Growth Behavior of Human Adipose Tissue-Derived Stromal/Stem Cells at Small Scale: Numerical and Experimental Investigations




Abstract
1. Introduction
2. Materials and Methods
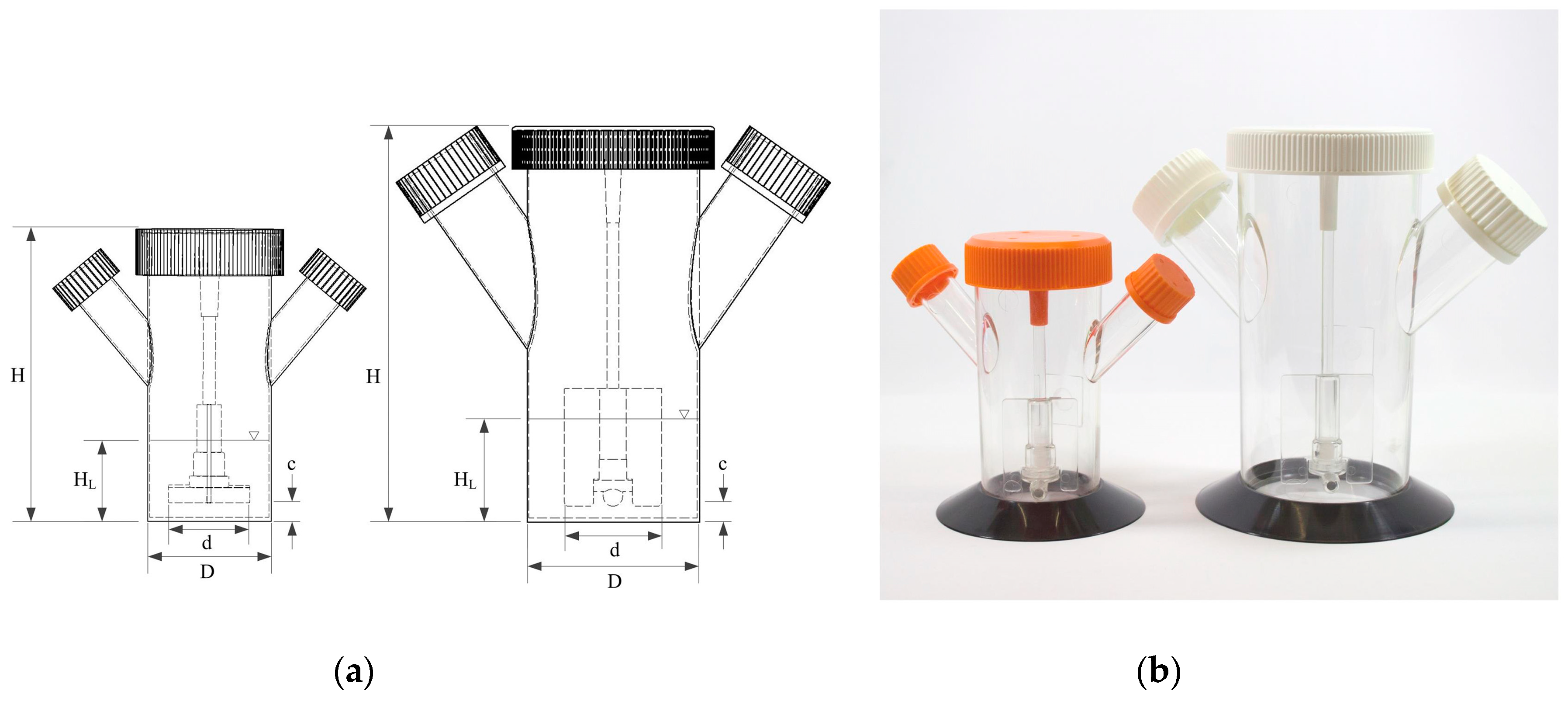
2.1. Bioreactor Systems: 125 mL and 500 mL Disposable Conring® Spinner Flasks
2.2. Numerical Investigations
2.2.1. CFD
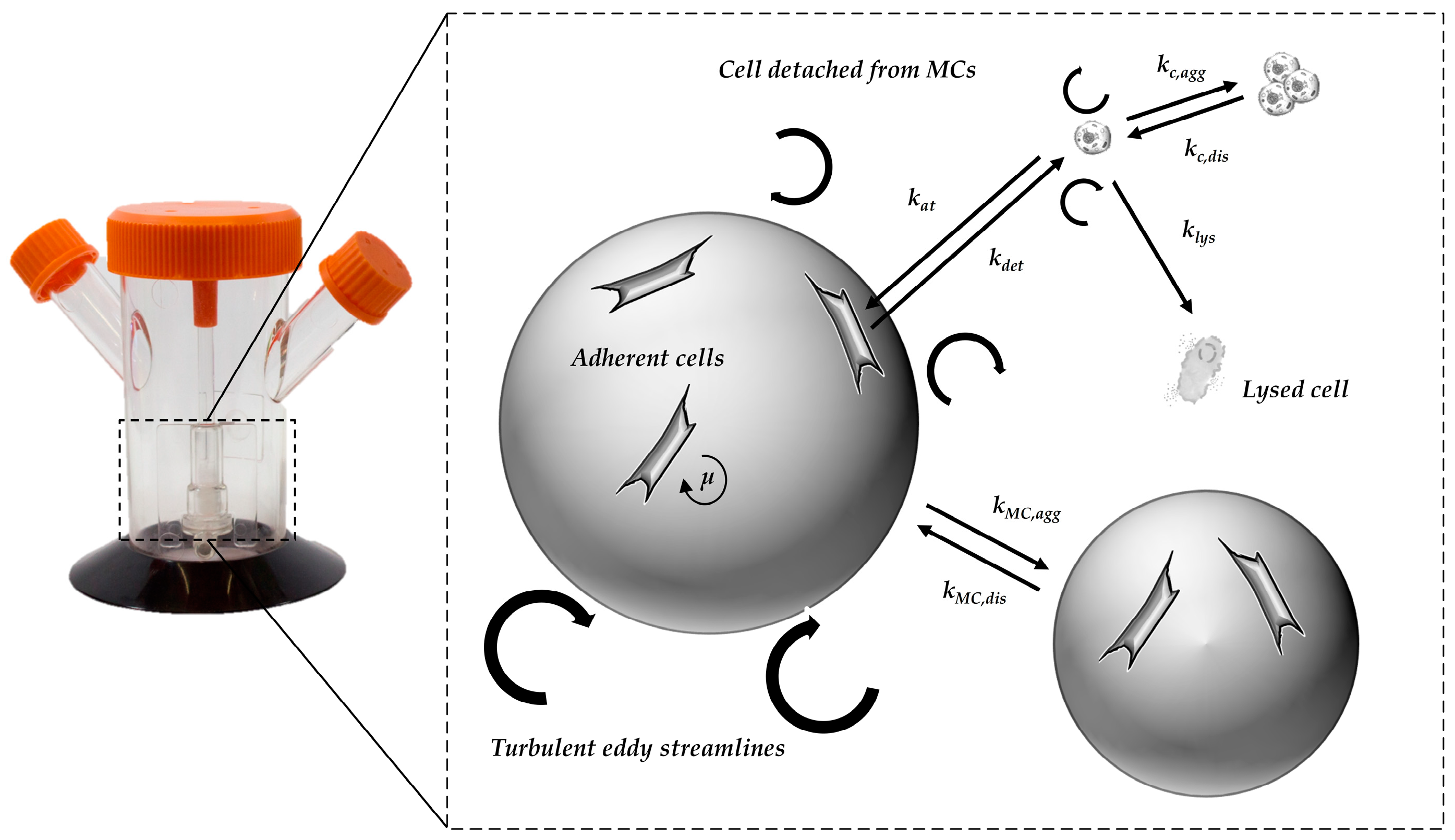
2.2.2. Segregated Growth Model (SGM)
2.3. Biochemical Engineering Investigations
2.3.1. Suspension Studies
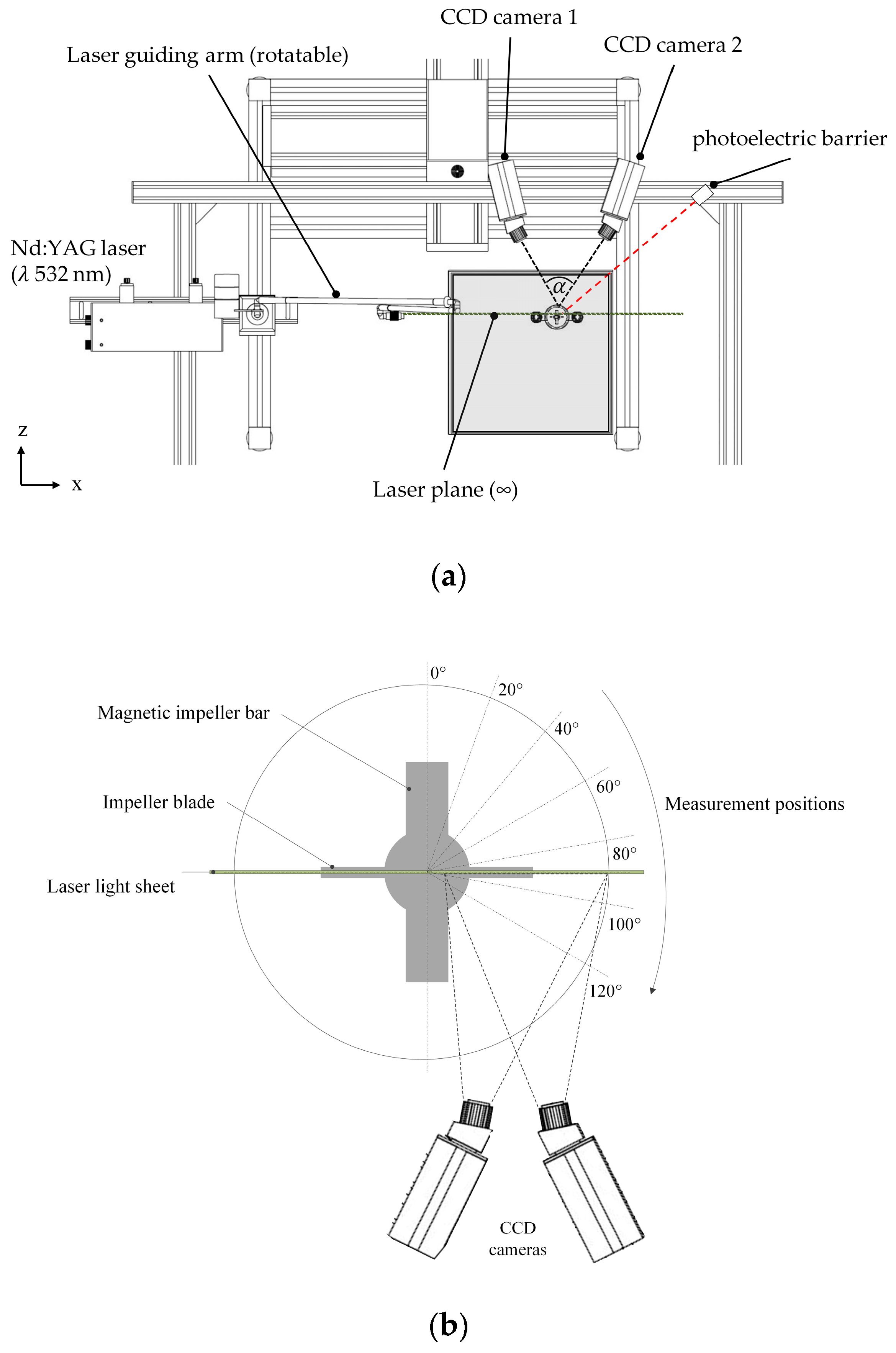
2.3.2. Particle Image Velocimetry (PIV)
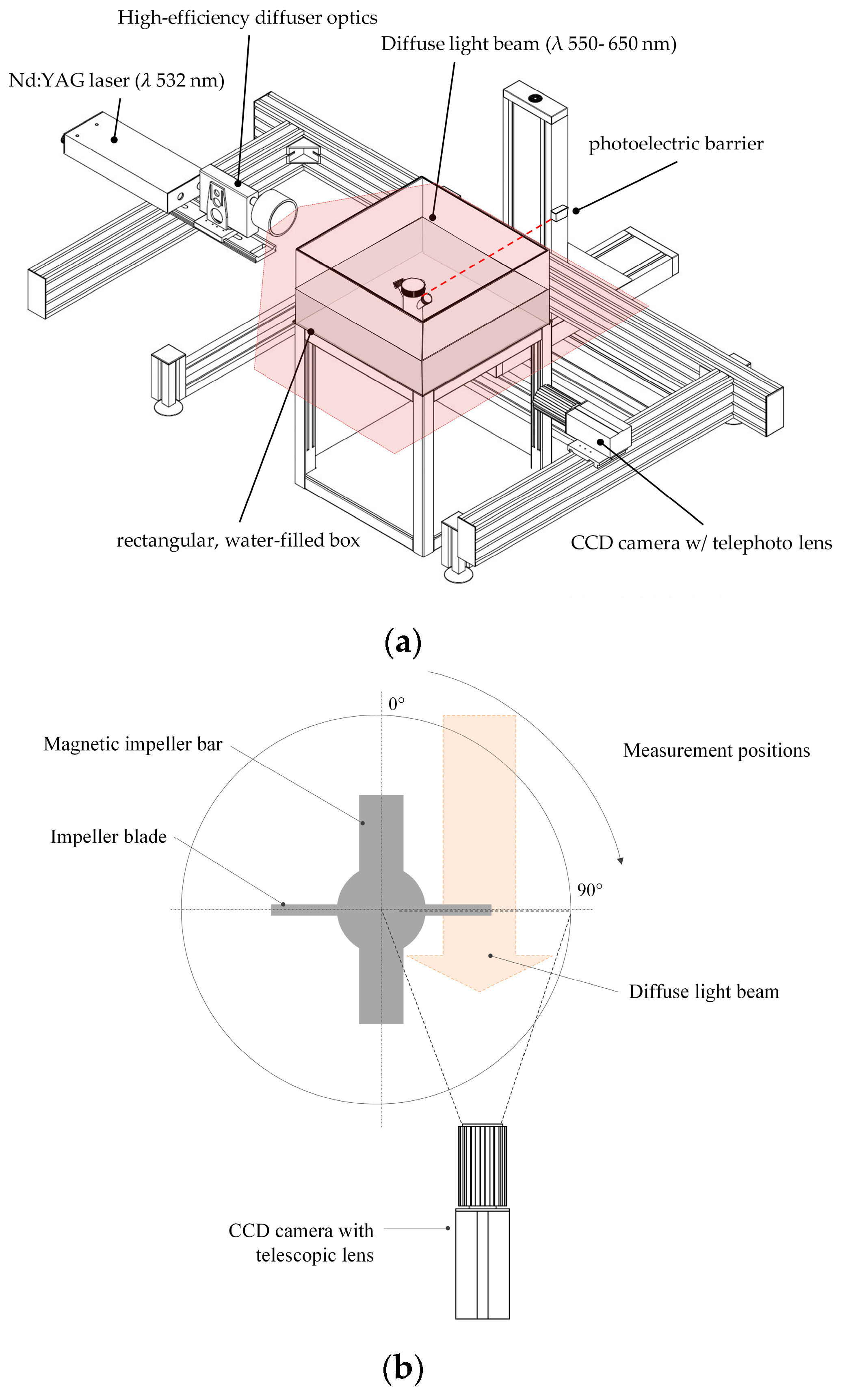
2.3.3. Microcarrier Measurement by Shadow Imaging (Shadowgraphy)
2.4. Cultivation Studies
2.4.1. Cells, Microcarriers, and Medium
2.4.2. Analytics
2.4.3. Spinner Flask Cultivations
3. Results and Discussion
3.1. Suspension Studies
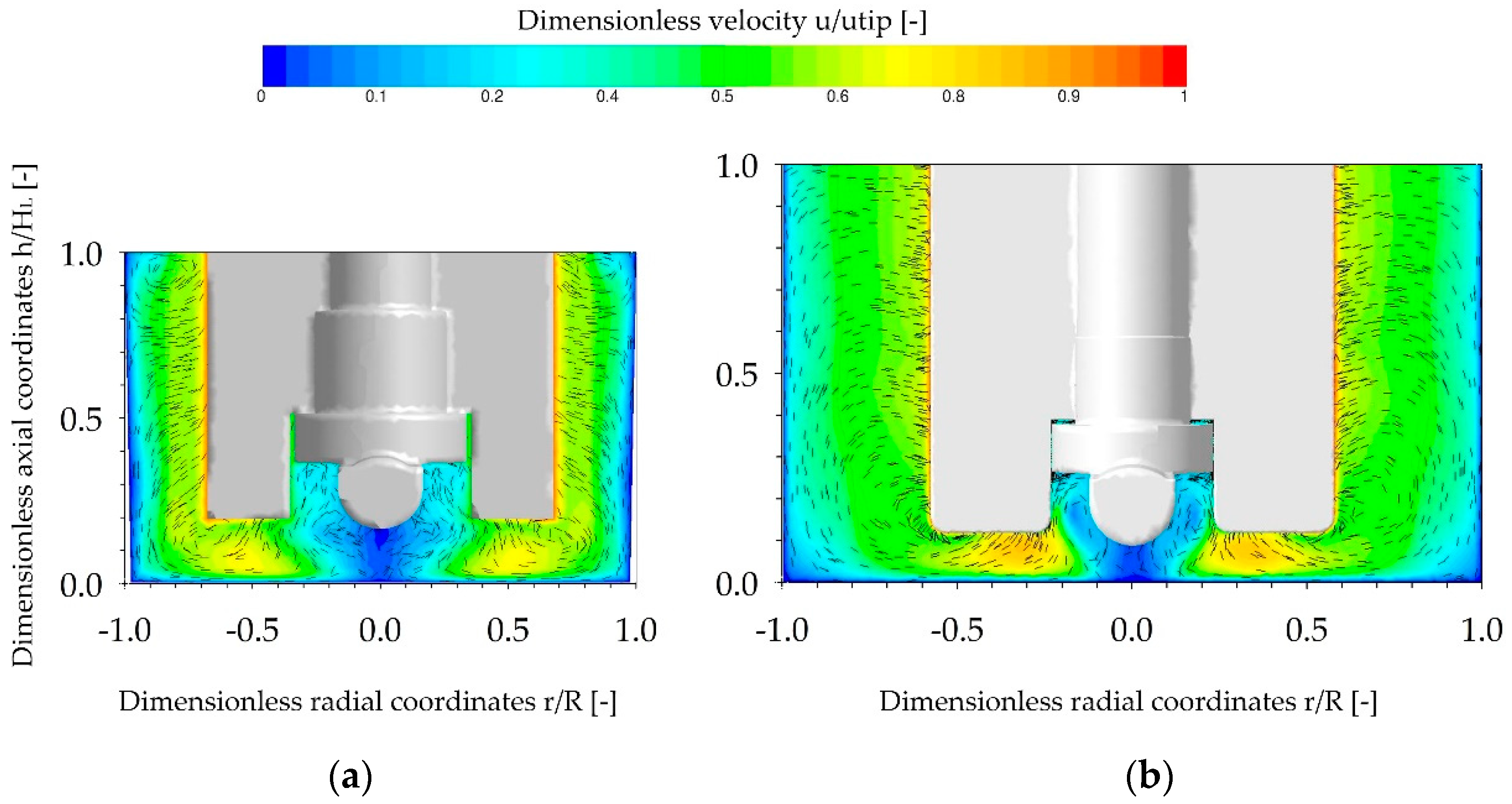
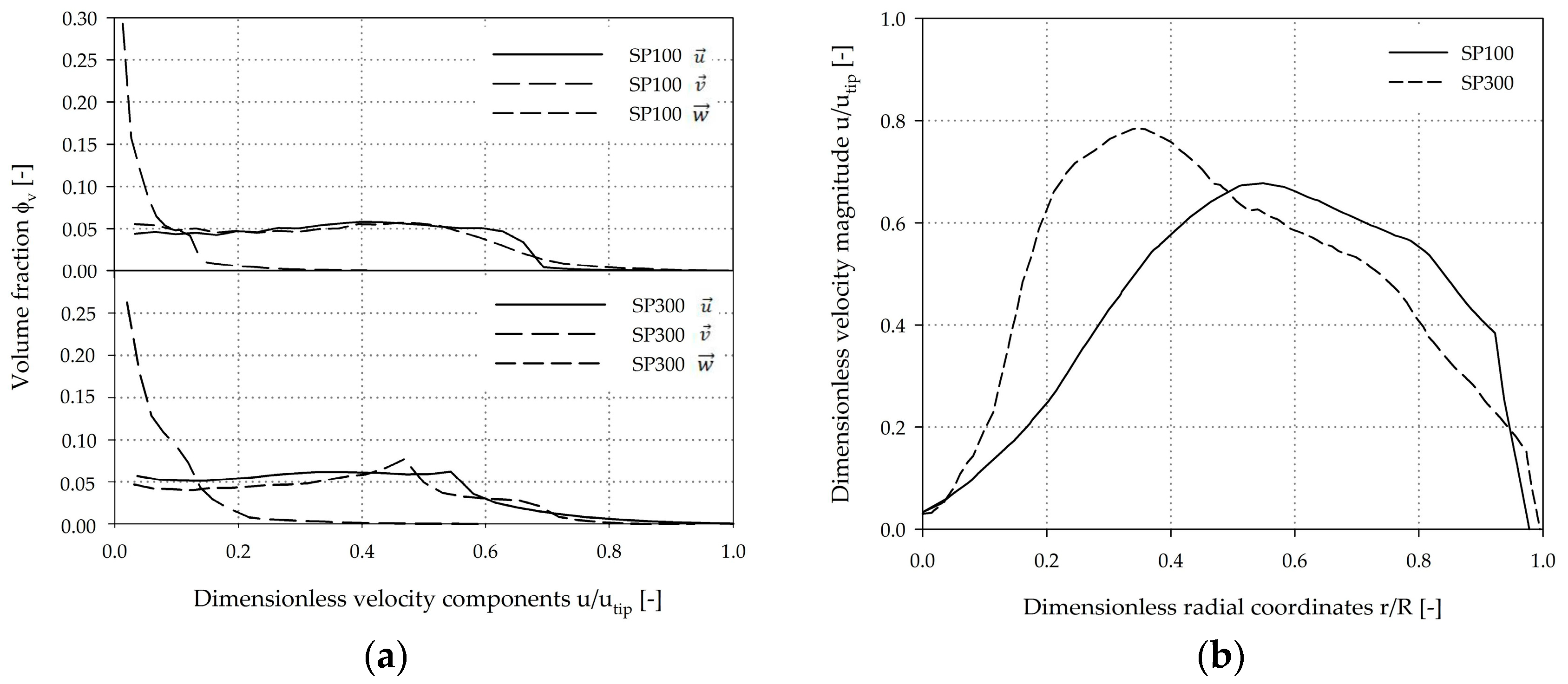
3.2. Single-Phase Fluid Flow Pattern
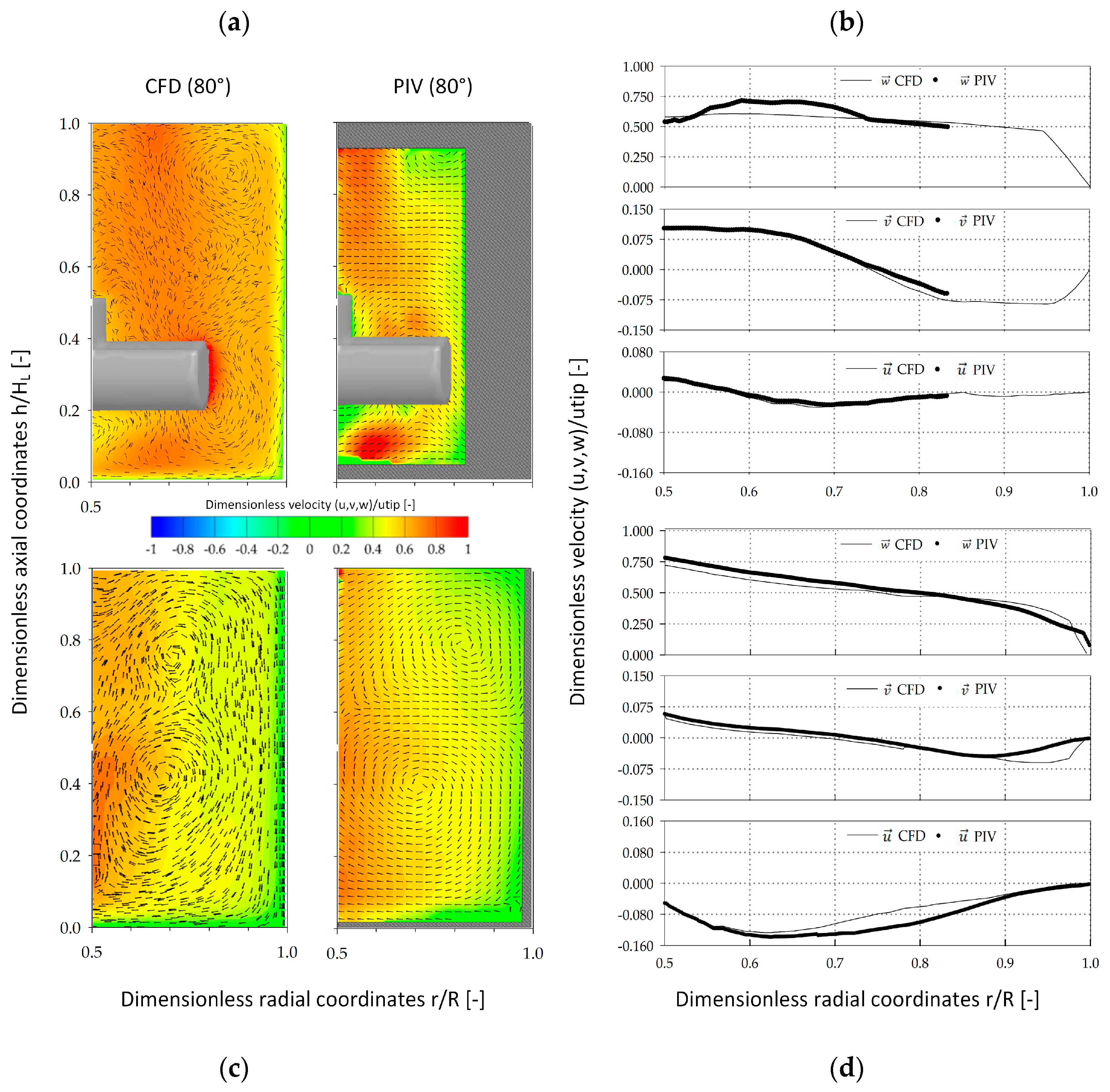
3.3. Fluid Flow Field Verification
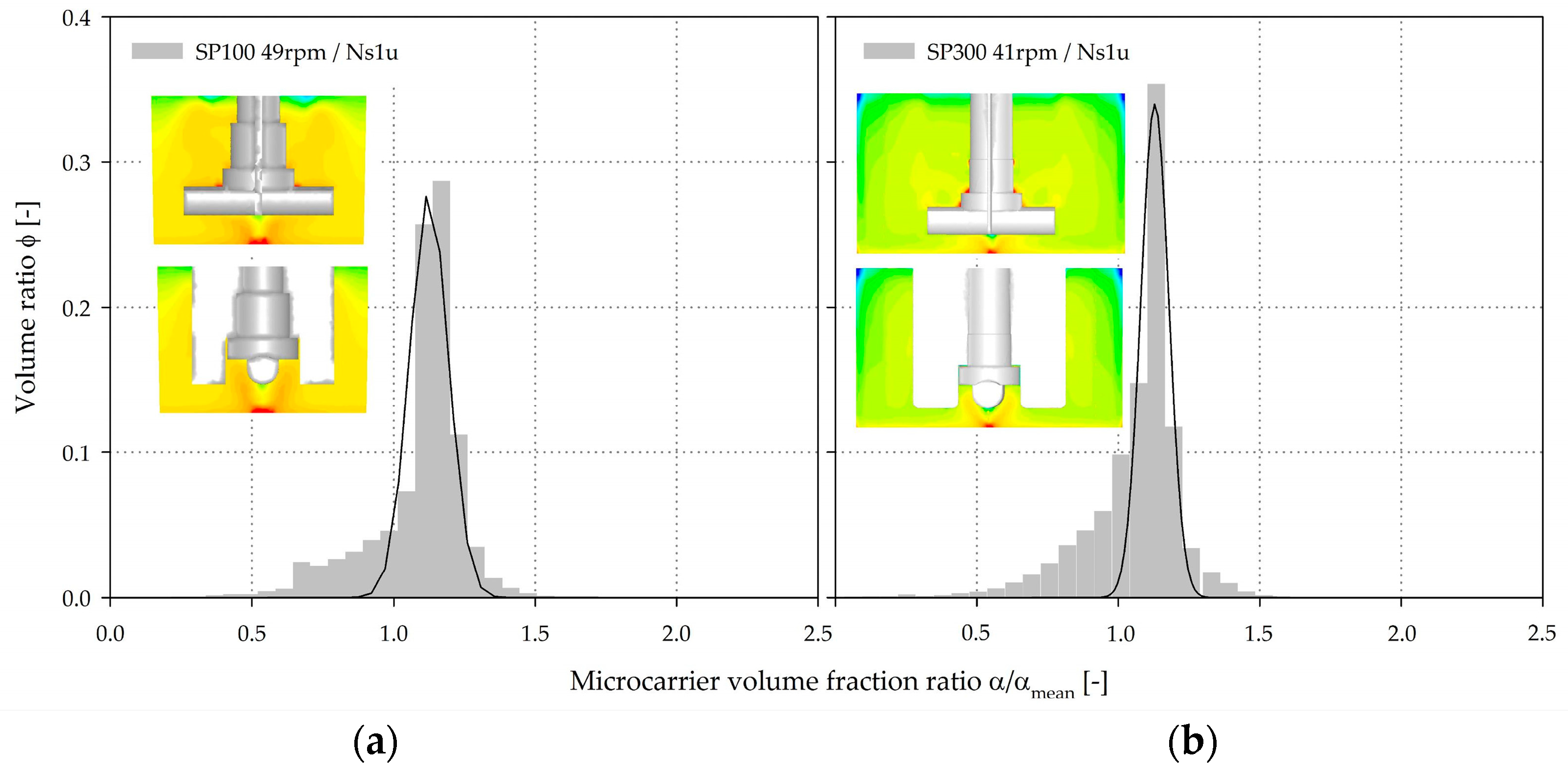
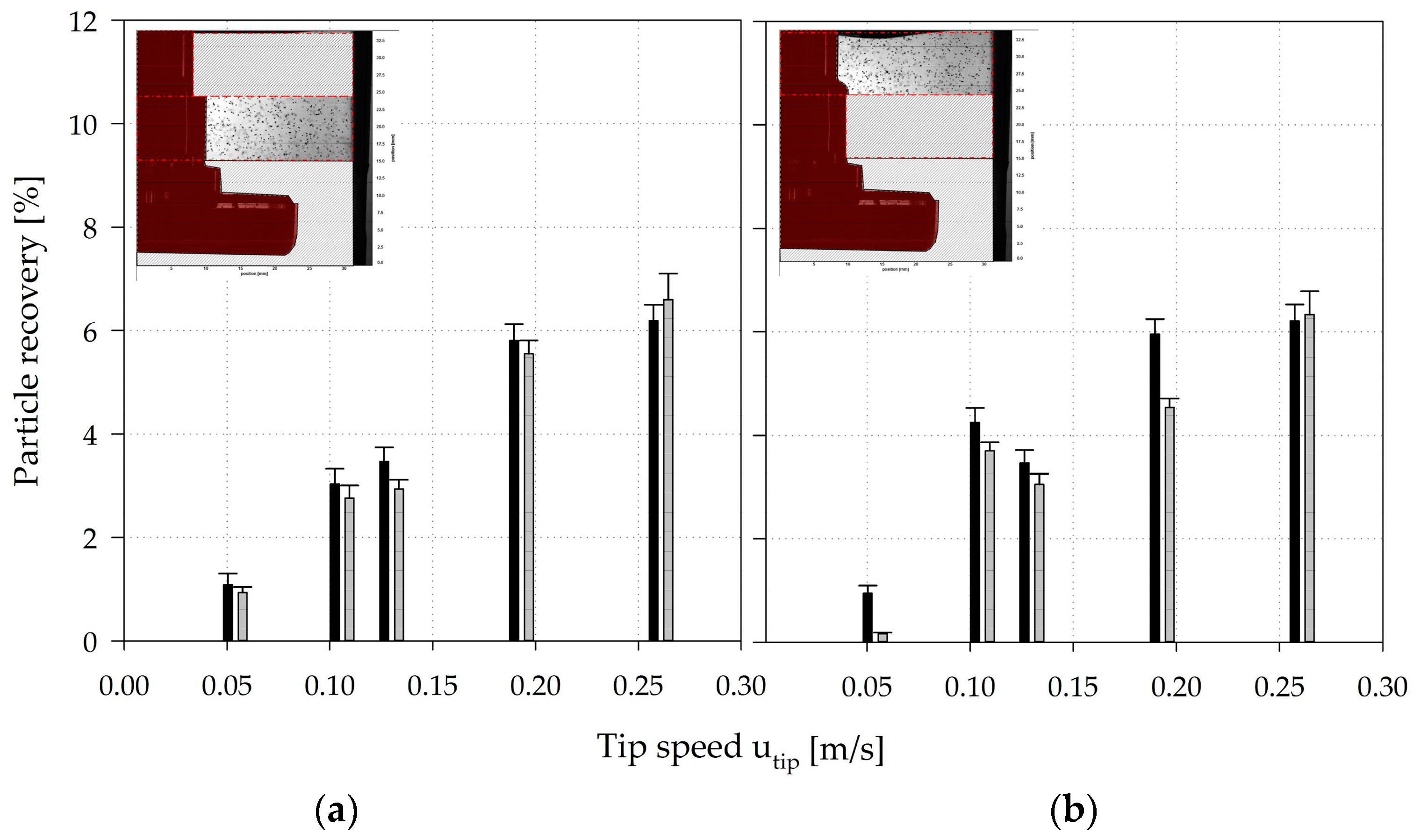
3.4. Microcarrier Distribution
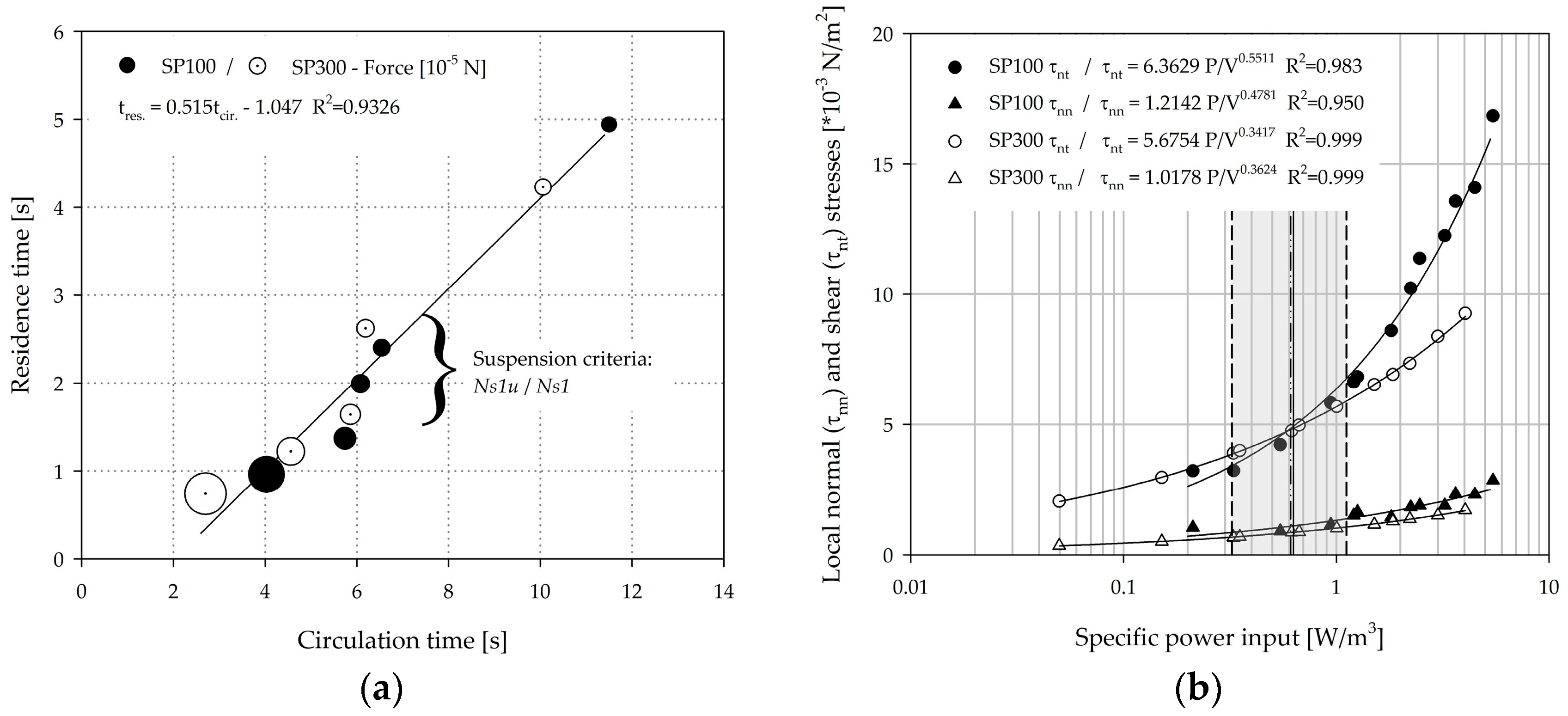
3.5. Circulation Times, Residence Times, and Hydrodynamic Stresses
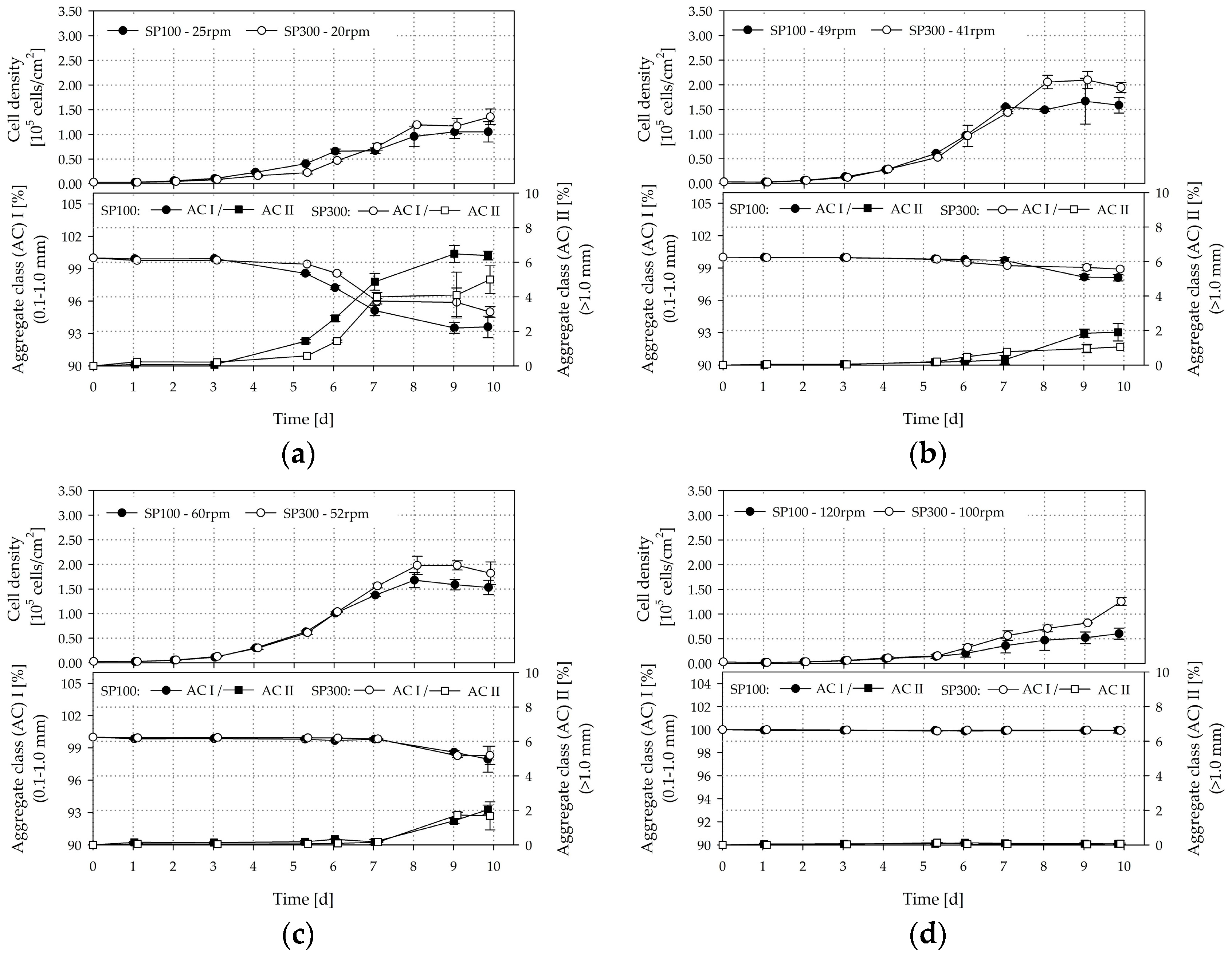
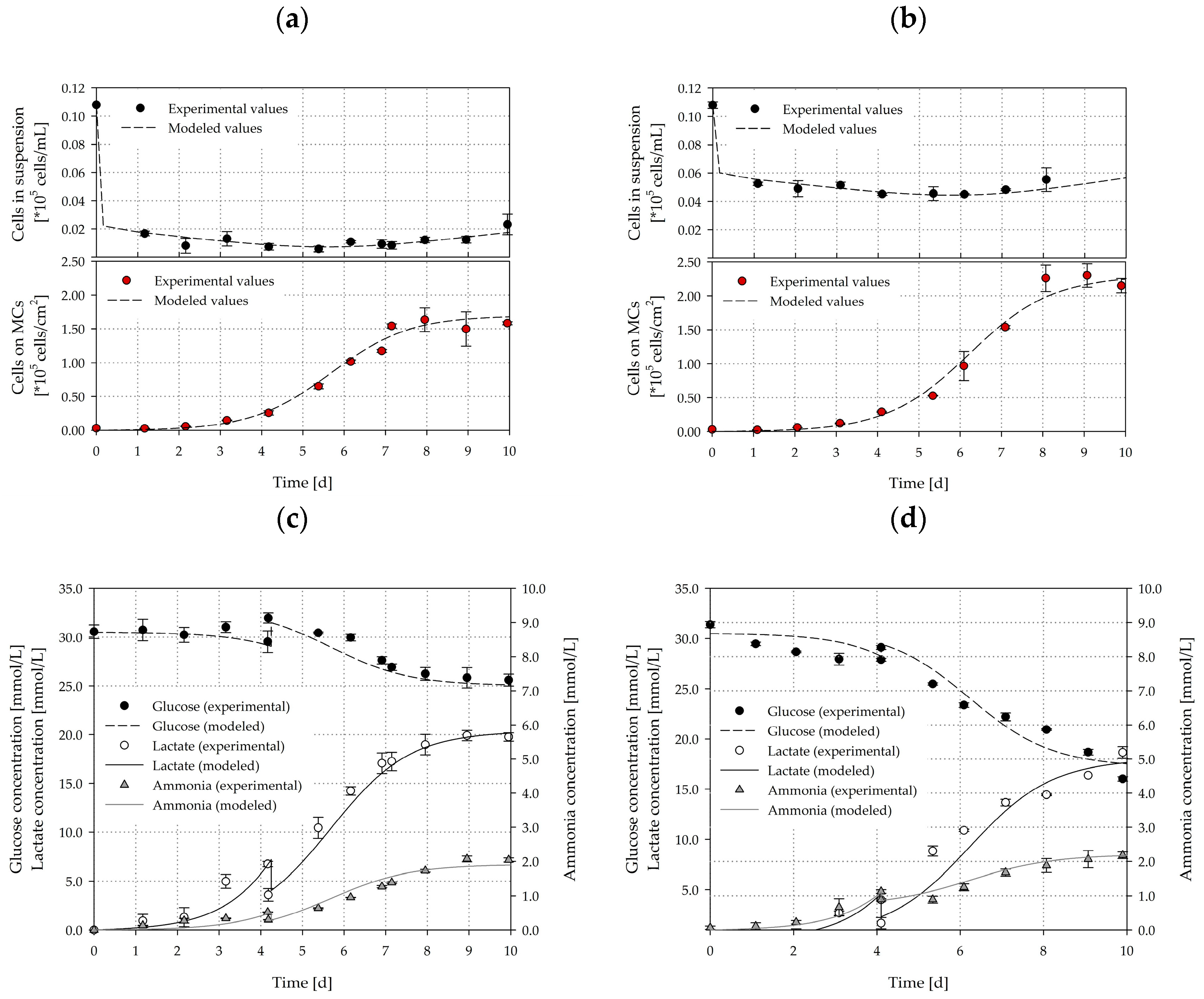
3.6. Cultivation Studies
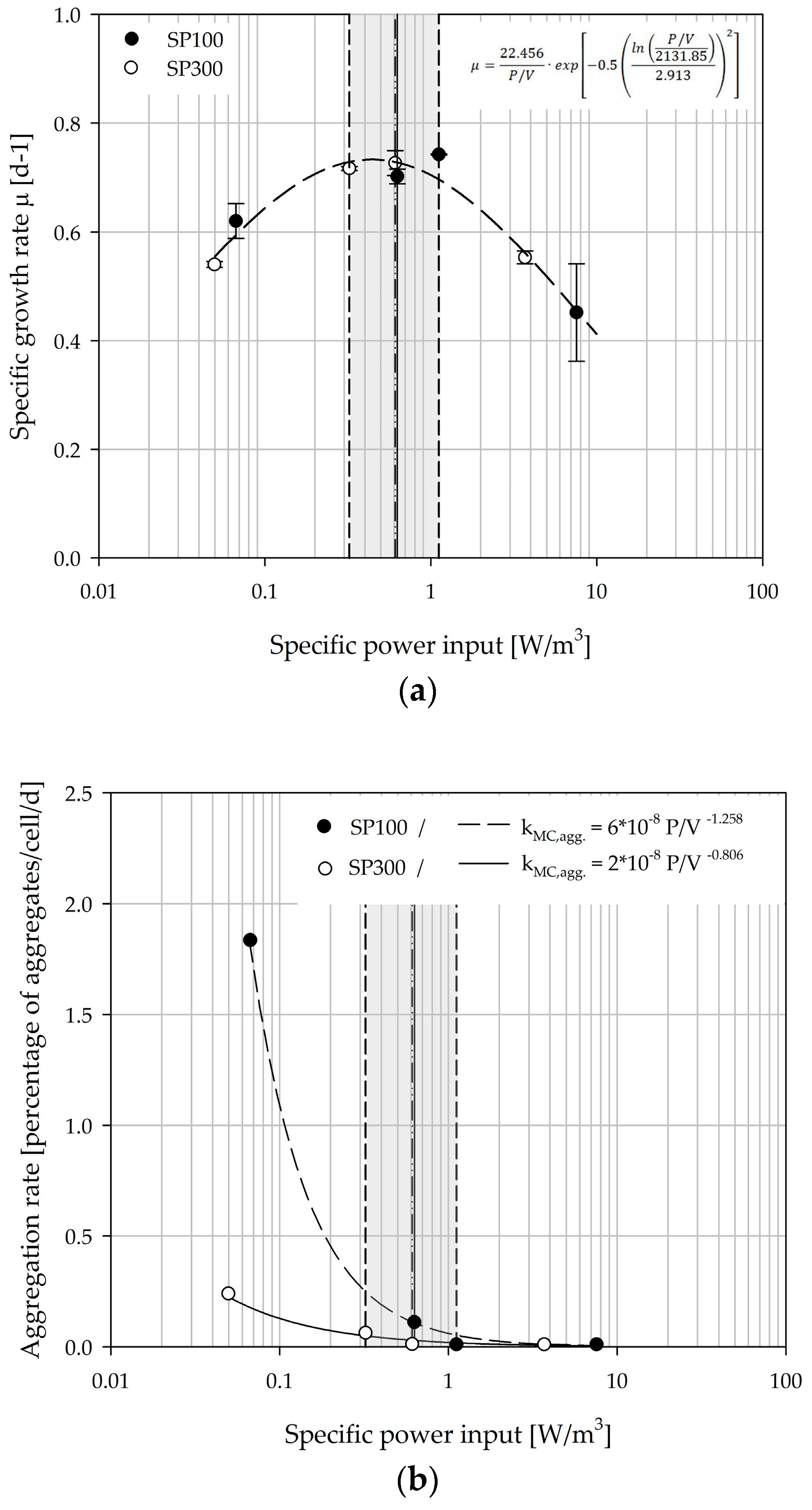
3.7. Growth Modelling
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Latin Symbols
| Amn | (mmol/L) | Ammonia concentration |
| Ap | (cm2/g) | Specific surface per unit mass MC |
| c | (m) | Off-bottom clearance |
| cMC | (g/L) | Microcarrier concentration |
| CD | (-) | Drag coefficient |
| d | (m) | Impeller diameter |
| dP | (m) | Particle diameter |
| D | (m) | Vessel diameter |
| EF | (-) | Expansion factor |
| (N) | Force (vector) | |
| (N) | Drag force (vector) | |
| g | (m/s2) | Gravitational acceleration |
| Glc | (mmol/L) | Glucose concentration |
| H | (m) | Vessel height |
| HL | (m) | Liquid height |
| kat | (d−1) | Cell attachment constant |
| kdet | (d−1) | Cell detachment constant |
| KAmn | (mmol/L) | Inhibition constant of ammonia |
| KGlc | (mmol/L) | Monod constant of glucose |
| KLac | (mmol/L) | Inhibition constant of lactate |
| Lac | (mmol/L) | Lactate concentration |
| mGlc | (mmol/cell/d) | Glucose consumption rate for maintenance |
| Ns1u, Ns1 | (rpm) | Suspension criteria |
| p | (Pa) | Pressure |
| PDL | (-) | Population doubling level |
| qAmn | (mmol/cell/d) | Specific ammonia production rate |
| −qGlc | (mmol/cell/d) | Specific glucose consumption rate |
| qLac | (mmol/cell/d) | Specific lactate production rate |
| Rep | (-) | Reynolds number for a particle |
| (m/s) | Velocity (vector) | |
| utip | (m/s) | Tip speed |
| (-) | Terminal velocity correlation for the solid phase (acc. to Symlal and O’Brien) | |
| Vmax | (m3) | Maximum working volume |
| XMC | (cells/cm2) | Cells on microcarrier |
| Xmax | (cm2) | Maximum growth surface |
| XSus | (cells/mL) | Cell in suspension |
| XV | (cells/mL) | Viable cells |
| x, y, z | (m) | Spatial co-ordinates |
| Yx/Glc | (cells/mmol) | Cell specific yield factor of glucose |
| Greek symbols | ||
| α | (-) | Phase volume fraction |
| αmean | (-) | Mean phase volume fraction |
| β | (°) | Impeller blade angle |
| μ | (d−1) | Specific growth rate |
| μmax | (d−1) | Maximum specific growth rate |
| ρ | (kg/m3) | Density |
| σ(Glc) | (-) | Simulation step response |
| (N/m2) | Reynolds stress tensor | |
| (N/m2) | Local shear stress | |
| (-) | Nabla operator |
References
- Heathman, T.R.J.; Nienow, A.W.; McCall, M.J.; Coopman, K.; Kara, B.; Hewitt, C.J. The translation of cell-based therapies: Clinical landscape and manufacturing challenges. Regen. Med. 2015, 10, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Ratcliffe, E.; Glen, K.E.; Naing, M.W.; Williams, D.J. Current status and perspectives on stem cell-based therapies undergoing clinical trials for regenerative medicine: Case studies. Br. Med. Bull. 2013, 108, 73–94. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.L.; Rao, M.S.; Deans, R.; Czermak, P. Manufacturing Cells for Clinical Use. Stem Cells Int. 2016, 2016, 1750697. [Google Scholar] [CrossRef]
- Trounson, A.; McDonald, C. Stem Cell Therapies in Clinical Trials: Progress and Challenges. Cell Stem Cell 2015, 17, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Yang, X.; Han, Z.; Qu, F.; Shao, L.; Shi, Y. Mesenchymal stem cells: A new trend for cell therapy. Acta Pharmacol. Sin. 2013, 34, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, B.M.; Kassem, M. Human mesenchymal stem cells: From basic biology to clinical applications. Gene Ther. 2008, 15, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Malik, N.N.; Durdy, M.B. Cell Therapy Landscape. In Translational Regenerative Medicine; Elsevier: Amsterdam, The Netherlands, 2015; pp. 87–106. Available online: http://linkinghub.elsevier.com/retrieve/pii/B9780124103962000074 (accessed on 3 January 2018).
- Simaria, A.S.; Hassan, S.; Varadaraju, H.; Rowley, J.; Warren, K.; Vanek, P.; Farid, S.S. Allogeneic cell therapy bioprocess economics and optimization: Single-use cell expansion technologies. Biotechnol. Bioeng. 2014, 111, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Jossen, V.; van den Bos, C.; Eibl, R.; Eibl, D. Manufacturing human mesenchymal stem cells at clinical scale: Process and regulatory challenges. Appl. Microbiol. Biotechnol. 2018, 102, 3981–3994. [Google Scholar] [CrossRef]
- Lodge, A.; Detela, G.; Barry, J.; Ginty, P.; Mount, N. Global Regulatory Perspective for MSCs. In Mesenchymal Stem Cells; Elsevier: Amsterdam, The Netherlands, 2017; pp. 243–287. [Google Scholar]
- Eibes, G.; dos Santos, F.; Andrade, P.Z.; Boura, J.S.; Abecasis, M.M.; da Silva, C.L.; Cabral, J.M. Maximizing the ex vivo expansion of human mesenchymal stem cells using a microcarrier-based stirred culture system. J. Biotechnol. 2010, 146, 194–197. [Google Scholar] [CrossRef]
- Lipsitz, Y.Y.; Milligan, W.D.; Fitzpatrick, I.; Stalmeijer, E.; Farid, S.S.; Tan, K.Y.; Smith, D.; Perry, R.; Carmen, J.; Chen, A.; et al. A roadmap for cost-of-goods planning to guide economic production of cell therapy products. Cytotherapy 2017, 19, 1383–1391. [Google Scholar] [CrossRef]
- Sharma, S.; Raju, R.; Shiu, S. Stem cell culture engineering—Process scale-up and beyond. Biotechnol. J. 2011, 6, 1317–1329. [Google Scholar] [CrossRef] [PubMed]
- Kino-Oka, M.; Mizutani, M. Cell Production System Based on Flexible Modular Platform. In Stem Cell Manufacturing; Elsevier: Amsterdam, The Netherlands, 2016; Chapter 7; pp. 161–169. [Google Scholar]
- Badenes, S.M.; Fernandes-Platzgummer, A.; Rodrigues, C.A.V.; Diogo, M.M.; da Silva, C.L.; Cabral, J.M.S. Microcarrier Culture Systems for Stem Cell Manufacturing. In Stem Cell Manufacturing; Elsevier: Amsterdam, The Netherlands, 2016; Chapter 4; pp. 77–104. [Google Scholar]
- Schirmaier, C.; Jossen, V.; Kaiser, S.C.; Jüngerkes, F.; Brill, S.; Safavi-Nab, A.; Siehoff, A.; van den Bos, C.; Eibl, D.; Eibl, R. Scale-up of adipose tissue-derived mesenchymal stem cell production in stirred single-use bioreactors under low-serum conditions. Eng. Life Sci. 2014, 14, 292–303. [Google Scholar] [CrossRef]
- Lawson, T.; Kehoe, D.E.; Schnitzler, A.C.; Rapiejko, P.J.; Der, K.A.; Philbrick, K.; Punreddy, S.; Rigby, S.; Smith, R.; Feng, Q.; et al. Process development for expansion of human mesenchymal stromal cells in a 50L single-use stirred tank bioreactor. Biochem. Eng. J. 2017, 120, 49–62. [Google Scholar] [CrossRef]
- Abraham, E.; Gupta, S.; Jung, S.; McAfee, E. Bioreactor for Scale-Up: Process Control. In Mesenchymal Stem Cells; Elsevier: Amsterdam, The Netherlands, 2017; pp. 139–177. [Google Scholar]
- Werner, S.; Kaiser, S.C.; Kraume, M.; Eibl, D. Computational fluid dynamics as a modern tool for engineering characterization of bioreactors. Pharm. Bioprocess 2014, 2, 85–99. [Google Scholar] [CrossRef]
- Kaiser, S.; Löffelholz, C.; Werner, S.; Eibl, D. CFD for characterizing standard and single-use stirred cell culture bioreactors. In Computational Fluid Dynamics; Minin, I.V., Minin, O.V., Eds.; InTech: Rijeka, Croatia, 2011; pp. 97–122. [Google Scholar]
- Eibl, R.; Kaiser, S.; Lombriser, R.; Eibl, D. Disposable bioreactors: The current state-of-the-art and recommended applications in biotechnology. Appl. Microbiol. Biotechnol. 2010, 86, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Sucosky, P.; Osorio, D.F.; Brown, J.B.; Neitzel, G.P. Fluid mechanics of a spinner-flask bioreactor. Biotechnol. Bioeng. 2004, 85, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Nienow, A.W.; Rielly, C.D.; Brosnan, K.; Bargh, N.; Lee, K.; Coopman, K.; Hewitt, C.J. The physical characterisation of a microscale parallel bioreactor platform with an industrial CHO cell line expressing an IgG4. Biochem. Eng. J. 2013, 76, 25–36. [Google Scholar] [CrossRef]
- Sharma, C.; Malhotra, D.; Rathore, A.S. Review of Computational fluid dynamics applications in biotechnology processes. Biotechnol. Prog. 2011, 27, 1497–1510. [Google Scholar] [CrossRef]
- Kaiser, S.; Jossen, V.; Schirmaier, C.; Eibl, D.; Brill, S.; van den Bos, C.; Eibl, R. Fluid Flow and Cell Proliferation of Mesenchymal Adipose-Derived Stem Cells in Small-Scale, Stirred, Single-Use Bioreactors. Chem. Ing. Tech. 2013, 85, 95–102. [Google Scholar] [CrossRef]
- Jossen, V.; Kaiser, S.C.; Schirmaier, C.; Herrmann, J.; Tappe, A.; Eibl, D.; Siehoff, A.; den Bos, C.V.; Eibl, R. Modification and qualification of a stirred single-use bioreactor for the improved expansion of human mesenchymal stem cells at benchtop scale. Pharm. Bioprocess 2014, 2, 311–322. [Google Scholar] [CrossRef]
- Jossen, V.; Schirmer, C.; Mostafa Sindi, D.; Eibl, R.; Kraume, M.; Pörtner, R.; Eibl, D. Theoretical and Practical Issues That Are Relevant When Scaling Up hMSC Microcarrier Production Processes. Stem Cells Int. 2016, 2016, 4760414. [Google Scholar] [CrossRef] [PubMed]
- Liovic, P.; Šutalo, I.D.; Stewart, R.; Glattauer, V.; Meagher, L. Fluid flow and stresses on microcarriers in spinner flask bioreactors. In Proceedings of the Ninth International Conference on CFD in the Minerals and Process Industries, Melbourne, Australia, 10–12 December 2012; pp. 1–6. [Google Scholar]
- Ismadi, M.-Z.; Hourigan, K.; Fouras, A. Experimental Characterisation of Fluid Mechanics in a Spinner Flask Bioreactor. Processes 2014, 2, 753–772. [Google Scholar] [CrossRef]
- Hutmacher, D.W.; Singh, H. Computational fluid dynamics for improved bioreactor design and 3D culture. Trends Biotechnol. 2008, 26, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.D.; Liovic, P.; Šutalo, I.D.; Stewart, R.L.; Glattauer, V.; Meagher, L. Characterisation of stresses on microcarriers in a stirred bioreactor. Appl. Math. Model. 2016, 40, 6787–6804. [Google Scholar] [CrossRef]
- Liovic, P.; Šutalo, I.D.; Meagher, L.; Lovrecz, G.O. Computations of flow environments in medium-scale stirred tank bioreactors for stem cell expansion. In Proceedings of the 2014 12th International Conference on Nanochannels, Microchannels, and Minichannels, Chicago, IL, USA, 3–7 August 2014; pp. 1–7. [Google Scholar]
- Julaey, M.; Hosseini, M.; Amani, H. Stem Cells Culture Bioreactor Fluid Flow, Shear Stress and Microcarriers Dispersion Analysis Using Computational Fluid Dynamics. J. Appl. Biotechnol. Rep. 2016, 3, 425–431. [Google Scholar]
- Symlal, M.; Rogers, W.; O’Brien, T.J. Computer Simulation of Bubbles in a Fluidized Bed. AIChE Symp. 1989, 85, 22–31. [Google Scholar]
- Schiller, L.; Naumann, Z. A drag coefficient correlation. Z. Ver. Deutsch. Ing. 1935, 77, 318. [Google Scholar]
- Ansys Fluent 13.0. Theory Guide; Ansys Inc.: Canonsburg, PA, USA, 2010; pp. 133–174.
- Möhler, L.; Bock, A.; Reichl, U. Segregated mathematical model for growth of anchorage-dependent MDCK cells in microcarrier culture. Biotechnol. Prog. 2008, 24, 110–119. [Google Scholar] [CrossRef]
- Bock, A.; Sann, H.; Schulze-Horsel, J.; Genzel, Y.; Reichl, U.; Möhler, L. Growth behavior of number distributed adherent MDCK cells for optimization in microcarrier cultures. Biotechnol. Prog. 2009, 25, 1717–1731. [Google Scholar] [CrossRef]
- Schop, D.; Janssen, F.W.; van Rijn, L.D.; Fernandes, H.; Bloem, R.M.; de Bruijn, J.D.; van Dijkhuizen-Radersma, R. Growth, metabolism, and growth inhibitors of mesenchymal stem cells. Tissue Eng. Part A 2009, 15, 1877–1886. [Google Scholar] [CrossRef]
- Zwietering, T.N. Suspending solid particles in liquid by agitators. Chem. Eng. Sci. 1958, 8, 244–253. [Google Scholar] [CrossRef]
- Liepe, F.; Sperling, R.; Jembere, S. Rührwerke: Theoretische Grundlagen, Auslegung und Bewertung; Fachhochschule Köthen Eigenverlag: Köthen, Germany, 1998; 548p. [Google Scholar]
- LaVision. ParticleMaster Shadow: Product-Manual; LaVision: Göttingen, Germany, 2015; pp. 1–110. [Google Scholar]
- Venkat, R.V.; Stock, L.R.; Chalmers, J.J. Study of hydrodynamics in microcarrier culture spinner vessels: A particle tracking velocimetry approach. Biotechnol. Bioeng. 1996, 49, 456–466. [Google Scholar] [CrossRef]
- Wollny, S. Experimentelle und Numerische Untersuchungen zur Partikelbeanspruchung in Gerührten (Bio-) Reaktoren. Ph.D. Thesis, Technische Universitaot Berlin, Berlin, Germany, 2010. [Google Scholar]
- Grein, T.A.; Leber, J.; Blumenstock, M.; Petry, F.; Weidner, T.; Salzig, D. Multiphase mixing characteristics in a microcarrier-based stirred tank bioreactor suitable for human mesenchymal stem cell expansion. Process Biochem. 2016, 51, 1109–1119. [Google Scholar] [CrossRef]
- Cierpka, K.; Elseberg, C.L.; Niss, K.; Kassem, M.; Salzig, D.; Czermak, P. hMSC Production in Disposable Bioreactors with Regards to GMP and PAT. Chem. Ing. Tech. 2013, 85, 67–75. [Google Scholar] [CrossRef]
- Langer, G.; Deppe, A. Zum Veständnis der hydrodynamischen Beanspruchung von Partikeln in turbulenten Rührerströmungen. Chem. Ing. Tech. 2000, 72, 31–41. [Google Scholar] [CrossRef]
- Weyand, B.; Reimers, K.; Vogt, P.M. Influences of Extracellular Matrix Properties and Flow Shear Stresses on Stem Cell Shape in a Three-Dimensional Dynamic Environment. In Proceedings of the 8th International Conference on Cell & Stem Cell Engineering (ICCE), Dublin, Ireland, 11–12 June 2011; pp. 47–50. [Google Scholar]
- Weyand, B.; Israelowitz, M.; von Schroeder, H.P.; Vogt, P.M. Fluid Dynamics in Bioreactor Design: Considerations for the Theoretical and Practical Approach. Adv. Biochem. Eng. Biotechnol. 2009, 112, 251–268. [Google Scholar] [PubMed]
- Weyand, B.; Kasper, C.; Israelowitz, M.; Gille, C.; von Schroeder, H.P.; Reimers, K.; Vogt, P.M. A Differential Pressure Laminar Flow Reactor Supports Osteogenic Differentiation and Extracellular Matrix Formation from Adipose Mesenchymal Stem Cells in a Macroporous Ceramic Scaffold. Biores. Open Access 2012, 1, 145–157. [Google Scholar] [CrossRef]
- Yeatts, A.B.; Choquette, D.T.; Fisher, J.P. Bioreactors to influence stem cell fate: Augmentation of mesenchymal stem cell signaling pathways via dynamic culture systems. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 2470–2480. [Google Scholar] [CrossRef]
- Yeatts, A.B.; Fisher, J.P. Bone tissue engineering bioreactors: Dynamic culture and the influence of shear stress. Bone 2011, 48, 171–181. [Google Scholar] [CrossRef]
- Nienow, A. Scale-Up, Stirred Tank Reactors. In Encyclopedia of Industrial Biotechnology: Bioprocess, Bioseparation, and Cell Technology; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2010; Volume 16, pp. 1–14. [Google Scholar]
- Nienow, A.W. Reactor engineering in large scale animal cell culture. Cytotechnology 2006, 50, 9–33. [Google Scholar] [CrossRef]
- Thomas, C.R.; Zhang, Z. The Effect of Hydrodynamics on Biological Materials. In Advances in Bioprocess Engineering; Galindo, E., Ramirez, O.T., Eds.; Springer: Dordrecht, The Netherlands, 1998; pp. 137–170. [Google Scholar]
- Croughan, M.S.; Hamel, J.-F.; Wang, D.I.C. Hydrodynamic effects on animal cells grown in microcarrier cultures. Biotechnol. Bioeng. 2006, 95, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Ponnuru, K.; Wu, J.; Ashok, P.; Tzanakakis, E.; Furlani, E.P. Analysis of Stem Cell Culture Performance in a Microcarrier Bioreactor System. Proc. Int. NSTI Nanotech. Conf. 2014, 2, 132–135. [Google Scholar]
- Takahashi, I.; Sato, K.; Mera, H.; Wakitani, S.; Takagi, M. Effects of agitation rate on aggregation during beads-to-beads subcultivation of microcarrier culture of human mesenchymal stem cells. Cytotechnology 2016, 29, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, C.; Balandras, F.; Guedon, E.; Olmos, E.; Chevalot, I.; Marc, A. Limiting cell aggregation during mesenchymal stem cell expansion on microcarriers. Biotechnol. Prog. 2012, 28, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, Q.A.; Ruck, S.; Hanga, M.P.; Heathman, T.R.; Coopman, K.; Nienow, A.W.; Williams, D.J.; Hewitt, C.J. Qualiative and quantitative demonstration of bead-to-bead transfer with bone marrow-derived human mesenchymal stem cells on microcarriers: Utilising the phenomenon to improve culture performance. Biochem. Eng. J. 2017, 15, 11–21. [Google Scholar]
- Schop, D.; van Dijkhuizen-Radersma, R.; Borgart, E.; Janssen, F.W.; Rozemuller, H.; Prins, H.J.; de Bruijn, J.D. Expansion of human mesenchymal stromal cells on microcarriers: Growth and metabolism. J. Tissue Eng. Regen. Med. 2010, 4, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Higuera, G.; Schop, D.; Janssen, F.; van Dijkhuizen-Radersma, R.; van Boxtel, T.; van Blitterswijk, C.A. Quantifying In Vitro Growth and Metabolism Kinetics of Human Mesenchymal Stem Cells Using a Mathematical Model. Tissue Eng. Part A 2009, 15, 2653–2663. [Google Scholar] [CrossRef]
- Balducci, L.; Blasi, A.; Saldarelli, M.; Soleti, A.; Pessina, A.; Bonomi, A.; Coccè, V.; Dossena, M.; Tosetti, V.; Ceserani, V.; et al. Immortalization of human adipose-derived stromal cells: Production of cell lines with high growth rate, mesenchymal marker expression and capability to secrete high levels of angiogenic factors. Stem Cell Res. Ther. 2014, 5, 63. [Google Scholar] [CrossRef]
- Leber, J.; Barekzai, J.; Blumenstock, M.; Pospisil, B.; Salzig, D.; Czermak, P. Microcarrier choice and bead-to-bead transfer for human mesenchymal stem cells in serum-containing and chemically defined media. Process Biochem. 2017, 59, 255–265. [Google Scholar] [CrossRef]
- Yin, D.; Wells, J.; Clinton, J.; Zou, C. Comparative analysis of cell proliferation, immunosuppressive action, and multi-lineage differentiation of immortalized MSC and MSC from bone marrow, adipose tissue, and umbilical cord blood. In Proceedings of the International Society for Stem Cell Research Conference, Toronto, ON, Canada, 15–18 June 2010. [Google Scholar]
- Wolbank, S.; Stadler, G.; Peterbauer, A.; Gillich, A.; Karbiener, M.; Streubel, B.; Wieser, M.; Katinger, H.; van Griensven, M.; Redl, H.; et al. Telomerase immortalized human amnion- and adipose-derived mesenchymal stem cells: Maintenance of differentiation and immunomodulatory characteristics. Tissue Eng. Part A 2009, 15, 1843–1854. [Google Scholar] [CrossRef]
- Bourin, P.; Bunnell, B.A.; Casteilla, L.; Dominici, M.; Katz, A.J.; March, K.L.; Redl, H.; Rubin, J.P.; Yoshimura, K.; Gimble, J.M. Stromal cells from the adipose tissue-derived stromal vascular fraction and culture expanded adipose tissue-derived stromal/stem cells: A joint statement of the International Federation for Adipose Therapeutics and Science (IFATS) and the International Society for Cellular Therapy. Cytotherapy 2013, 15, 641–648. [Google Scholar] [CrossRef] [PubMed]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physical Dimension/Ratios | Corning® 125 mL Spinner | Corning® 500 mL Spinner | |
|---|---|---|---|
| D * | [mm] | 63.5 | 87.3 |
| HL ** | [mm] | 41 | 52 |
| H * | [mm] | 145 | 203 |
| Vmax * | [mL] | 100 | 300 |
| dmax * | [mm] | 41.5 | 50.3 |
| C ** | [mm] | 8.0 | 8.0 |
| β | [°] | 90 | 90 |
| H/D | [-] | 0.65 | 0.60 |
| d/D | [-] | 0.68 | 0.58 |
| c/D | [-] | 0.12 | 0.07 |
| Parameter | Values | Reference |
|---|---|---|
| −qGlc (pmol/cell/d) | 9.8–35 | This study |
| qLac (pmol/cell/d) | 20–89 | This study |
| qAmn (pmol/cell/d) | 6–19 | This study |
| kat (d−1) | 0.033–0.05 | This study |
| kdet (d−1) | 0.002–0.01 | This study |
| KGlc (mmol/L) | 0.4 | [37,39] |
| KLac (mmol/L) | 35–50 | [37,39] |
| KAmn (mmol/L) | 8–10 | [37,39] |
| N | utip | Re | P/V | lλ (a) | LSS (b) | LNS (c) | F (d) |
|---|---|---|---|---|---|---|---|
| (rpm) | (m/s) | (-) | (W/m3) | (μm) | (10−3 N/m2) | (10−3 N/m2) | (10−5 N) |
| Corning® 125 mL spinner (SP100): | |||||||
| 25 | 0.054 | 715 | 0.07 | 130/530 | 3.21/69.41 | 1.04/43.17 | 0.75 |
| 49 | 0.106 | 1402 | 0.63 | 66/228 | 4.96/187.00 | 1.15/109.00 | 0.85 |
| 60 | 0.130 | 1717 | 1.12 | 60/191 | 6.62/232.37 | 1.51/127.20 | 0.91 |
| 120 | 0.261 | 3434 | 7.56 | 30/111 | 13.55/437.69 | 2.33/277.44 | 1.82 |
| Corning® 500 mL spinner (SP300): | |||||||
| 20 | 0.053 | 841 | 0.05 | 136/546 | 2.05/214.40 | 0.35/138.86 | 0.83 |
| 41 | 0.108 | 1724 | 0.33 | 76/295 | 4.00/481.99 | 0.69/362.76 | 0.89 |
| 52 | 0.137 | 2186 | 0.61 | 66/282 | 4.98/680.55 | 0.88/473.87 | 1.04 |
| 100 | 0.263 | 4204 | 3.70 | 47/181 | 9.28/1352.86 | 1.71/874.34 | 2.10 |
| N | Living xmax | EF | µ/td | −qgluc | qlac | qNH4+ |
|---|---|---|---|---|---|---|
| (rpm) | (105 Cells/mL) (105 cells/cm2) | (-) | (d−1) (d) | (pmol cell−1 d−1) | ||
| Corning® 125 mL Spinner (SP100): | ||||||
| 25 | 4.49 ± 0.06 1.05 ± 0.06 | 35.03 | 0.62 ± 0.03 1.12 ± 0.06 | −13.21 ± 2.27 | 20.65 ± 2.73 | 8.78 ± 0.28 |
| 49 | 6.01 ± 0.12 1.67 ± 0.12 | 55.62 | 0.70 ± 0.01 0.99 ± 0.02 | −10.55 ± 1.59 | 35.22 ± 1.91 | 6.09 ± 0.42 |
| 60 | 6.25 ± 0.35 1.68 ± 0.36 | 56.01 | 0.74 ± 0.01 0.93 ± 0.01 | −9.80 ± 0.76 | 30.28 ± 1.01 | 6.20 ± 0.34 |
| 120 | 2.17 ± 0.40 0.60 ± 0.04 | 20.11 | 0.45 ± 0.09 1.53 ± 0.38 | −35.00 ± 1.61 | 88.78 ± 5.21 | 16.48 ± 0.25 |
| Corning® 500 mL spinner (SP300): | ||||||
| 20 | 4.88 ± 0.57 1.36 ± 0.57 | 45.20 | 0.54 ± 0.01 1.28 ± 0.01 | −20.98 ± 0.93 | 28.60 ± 9.86 | 14.71 ± 0.15 |
| 41 | 8.51 ± 0.16 2.46 ± 0.16 | 81.92 | 0.72 ± 0.01 0.97 ± 0.01 | −15.47 ± 0.59 | 40.63 ± 1.78 | 10.64 ± 0.54 |
| 52 | 8.77 ± 0.66 2.43 ± 0.66 | 81.14 | 0.73 ± 0.02 0.95 ± 0.03 | −11.75 ± 1.23 | 35.29 ± 3.28 | 9.73 ± 0.42 |
| 100 | 4.51 ± 0.29 1.25 ± 0.29 | 41.76 | 0.55 ± 0.01 1.25 ± 0.03 | −20.76 ± 9.84 | 88.56 ± 2.09 | 18.96 ± 1.39 |
| Marker | Inoculum | SP100 | SP300 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| T-flasks | 25 rpm | 49 rpm | 60 rpm | 120 rpm | 20 rpm | 41 rpm | 52 rpm | 100 rpm | |
| (%) | (%) | (%) | |||||||
| CD14− | 2.7 | 1.7 ± 0.2 | 1.3 ± 0.2 | 1.0 ± 0.3 | 1.2 ± 0.0 | 1.9 ± 0.3 | 1.5 ± 0.3 | 1.3 ± 0.1 | 1.2 ± 0.1 |
| (p 0.518) | (p 0.100) | (p 0.442) | (p 0.493) | (p > 0.05) | (p > 0.05) | (p > 0.05) | (p > 0.05) | ||
| CD20− | 2.7 | 1.7 ± 0.2 | 1.3 ± 0.2 | 1.0 ± 0.3 | 1.2 ± 0.0 | 1.9 ± 0.3 | 1.5 ± 0.3 | 1.3 ± 0.1 | 1.2 ± 0.1 |
| (p 0.518) | (p 0.100) | (p 0.442) | (p 0.493) | (p > 0.05) | (p > 0.05) | (p > 0.05) | (p > 0.05) | ||
| CD34− | 2.7 | 1.7 ± 0.2 | 1.3 ± 0.2 | 1.0 ± 0.3 | 1.2 ± 0.0 | 1.9 ± 0.3 | 1.5 ± 0.3 | 1.3 ± 0.1 | 1.2 ± 0.1 |
| (p 0.518) | (p 0.100) | (p 0.442) | (p 0.493) | (p > 0.05) | (p > 0.05) | (p > 0.05) | (p > 0.05) | ||
| CD45− | 2.7 | 1.7 ± 0.2 | 1.3 ± 0.2 | 1.0 ± 0.3 | 1.2 ± 0.0 | 1.9 ± 0.3 | 1.5 ± 0.3 | 1.3 ± 0.1 | 1.2 ± 0.1 |
| (p 0.518) | (p 0.100) | (p 0.442) | (p 0.493) | (p > 0.05) | (p > 0.05) | (p > 0.05) | (p > 0.05) | ||
| CD73+ | 99.4 | 99.6 ± 0.0 | 99.8 ± 0.0 | 97.1 ± 2.8 | 99.6 ± 0.2 | 99.5 ± 0.1 | 99.7 ± 0.0 | 99.8 ± 0.1 | 99.7 ± 0.0 |
| (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | ||
| CD90+ | 96.2 | 97.7 ± 0.4 | 98.9± 0.0 | 98.7 ± 0.2 | 97.6 ± 0.7 | 95.0 ± 0.0 | 96.1 ± 0.8 | 97.5 ± 0.4 | 96.8 ± 0.1 |
| (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | ||
| CD105+ | 99.3 | 91.7 ± 1.3 | 94.6 ± 0.4 | 94.0 ± 0.8 | 96.4 ± 2.1 | 94.0 ± 0.5 | 91.4 ± 2.4 | 97.6 ± 0.2 | 98.7 ± 0.4 |
| (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | (p < 0.05) | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jossen, V.; Eibl, R.; Kraume, M.; Eibl, D. Growth Behavior of Human Adipose Tissue-Derived Stromal/Stem Cells at Small Scale: Numerical and Experimental Investigations. Bioengineering 2018, 5, 106. https://doi.org/10.3390/bioengineering5040106
Jossen V, Eibl R, Kraume M, Eibl D. Growth Behavior of Human Adipose Tissue-Derived Stromal/Stem Cells at Small Scale: Numerical and Experimental Investigations. Bioengineering. 2018; 5(4):106. https://doi.org/10.3390/bioengineering5040106
Chicago/Turabian StyleJossen, Valentin, Regine Eibl, Matthias Kraume, and Dieter Eibl. 2018. "Growth Behavior of Human Adipose Tissue-Derived Stromal/Stem Cells at Small Scale: Numerical and Experimental Investigations" Bioengineering 5, no. 4: 106. https://doi.org/10.3390/bioengineering5040106
APA StyleJossen, V., Eibl, R., Kraume, M., & Eibl, D. (2018). Growth Behavior of Human Adipose Tissue-Derived Stromal/Stem Cells at Small Scale: Numerical and Experimental Investigations. Bioengineering, 5(4), 106. https://doi.org/10.3390/bioengineering5040106