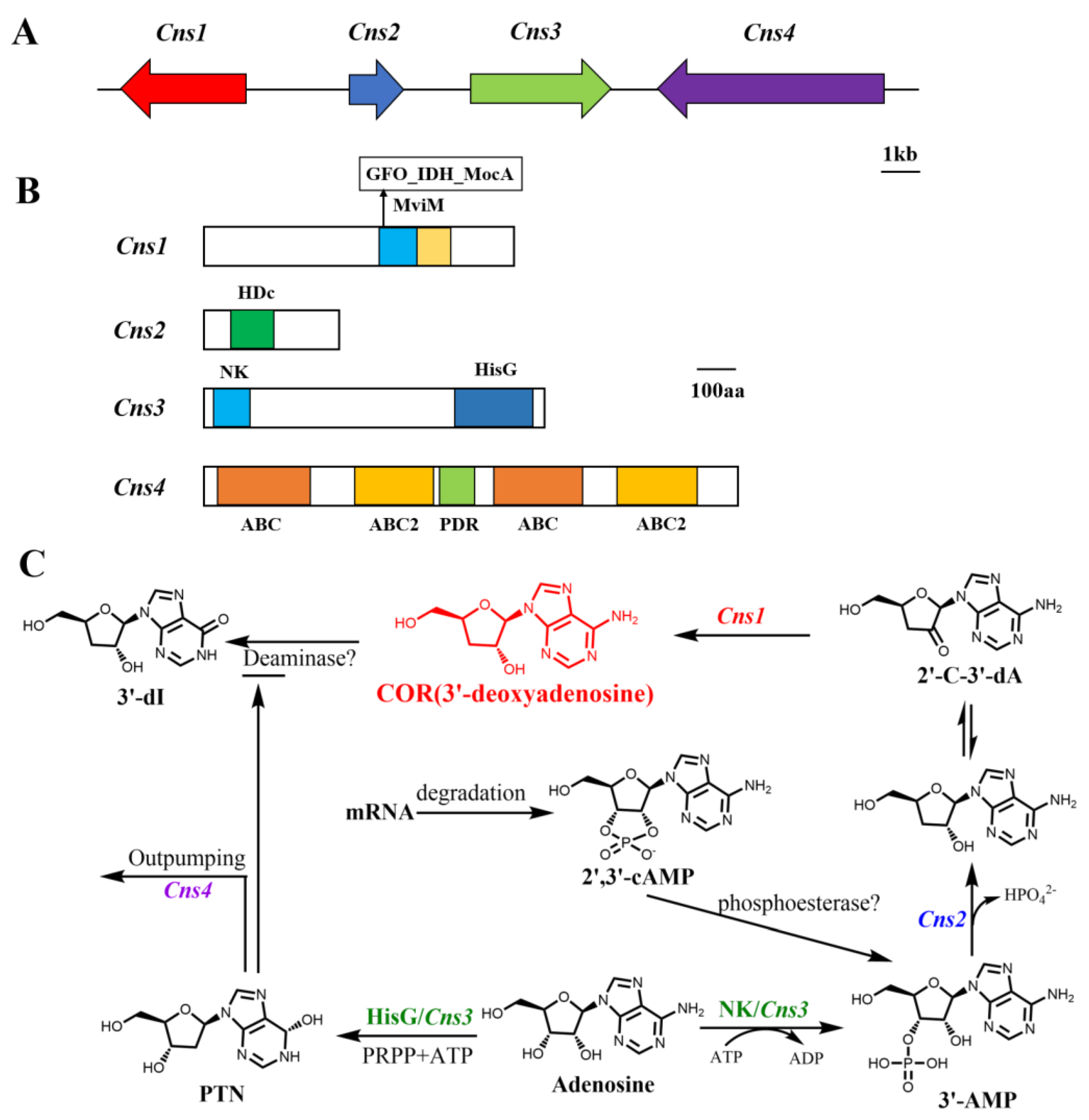
3. Biosynthesis of COR
4. Liquid Fermentation to Produce COR
5. Solid-State Fermentation to Produce COR
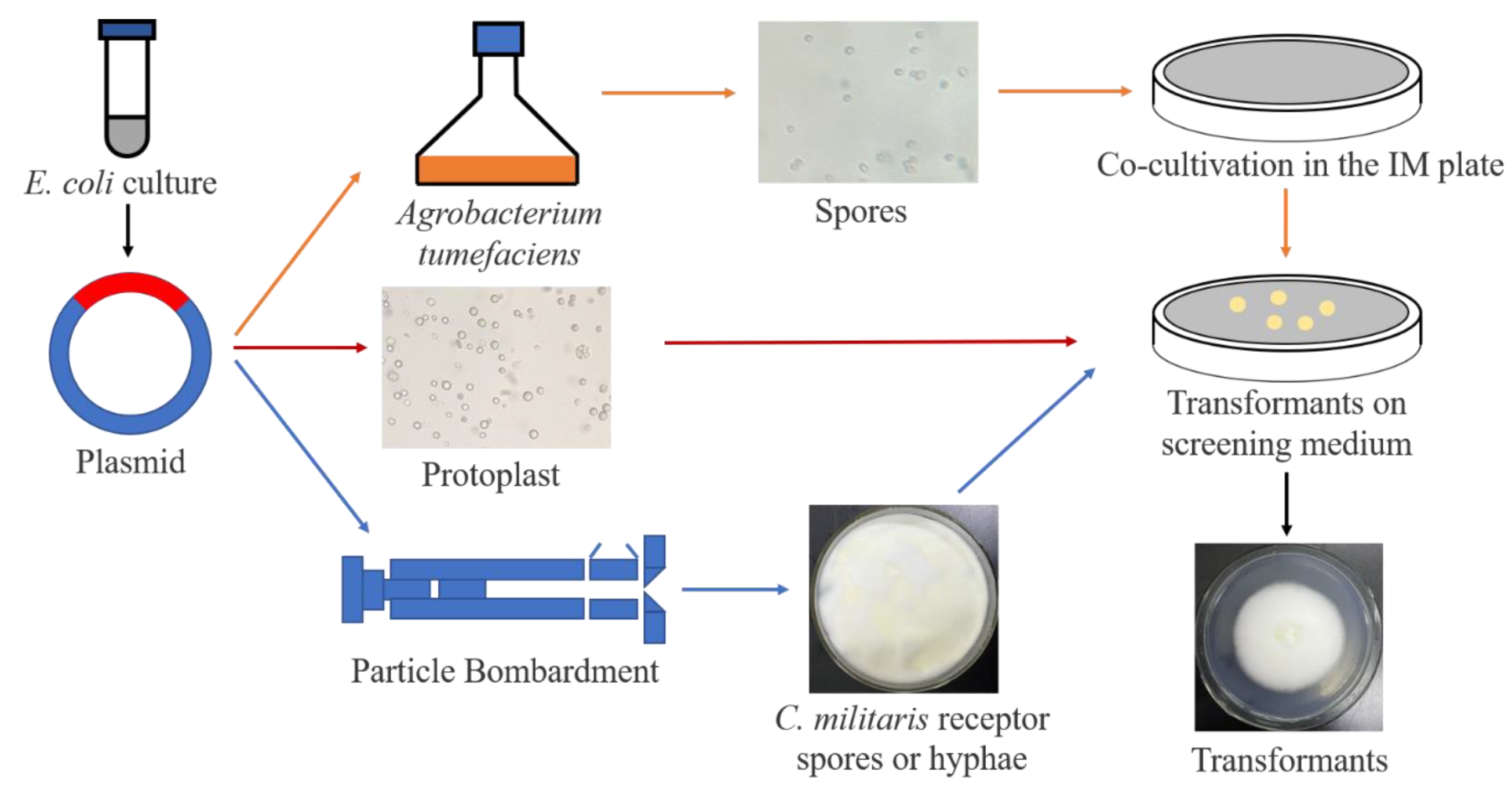
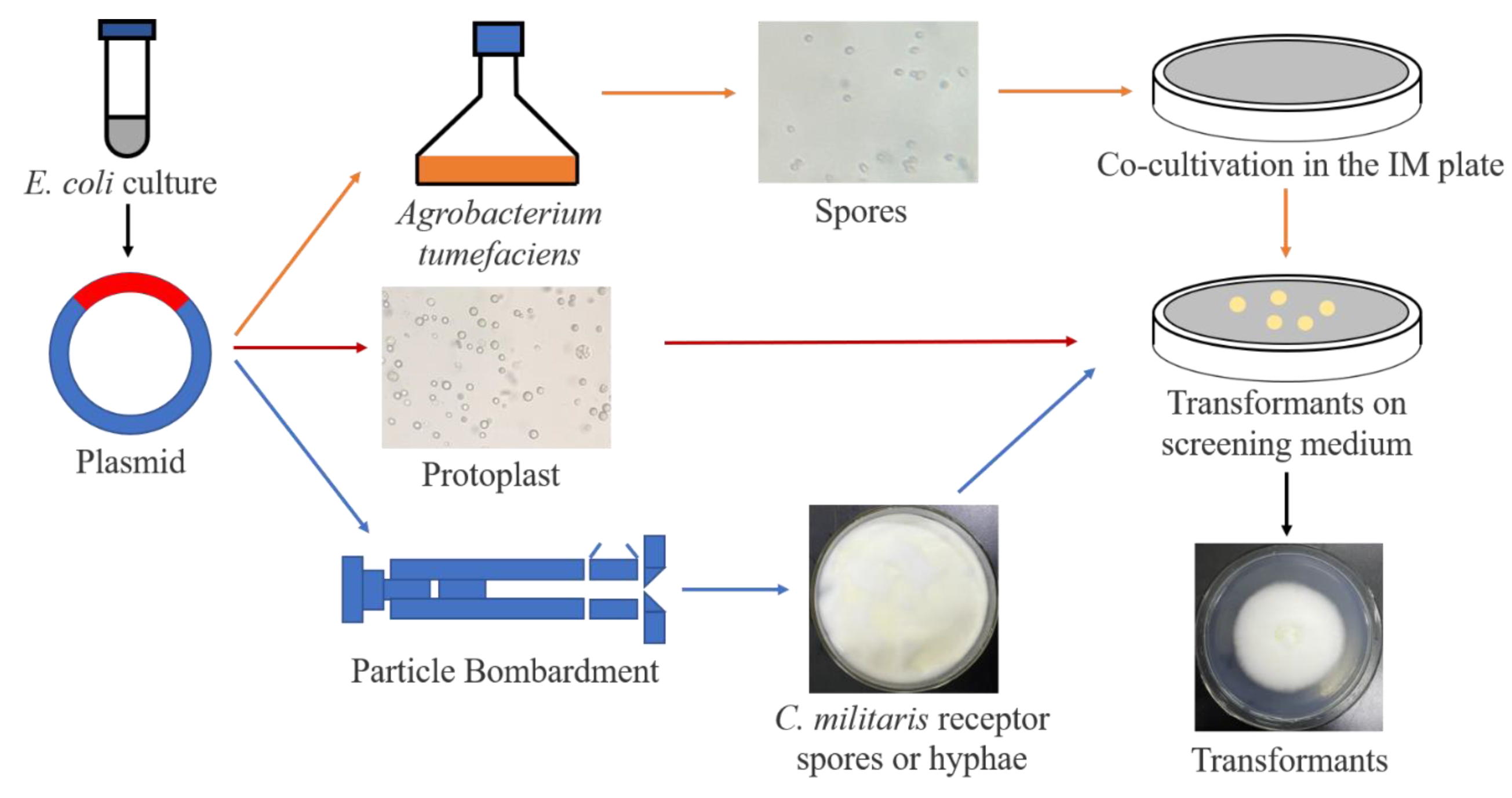
6. Genetic Modification of C. militaris
7. Genetic Modification of C. militaris to Increase COR Content
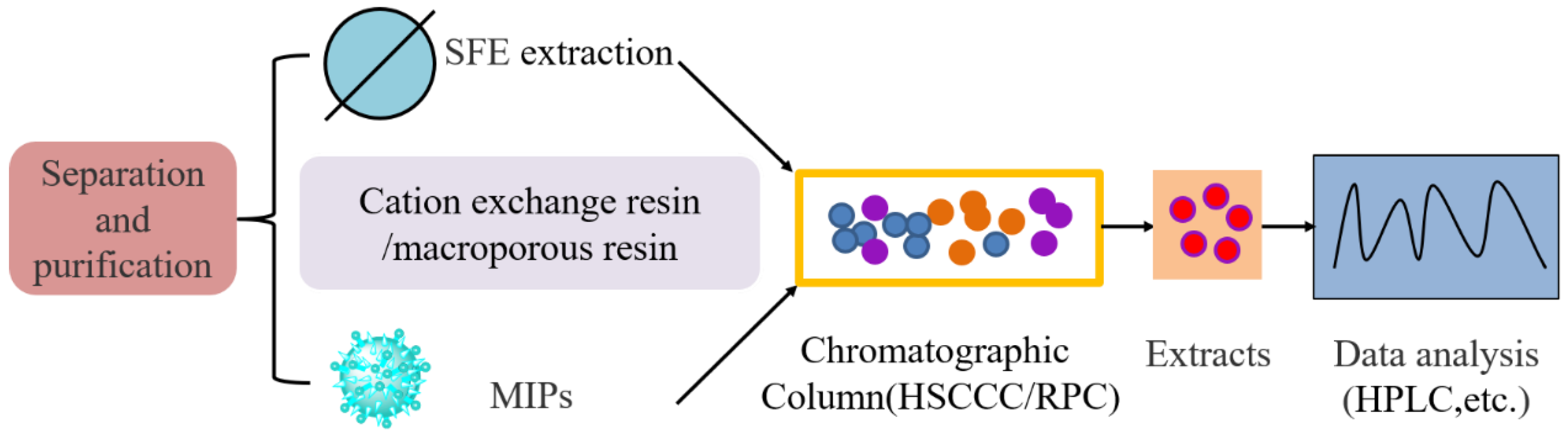
8. Separation and Purification of COR
9. Conclusions and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, F.; Zhou, L.W.; Yang, Z.L.; Bau, T.; Dai, Y.C. Resource diversity of Chinese macrofungi: Edible, medicinal and poisonous species. Fungal Divers. 2019, 98, 1–76. [Google Scholar] [CrossRef]
- Tian, L.H.; Hu, B.; Zhou, H.; Zhang, W.M.; Qu, L.H.; Chen, Y.Q. Molecular phylogeny of the entomopathogenic fungi of the genus Cordyceps (Ascomycota: Clavicipitaceae) and its evolutionary implications. J. Syst. Evol. 2010, 6, 47–56. [Google Scholar] [CrossRef]
- Wei, H.P.; Ye, X.L.; Chen, Z.; Zhong, Y.J.; Li, P.M.; Pu, S.C.; Li, X.G. Synthesis and pharmacokinetic evaluation of novel N-acyl-cordycepin derivatives with a normal alkyl chain. Eur. J. Med. Chem. 2009, 44, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.C.; Kim, K.D.; Chan, H.B.; Ju, C.K.; Kim, D.K.; Kim, H.H. A mushroom lectin from ascomycete Cordyceps militaris. Biochim. Biophys. Acta Gen. Subj. 2007, 1770, 833–838. [Google Scholar] [CrossRef]
- Quy, T.N.; Xuan, T.D.; Andriana, Y.; Tran, H.D.; Khanh, T.D.; Teschke, R. Cordyceps militaris Cordycepin Isolated from: Its Newly Discovered Herbicidal Property and Potential Plant-Based Novel Alternative to Glyphosate. Molecules 2019, 24, 2901. [Google Scholar] [CrossRef] [Green Version]
- Lan, D.H.; Thu, N.; Lan, P.T.; Nha, P.V.; Tung, B.T. Cordyceps militaris (L.) Link: Chemical Bioactive Compounds and Pharmacological Activities. J. Pharm. Nutr. Sci. 2016, 6, 153–159. [Google Scholar]
- Yue, K.; Ye, M.; Zhou, Z.; Sun, W.; Lin, X. The genus Cordyceps: A chemical and pharmacological review. J. Pharm. Pharmacol. 2012, 65, 474–493. [Google Scholar] [CrossRef]
- Woolley, V.C.; Teakle, G.R.; Prince, G.; de Moor, C.H.; Chandler, D. Cordycepin, a metabolite of Cordyceps militaris, reduces immune-related gene expression in insects. J. Invertebr. Pathol. 2020, 177, 107480. [Google Scholar] [CrossRef]
- Lin, L.-T.; Lai, Y.-J.; Wu, S.-C.; Hsu, W.-H.; Tai, C.-J. Optimal conditions for cordycepin production in surface liquid-cultured Cordyceps militaris treated with porcine liver extracts for suppression of oral cancer. J. Food Drug Anal. 2018, 26, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Ahn, Y.J.; Park, S.J.; Lee, S.G.; Shin, S.C.; Choi, D.H. Cordycepin: Selective Growth Inhibitor Derived from Liquid Culture of Cordyceps militaris against Clostridium spp. J. Agric. Food Chem. 2000, 48, 2744–2748. [Google Scholar] [CrossRef]
- Cunningham, K.G.; Manson, W.; Spring, F.S.; Hutchinson, S.A. Cordycepin, a Metabolic Product isolated from Cultures of Cordyceps militaris (Linn.) Link. Nature 1950, 166, 949-949. [Google Scholar] [CrossRef] [PubMed]
- Kaczka, E.A.; Trenner, N.R.; Arison, B.; Walker, R.W.; Folkers, K. Identification of cordycepin, a metabolite of Cordyceps militaris, as 3’-deoxyadenosine. Biochem. Biophys. Res. Commun. 1964, 14, 456–457. [Google Scholar] [CrossRef]
- Kaczka, E.A.; Dulaney, E.L.; Gitterman, C.O.; Woodruff, H.B.; Folkers, K. Isolation and inhibitory effects of KB cell cultures of 3’-deoxyandenosine from Aspergillus nidulans (Eidam) Wint. Biochem. Biophys. Res. Commun. 1964, 14, 452–455. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, G.; Li, C.; Ling, J. Cordycepin and pentostatin biosynthesis gene identified through transcriptome and proteomics analysis of Cordyceps kyushuensis Kob. Microbiol. Res. 2019, 218, 12–21. [Google Scholar] [CrossRef]
- Liu, T.; Liu, Z.; Yao, X.; Huang, Y.; Qu, Q.; Shi, X.; Zhang, H.; Shi, X. Identification of cordycepin biosynthesis-related genes through de novo transcriptome assembly and analysis in Cordyceps cicadae. R. Soc. Open Sci. 2018, 5, 181247. [Google Scholar] [CrossRef] [Green Version]
- Tuli, H.S.; Kashyap, D.; Sharma, A.K. Cordycepin: A Cordyceps Metabolite with Promising Therapeutic Potential; Springer International Publishing: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Tuli, H.S.; Sharma, A.K.; Sandhu, S.S.; Kashyap, D. Cordycepin: A bioactive metabolite with therapeutic potential. Life Sci. 2013, 93, 863–869. [Google Scholar] [CrossRef]
- Tuli, H.S.; Sandhu, S.S.; Sharma, A.K. Pharmacological and therapeutic potential of Cordyceps with special reference to Cordycepin. Biotech 2014, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Xue, M.; Qiu, Z.; Su, Y.; Yu, P.; Peng, Q. Anti-tumor and anti-metastatic roles of cordycepin, one bioactive compound of Cordyceps militaris. Saudi J. Biol. Sci. 2018, 25, 991–995. [Google Scholar] [CrossRef]
- Zhou, X.; Meyer, C.U.; Schmidtke, P.; Zepp, F. Effect of cordycepin on interleukin-10 production of human peripheral blood mononuclear cells. Eur. J. Pharmacol. 2002, 453, 309–317. [Google Scholar] [CrossRef]
- Jeong, M.H.; Lee, C.M.; Lee, S.W.; Seo, S.Y.; Seo, M.J.; Kang, B.W.; Jeong, Y.K.; Choi, Y.J.; Yang, K.M.; Jo, W.S. Cordycepin-enriched Cordyceps militaris induces immunomodulation and tumor growth delay in mouse-derived breast cancer. Oncol. Rep. 2013, 30, 1996–2002. [Google Scholar] [CrossRef] [Green Version]
- Cao, T.; Xu, R.; Xu, Y.; Liu, Y.; Wan, Q. The protective effect of Cordycepin on diabetic nephropathy through autophagy induction in vivo and in vitro. Int. Urol. Nephrol. 2019, 51, 1883–1892. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zhang, S.; Du, M. Cordycepin from Cordyceps militaris prevents hyperglycemia in alloxan-induced diabetic mice. Nutr. Res. 2015, 35, 431–439. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Li, Y.; Wang, X.; Chen, Z.; Xu, H.; Wu, L.; Li, S.; Wang, C.; Luan, W.; Wang, X. Cordycepin reduces weight through regulating gut microbiota in high-fat diet-induced obese rats. Lipids Health Dis. 2018, 17, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahy, B.W.J.; Cox, N.J.; Armstrong, S.J.; Barry, R.D. Multiplication of Influenza Virus in the Presence of Cordycepin, an Inhibitor of Cellular RNA Synthesis. Nat. New Biol. 1973, 243, 172–174. [Google Scholar] [CrossRef] [PubMed]
- White, J.L.; Dawson, W.O. Effect of cordycepin triphosphate on in vitro RNA synthesis by plant viral replicases. J. Virol. 1979, 29, 811–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, W.E.; Weiler, B.E.; Charubala, R.; Pfleiderer, W.; Leserman, L.; Sobol, R.W.; Suhadolnik, R.J.; Schröder, H.C. Cordycepin analogues of 2′,5′-oligoadenylate inhibit human immunodeficiency virus infection via inhibition of reverse transcriptase. Biochemistry 1991, 30, 2027–2033. [Google Scholar] [CrossRef]
- Lonai, P.; Declève, A.; Kaplan, H.S. Spontaneous induction of endogenous murine leukemia virus-related antigen expression during short-term in vitro incubation of mouse lymphocytes. Proc. Natl. Acad. Sci. USA 1974, 71, 2008–2012. [Google Scholar] [CrossRef] [Green Version]
- Doetsch, P.W.; Suhadolnik, R.J.; Sawada, Y.; Mosca, J.D.; Flick, M.B.; Reichenbach, N.L.; Dang, A.Q.; Wu, J.M.; Charubala, R.; Pfleiderer, W.; et al. Core (2′-5′)oligoadenylate and the cordycepin analog: Inhibitors of Epstein--Barr virus-induced transformation of human lymphocytes in the absence of interferon. Proc. Natl. Acad. Sci. USA 1981, 78, 6699–6703. [Google Scholar] [CrossRef] [Green Version]
- de Julián-Ortiz, J.V.; Gálvez, J.; Muñoz-Collado, C.; García-Domenech, R.; Gimeno-Cardona, C. Virtual combinatorial syntheses and computational screening of new potential anti-herpes compounds. J. Med. Chem. 1999, 42, 3308–3314. [Google Scholar] [CrossRef]
- Ryu, E.; Son, M.; Lee, M.; Lee, K.; Cho, J.Y.; Cho, S.; Lee, S.K.; Lee, Y.M.; Cho, H.; Sung, G.H. Cordycepin is a novel chemical suppressor of Epstein-Barr virus replication. Oncoscience 2014, 1, 866. [Google Scholar] [CrossRef] [Green Version]
- Pridgen, C.L. Influenza virus RNA’s in the cytoplasm of chicken embryo cells treated with 3′-deoxyadenosine. J. Virol. 1976, 18, 356–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Q.; Lou, Z.; Wang, H.; Chen, C. Antimicrobial effect and proposed action mechanism of cordycepin against Escherichia coli and Bacillus subtilis. J. Microbiol. 2019, 57, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Sugar, A.M.; Mccaffrey, R.P. Antifungal Activity of 3′-Deoxyadenosine (Cordycepin). Antimicrob. Antimicrob. Agents Chemother. 1998, 42, 1424–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.J.; Cho, J.Y.; Man, H.R.; Park, H.J. Cordycepin (3′-deoxyadenosine) inhibits human platelet aggregation in a cyclic AMP- and cyclic GMP-dependent manner. Eur. J. Pharmacol. 2007, 558, 43–51. [Google Scholar] [CrossRef]
- Lei, J.; Wei, Y.; Song, P.; Li, Y.; Xu, G. Cordycepin inhibits LPS-induced acute lung injury by inhibiting inflammation and oxidative stress. Eur. J. Pharmacol. 2017, 818, 110. [Google Scholar] [CrossRef] [PubMed]
- Fei, X.; Zhang, X.; Zhang, G.Q.; Bao, W.P.; Zhang, Y.Y.; Zhang, M.; Zhou, X. Cordycepin inhibits airway remodeling in a rat model of chronic asthma. Biomed. Pharmacother. 2017, 88, 335. [Google Scholar] [CrossRef]
- Tan, L.; Song, X.; Ren, Y.; Wang, M.; Guo, C.; Guo, D.; Gu, Y.; Li, Y.; Cao, Z.; Deng, Y. Anti-inflammatory effects of cordycepin: A review. Phytother. Res. 2021, 35, 1284–1297. [Google Scholar] [CrossRef]
- Lee, Y.R.; Noh, E.M.; Jeong, E.Y.; Yun, S.K.; Jeong, Y.J.; Kim, J.H.; Kwon, K.B.; Kim, B.S.; Lee, S.H.; Park, C.S. Cordycepin inhibits UVB-induced matrix metalloproteinase expression by suppressing the NF-kappaB pathway in human dermal fibroblasts. Exp. Mol. Med. 2009, 41, 548–554. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Park, S.; Kim, Y.; Park, G.; Son, H.; Lee, S. Suppression of α-MSH and IBMX-induced melanogenesis by cordycepin via inhibition of CREB and MITF, and activation of PI3K/Akt and ERK-dependent mechanisms. Int. J. Mol. Med. 2012, 29, 119–124. [Google Scholar]
- Teng, X.; Zhang, M.; Bhandri, B. 3D printing of Cordyceps flower powder. J. Food Process Eng. 2019, 42, e13179. [Google Scholar] [CrossRef]
- Kunhorm, P.; Chaicharoenaudomrung, N.; Noisa, P. Enrichment of cordycepin for cosmeceutical applications: Culture systems and strategies. Appl. Microbiol. Biotechnol. 2019, 103, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Yoon, D.H.; Shrestha, B.; Choi, H.K.; Sung, G.H. Metabolomic profiling reveals enrichment of cordycepin in senescence process of Cordyceps militaris fruit bodies. J. Microbiol. 2019, 57, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Pang, F.; Wang, L.; Jin, Y.; Guo, L.; Song, L.; Liu, G.; Feng, C. Transcriptome analysis of Paecilomyces hepiali at different growth stages and culture additives to reveal putative genes in cordycepin biosynthesis. Genomics 2018, 110, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Zhong, J. Hyperproduction of Cordycepin by Two-Stage Dissolved Oxygen Control in Submerged Cultivation of Medicinal Mushroom Cordyceps militaris in Bioreactors. Biotechnol. Prog. 2010, 20, 1408–1413. [Google Scholar] [CrossRef] [PubMed]
- Kodama, E.N.; McCaffrey, R.P.; Yusa, K.; Mitsuya, H. Antileukemic activity and mechanism of action of cordycepin against terminal deoxynucleotidyl transferase-positive (TdT+) leukemic cells. Biochem. Pharmacol. 2000, 59, 273–281. [Google Scholar] [CrossRef]
- Qin, P.; Li, X.; Yang, H.; Wang, Z.Y.; Lu, D. Therapeutic Potential and Biological Applications of Cordycepin and Metabolic Mechanisms in Cordycepin-Producing Fungi. Molecules 2019, 24, 2231. [Google Scholar] [CrossRef] [Green Version]
- Sir, T.A.; Ulbricht, T.L.V. Deoxynucleosides and related compounds. Part IX. A synthesis of 3′-deoxyadenosine. J. Chem. Soc. 1960, 82, 3275–3277. [Google Scholar]
- Hansske, F.; Robins, M.J. Regiospecific and stereoselective conversion of ribonucleosides to 3′-deoxynucleosides. A high yield three-stage synthesis of cordycepin from adenosine. Tetrahedron Lett. 1985, 26, 4295–4298. [Google Scholar] [CrossRef]
- Meier, C.; Huynh-Dinh, T. Improved Conversion of Adenosine to 3′-Deoxyadenosine. Synlett 1991, 4, 227–228. [Google Scholar] [CrossRef]
- Meier, C.; Tam, H.D. ChemInform Abstract: Improved Conversion of Adenosine to 3′-Deoxyadenosine. ChemInform 1992, 23, 1. [Google Scholar] [CrossRef]
- Aman, S.; Anderson, D.J.; Connolly, T.J.; Crittall, A.J.; Ji, G. From Adenosine to 3′-Deoxyadenosine: Development and Scale Up. Org. Process Res. Dev. 2000, 4, 601–605. [Google Scholar] [CrossRef]
- Huang, S.; Liu, H.; Sun, Y.; Chen, J.; Li, X.; Xu, J.; Hu, Y.; Li, Y.; Deng, Z.; Zhong, S. An effective and convenient synthesis of cordycepin from adenosine. Chem. Pap. 2018, 72, 149–160. [Google Scholar] [CrossRef]
- Mcdonald, F.E.; Gleason, M.M. Asymmetric Syntheses of Stavudine(d4T) and Cordycepin by Cycloisomerization of Alkynyl Alcohols to Endocyclic Enol Ethers. Angew. Chem. Int. Ed. Engl. 1995, 34, 350–352. [Google Scholar] [CrossRef]
- Li, Q.; Yang, R.; Ruan, Z.; Tao, H.; Qiang, X. Total Synthesis of Cordycepin. Chin. J. Org. Chem. 2013, 33, 1340. [Google Scholar] [CrossRef] [Green Version]
- Kredich, N.M.; Guarino, A.J. Studies on the biosynthesis of cordycepin. Biochim. Biophys. Acta 1961, 47, 499–535. [Google Scholar] [CrossRef] [Green Version]
- Suhadolnik, R.J.; Cory, J.G. Further evidence for the biosynthesis of cordycepin and proof of the structure of 3-deoxyribose. Biochim. Et Biophys. Acta 1964, 91, 661–662. [Google Scholar] [CrossRef]
- Lennon, M.B.; Suhadolnik, R.J. Biosynthesis of 3′-deoxyadenosine by Cordyceps militaris. Mechanism of reduction. Biochim. Et Biophys. Acta 1976, 425, 532–536. [Google Scholar] [CrossRef]
- Masuda, M.; Urabe, E.; Honda, H.; Sakurai, A.; Sakakibara, M. Enhanced production of cordycepin by surface culture using the medicinal mushroom Cordyceps militaris. Enzym. Microb. Technol. 2007, 39, 641–646. [Google Scholar] [CrossRef]
- Zheng, P.; Xia, Y.; Xiao, G.; Xiong, C.; Hu, X.; Zhang, S.; Zheng, H.; Huang, Y.; Zhou, Y.; Wang, S.; et al. Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional Chinese medicine. Genome Biol. 2011, 12, R116. [Google Scholar] [CrossRef] [Green Version]
- Xiang, L.; Li, Y.; Zhu, Y.; Luo, H.; Li, C.; Xu, X.; Sun, C.; Song, J.; Shi, L.; He, L. Transcriptome analysis of the Ophiocordyceps sinensis fruiting body reveals putative genes involved in fruiting body development and cordycepin biosynthesis. Genomics 2014, 103, 154–159. [Google Scholar] [CrossRef]
- Lin, S.; Liu, Z.Q.; Xue, Y.P.; Baker, P.J.; Wu, H.; Xu, F.; Teng, Y.; Brathwaite, M.E.; Zheng, Y.G. Biosynthetic Pathway Analysis for Improving the Cordycepin and Cordycepic Acid Production in Hirsutella sinensis. Appl. Biochem. Biotechnol. 2016, 179, 633–649. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Luo, F.; Shang, Y.; Chen, P.; Lu, Y.; Wang, C. Fungal Cordycepin Biosynthesis Is Coupled with the Production of the Safeguard Molecule Pentostatin. Cell Chem. Biol. 2017, 24, 1479–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wongsa, B.; Raethong, N.; Chumnanpuen, P.; Wong-Ekkabut, J.; Laoteng, K.; Vongsangnak, W. Alternative metabolic routes in channeling xylose to cordycepin production of Cordyceps militaris identified by comparative transcriptome analysis. Genomics 2020, 112, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Jackson, E.K.; Ren, J.; Mi, Z. Extracellular 2′,3′-cAMP Is a Source of Adenosine. J. Biol. Chem. 2009, 284, 33097. [Google Scholar] [CrossRef] [Green Version]
- Yakunin, A.F.; Proudfoot, M.; Kuznetsova, E.; Savchenko, A.; Brown, G.; Arrowsmith, C.H.; Edwards, A.M. The HD Domain of the Escherichia coli tRNA Nucleotidyltransferase Has 2′,3′-Cyclic Phosphodiesterase, 2′-Nucleotidase, and Phosphatase Activities. J. Biol. Chem. 2004, 279, 36819–36827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.K.; Masuda, M.; Sakurai, A.; Sakakibara, M. Medicinal uses of the mushroom Cordyceps militaris: Current state and prospects. Fitoterapia 2010, 81, 961–968. [Google Scholar] [CrossRef]
- Kang, N.; Lee, H.H.; Park, I.; Seo, Y.S. Development of High Cordycepin-Producing Cordyceps militaris Strains. Mycobiology 2017, 45, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Das, S.K.; Masuda, M.; Hatashita, M.; Sakurai, A.; Sakakibara, M. Optimization of culture medium for cordycepin production using Cordyceps militaris mutant obtained by ion beam irradiation. Process Biochem. 2010, 45, 129–132. [Google Scholar] [CrossRef]
- Masuda, M.; Das, S.K.; Fujihara, S.; Hatashita, M.; Sakurai, A. Production of cordycepin by a repeated batch culture of a Cordyceps militaris mutant obtained by proton beam irradiation. J. Biosci. Bioeng. 2011, 111, 55–60. [Google Scholar] [CrossRef]
- Mao, X.B.; Eksriwong, T.; Chauvatcharin, S.; Zhong, J.J. Optimization of carbon source and carbon/nitrogen ratio for cordycepin production by submerged cultivation of medicinal mushroom Cordyceps militaris. Process Biochem. 2005, 40, 1667–1672. [Google Scholar] [CrossRef]
- Shih, I.L.; Tsai, K.L.; Hsieh, C. Effects of culture conditions on the mycelial growth and bioactive metabolite production in submerged culture of Cordyceps militaris. Biochem. Eng. J. 2007, 33, 193–201. [Google Scholar] [CrossRef]
- Hung, Y.P.; Wang, J.J.; Wei, B.L.; Lee, C.L. Effect of the salts of deep ocean water on the production of cordycepin and adenosine of Cordyceps militaris-fermented product. AMB Express 2015, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, D.-D.; Wang, W.; Zhong, J.-J. Enhancement of cordycepin production in submerged cultures of Cordyceps militaris by addition of ferrous sulfate. Biochem. Eng. J. 2012, 60, 30–35. [Google Scholar] [CrossRef]
- Cai, X.; Jin, J.Y.; Zhang, B.; Liu, Z.Q.; Zheng, Y.G. Improvement of cordycepin production by an isolated Paecilomyces hepiali mutant from combinatorial mutation breeding and medium screening. Bioprocess Biosyst. Eng. 2021, 44, 2387–2398. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.J.; Huang, F.K.; Li, Y.S.; Shu-Yao, T. The Quality Improvement of Solid-State Fermentation with Cordyceps militaris by UVB Irradiation. Food Technol. Biotechnol 2017, 55, 445. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Ashraf, S.A.; Khan, S.; Alshammari, E.; Awadelkareem, A.M. Effect of pH, temperature and incubation time on cordycepin production from Cordyceps militaris using solid-state fermentation on various substrates. CyTA-J. Food 2017, 15, 617–621. [Google Scholar] [CrossRef] [Green Version]
- Wen, T.C.; Li, G.R.; Kang, J.C.; Kang, C.; Hyde, K.D. Optimization of solid-state fermentation for fruiting body growth and cordycepin production by Cordyceps militaris. Chiang Mai J. Sci. 2014, 41, 858–872. [Google Scholar]
- Dong, J.Z.; Lei, C.; Zheng, X.J.; Ai, X.R.; Wang, Y.; Wang, Q. Light Wavelengths Regulate Growth and Active Components of Cordyceps militaris Fruit Bodies. J. Food Biochem. 2013, 37, 578–584. [Google Scholar] [CrossRef]
- Chiang, S.S.; Liang, Z.C.; Wang, Y.C.; Liang, C.H. Effect of light-emitting diodes on the production of cordycepin, mannitol and adenosine in solid-state fermented rice by Cordyceps militaris. J. Food Compos. Anal. 2017, 60, 51–56. [Google Scholar] [CrossRef]
- Yang, L.; Li, G.; Chai, Z.; Gong, Q.; Guo, J. Synthesis of cordycepin: Current scenario and future perspectives. Fungal Genet. Biol. 2020, 143, 103431. [Google Scholar] [CrossRef]
- Zheng, Z.; Huang, C.; Li, C.; Xie, C.; Han, R. Agrobacterium tumefaciens-mediated transformation as a tool for insertional mutagenesis in medicinal fungus Cordyceps militaris. Fungal Biol. 2011, 115, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Rachmawati, R.; Kinoshita, H.; Nihira, T. Establishment of Transformation System in Cordyceps militaris by using Integration Vector with Benomyl Resistance Gene. Procedia Environ. Sci. 2013, 17, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Mao, W.J.; Bao, D.P.; Zhou, C.L.; Yan, L.I.; Tan, Q.; Wang, Y. Effective Transformation of Cordyceps militaris by Particle Bombardment. Acta Hortic. Sin. 2015, 42, 1837–1842. [Google Scholar]
- Lou, H.W.; Ye, Z.W.; Yun, F.; Lin, J.F.; Guo, L.Q.; Chen, B.X.; Mu, Z.X. Targeted Gene Deletion in Cordyceps militaris Using the Split-Marker Approach. Mol. Biotechnol. 2018, 60, 380–385. [Google Scholar] [CrossRef]
- Lou, H.W.; Zhao, Y.; Chen, B.X.; Yu, Y.H.; Guo, L.Q. Cmfhp Gene Mediates Fruiting Body Development and Carotenoid Production in Cordyceps militaris. Biomolecules 2020, 10, 410. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.X.; Wei, T.; Ye, Z.W.; Yun, F.; Kang, L.Z.; Tang, H.B.; Guo, L.Q.; Lin, J.F. Efficient CRISPR-Cas9 Gene Disruption System in Edible-Medicinal Mushroom Cordyceps militaris. Front. Microbiol. 2018, 9, 1157. [Google Scholar] [CrossRef]
- Takano, H. The regulatory mechanism underlying light-inducible production of carotenoids in nonphototrophic bacteria. Bioscience Biotechnol. Biochem. 2016, 80, 1264–1273. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Liu, Q.; Zhang, J.; Liu, K.; Li, K.; Liu, G.; Dong, C. Comparative Transcriptome Analysis Between a Spontaneous Albino Mutant and Its Sibling Strain of Cordyceps militaris in Response to Light Stress. Front. Microbiol. 2018, 9, 1237. [Google Scholar] [CrossRef]
- Tao, Y.; Guo, M.; Yang, H.; Guo, S.; Dong, C. The blue-light receptor CmWC-1 mediates fruit body development and secondary metabolism in Cordyceps militaris. Appl. Microbiol. Biotechnol. 2016, 100, 743–755. [Google Scholar]
- Zhang, H.; Wang, Y.; Tong, X.; Yokoyama, W.; Cao, J.; Wang, F.; Peng, C.; Guo, J. Overexpression of ribonucleotide reductase small subunit, RNRM, increases cordycepin biosynthesis in transformed Cordyceps militaris. Chin. J. Nat. Med. 2020, 18, 393–400. [Google Scholar] [CrossRef]
- Lin, P.-J.; Ye, Z.-W.; Wei, T.; Wu, J.-Y.; Zheng, Q.-W.; Chen, B.-X.; Guo, L.-Q.; Lin, J.-F. Cross breeding of novel Cordyceps militaris strains with high contents of cordycepin and carotenoid by using MAT genes as selectable markers. Sci. Hortic. 2021, 290, 110492. [Google Scholar] [CrossRef]
- Chen, B.-X.; Wei, T.; Xue, L.-N.; Zheng, Q.-W.; Ye, Z.-W.; Zou, Y.; Yang, Y.; Yun, F.; Guo, L.-Q.; Lin, J.-F. Transcriptome Analysis Reveals the Flexibility of Cordycepin Network in Cordyceps militaris Activated by L-Alanine Addition. Front. Microbiol. 2020, 11, 577. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.Y.; Zhang, G.Y.; Lin, J.Q.; Cui, Z.J.; Zhang, C.K. Supercritical fluid extraction of cordycepin and adenosine from Cordyceps kyushuensis and purification by high-speed counter-current chromatography. Sep. Purif. Technol. 2009, 66, 625–629. [Google Scholar] [CrossRef]
- Zhu, L.; Liang, Y.; Lao, D.; Zhang, T.; Ito, Y. Preparative separation of high-purity cordycepin from Cordyceps militaris(L.) Link by high-speed countercurrent chromatography. J. Liq. Chromatogr. Relat. Technol. 2011, 34, 491–499. [Google Scholar]
- Zhang, Z.; Tudi, T.; Liu, Y.; Zhou, S.; Feng, N.; Yang, Y.; Tang, C.; Tang, Q.; Zhang, J. Preparative isolation of cordycepin, N6-(2-hydroxyethyl)-adenosine and adenosine from Cordyceps militaris by macroporous resin and purification by recycling high-speed counter-current chromatography. J. Chromatogr. B 2016, 1033–1034, 218–225. [Google Scholar] [CrossRef]
- Zhang, H.C.; Fan, H.T.; Wang, X.J.; Zhang, Z.T.; Yang, J.; Yang, G.W.; Wang, W.B.; Jia, L.I.; Ding, Q.; Hua-Xin, L.I. Purification of cordycepin from fermentation broth of Cordyceps militaris by use of macroporous resin AB-8 and octadecyl bonded silica chromatography. Mycosystema 2015, 34, 490–498. [Google Scholar]
- Zhang, Y.; Wan, J.; Cao, X. Synthesis of surface molecularly imprinting polymers for cordycepin and its application in separating cordycepin. Process Biochem. 2016, 51, 517–527. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Starting Material | Final Product | Yield | (Dis)Advantages | Refs |
|---|---|---|---|---|
1  | 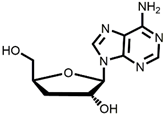 Cordycepin, COR | 14% | Available raw materials and simple procedure with low yield | [48] |
2 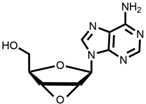 | 90% | High yield but expensive raw materials | [49] | |
3 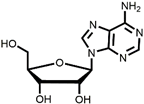 | 56% (90% per step) | High residual tin | [50,51] | |
| 20% | Mild and controllable reaction conditions, high purity of COR; low yield and high cost | [52] | ||
| 13.5% | Simple route with low yield | [53] | ||
| 36% | Acceptable total product yield and commercial availability of all starting materials | |||
4 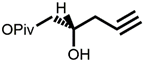 | 14% | Expensive chemical reagents and low yield | [54] | |
5 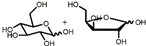 | 37%; 40% | Cheap raw materials and appropriate yield | [55] |
| Strains | Method | COR (g/L) | Increase Rate | Refs |
|---|---|---|---|---|
| KSP8 | Brown rice medium Silk worm pupae medium | 6.63 8.10 | 116% 164% | [68] |
| G81-3 | Medium composition optimization | 6.84 | 179% | [69] |
| Adenosine addition | 8.60 | 251% | [70] | |
| NBRC 9787 | Adenine and glycine | 2.50 | 400% | [59] |
| C. militaris (Hubei) | Carbon and nitrogen sources optimization | 0.35 | 40% | [71] |
| CCRC 32219 | Two-stage shaking-static culture | 2.21 | 112% | [72] |
| C. militaris (Hubei) | FeSO4 addition | 0.59 | 70% | [74] |
| C. militaris BCRC34380 | Porcine liver extracts | 2.45 | 390% | [9] |
| Combined with blue light LED irradiation | 3.48 | 700% | [9] | |
| P. hepiali ZJB18001 | Mutant from 60Co γ-ray and UV irradiation | 0.61 | 134% | [75] |
| Medium composition optimization | 0.96 | 269% | [75] |
| Strains | Method | COR (mg/g) | Increase Rate | Refs |
|---|---|---|---|---|
| C. militaris (Taiwan) | Ultraviolet B (UVB) irradiation | 0.08 | 10% | [76] |
| C. militaris 34164 | Wheat medium | 565.20 | Control | [77] |
| Oat medium | 638.85 | 13% | ||
| Rice medium | 814.60 | 44% | ||
| C. militaris CGMCC2459 | Optimization medium composition | 9.17 | 63% | [78] |
| C. militaris 101 | Fluorescent lamps | 2.89 | Control | [80] |
| 12 h/day illumination | 3.97 | 37% | ||
| LED wavelengths combinations (3R:3B) | 30 | 938% |
| Strains | Method | Selection Marker | Transformation Efficiency | Refs |
|---|---|---|---|---|
| JM4 | Agrobacterium tumefaciens-mediated transformation (ATMT) | Hygromycin B | 30–600 cfu/1 × 105 spores | [82] |
| HF 374-1, HF 438, and CM 001-5 | Protoplast-mediated transformation (PMT) | Benomyl | 7 cfu/μg | [83] |
| CM01 | Particle Bombardment | Basta | 0.4 cfu/µg | [84] |
| CM10 | Split-Marker | Basta | 4.53 cfu/µg | [85] |
| CM10 | CRISPR-Cas9 | Basta | 1.7 cfu/µg | [87] |
| Strains | Genetic Modification | COR (mg/g) | Increase Rate | Refs |
|---|---|---|---|---|
| CGMCC 5.2191 | Albino mutant strain | 6.700 | 117% | [89] |
| C. militaris 40 (CGMCC 3.16322) | CmWC-1c deletion | 0.005 | −58% | [90] |
| C. militaris | The RNR subunit RNRM overexpression | 3.750 | 50% | [91] |
| The RNR subunit RNRL overexpression | 2.600 | 4% | [91] | |
| ZA10-C4 | Hybrid from ZGMM and CM 17 strains | 357.166 | 136–209% | [92] |
| C. militaris CM10 | Overexpression of CmTf1 and CmTf2 | 0.099 | 100% | [93] |
| S. cerevisiae | Overexpression Cns1 and Cns2 | 0.021 | S. cerevisiae can’t synthesis COR | [63] |
| Strains | Method | COR (mg/g) | Purity | Refs |
|---|---|---|---|---|
| C. kyushuensis | supercritical fluid extraction (SFE) and high-speed counter-current chromatography (HSCCC) | 22.3 | 98.5% | [94] |
| C. militaris (Link) | HSCCC and cation-exchange resin | 299.7 | 98.9% | [95] |
| C. militaris (Jiangsu) | Macroporous resin and HSCCC | 31.2 | 98.5% | [96] |
| C. militaris | Silica gel column chromatography and reversed-phase (RPC) | 0.4 | 99% | [97] |
| C. militaris (Shanghai) | Surface imprinting technology | 95.4 | 98% | [98] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Yan, H.; Zeng, B.; Hu, Z. Research Progress on Cordycepin Synthesis and Methods for Enhancement of Cordycepin Production in Cordyceps militaris. Bioengineering 2022, 9, 69. https://doi.org/10.3390/bioengineering9020069
Wang L, Yan H, Zeng B, Hu Z. Research Progress on Cordycepin Synthesis and Methods for Enhancement of Cordycepin Production in Cordyceps militaris. Bioengineering. 2022; 9(2):69. https://doi.org/10.3390/bioengineering9020069
Chicago/Turabian StyleWang, Li, Huanhuan Yan, Bin Zeng, and Zhihong Hu. 2022. "Research Progress on Cordycepin Synthesis and Methods for Enhancement of Cordycepin Production in Cordyceps militaris" Bioengineering 9, no. 2: 69. https://doi.org/10.3390/bioengineering9020069
APA StyleWang, L., Yan, H., Zeng, B., & Hu, Z. (2022). Research Progress on Cordycepin Synthesis and Methods for Enhancement of Cordycepin Production in Cordyceps militaris. Bioengineering, 9(2), 69. https://doi.org/10.3390/bioengineering9020069