Assessment of the Inner Surface Microstructure of Decellularized Cortical Bone by a Scanning Electron Microscope

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Femoral Bone Harvest
2.2. MicroCT Observation
2.3. Decellularization Treatment
2.4. Hematoxylin and Eosin Staining
2.5. Scanning Electron Microscopy Observation
2.6. Image and Statistical Analysis
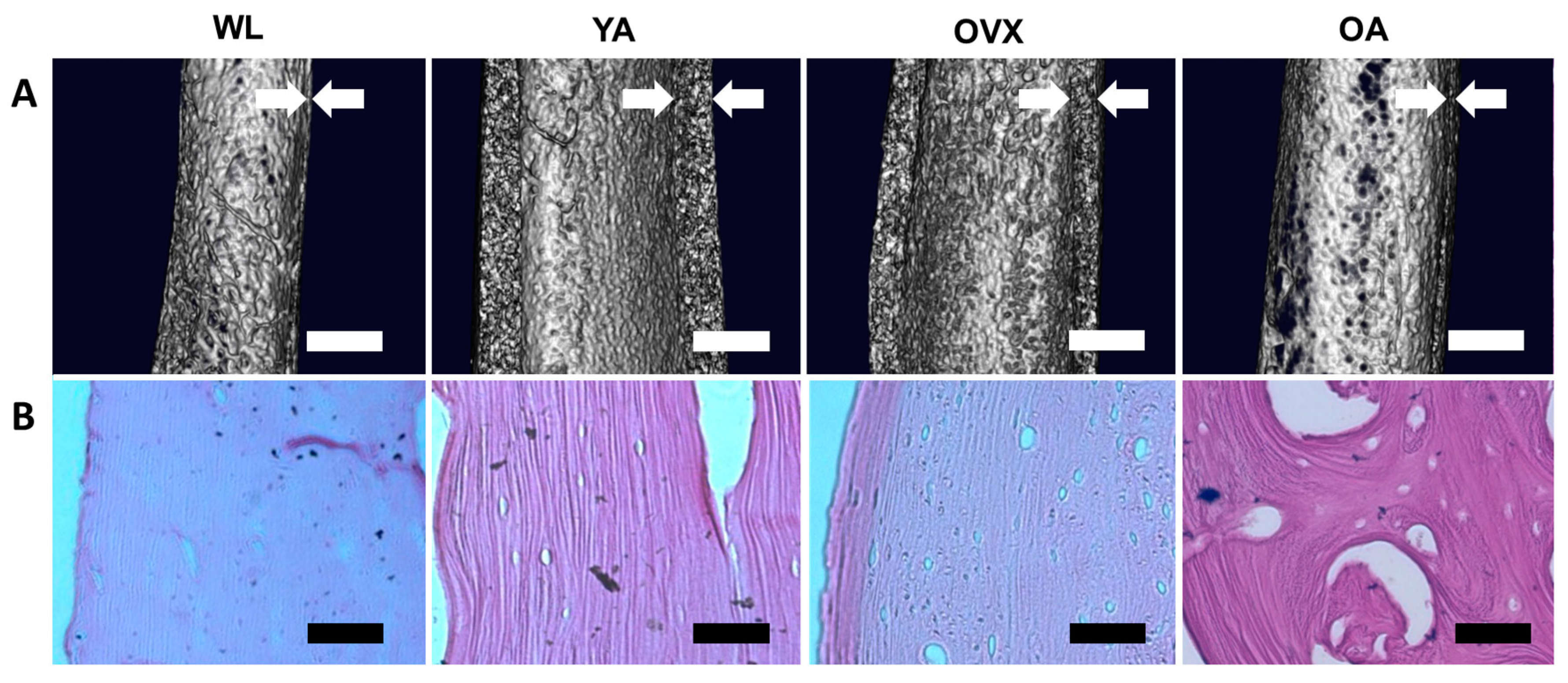
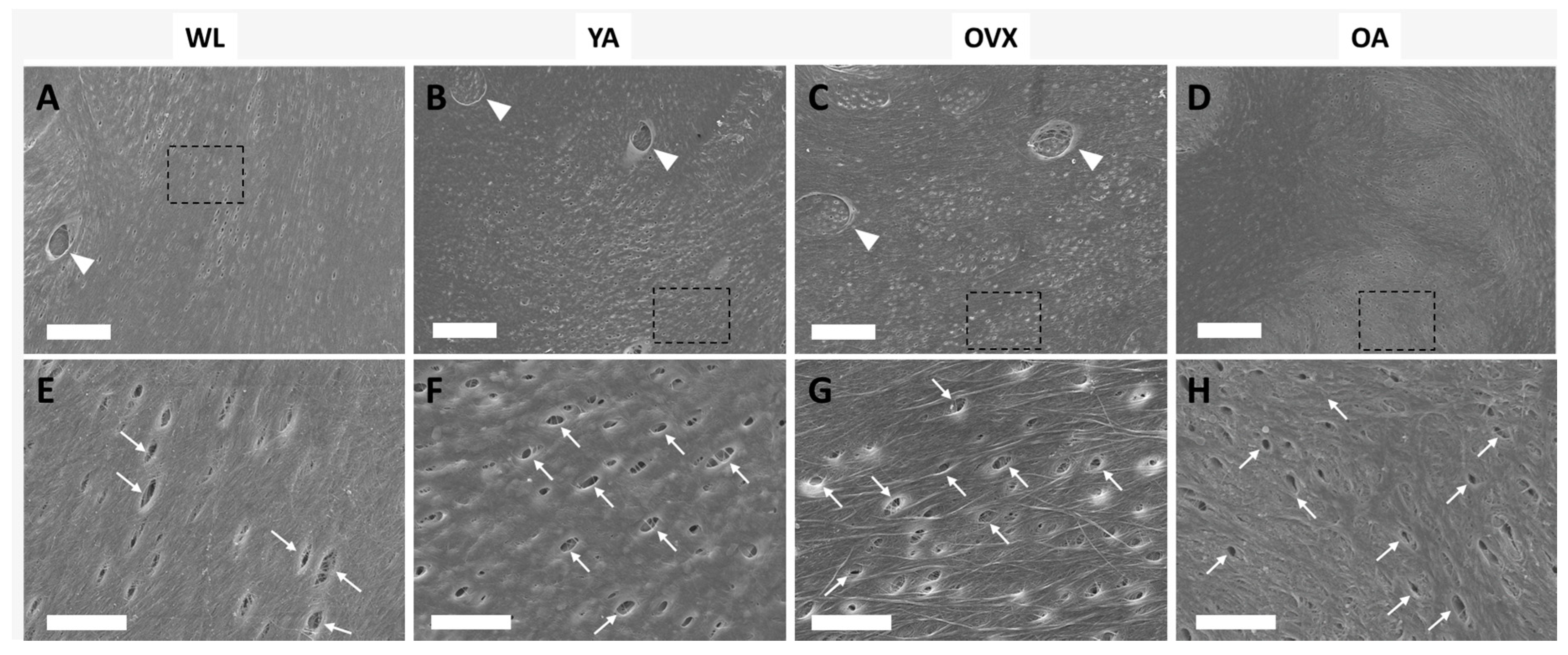
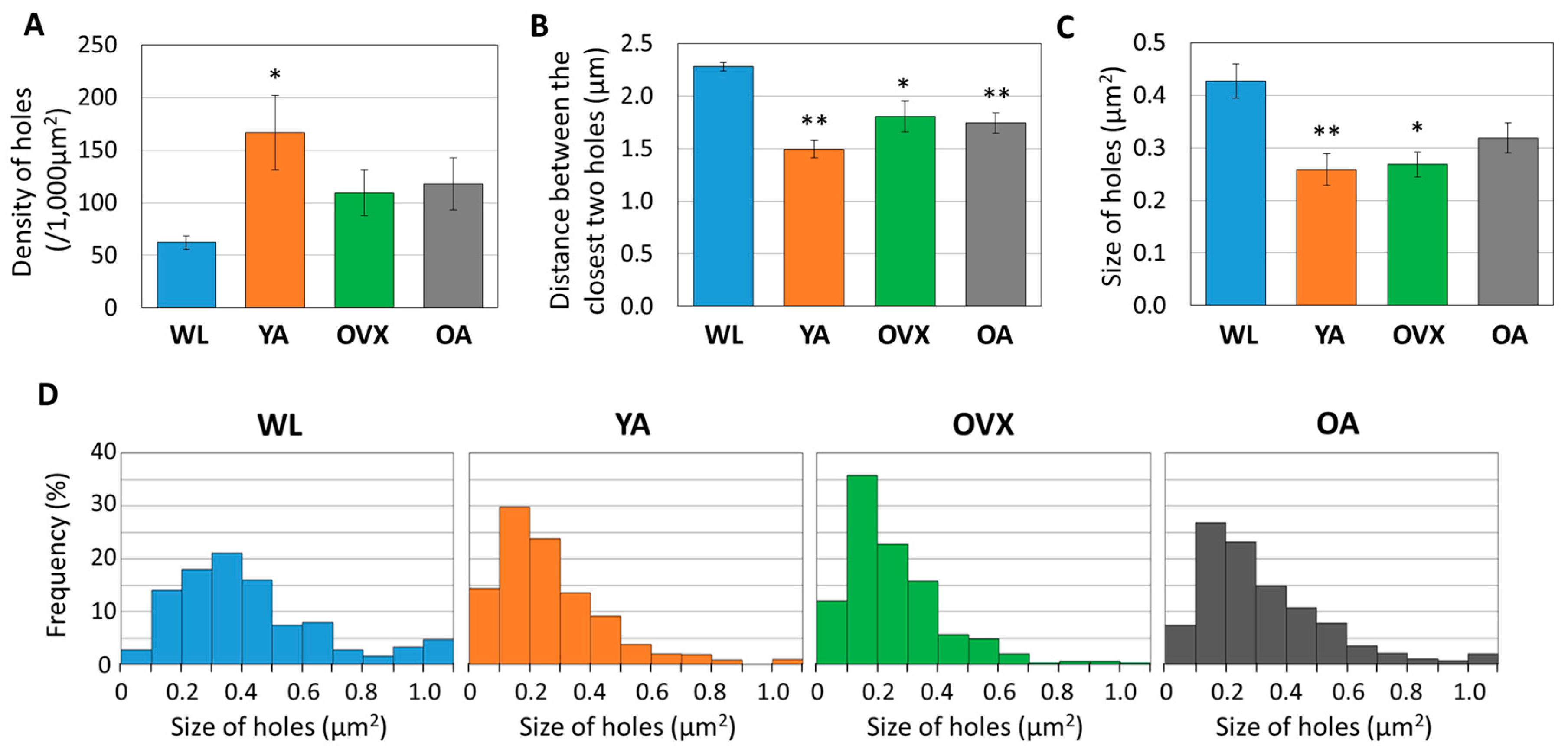
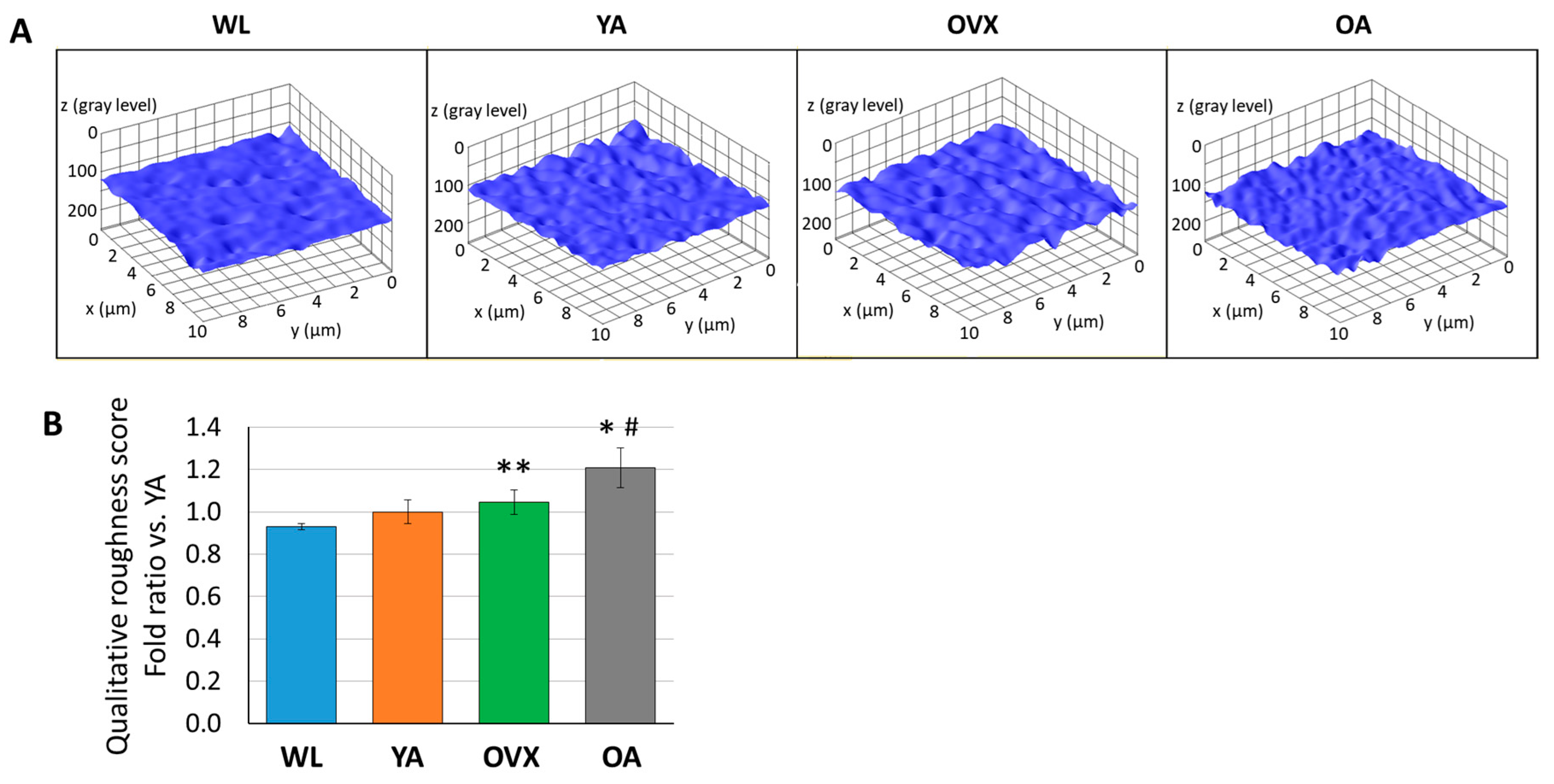
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weiner, S.; Wagner, H.D. THE MATERIAL BONE: Structure-Mechanical Function Relations. Annu. Rev. Mater. Sci. 1998, 28, 271–298. [Google Scholar] [CrossRef]
- Reznikov, N.; Shahar, R.; Weiner, S. Three-dimensional structure of human lamellar bone: The presence of two different materials and new insights into the hierarchical organization. Bone 2014, 59, 93–104. [Google Scholar] [CrossRef]
- Hattner, R.; Epker, B.N.; Frost, H. Suggested Sequential Mode of Control of Changes in Cell Behaviour in Adult Bone Remodelling. Nature 1965, 206, 489–490. [Google Scholar] [CrossRef]
- Martin, T.J.; Sims, N.A. Osteoclast-derived activity in the coupling of bone formation to resorption. Trends Mol. Med. 2005, 11, 76–81. [Google Scholar] [CrossRef]
- Phan, T.C.A.; Xu, J.; Ming, H. Zheng Interaction between osteoblast and osteoclast: Impact in bone disease. Histol. Histopathol. 2004, 19, 1325–1344. [Google Scholar]
- Buckwalter, J.A.; Glimcher, M.J.; Cooper, R.R.; Recker, R.R. Bone biology. I: Structure, blood supply, cells, matrix, and mineralization. J. Bone Jt. Surg. Ser. A 1995, 77, 1256–1275. [Google Scholar] [CrossRef]
- Downey, P.A.; Siegel, M.I. Bone biology and the clinical implications for osteoporosis. Phys. Ther. 2006, 86, 77–91. [Google Scholar] [CrossRef]
- Parfitt, A.M. Age-related structural changes in trabecular and cortical bone: Cellular mechanisms and biomechanical consequences. Calcif. Tissue Int. 1984, 36. [Google Scholar] [CrossRef]
- Pazzaglia, U.E.; Congiu, T.; Raspanti, M.; Ranchetti, F.; Quacci, D. Anatomy of the intracortical canal system: Scanning electron microscopy study in rabbit femur. Clin. Orthop. Relat. Res. 2009, 467, 2446–2456. [Google Scholar] [CrossRef]
- Pazzaglia, U.E.; Sibilia, V.; Congiu, T.; Pagani, F.; Ravanelli, M.; Zarattini, G. Setup of a bone aging experimental model in the rabbit comparing changes in cortical and trabecular bone: Morphological and morphometric study in the femur. J. Morphol. 2015, 276, 733–747. [Google Scholar] [CrossRef]
- You, L.D.; Weinbaum, S.; Cowin, S.C.; Schaffler, M.B. Ultrastructure of the osteocyte process and its pericellular matrix. Anat. Rec. Part A 2004, 278, 505–513. [Google Scholar] [CrossRef]
- Reznikov, N.; Bilton, M.; Lari, L.; Stevens, M.M.; Kröger, R. Fractal-like hierarchical organization of bone begins at the nanoscale. Science 2018, 360, eaao2189. [Google Scholar] [CrossRef]
- Wronski, T.J.; Cintrón, M.; Dann, L.M. Temporal relationship between bone loss and increased bone turnover in ovariectomized rats. Calcif. Tissue Int. 1988, 43, 179–183. [Google Scholar] [CrossRef]
- Lee, D.J.; Diachina, S.; Lee, Y.T.; Zhao, L.; Zou, R.; Tang, N.; Han, H.; Chen, X.; Ko, C.-C. Decellularized bone matrix grafts for calvaria regeneration. J. Tissue Eng. 2016, 7, 204173141668030. [Google Scholar] [CrossRef]
- Bertelsen, P.K.; Clement, J.G.; Thomas, C.D.L. A morphometric study of the cortex of the human femur from early childhood to advanced old age. Forensic Sci. Int. 1995, 74, 63–77. [Google Scholar] [CrossRef]
- Garn, S.M. The Earlier Gain and the Later Loss of Cortical Bone, in Nutritional Perspective; Charles C Thomas Publisher: Springfield, IL, USA, 1970. [Google Scholar]
- Shah, F.A.; Zanghellini, E.; Matic, A.; Thomsen, P.; Palmquist, A. The Orientation of Nanoscale Apatite Platelets in Relation to Osteoblastic-Osteocyte Lacunae on Trabecular Bone Surface. Calcif. Tissue Int. 2016, 98, 193–205. [Google Scholar] [CrossRef]
- Tate, M.L.K.; Adamson, J.R.; Tami, A.E.; Bauer, T.W. The Osteocyte. Int. J. Biochem. Cell Biol. 2004, 36, 1–8. [Google Scholar] [CrossRef]
- Banerjee, S.; Yang, R.; Courchene, C.E.; Conners, T.E. Scanning electron microscopy measurements of the surface roughness of paper. Ind. Eng. Chem. Res. 2009, 48, 4322–4325. [Google Scholar] [CrossRef]
- Castle, J.E.; Zhdan, P.A. Characterization of surface topography by SEM and SFM: Problems and solutions. J. Phys. D. Appl. Phys. 1997, 30, 722–740. [Google Scholar] [CrossRef]
- Makihira, S.; Mine, Y.; Kosaka, E.; Nikawa, H. Titanium surface roughness accelerates RANKL-dependent differentiation in the osteoclast precursor cell line, RAW264. 7. Dent. Mater. J. 2007, 26, 739–745. [Google Scholar] [CrossRef]
- Costa-Rodrigues, J.; Carmo, S.; Perpétuo, I.P.; Monteiro, F.J.; Fernandes, M.H. Osteoclastogenic differentiation of human precursor cells over micro- and nanostructured hydroxyapatite topography. Biochim. Biophys. Acta Gen. Subj. 2016, 1860, 825–835. [Google Scholar] [CrossRef]
- Ciapetti, G.; Di Pompo, G.; Avnet, S.; Martini, D.; Diez-Escudero, A.; Montufar, E.B.; Ginebra, M.P.; Baldini, N. Osteoclast differentiation from human blood precursors on biomimetic calcium-phosphate substrates. Acta Biomater. 2017, 50, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Deng, Y.; Liu, X.; Xu, A.; Wang, L.; Luo, Z.; Zheng, Y.; Deng, F.; Tang, Z.; Wei, J. Effect of surface roughness on osteogenesis in vitro and osseointegration in vivo of carbon fiber-reinforced polyetheretherketone– nanohydroxyapatite composite. Int. J. Nanomed. 2015, 10, 1425–1447. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-N.; Lee, J.-Y.; Shin, K.-J.; Gil, Y.-C.; Koh, K.-S.; Song, W.-C. Morphological characteristics of the Haversian system. Anat. Cell Biol. 2015, 48, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Georgiadis, M.; Müller, R.; Schneider, P. Techniques to assess bone ultrastructure organization: orientation and arrangement of mineralized collagen fibrils. J. R. Soc. Interface 2016, 13, 20160088. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, A.; Hewitt, R.N.; McNamara, L.E.; McCloy, D.; Dominic Meek, R.M.; Dalby, M.J. Biomimetic microtopography to enhance osteogenesis in vitro. Acta Biomater. 2011, 7, 2919–2925. [Google Scholar] [CrossRef] [PubMed]
- Seo, C.H.; Jeong, H.; Feng, Y.; Montagne, K.; Ushida, T.; Suzuki, Y.; Furukawa, K.S. Micropit surfaces designed for accelerating osteogenic differentiation of murine mesenchymal stem cells via enhancing focal adhesion and actin polymerization. Biomaterials 2014, 35, 2245–2252. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Chen, Y.; Tang, C.; Fei, Y.; Wu, H.; Ruan, D.; Paul, M.E.; Chen, X.; Yin, Z.; Heng, B.C.; et al. The relationship between substrate topography and stem cell differentiation in the musculoskeletal system. Cell. Mol. Life Sci. 2019, 76, 505–521. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, D.; Costa, P.; Neves, N.; Gomes, M.E.; Reis, R.L. Micro- and nanotechnology in tissue engineering. Nanotechnology 2011, 22, 3–29. [Google Scholar]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, H.; Asai, J.; Ushida, T.; Furukawa, K.S. Assessment of the Inner Surface Microstructure of Decellularized Cortical Bone by a Scanning Electron Microscope. Bioengineering 2019, 6, 86. https://doi.org/10.3390/bioengineering6030086
Jeong H, Asai J, Ushida T, Furukawa KS. Assessment of the Inner Surface Microstructure of Decellularized Cortical Bone by a Scanning Electron Microscope. Bioengineering. 2019; 6(3):86. https://doi.org/10.3390/bioengineering6030086
Chicago/Turabian StyleJeong, Heonuk, Jungo Asai, Takashi Ushida, and Katsuko S. Furukawa. 2019. "Assessment of the Inner Surface Microstructure of Decellularized Cortical Bone by a Scanning Electron Microscope" Bioengineering 6, no. 3: 86. https://doi.org/10.3390/bioengineering6030086
APA StyleJeong, H., Asai, J., Ushida, T., & Furukawa, K. S. (2019). Assessment of the Inner Surface Microstructure of Decellularized Cortical Bone by a Scanning Electron Microscope. Bioengineering, 6(3), 86. https://doi.org/10.3390/bioengineering6030086