
Impact of Electrospun Piezoelectric Core–Shell PVDFhfp/PDMS Mesh on Tenogenic and Inflammatory Gene Expression in Human Adipose-Derived Stem Cells: Comparison of Static Cultivation with Uniaxial Cyclic Tensile Stretching

 , ,
, ,
Abstract
:1. Introduction
- PVDFhfp/PDMS electrospun fibers impact gene expression of human adipose-derived stem cells with an upregulation of tenogenic and downregulation of pro-inflammatory marker genes in supplement-free basal Dulbecco’s modified eagle medium (DMEM).
- Aligned PVDFhfp/PDMS electrospun fibers lead to a higher tenogenic induction than corresponding random electrospun fibers [36].
- Tenogenic gene upregulation is more pronounced after uniaxial cyclic stretching than under mere static cell cultivation [25].
2. Materials and Methods
2.1. Cells
2.2. Multilineage Cell Differentiation
2.3. Scaffolds
2.4. Tissue Engineered Constructs: Cultivation
2.5. Mechanics of Tissue Engineered Constructs
2.6. Gene Expression
2.7. Statistics
3. Results
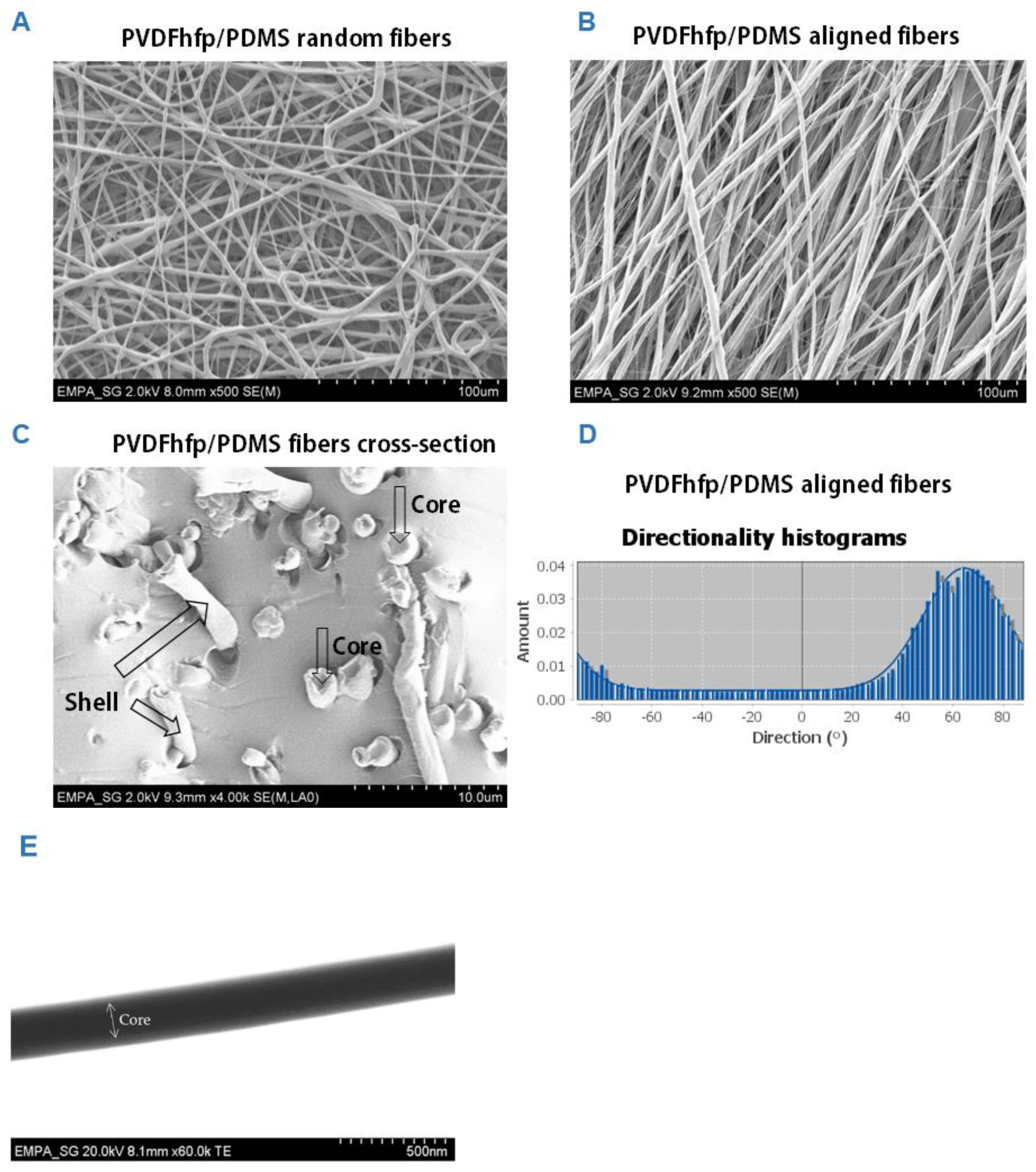
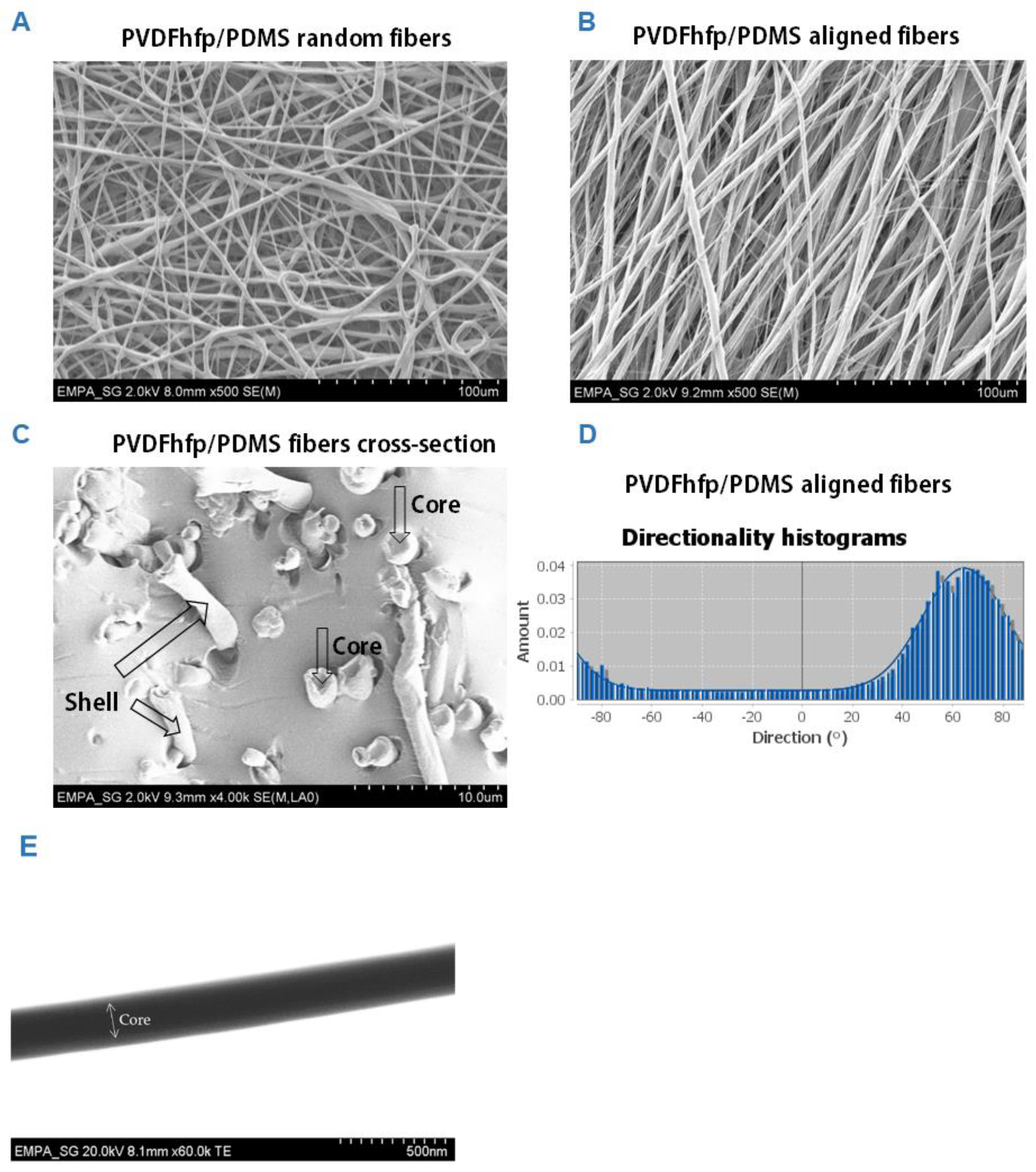
3.1. PVDFhfp/PDMS Electrospun Membranes
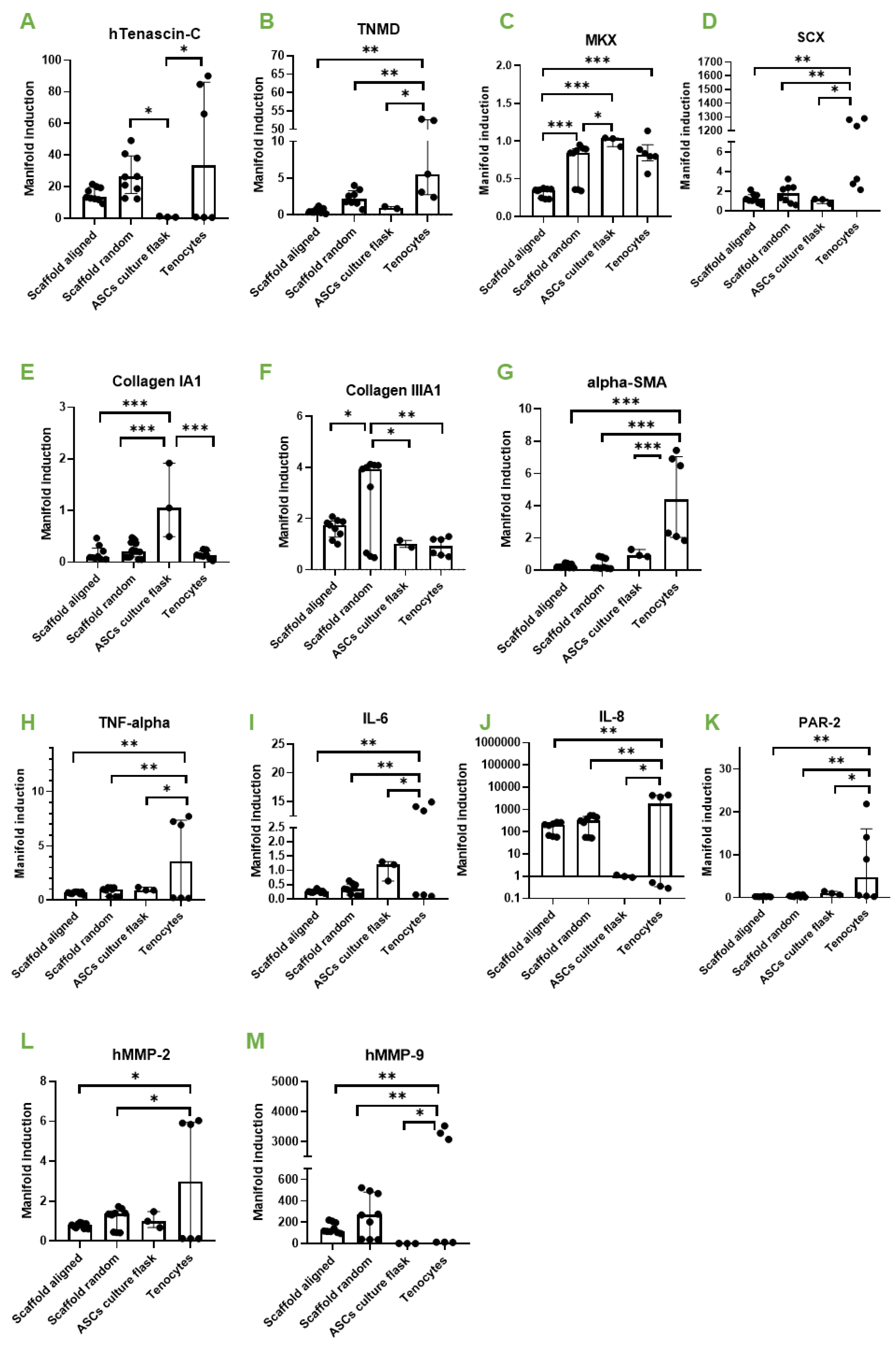
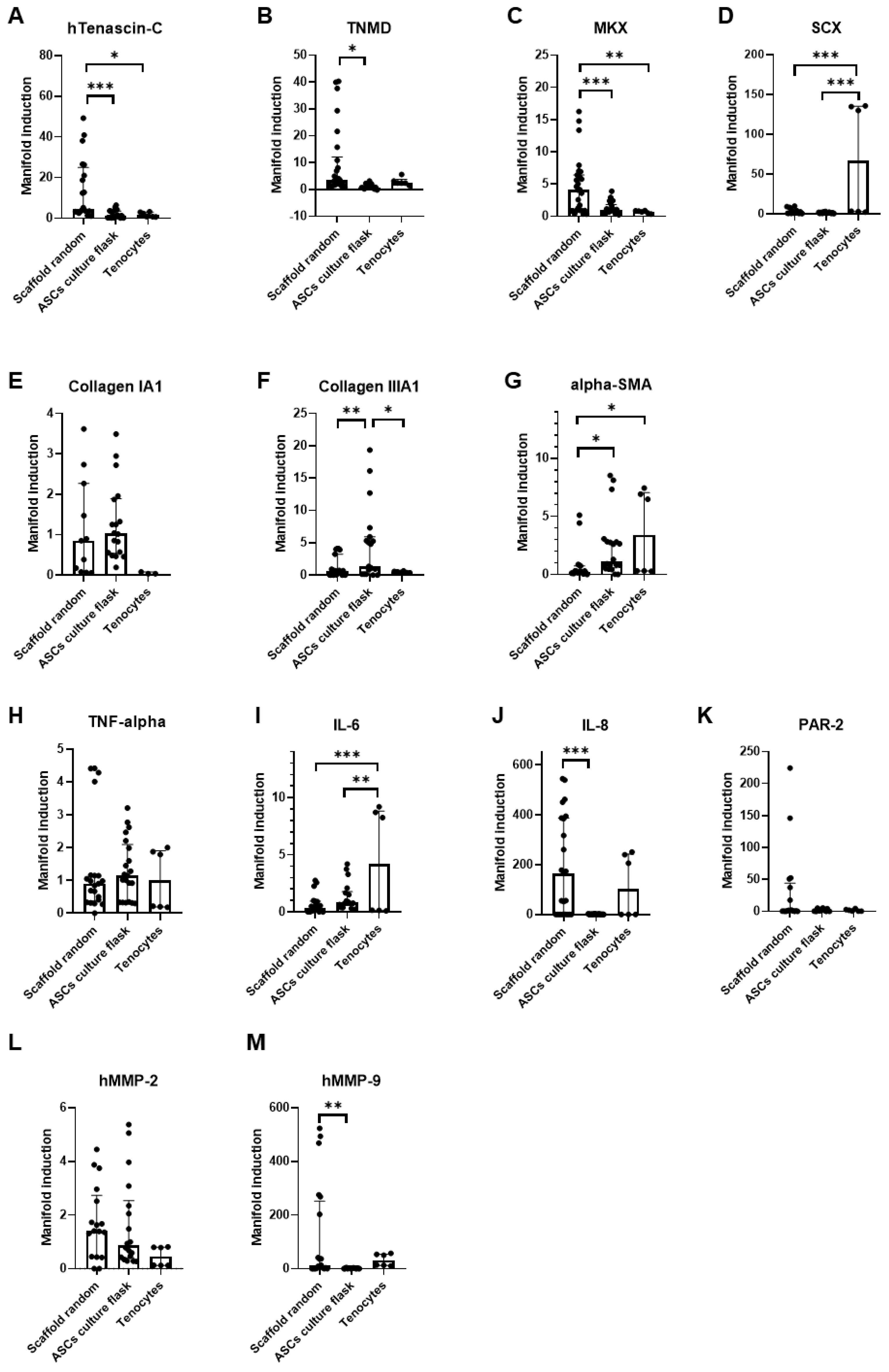
3.2. Random Versus Aligned Scaffolds; Impact on Gene Expression under Static Conditions
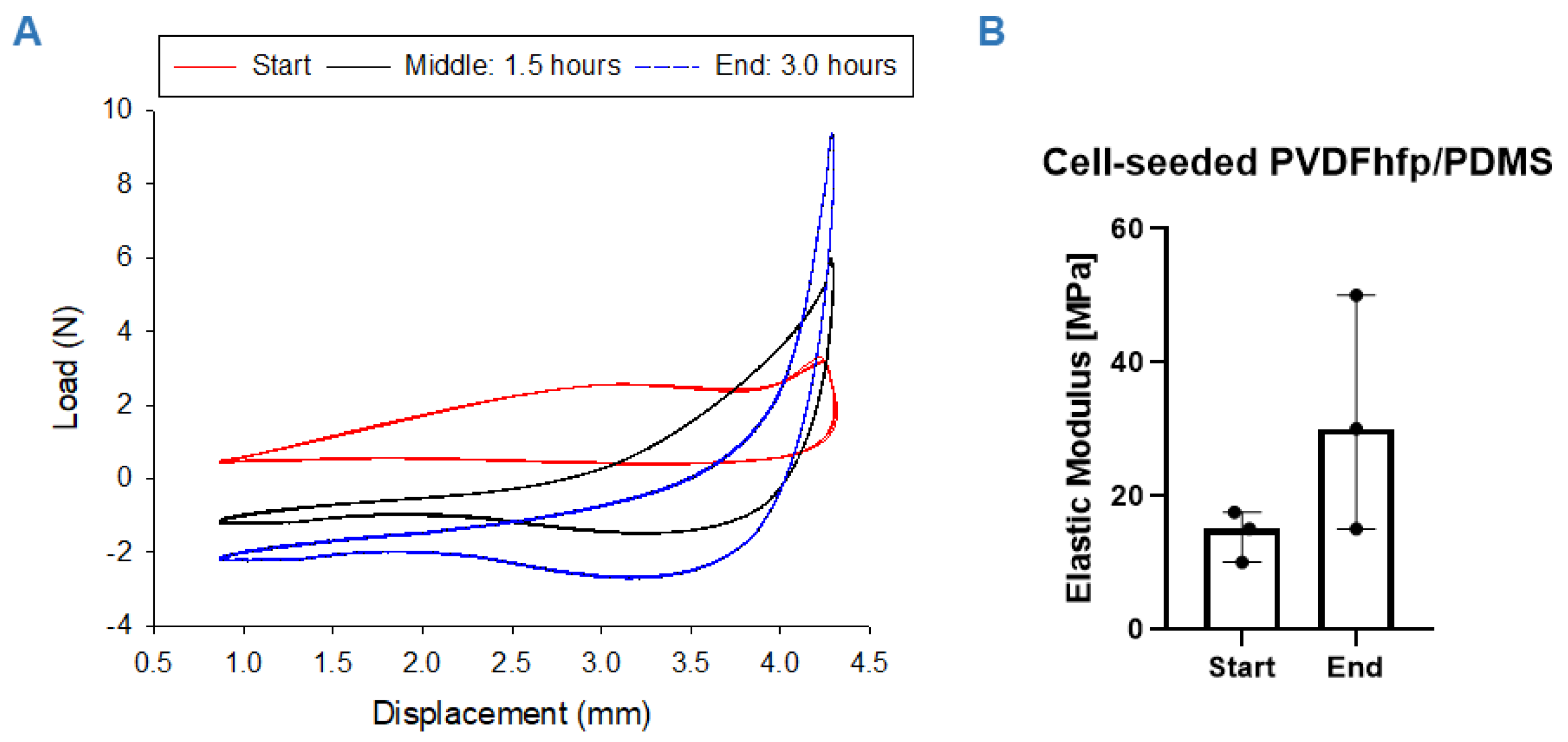
3.3. Mechanical Properties of Tissue-Engineered Constructs
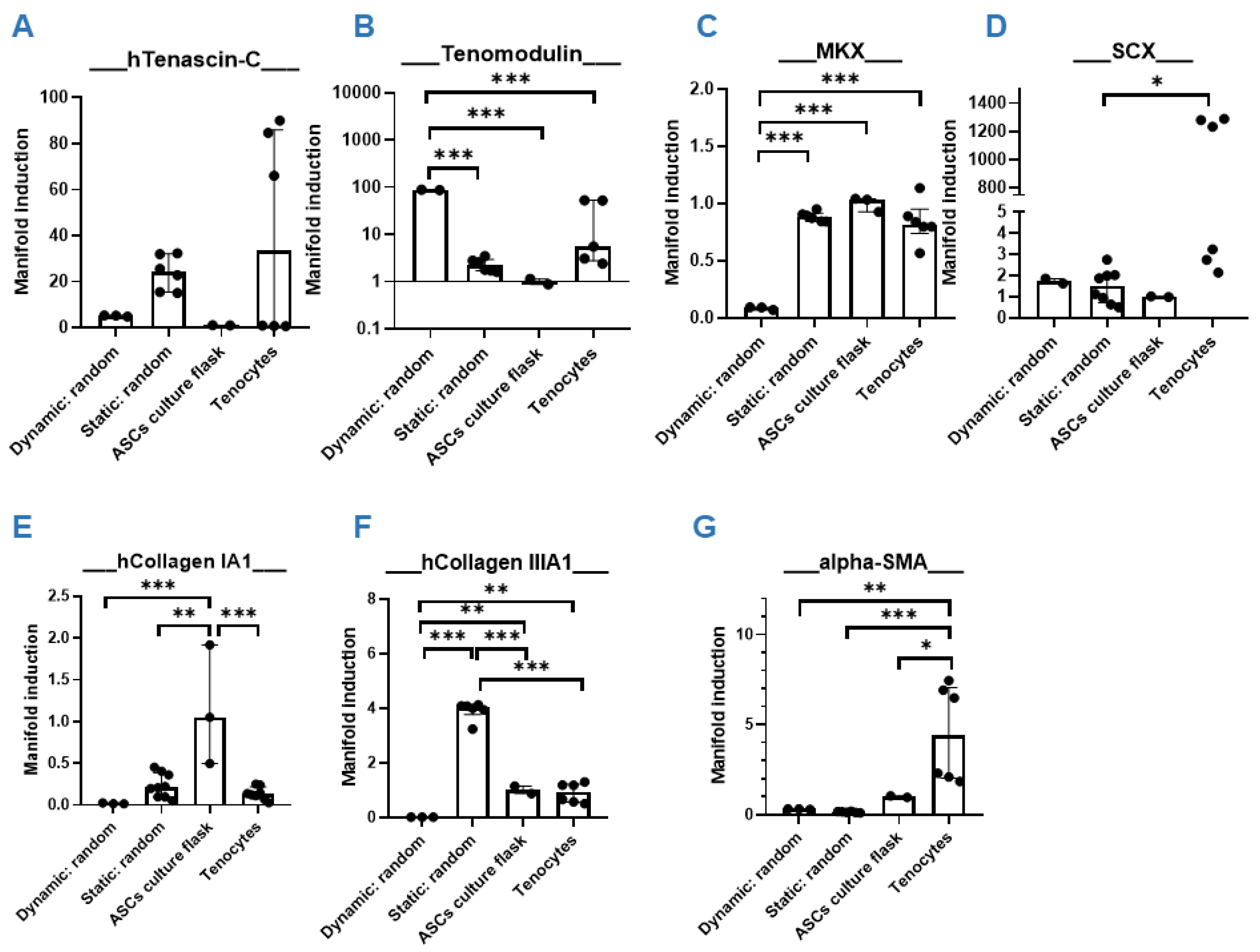
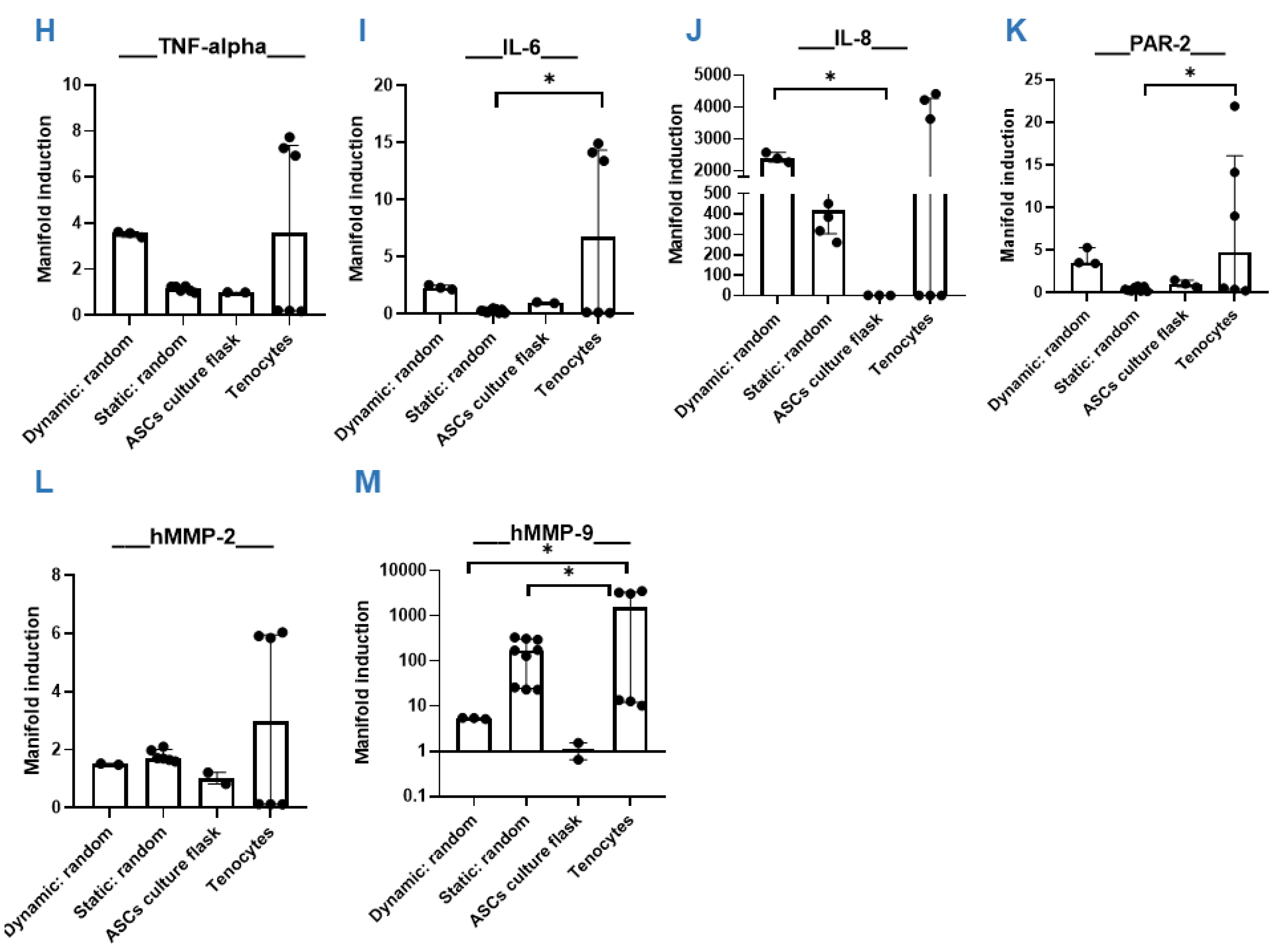
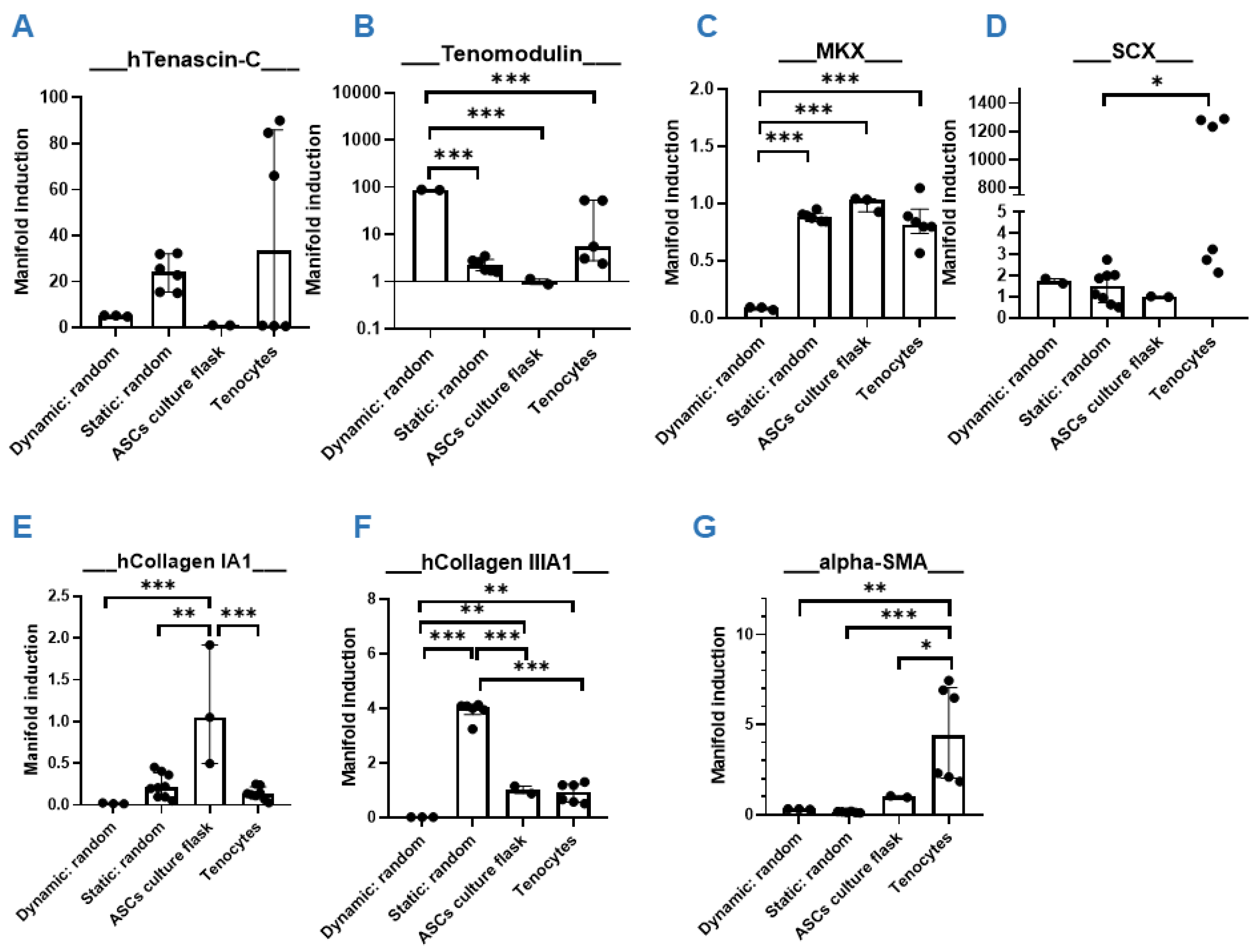
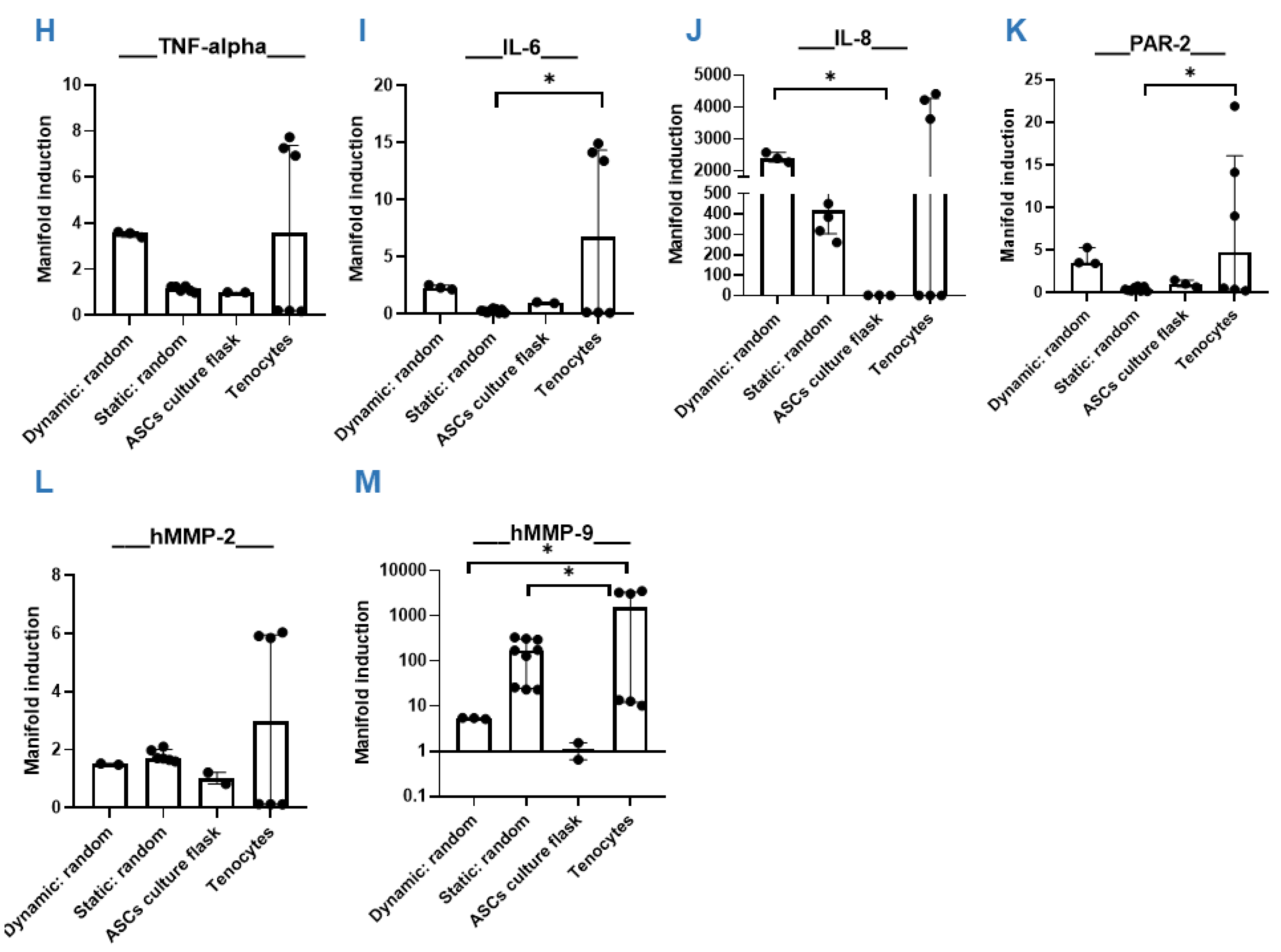
3.4. Dynamic Compared to Static Cultivation
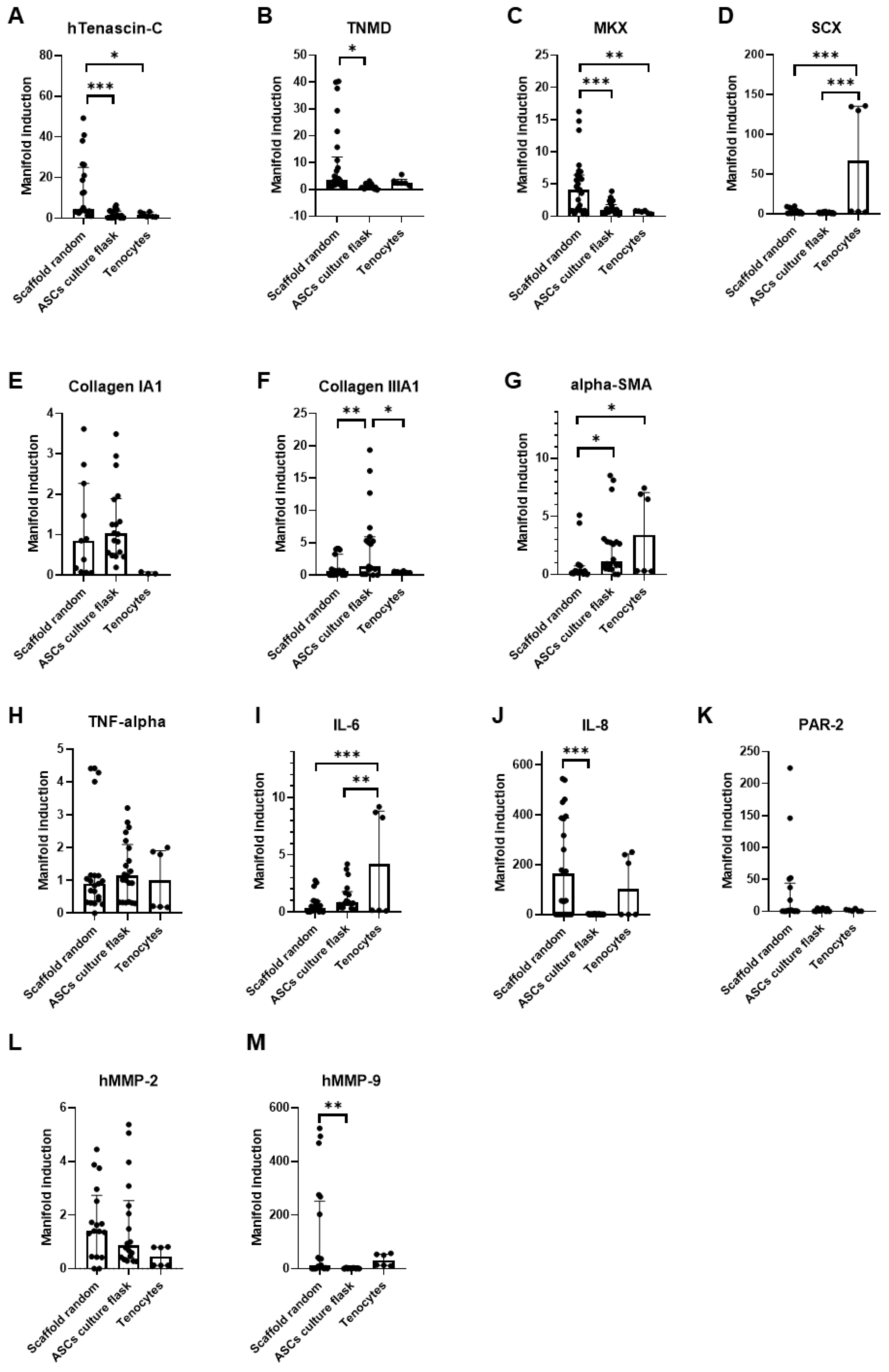
3.5. Static Cultivation on Random PVDFhfp/PDMS—The Favorite
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shapiro, E.; Grande, D.; Drakos, M. Biologics in Achilles tendon healing and repair: A review. Curr. Rev. Musculoskelet. Med. 2015, 8, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Graham, D.J.; Clitherow, H.D.S.; Singh, H.P.; Clarke, E.C.; Smith, B.J.; Tonkin, M.A. The Effect of Extensor Tendon Adhesions on Finger Motion. J. Hand Surg. 2019, 44, 903.e1–903.e5. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Maffulli, N. Tendon injury and tendinopathy: Healing and repair. J. Bone Joint Surg. Am. 2005, 87, 187–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Wang, J.; Chen, Y.; Mo, X.; Fan, C. Tenogenic adipose-derived stem cell sheets with nanoyarn scaffolds for tendon regeneration. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 119, 111506. [Google Scholar] [CrossRef]
- Xu, Y.; Yin, H.; Chu, J.; Eglin, D.; Serra, T.; Docheva, D. An anisotropic nanocomposite hydrogel guides aligned orientation and enhances tenogenesis of human tendon stem/progenitor cells. Biomater. Sci. 2021, 9, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- James, R.; Kumbar, S.G.; Laurencin, C.T.; Balian, G.; Chhabra, A.B. Tendon tissue engineering: Adipose-derived stem cell and GDF-5 mediated regeneration using electrospun matrix systems. Biomed. Mater. 2011, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evrova, O.; Bürgisser, G.M.; Ebnöther, C.; Adathala, A.; Calcagni, M.; Bachmann, E.; Snedeker, J.G.; Scalera, C.; Giovanoli, P.; Vogel, V.; et al. Elastic and surgeon friendly electrospun tubes delivering PDGF-BB positively impact tendon rupture healing in a rabbit Achilles tendon model. Biomaterials 2020, 232, 119722. [Google Scholar] [CrossRef]
- Monavarian, M.; Kader, S.; Moeinzadeh, S.; Jabbari, E. Regenerative Scar-Free Skin Wound Healing. Tissue Eng. Part B Rev. 2019, 25, 294–311. [Google Scholar] [CrossRef]
- Schulze-Tanzil, G.; Al-Sadi, O.; Wiegand, E.; Ertel, W.; Busch, C.; Kohl, B.; Pufe, T. The role of pro-inflammatory and immunoregulatory cytokines in tendon healing and rupture: New insights. Scand. J. Med. Sci. Sports 2011, 21, 337–351. [Google Scholar] [CrossRef]
- Brandt, L.; Schubert, S.; Scheibe, P.; Brehm, W.; Franzen, J.; Gross, C.; Burk, J. Tenogenic Properties of Mesenchymal Progenitor Cells Are Compromised in an Inflammatory Environment. Int. J. Mol. Sci. 2018, 19, 2549. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Wu, J.; Wang, S.; Kun, L.; Gao, J.; Chen, B.; Ye, Y.; Wang, F.; Tong, F.; Jiang, J.; et al. Control the Neural Stem Cell Fate with Biohybrid Piezoelectrical Magnetite Micromotors. Nano Lett. 2021, 21, 3518–3526. [Google Scholar] [CrossRef]
- Tang, Y.F.; Wu, C.; Wu, Z.X.; Hu, L.; Zhang, W.; Zhao, K. Fabrication and in vitro biological properties of piezoelectric bioceramics for bone regeneration. Sci. Rep. 2017, 7, 43360. [Google Scholar] [CrossRef] [Green Version]
- Park, S.E.; Yeon, G.B.; Goo, H.G.; Seo, D.S.; Dayem, A.A.; Lee, K.E.; Park, H.M.; Cho, S.G.; Kim, D.S. Maintenance and differentiation of human ES cells on polyvinylidene fluoride scaffolds immobilized with a vitronectin-derived peptide. J. Cell. Physiol. 2021, 236, 3510–3520. [Google Scholar] [CrossRef] [PubMed]
- Guillot-Ferriols, M.; Rodríguez-Hernández, J.C.; Correia, D.M.; Carabineiro, S.A.C.; Lanceros-Méndez, S.; Gómez Ribelles, J.L.; Gallego Ferrer, G. Poly(vinylidene) fluoride membranes coated by heparin/collagen layer-by-layer, smart biomimetic approaches for mesenchymal stem cell culture. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 117, 111281. [Google Scholar] [CrossRef]
- Ponnamma, D.; Chamakh, M.M.; Alahzm, A.M.; Salim, N.; Hameed, N.; AlMaadeed, M.A.A. Core-Shell Nanofibers of Polyvinylidene Fluoride-based Nanocomposites as Piezoelectric Nanogenerators. Polymers 2020, 12, 2344. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.; Zhao, K.; Ma, L.N.; Tang, Y.F.; Liu, X.; Bian, T.R. Preparation and characterization of BaTiO3/HA nanocomposite materials by hydrothermal synthesis. J. Alloy. Compd. 2017, 693, 221–225. [Google Scholar] [CrossRef]
- Sappati, K.K.; Bhadra, S. Piezoelectric Polymer and Paper Substrates: A Review. Sensors 2018, 18, 3605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urdeitx, P.; Doweidar, M.H. Enhanced Piezoelectric Fibered Extracellular Matrix to Promote Cardiomyocyte Maturation and Tissue Formation: A 3D Computational Model. Biology 2021, 10, 135. [Google Scholar] [CrossRef]
- Deniz, P.; Guler, S.; Çelik, E.; Hosseinian, P.; Aydin, H.M. Use of cyclic strain bioreactor for the upregulation of key tenocyte gene expression on Poly(glycerol-sebacate) (PGS) sheets. Mater. Sci. Eng. C 2020, 106, 110293. [Google Scholar] [CrossRef] [PubMed]
- Bacakova, L.; Zarubova, J.; Travnickova, M.; Musilkova, J.; Pajorova, J.; Slepicka, P.; Kasalkova, N.S.; Svorcik, V.; Kolska, Z.; Motarjemi, H.; et al. Stem cells: Their source, potency and use in regenerative therapies with focus on adipose-derived stem cells—A review. Biotechnol. Adv. 2018, 36, 1111–1126. [Google Scholar] [CrossRef]
- Nam, H.Y.; Pingguan-Murphy, B.; Abbas, A.A.; Merican, A.M.; Kamarul, T. Uniaxial Cyclic Tensile Stretching at 8% Strain Exclusively Promotes Tenogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stromal Cells. Stem Cells Int. 2019, 2019, 9723025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyment, N.A.; Barrett, J.G.; Awad, H.A.; Bautista, C.A.; Banes, A.J.; Butler, D.L. A brief history of tendon and ligament bioreactors: Impact and future prospects. J. Orthop. Res. 2020, 38, 2318–2330. [Google Scholar] [CrossRef]
- Kubo, Y.; Hoffmann, B.; Goltz, K.; Schnakenberg, U.; Jahr, H.; Merkel, R.; Schulze-Tanzil, G.; Pufe, T.; Tohidnezhad, M. Different Frequency of Cyclic Tensile Strain Relates to Anabolic/Catabolic Conditions Consistent with Immunohistochemical Staining Intensity in Tenocytes. Int. J. Mol. Sci. 2020, 21, 1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talò, G.; D’Arrigo, D.; Lorenzi, S.; Moretti, M.; Lovati, A.B. Independent, Controllable Stretch-Perfusion Bioreactor Chambers to Functionalize Cell-Seeded Decellularized Tendons. Ann. Biomed. Eng. 2020, 48, 1112–1126. [Google Scholar] [CrossRef] [Green Version]
- Engebretson, B.; Mussett, Z.R.; Sikavitsas, V.I. The effects of varying frequency and duration of mechanical stimulation on a tissue-engineered tendon construct. Connect. Tissue Res. 2018, 59, 167–177. [Google Scholar] [CrossRef]
- Weidenbacher, L.; Abrishamkar, A.; Rottmar, M.; Guex, A.G.; Maniura-Weber, K.; deMello, A.J.; Ferguson, S.J.; Rossi, R.M.; Fortunato, G. Electrospraying of microfluidic encapsulated cells for the fabrication of cell-laden electrospun hybrid tissue constructs. Acta Biomater. 2017, 64, 137–147. [Google Scholar] [CrossRef]
- Ahmed, F.; Dutta, N.K.; Zannettino, A.; Vandyke, K.; Choudhury, N.R. Engineering interaction between bone marrow derived endothelial cells and electrospun surfaces for artificial vascular graft applications. Biomacromolecules 2014, 15, 1276–1287. [Google Scholar] [CrossRef]
- Kamberi, M.; Pinson, D.; Pacetti, S.; Perkins, L.E.L.; Hossainy, S.; Mori, H.; Rapoza, R.J.; Kolodgie, F.; Virmani, R. Evaluation of chemical stability of polymers of XIENCE everolimus-eluting coronary stents in vivo by pyrolysis-gas chromatography/mass spectrometry. J. Biomed. Mater. Res. B Appl. Biomater. 2018, 106, 1721–1729. [Google Scholar] [CrossRef]
- Brunelli, M.; Alther, S.; Rossi, R.M.; Ferguson, S.J.; Rottmar, M.; Fortunato, G. Nanofiber membranes as biomimetic and mechanically stable surface coatings. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 108, 110417. [Google Scholar] [CrossRef]
- Najjar, R.; Luo, Y.; Jao, D.; Brennan, D.; Xue, Y.; Beachley, V.; Hu, X.; Xue, W. Biocompatible Silk/Polymer Energy Harvesters Using Stretched Poly (vinylidene fluoride-co-hexafluoropropylene) (PVDF-HFP) Nanofibers. Polymers 2017, 9, 479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.Y.; Bandekar, A.; Chase, G.G. Evaluation of electrowet coalescer in series with PVDF-HFP electrospun fiber membranes for separation of water from ULSD. Fuel 2018, 225, 111–117. [Google Scholar] [CrossRef]
- Wei, Z.; Reisdorf, R.L.; Thoreson, A.R.; Jay, G.D.; Moran, S.L.; An, K.N.; Amadio, P.C.; Zhao, C. Comparison of Autograft and Allograft with Surface Modification for Flexor Tendon Reconstruction: A Canine in Vivo Model. J. Bone Joint Surg. Am. 2018, 100, e42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Lu, C.C.; Reisdorf, R.L.; Thoreson, A.R.; Gingery, A.; Moran, S.L.; Amadio, P.C.; Zhao, C. Revitalized and synovialized allograft for intrasynovial flexor tendon reconstruction in an in vivo canine model. J. Orthop. Res. 2018, 36, 2218–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lineberry, K.D.; Shue, S.; Chepla, K.J. The Management of Partial Zone II Intrasynovial Flexor Tendon Lacerations: A Literature Review of Biomechanics, Clinical Outcomes, and Complications. Plast. Reconstr. Surg. 2018, 141, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.R.; Smith, R.K.; David, F.; Lam, R.; Hughes, G.; De Godoy, R.; Carr, A.J.; Goodship, A.E.; Dudhia, J. Evaluation of the Effects of Synovial Multipotent Cells on Deep Digital Flexor Tendon Repair in a Large Animal Model of Intra-Synovial Tendinopathy. J. Orthop. Res. 2020, 38, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Schoenenberger, A.D.; Tempfer, H.; Lehner, C.; Egloff, J.; Mauracher, M.; Bird, A.; Widmer, J.; Maniura-Weber, K.; Fucentese, S.F.; Traweger, A.; et al. Macromechanics and polycaprolactone fiber organization drive macrophage polarization and regulate inflammatory activation of tendon in vitro and in vivo. Biomaterials 2020, 249, 120034. [Google Scholar] [CrossRef] [PubMed]
- Buschmann, J.; Gao, S.; Härter, L.; Hemmi, S.; Welti, M.; Werner, C.M.L.; Calcagni, M.; Cinelli, P.; Wanner, G.A. Yield and proliferation rate of adipose-derived stem cells as a function of age, BMI and harvest site: Increasing the yield by using adherent and supernatant fractions? Cytotherapy 2013, 15, 1098–1105. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronthos, S.; Franklin, D.M.; Leddy, H.A.; Robey, P.G.; Storms, R.W.; Gimble, J.M. Surface protein characterization of human adipose tissue-derived stromal cells. J. Cell. Physiol. 2001, 189, 54–63. [Google Scholar] [CrossRef]
- Buschmann, J.; Härter, L.; Gao, S.; Hemmi, S.; Welti, M.; Hild, N.; Schneider, O.D.; Stark, W.J.; Lindenblatt, N.; Werner, C.M.L.; et al. Tissue engineered bone grafts based on biomimetic nanocomposite PLGA/amorphous calcium phosphate scaffold and human adipose-derived stem cells. Injury 2012, 43, 1689–1697. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.P.; Calcagni, M.; Welti, M.; Hemmi, S.; Hild, N.; Stark, W.J.; Meier Buergisser, G.; Wanner, G.A.; Cinelli, P.; Buschmann, J. Proliferation of ASC-derived endothelial cells in a 3D electrospun mesh: Impact of bone-biomimetic nanocomposite and co-culture with ASC-derived osteoblasts. Injury 2014, 45, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Hess, S.C.; Stark, W.J.; Mohn, D.; Cohrs, N.H.; Märsmann, S.; Calcagni, M.; Cinelli, P.; Buschmann, J. Gene expression in human adipose-derived stem cells: Comparison of 2D films, 3D electrospun meshes or co-cultured scaffolds with two-way paracrine effects. Eur. Cells Mater. 2017, 34, 232–248. [Google Scholar] [CrossRef]
- D’Addona, A.; Maffulli, N.; Formisano, S.; Rosa, D. Inflammation in tendinopathy. Surgeon 2017, 15, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Galatz, L.M.; Gerstenfeld, L.; Heber-Katz, E.; Rodeo, S.A. Tendon regeneration and scar formation: The concept of scarless healing. J. Orthop. Res. 2015, 33, 823–831. [Google Scholar] [CrossRef]
- Ciardulli, M.C.; Marino, L.; Lamparelli, E.P.; Guida, M.; Forsyth, N.R.; Selleri, C.; Della Porta, G.; Maffulli, N. Dose-Response Tendon-Specific Markers Induction by Growth Differentiation Factor-5 in Human Bone Marrow and Umbilical Cord Mesenchymal Stem Cells. Int. J. Mol. Sci. 2020, 21, 5905. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, X.; Zhang, E.; Yang, L.; Yuan, H.; Tu, W.; Zhang, H.; Yin, Z.; Shen, W.; Chen, X.; et al. An Epigenetic Bioactive Composite Scaffold with Well-aligned Nanofibers for Functional Tendon Tissue Engineering. Acta Biomater. 2017, 66, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.X.; Wang, Y.S.; Ma, C.; Zheng, W.; Li, L.; Zheng, Y.F. Electrospinning of PLGA/gelatin randomly-oriented and aligned nanofibers as potential scaffold in tissue engineering. Mater. Sci. Eng. C Mater. Biol. Appl. 2010, 30, 1204–1210. [Google Scholar] [CrossRef]
- Beason, D.P.; Connizzo, B.K.; Dourte, L.M.; Mauck, R.L.; Soslowsky, L.J.; Steinberg, D.R.; Bernstein, J. Fiber-aligned polymer scaffolds for rotator cuff repair in a rat model. J. Shoulder Elb. Surg. 2012, 21, 245–250. [Google Scholar] [CrossRef]
- Xie, J.; Shen, H.; Yuan, G.; Lin, K.; Su, J. The effects of alignment and diameter of electrospun fibers on the cellular behaviors and osteogenesis of BMSCs. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 120, 111787. [Google Scholar] [CrossRef]
- Leite, M.L.; Soares, D.G.; Anovazzi, G.; Mendes Soares, I.P.; Hebling, J.; de Souza Costa, C.A. Development of fibronectin-loaded nanofiber scaffolds for guided pulp tissue regeneration. J. Biomed. Mater. Res. B Appl. Biomater. 2021, 109, 1244–1258. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.; Mbimba, T.; Younesi, M.; Akkus, O. Effects of substrate stiffness on the tenoinduction of human mesenchymal stem cells. Acta Biomater. 2017, 58, 244–253. [Google Scholar] [CrossRef]
- Perikamana, S.K.M.; Lee, J.; Ahmad, T.; Kim, E.M.; Byun, H.; Lee, S.; Shin, H. Harnessing biochemical and structural cues for tenogenic differentiation of adipose derived stem cells (ADSCs) and development of an in vitro tissue interface mimicking tendon-bone insertion graft. Biomaterials 2018, 165, 79–93. [Google Scholar] [CrossRef]
- Midwood, K.S.; Chiquet, M.; Tucker, R.P.; Orend, G. Tenascin-C at a glance. J. Cell Sci. 2016, 129, 4321–4327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kai, D.; Prabhakaran, M.P.; Jin, G.; Ramakrishna, S. Guided orientation of cardiomyocytes on electrospun aligned nanofibers for cardiac tissue engineering. J. Biomed. Mater. Res. B Appl. Biomater. 2011, 98, 379–386. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, C.; Zhu, S.; Lu, P.; Zhu, T.; Gong, X.; Zhang, Z.; Hu, J.; Yin, Z.; Heng, B.C.; et al. Mohawk Promotes the Tenogenesis of Mesenchymal Stem Cells through Activation of the TGF beta Signaling Pathway. Stem Cells 2015, 33, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Ito, Y.; Shinohara, M.; Yamashita, S.; Ichinose, S.; Kishida, A.; Oyaizu, T.; Kayama, T.; Nakamichi, R.; Koda, N.; et al. Gene targeting of the transcription factor Mohawk in rats causes heterotopic ossification of Achilles tendon via failed tenogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, 7840–7845. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Gao, J.; Mi, L.; Zhang, G.; Zhang, L.; Zhang, N.; Huo, R.; Hu, J.; Xu, K. Synovial membrane mesenchymal stem cells: Past life, current situation, and application in bone and joint diseases. Stem Cell. Res. Ther. 2020, 11, 381. [Google Scholar] [CrossRef]
- Alexandrov, V.P.; Naimov, S.I. A Prospectus of Tenomodulin. Folia Med. 2016, 58, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Docheva, D.; Hunziker, E.B.; Fassler, R.; Brandau, O. Tenomodulin is necessary for tenocyte proliferation and tendon maturation. Mol. Cell. Biol. 2005, 25, 699–705. [Google Scholar] [CrossRef] [Green Version]
- Dex, S.; Alberton, P.; Willkomm, L.; Söllradl, T.; Bago, S.; Milz, S.; Shakibaei, M.; Ignatius, A.; Bloch, W.; Clausen-Schaumann, H.; et al. Tenomodulin is Required for Tendon Endurance Running and Collagen I Fibril Adaptation to Mechanical Load. EBioMedicine 2017, 20, 240–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.; Alberton, P.; Caceres, M.D.; Volkmer, E.; Schieker, M.; Docheva, D. Tenomodulin is essential for prevention of adipocyte accumulation and fibrovascular scar formation during early tendon healing. Cell Death Dis. 2017, 8, e3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, N.L.; Silbernagel, K.G.; Thorborg, K.; Kirwan, P.D.; Galatz, L.M.; Abrams, G.D.; Murrell, G.A.C.; McInnes, I.B.; Rodeo, S.A. Tendinopathy. Nat. Rev. Dis. Primers 2021, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Heinemeier, K.M.; Kjaer, M. In vivo investigation of tendon responses to mechanical loading. J. Musculoskelet. Neuronal Interact. 2011, 11, 115–123. [Google Scholar] [PubMed]
- Giz, A.S.; Aydelik-Ayazoglu, S.; Catalgil-Giz, H.; Bayraktar, H.; Alaca, B.E. Stress relaxation and humidity dependence in sodium alginate-glycerol films. J. Mech. Behav. Biomed. Mater. 2019, 100, 103374. [Google Scholar] [CrossRef]
- Brunelli, M.; Perrault, C.M.; Lacroix, D. Short bursts of cyclic mechanical compression modulate tissue formation in a 3D hybrid scaffold. J. Mech. Behav. Biomed. Mater. 2017, 71, 165–174. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | 5′-3′ | Primers |
|---|---|---|
| hTenascin-C | Forward | GGTGGATGGATTGTGTTCCTGAGA |
| Reverse | CTGTGTCCTTGTCAAAGGTGGAGA | |
| hMMP-2 | Forward | TGCGACCACAGCCAACTACG |
| Reverse | TGGGACAGACGGAAGTTCTTGG | |
| hMMP-9 | Forward | GACGCCGCTCACCTTCACTC |
| Reverse | TTGGAACCACGACGCCCTTG | |
| hCOL3A1 | Forward | CAGCGGTTCTCCAGGCAAGG |
| Reverse | CTCCAGTGATCCCAGCAATCCC | |
| hCOL1A1 | Forward | TGA CGA GAC CAA GAA CTG |
| Reverse | CCA TCC AAA CCA CTG AAA CC | |
| hTNMD | Forward | CCATGCTGGATGAGAGAGGTT |
| Reverse | TTGGTAGCAGTATGGATATGGGT | |
| hTNF-α | Forward | CGGACACCATGGACAAGTTT |
| Reverse | GAAAGCCTTGCAGAGGTCAG | |
| hα-SMA | Forward | ACTGAGCGTGGCTATTCCTCCGTT |
| Reverse | GCAGTGGCCATCTCATTTTCA | |
| hMKX | Forward | TCAAGGACAACCTCGGCCTG |
| Reverse | ACGGGTTGTCACGGTGCTTG | |
| hSCX | Forward | AGAACACCCAGCCCAAACAG |
| Reverse | GGCCACCTCCTAACTGCGAATC | |
| hIL-6 | Forward | GTAGCCGCCCACACAGACAGCC |
| Reverse | GCCATCTTTGGAAGGTTC | |
| hIL-8 | Forward | TCTGCAGCTCTGTGTGAAGGT |
| Reverse | TGAATTCTCAGCCCTCTTCAA | |
| hPAR2 | Forward | GTTGATGGCACATCCCACGTC |
| Reverse | GTACAGGGCATAGACATGGC | |
| gapdH | Forward | ACCACAGTCCATGCCATCAC |
| Reverse | TCCACCACCCTGTTGCTGTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baumgartner, W.; Wolint, P.; Hofmann, S.; Nüesch, C.; Calcagni, M.; Brunelli, M.; Buschmann, J. Impact of Electrospun Piezoelectric Core–Shell PVDFhfp/PDMS Mesh on Tenogenic and Inflammatory Gene Expression in Human Adipose-Derived Stem Cells: Comparison of Static Cultivation with Uniaxial Cyclic Tensile Stretching. Bioengineering 2022, 9, 21. https://doi.org/10.3390/bioengineering9010021
Baumgartner W, Wolint P, Hofmann S, Nüesch C, Calcagni M, Brunelli M, Buschmann J. Impact of Electrospun Piezoelectric Core–Shell PVDFhfp/PDMS Mesh on Tenogenic and Inflammatory Gene Expression in Human Adipose-Derived Stem Cells: Comparison of Static Cultivation with Uniaxial Cyclic Tensile Stretching. Bioengineering. 2022; 9(1):21. https://doi.org/10.3390/bioengineering9010021
Chicago/Turabian StyleBaumgartner, Walter, Petra Wolint, Silvan Hofmann, Cléa Nüesch, Maurizio Calcagni, Marzia Brunelli, and Johanna Buschmann. 2022. "Impact of Electrospun Piezoelectric Core–Shell PVDFhfp/PDMS Mesh on Tenogenic and Inflammatory Gene Expression in Human Adipose-Derived Stem Cells: Comparison of Static Cultivation with Uniaxial Cyclic Tensile Stretching" Bioengineering 9, no. 1: 21. https://doi.org/10.3390/bioengineering9010021
APA StyleBaumgartner, W., Wolint, P., Hofmann, S., Nüesch, C., Calcagni, M., Brunelli, M., & Buschmann, J. (2022). Impact of Electrospun Piezoelectric Core–Shell PVDFhfp/PDMS Mesh on Tenogenic and Inflammatory Gene Expression in Human Adipose-Derived Stem Cells: Comparison of Static Cultivation with Uniaxial Cyclic Tensile Stretching. Bioengineering, 9(1), 21. https://doi.org/10.3390/bioengineering9010021