Honey-Based Templates in Wound Healing and Tissue Engineering



Abstract
1. Introduction
2. In Vitro and In Vivo Evidence of the Beneficial Effects of Honey in Wounds
2.1. Antibacterial and Antibiofilm Effects of Honey
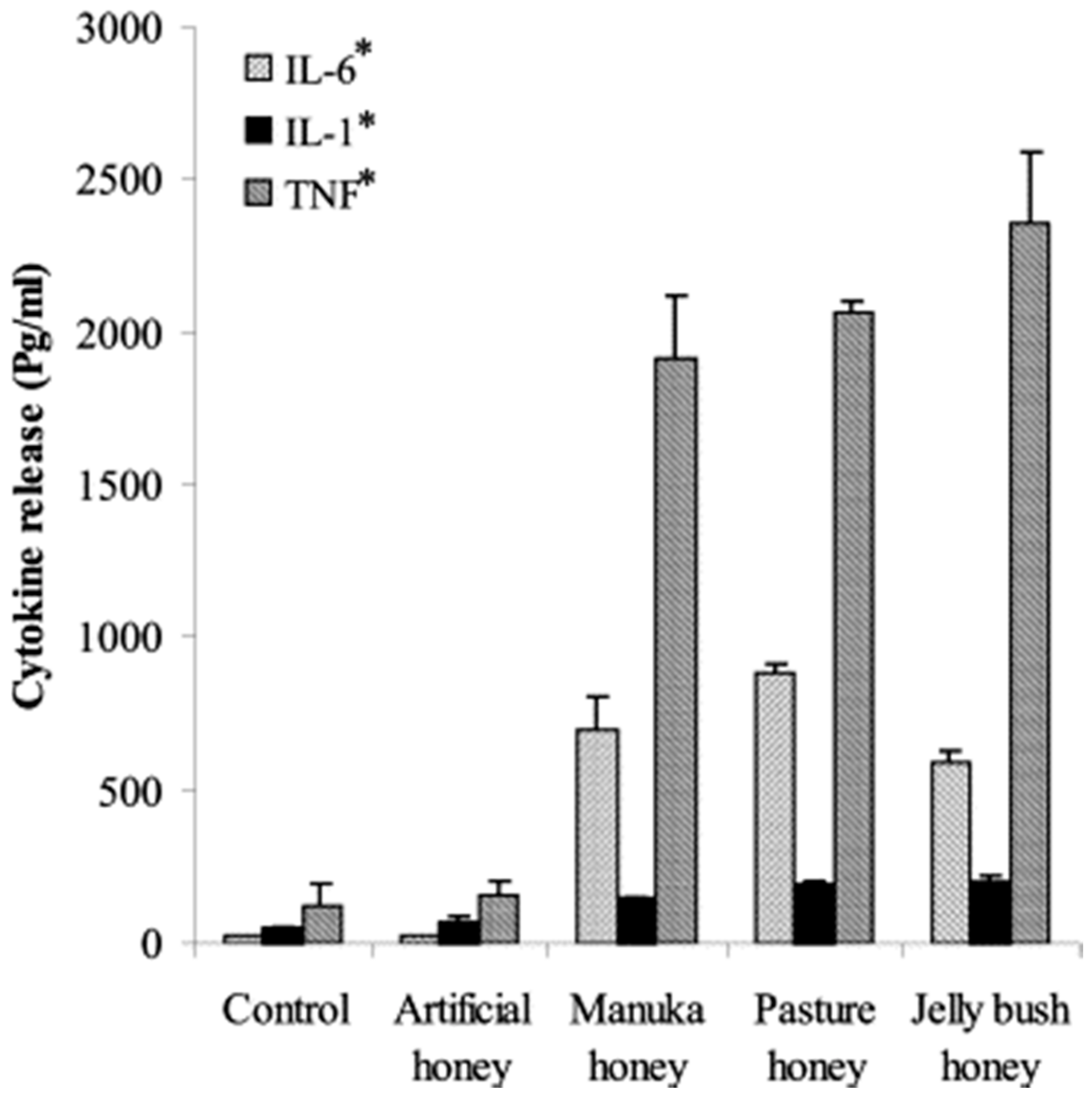
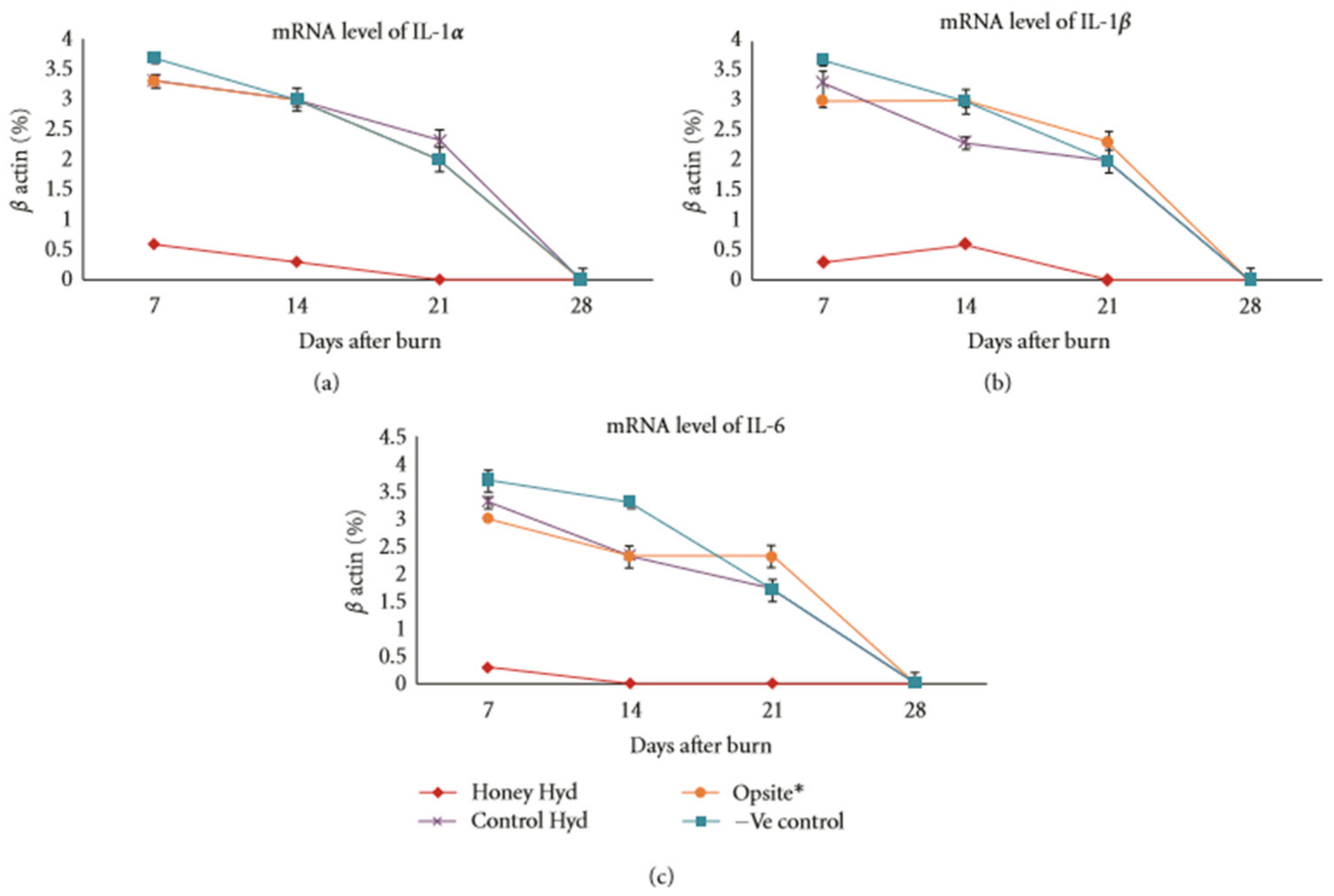
2.2. Immunomodulatory Effects of Honey
2.3. Wound Closure Effects of Honey
3. Cautionary Evidence of Cytotoxicity
4. Honey in Tissue Engineering
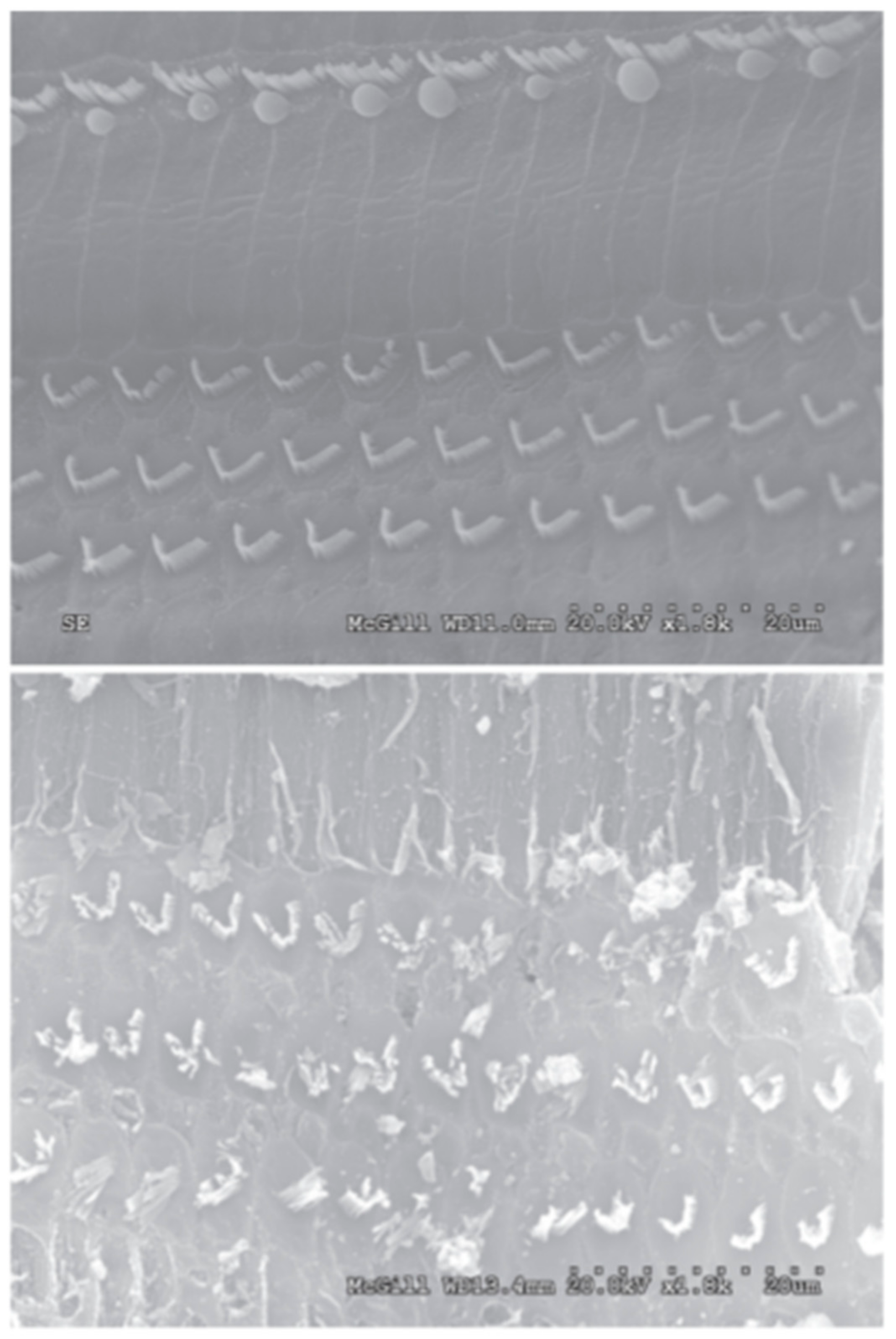
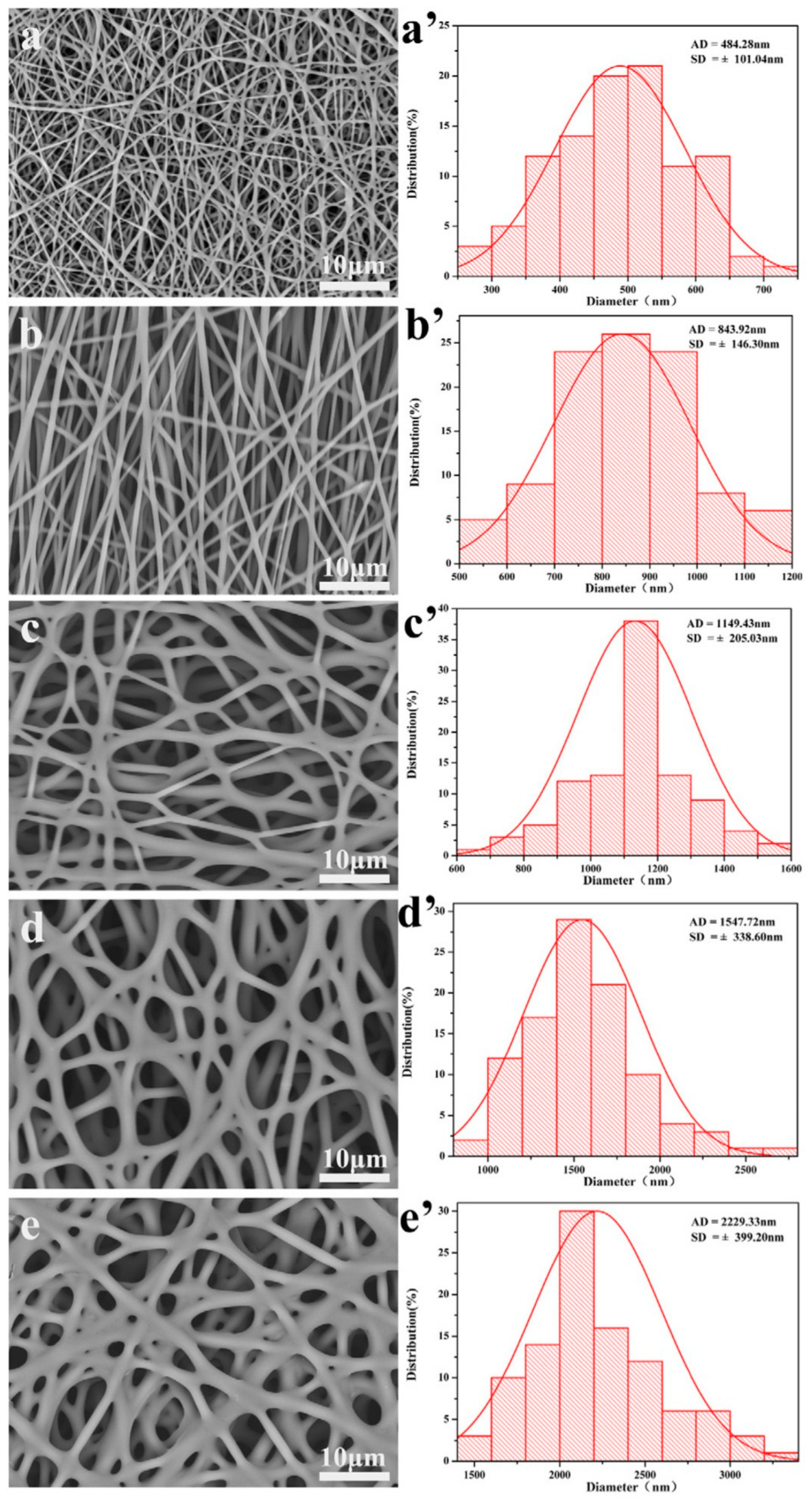
4.1. Electrospun Templates
4.2. Cryogels
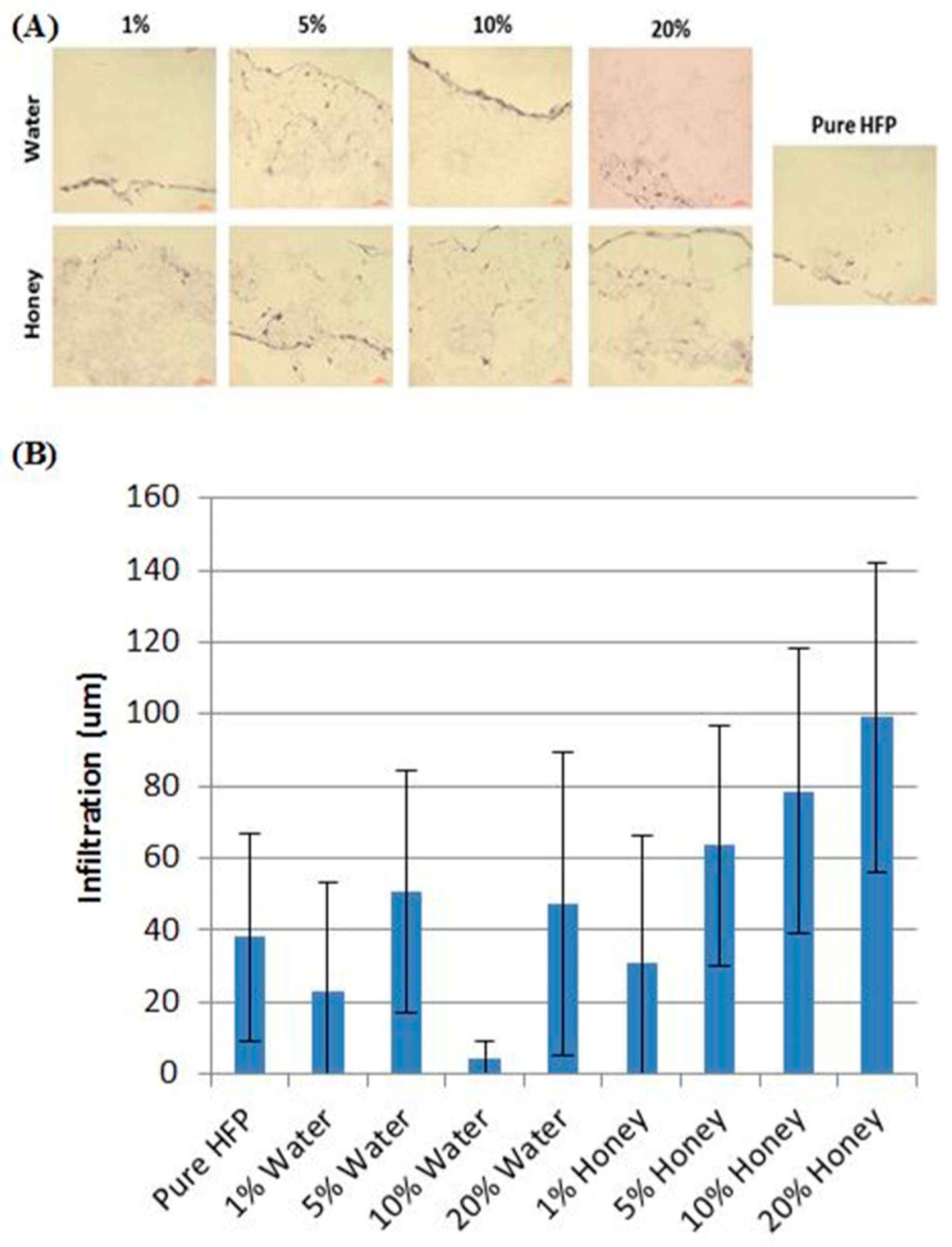
4.3. Hydrogels
5. Commercialization
6. The Future of Honey in Tissue Engineering
7. Conclusions
Acknowledgments
Conflicts of Interest
References
- Forrest, R.D. Early history of wound treatment. J. R. Soc. Med. 1982, 75, 198. [Google Scholar] [PubMed]
- Clardy, J.; Fischbach, M.A.; Currie, C.R. The natural history of antibiotics. Curr. Biol. 2009, 19, R437–R441. [Google Scholar] [CrossRef] [PubMed]
- Gethin, G.T.; Cowman, S.; Conroy, R.M. The impact of Manuka honey dressings on the surface pH of chronic wounds. Int. Wound J. 2008, 5, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Ball, D.W. The chemical composition of honey. J. Chem. Educ. 2007, 84, 1643. [Google Scholar] [CrossRef]
- White, J.; Doner, L.W. Honey composition and properties. Beekeep. U. S. Agric. Handb. 1980, 335, 82–91. [Google Scholar]
- Kwakman, P.H.; Zaat, S.A. Antibacterial components of honey. IUBMB Life 2012, 64, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Molan, P.; Cooper, R.; Molan, P.; White, R. Why honey works. In Honey in Modern Wound Management; Wounds UK Ltd.: Aberdeen, UK, 2009; Volume 9, pp. 36–37. [Google Scholar]
- Sanz, M.; Gonzalez, M.; De Lorenzo, C.; Sanz, J.; Martınez-Castro, I. A contribution to the differentiation between nectar honey and honeydew honey. Food Chem. 2005, 91, 313–317. [Google Scholar] [CrossRef]
- Földházi, G. Analysis and quantitation of sugars in honey of different botanical origin using high performance liquid chromatography. Acta Aliment. 1994, 23, 299–311. [Google Scholar]
- Majtan, J.; Bohova, J.; Garcia-Villalba, R.; Tomas-Barberan, F.A.; Madakova, Z.; Majtan, T.; Majtan, V.; Klaudiny, J. Fir honeydew honey flavonoids inhibit TNF-α-induced MMP-9 expression in human keratinocytes: A new action of honey in wound healing. Arch. Dermatol. Res. 2013, 305, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the total phenolic, flavonoid and proline contents in Burkina Fasan honey, as well as their radical scavenging activity. Food Chem. 2005, 91, 571–577. [Google Scholar] [CrossRef]
- Jenkins, R.; Burton, N.; Cooper, R. Effect of manuka honey on the expression of universal stress protein A in meticillin-resistant Staphylococcus aureus. Int. J. Antimicrob. Agents 2011, 37, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Leong, A.G.; Herst, P.M.; Harper, J.L. Indigenous New Zealand honeys exhibit multiple anti-inflammatory activities. Innate Immun. 2012, 18, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.J.; Manley-Harris, M.; Molan, P.C. The origin of methylglyoxal in New Zealand manuka (Leptospermum scoparium) honey. Carbohydr. Res. 2009, 344, 1050–1053. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.; Molan, P.; Harding, K. The sensitivity to honey of Gram-positive cocci of clinical significance isolated from wounds. J. Appl. Microbiol. 2002, 93, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Molan, P.C. Re-introducing honey in the management of wounds and ulcers-theory and practice. Ostomy Wound Manag. 2002, 48, 28–40. [Google Scholar]
- Molan, P.C. Potential of honey in the treatment of wounds and burns. Am. J. Clin. Dermatol. 2001, 2, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Gleiter, R.; Horn, H.; Isengard, H.-D. Influence of type and state of crystallisation on the water activity of honey. Food Chem. 2006, 96, 441–445. [Google Scholar] [CrossRef]
- Sundoro, A.; Nadia, K.; Nur, A.; Sudjatmiko, G.; Tedjo, A. Comparison of physical–chemical characteristic and antibacterial effect between Manuka honey and local honey. J. Plast. Rekonstr. 2012, 1. [Google Scholar] [CrossRef]
- Rockland, L.B.; Beuchat, L.R. Water Activity: Theory and Applications to Food; M. Dekker: New York, NY, USA, 1987. [Google Scholar]
- Molan, P.C. The evidence supporting the use of honey as a wound dressing. Int. J. Lower Extrem. Wounds 2006, 5, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Kwakman, P.H.; Te Velde, A.A.; De Boer, L.; Vandenbroucke-Grauls, C.M.; Zaat, S.A. Two major medicinal honeys have different mechanisms of bactericidal activity. PLoS ONE 2011, 6, e17709. [Google Scholar] [CrossRef] [PubMed]
- Brudzynski, K.; Abubaker, K.; Castle, A. Re-examining the role of hydrogen peroxide in bacteriostatic and bactericidal activities of honey. Front. Microbiol. 2011, 2, 213. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.E.; Maddocks, S.E.; Cooper, R.A. Manuka honey is bactericidal against Pseudomonas aeruginosa and results in differential expression of oprF and algD. Microbiology 2012, 158, 3005–3013. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Starkey, M.; Hazan, R.; Rahme, L. Honey’s ability to counter bacterial infections arises from both bactericidal compounds and QS inhibition. Front. Microbiol. 2012, 3, 144. [Google Scholar] [CrossRef] [PubMed]
- Molan, P.C. The antibacterial activity of honey: 2. Variation in the potency of the antibacterial activity. Bee World 1992, 73, 59–76. [Google Scholar] [CrossRef]
- Efem, S. Clinical observations on the wound healing properties of honey. Br. J. Surg. 1988, 75, 679–681. [Google Scholar] [CrossRef] [PubMed]
- Armon, P. The use of honey in the treatment of infected wounds. Trop. Dr. 1980, 10, 91. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.; Molan, P.; Harding, K. Antibacterial activity of honey against strains of Staphylococcus aureus from infected wounds. J. R. Soc. Med. 1999, 92, 283–285. [Google Scholar] [CrossRef] [PubMed]
- Kwakman, P.H.; te Velde, A.A.; de Boer, L.; Speijer, D.; Vandenbroucke-Grauls, C.M.; Zaat, S.A. How honey kills bacteria. FASEB J. 2010, 24, 2576–2582. [Google Scholar] [CrossRef] [PubMed]
- Bachanová, K.; Klaudiny, J.; Kopernický, J.; Šimúth, J. Identification of honeybee peptide active against Paenibacillus larvae larvae through bacterial growth-inhibition assay on polyacrylamide gel. Apidologie 2002, 33, 259–269. [Google Scholar] [CrossRef]
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710. [Google Scholar] [CrossRef] [PubMed]
- Rabie, E.; Serem, J.C.; Oberholzer, H.M.; Gaspar, A.R.M.; Bester, M.J. How methylglyoxal kills bacteria: An ultrastructural study. Ultrastruct. Pathol. 2016, 40, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Majtan, J.; Klaudiny, J.; Bohova, J.; Kohutova, L.; Dzurova, M.; Sediva, M.; Bartosova, M.; Majtan, V. Methylglyoxal-induced modifications of significant honeybee proteinous components in manuka honey: Possible therapeutic implications. Fitoterapia 2012, 83, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Majtan, J.; Bohova, J.; Prochazka, E.; Klaudiny, J. Methylglyoxal may affect hydrogen peroxide accumulation in manuka honey through the inhibition of glucose oxidase. J. Med. Food 2014, 17, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Blaser, G.; Santos, K.; Bode, U.; Vetter, H.; Simon, A. Effect of medical honey on wounds colonised or infected with MRSA. J. Wound Care 2007, 16, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Thawley, A. The components of honey and their effects on its properties: A review. Bee World 1969, 50, 51–60. [Google Scholar] [CrossRef]
- Garcia-Alvarez, M.; Huidobro, J.; Hermida, M.; Rodriguez-Otero, J. Major components of honey analysis by near-infrared transflectance spectroscopy. J. Agric. Food Chem. 2000, 48, 5154–5158. [Google Scholar] [CrossRef] [PubMed]
- Molan, P.C. The evidence and the rationale for the use of honey as wound dressing. Wound Pract. Res. 2011, 19, 204–220. [Google Scholar]
- Lusby, P.; Coombes, A.; Wilkinson, J. Honey: A potent agent for wound healing? J. Wound Ostomy Cont. Nurs. 2002, 29, 295–300. [Google Scholar] [CrossRef]
- Sherlock, O.; Dolan, A.; Athman, R.; Power, A.; Gethin, G.; Cowman, S.; Humphreys, H. Comparison of the antimicrobial activity of Ulmo honey from Chile and Manuka honey against methicillin-resistant Staphylococcus aureus, Escherichia coli and Pseudomonas aeruginosa. BMC Complement. Altern. Med. 2010, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Al Somal, N.; Coley, K.; Molan, P.; Hancock, B. Susceptibility of Helicobacter pylori to the antibacterial activity of manuka honey. J. R. Soc. Med. 1994, 87, 9–12. [Google Scholar] [PubMed]
- Watanabe, K.; Rahmasari, R.; Matsunaga, A.; Haruyama, T.; Kobayashi, N. Anti-influenza viral effects of honey in vitro: Potent high activity of manuka honey. Arch. Med. Res. 2014, 45, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Zeina, B.; Othman, O.; Al-Assad, S. Effect of honey versus thyme on Rubella virus survival in vitro. J. Altern. Complement. Med. 1996, 2, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, A.; Cohrs, R.J. In vitro antiviral activity of honey against varicella zoster virus (VZV): A translational medicine study for potential remedy for shingles. Transl. Biomed. 2012, 3. [Google Scholar] [CrossRef]
- Merckoll, P.; Jonassen, T.Ø.; Vad, M.E.; Jeansson, S.L.; Melby, K.K. Bacteria, biofilm and honey: A study of the effects of honey on ‘planktonic’and biofilm-embedded chronic wound bacteria. Scand. J. Infect. Dis. 2009, 41, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Jervis-Bardy, J.; Foreman, A.; Bray, S.; Tan, L.; Wormald, P.J. Methylglyoxal-infused honey mimics the anti-Staphylococcus aureus biofilm activity of manuka honey: Potential Implication in Chronic Rhinosinusitis. Laryngoscope 2011, 121, 1104–1107. [Google Scholar] [CrossRef] [PubMed]
- Alandejani, T.; Marsan, J.; Ferris, W.; Slinger, R.; Chan, F. Effectiveness of honey on Staphylococcus aureus and Pseudomonas aeruginosa biofilms. Otolaryngol. Head Neck Surg. 2009, 141, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Okhiria, O.; Henriques, A.; Burton, N.; Peters, A.; Cooper, R. Honey modulates biofilms of Pseudomonas aeruginosa in a time and dose dependent manner. J. ApiProd. ApiMed. Sci. 2009, 1, 6–10. [Google Scholar]
- Sojka, M.; Valachova, I.; Bucekova, M.; Majtan, J. Antibiofilm efficacy of honey and bee-derived defensin-1 on multispecies wound biofilm. J. Med. Microbiol. 2016, 65, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Moghazy, A.; Shams, M.; Adly, O.; Abbas, A.; El-Badawy, M.; Elsakka, D.; Hassan, S.; Abdelmohsen, W.; Ali, O.; Mohamed, B. The clinical and cost effectiveness of bee honey dressing in the treatment of diabetic foot ulcers. Diabetes Res. Clin. Pract. 2010, 89, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcus epidermidis—The ‘accidental’ pathogen. Nat. Rev. Microbiol. 2009, 7, 555. [Google Scholar] [CrossRef] [PubMed]
- Efem, S.E. Recent advances in the management of Fournier’s gangrene: Preliminary observations. Surgery 1993, 113, 200–204. [Google Scholar] [PubMed]
- Gethin, G.; Cowman, S. Bacteriological changes in sloughy venous leg ulcers treated with manuka honey or hydrogel: An RCT. J. Wound Care 2008, 17, 241–247. [Google Scholar] [CrossRef] [PubMed]
- The Science Behind Honey’s Eternal Shelf Life. Available online: https://www.smithsonianmag.com/science-nature/the-science-behind-honeys-eternal-shelf-life-1218690/ (accessed on 21 February 2018).
- Frequently Asked Questions. Available online: https://www.honey.com/faq (accessed on 21 February 2018).
- Antony, S.; Rieck, J.; Acton, J.; Han, I.; Halpin, E.; Dawson, P. Effect of dry honey on the shelf life of packaged turkey slices. Poult. Sci. 2006, 85, 1811–1820. [Google Scholar] [PubMed]
- Ergun, M.; Ergun, N. Extending shelf life of fresh-cut persimmon by honey solution dips. J. Food Process. Preserv. 2010, 34, 2–14. [Google Scholar] [CrossRef]
- Viuda-Martos, M.; Ruiz-Navajas, Y.; Fernández-López, J.; Pérez-Álvarez, J. Functional properties of honey, propolis, and royal jelly. J. Food Sci. 2008, 73, R117–R124. [Google Scholar] [CrossRef] [PubMed]
- Randall, W.A.; Welcb, H.; Hunter, A.C. The stability of penicillin sodium held at various temperatures. J. Pharm. Sci. 1945, 34, 110–113. [Google Scholar] [CrossRef]
- Dimins, F.; Kuka, P.; Kuka, M.; Cakste, I. The criteria of honey quality and its changes during storage and thermal treatment. Proc. Latvia Univ. Agric. 2006, 16, 73–78. [Google Scholar]
- Bloom, B.R. Vaccines for the third world. Nature 1989, 342, 115. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, J.; Chen, H.; Li, F.; Armstrong, G.L.; Nelson, C.; Ze, W.; Shapiro, C.N. Hepatitis B vaccination of newborn infants in rural China: Evaluation of a village-based, out-of-cold-chain delivery strategy. Bull. World Health Organ. 2007, 85, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Tonks, A.J.; Cooper, R.; Jones, K.; Blair, S.; Parton, J.; Tonks, A. Honey stimulates inflammatory cytokine production from monocytes. Cytokine 2003, 21, 242–247. [Google Scholar] [CrossRef]
- Sell, S.A.; Wolfe, P.S.; Spence, A.J.; Rodriguez, I.A.; McCool, J.M.; Petrella, R.L.; Garg, K.; Ericksen, J.J.; Bowlin, G.L. A preliminary study on the potential of manuka honey and platelet-rich plasma in wound healing. Int. J. Biomater. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Oryan, A.; Zaker, S. Effects of topical application of honey on cutaneous wound healing in rabbits. Transbound. Emerg. Dis. 1998, 45, 181–188. [Google Scholar] [CrossRef]
- Prakash, A.; Medhi, B.; Avti, P.; Saikia, U.; Pandhi, P.; Khanduja, K. Effect of different doses of Manuka honey in experimentally induced inflammatory bowel disease in rats. Phytother. Res. 2008, 22, 1511–1519. [Google Scholar] [CrossRef] [PubMed]
- Ranzato, E.; Martinotti, S.; Burlando, B. Epithelial mesenchymal transition traits in honey-driven keratinocyte wound healing: Comparison among different honeys. Wound Repair Regener. 2012, 20, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Suguna, L.; Chandrakasan, G.; Joseph, K.T. Influence of honey on collagen metabolism during wound healing in rats. J. Clin. Biochem. Nutr. 1992, 13, 7–12. [Google Scholar] [CrossRef]
- Gethin, G.; Cowman, S. Case series of use of Manuka honey in leg ulceration. Int. Wound J. 2005, 2, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Visavadia, B.G.; Honeysett, J.; Danford, M.H. Manuka honey dressing: An effective treatment for chronic wound infections. Br. J. Oral Maxillofac. Surg. 2008, 46, 55–56. [Google Scholar] [CrossRef] [PubMed]
- Al-Waili, N.; Salom, K.; Al-Ghamdi, A.A. Honey for wound healing, ulcers, and burns; data supporting its use in clinical practice. Sci. World J. 2011, 11, 766–787. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.; Sofka, K.; Wiszniewsky, G.; Blaser, G.; Bode, U.; Fleischhack, G. Wound care with antibacterial honey (Medihoney) in pediatric hematology–oncology. Support. Care Cancer 2006, 14, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Okeniyi, J.A.; Olubanjo, O.O.; Ogunlesi, T.A.; Oyelami, O.A. Comparison of healing of incised abscess wounds with honey and EUSOL dressing. J. Altern. Complement. Med. 2005, 11, 511–513. [Google Scholar] [CrossRef] [PubMed]
- Jull, A.B.; Cullum, N.; Dumville, J.C.; Westby, M.J.; Deshpande, S.; Walker, N. Honey as a topical treatment for wounds. Cochrane Database Syst. Rev. 2015, 6, CD005083. [Google Scholar] [CrossRef] [PubMed]
- Aron, M.; Akinpelu, O.V.; Dorion, D.; Daniel, S. Otologic safety of manuka honey. J. Otolaryngol.—Head Neck Surg. 2012, 41, S21–S30. [Google Scholar] [PubMed]
- Paramasivan, S.; Drilling, A.J.; Jardeleza, C.; Jervis-Bardy, J.; Vreugde, S.; Wormald, P.J. Methylglyoxal-augmented manuka honey as a topical anti–Staphylococcus aureus biofilm agent: Safety and efficacy in an in vivo model. In International Forum of Allergy & Rhinology; Wiley Online Library: Hoboken, NJ, USA, 2014; pp. 187–195. [Google Scholar]
- Vadodaria, K.; Stylios, G.K. Ultrafine Web Formation from Bee’s Sweet Treasure. In Advanced Materials Research; Trans Tech Publication: Stafa-Zurich, Switzerland, 2013; pp. 1784–1788. [Google Scholar]
- Maleki, H.; Gharehaghaji, A.; Dijkstra, P. A novel honey-based nanofibrous scaffold for wound dressing application. J. Appl. Polym. Sci. 2013, 127, 4086–4092. [Google Scholar] [CrossRef]
- Minden-Birkenmaier, B.A.; Neuhalfen, R.M.; Janowiak, B.E.; Sell, S.A. Preliminary Investigation and Characterization of Electrospun Polycaprolactone and Manuka Honey Scaffolds for Dermal Repair. J. Eng. Fabr. Fibers (JEFF) 2015, 10, 126–138. [Google Scholar]
- Murthy, M.B.; Murthy, B.K.; Bhave, S. Comparison of safety and efficacy of papaya dressing with hydrogen peroxide solution on wound bed preparation in patients with wound gape. Indian J. Pharmacol. 2012, 44, 784. [Google Scholar] [CrossRef] [PubMed]
- Sadek, K.M. Antioxidant and immunostimulant effect of Carica papaya Linn. aqueous extract in acrylamide intoxicated rats. Acta Inform. Med. 2012, 20, 180. [Google Scholar] [CrossRef] [PubMed]
- Balaji, A.; Jaganathan, S.K.; Ismail, A.F.; Rajasekar, R. Fabrication and hemocompatibility assessment of novel polyurethane-based bio-nanofibrous dressing loaded with honey and Carica papaya extract for the management of burn injuries. Int. J. Nanomed. 2016, 11, 4339. [Google Scholar]
- Kadakia, P.U.; Growney Kalaf, E.A.; Dunn, A.J.; Shornick, L.P.; Sell, S.A. Comparison of silk fibroin electrospun scaffolds with poloxamer and honey additives for burn wound applications. J. Bioact. Compat. Polym. 2016, 33. [Google Scholar] [CrossRef]
- Yang, X.; Fan, L.; Ma, L.; Wang, Y.; Lin, S.; Yu, F.; Pan, X.; Luo, G.; Zhang, D.; Wang, H. Green electrospun Manuka honey/silk fibroin fibrous matrices as potential wound dressing. Mater. Des. 2017, 119, 76–84. [Google Scholar] [CrossRef]
- Hixon, K.R.; Lu, T.; Carletta, M.N.; McBride-Gagyi, S.H.; Janowiak, B.E.; Sell, S.A. A preliminary in vitro evaluation of the bioactive potential of cryogel scaffolds incorporated with Manuka honey for the treatment of chronic bone infections. J. Biomed. Mater. Res. Part B Appl. Biomater. 2017. [Google Scholar] [CrossRef] [PubMed]
- Arnett, T.R. Extracellular pH regulates bone cell function. J. Nutr. 2008, 138, 415S–418S. [Google Scholar] [CrossRef] [PubMed]
- Arnett, T.R. Acidosis, hypoxia and bone. Arch. Biochem. Biophys. 2010, 503, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Hixon, K.R.; Lu, T.; McBride-Gagyi, S.H.; Janowiak, B.E.; Sell, S.A. A Comparison of Tissue Engineering Scaffolds Incorporated with Manuka Honey of Varying UMF. BioMed Res. Int. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhu, X.-K.; Xue, X.-T.; Wu, D.-Y. Hydrogel sheets of chitosan, honey and gelatin as burn wound dressings. Carbohydr. Polym. 2012, 88, 75–83. [Google Scholar] [CrossRef]
- Sasikala, L.; Durai, B. Development and evaluation of chitosan honey hydrogel sheets as wound dressing. Int. J. Pharm. Biol. Sci. 2015, 6, 26–37. [Google Scholar]
- Leveen, H.H.; Falk, G.; Borek, B.; Diaz, C.; Lynfield, Y.; Wynkoop, B.J.; Mabunda, G.A.; Rubricius, J.L.; Christoudias, G.C. Chemical acidification of wounds. An adjuvant to healing and the unfavorable action of alkalinity and ammonia. Ann. Surg. 1973, 178, 745. [Google Scholar] [CrossRef] [PubMed]
- Greener, B.; Hughes, A.; Bannister, N.; Douglass, J. Proteases and pH in chronic wounds. J. Wound Care 2005, 14, 59–61. [Google Scholar] [CrossRef] [PubMed]
- Phillips, I.; Lobo, A.; Fernandes, R.; Gundara, N. Acetic acid in the treatment of superficial wounds infected by Pseudomonas aeruginosa. Lancet 1968, 291, 11–13. [Google Scholar] [CrossRef]
- Giusto, G.; Beretta, G.; Vercelli, C.; Valle, E.; Iussich, S.; Borghi, R.; Odetti, P.; Monacelli, F.; Tramuta, C.; Grego, E. A simple method to produce pectin-honey hydrogels and its characterization as new biomaterial for surgical use. J. Biomed. Mater. Res. Part B 2018. under review. [Google Scholar]
- Giusto, G.; Beretta, G.; Vercelli, C.; Valle, E.; Iussich, S.; Borghi, R.; Odetti, P.; Monacelli, F.; Tramuta, C.; Grego, E. Pectin-honey hydrogel: Characterization, antimicrobial activity and biocompatibility. Biomed. Mater. Eng. 2018, 29, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Mohd Zohdi, R.; Abu Bakar Zakaria, Z.; Yusof, N.; Mohamed Mustapha, N.; Abdullah, M.N.H. Gelam (Melaleuca spp.) honey-based hydrogel as burn wound dressing. Evid.-Based Complement. Altern. Med. 2011, 2012. [Google Scholar] [CrossRef]
- Medihoney. Available online: http://www.dermasciences.com/medihoney (accessed on 4 December 2017).
- Müller, P.; Alber, D.G.; Turnbull, L.; Schlothauer, R.C.; Carter, D.A.; Whitchurch, C.B.; Harry, E.J. Synergism between Medihoney and rifampicin against methicillin-resistant Staphylococcus aureus (MRSA). PLoS ONE 2013, 8, e57679. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.; Jenkins, L.; Hooper, S. Inhibition of biofilms of Pseudomonas aeruginosa by Medihoney in vitro. J. Wound Care 2014, 23, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.W.; van Eps, C.; Mudge, D.W.; Wiggins, K.J.; Armstrong, K.; Hawley, C.M.; Campbell, S.B.; Isbel, N.M.; Nimmo, G.R.; Gibbs, H. Randomized, controlled trial of topical exit-site application of honey (Medihoney) versus mupirocin for the prevention of catheter-associated infections in hemodialysis patients. J. Am. Soc. Nephrol. 2005, 16, 1456–1462. [Google Scholar] [CrossRef] [PubMed]
- Robson, V.; Dodd, S.; Thomas, S. Standardized antibacterial honey (Medihoney™) with standard therapy in wound care: Randomized clinical trial. J. Adv. Nurs. 2009, 65, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.W.; Clark, C.; Isbel, N.M.; Hawley, C.M.; Beller, E.; Cass, A.; De Zoysa, J.; McTaggart, S.; Playford, G.; Rosser, B. The honeypot study protocol: A randomized controlled trial of exit-site application of medihoney antibacterial wound gel for the prevention of catheter-associated infections in peritoneal dialysis patients. Perit. Dial. Int. 2009, 29, 303–309. [Google Scholar] [PubMed]
- Biglari, B.; Vd Linden, P.; Simon, A.; Aytac, S.; Gerner, H.; Moghaddam, A. Use of Medihoney as a non-surgical therapy for chronic pressure ulcers in patients with spinal cord injury. Spinal Cord 2012, 50, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.; Legel, K.; Hanft, J.R. Topical Leptospermum honey (Medihoney) in recalcitrant venous leg wounds: A preliminary case series. Adv. Skin Wound Care 2009, 22, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Wound Dressing with Manuka Honey. Available online: http://honeywoundcare.com/product-range/product-61/Wound-Dressing-with-Manuka-Honey (accessed on 4 December 2017).
- ManukaMed. Available online: https://shop.manukamed.com/products/manukahd (accessed on 4 December 2017).
- Activon—Manuka Honey Dressings. Available online: http://www.advancis.co.uk/products/activon-manuka-honey (accessed on 4 December 2017).
- What We Do. Available online: http://sweetbio.com/what-we-do/ (accessed on 4 December 2017).
- Sun, Z.; Zussman, E.; Yarin, A.L.; Wendorff, J.H.; Greiner, A. Compound core–shell polymer nanofibers by co-electrospinning. Adv. Mater. 2003, 15, 1929–1932. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, Z.-M.; Xu, X.; Lim, C.T.; Ramakrishna, S. Preparation of core—Shell structured PCL-r-gelatin bi-component nanofibers by coaxial electrospinning. Chem. Mater. 2004, 16, 3406–3409. [Google Scholar] [CrossRef]
- Wang, P.; He, J.-H. Electrospun polyvinyl alcohol-honey nanofibers. Therm. Sci. 2013, 17, 1549–1550. [Google Scholar] [CrossRef]
- Tamura, D.Y.; Moore, E.E.; Partrick, D.A.; Johnson, J.L.; Offner, P.J.; Silliman, C.C. Acute hypoxemia in humans enhances the neutrophil inflammatory response. Shock 2002, 17, 269–273. [Google Scholar] [CrossRef] [PubMed]
- McCourt, M.; Wang, J.H.; Sookhai, S.; Redmond, H.P. Proinflammatory mediators stimulate neutrophil-directed angiogenesis. Arch. Surg. 1999, 134, 1325–1331. [Google Scholar] [CrossRef] [PubMed]
- Chavakis, T.; Cines, D.B.; Rhee, J.-S.; Liang, O.D.; Schubert, U.; Hammes, H.-P.; Higazi, A.A.-R.; Nawroth, P.P.; Preissner, K.T.; Bdeir, K. Regulation of neovascularization by human neutrophil peptides (α-defensins): A link between inflammation and angiogenesis. FASEB J. 2004, 18, 1306–1308. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration | 50% v/v | 25% v/v | 12.5% v/v | 6.3% v/v | ||||
|---|---|---|---|---|---|---|---|---|
| Isolates | Ulmo | Manuka | Ulmo | Manuka | Ulmo | Manuka | Ulmo | Manuka |
| MRSA ATCC 43300 | 30 (1.7) | 24 (1.5) | 26 (0.6) | 19 (2.1) | 18 (0.6) | 13 (1.0) | 10 (0.6) | - |
| MRSA 0791 * | 34 (1.5) | 23 (1.2) | 29 (1.7) | 17 (1.7) | 22 (2.1) | - | 14 (2.5) | - |
| MRSA 28965 * | 24 (1.0) | 17 (1.7) | 19 (1.5) | 15 (2.0) | - | - | - | - |
| MRSA 01322 * | 28 (5.8) | 22 (1.0) | 23 (4.2) | 18 (0.6) | 17 (2.9) | - | 11 (2.0) | - |
| MRSA 0745 * | 23 (2.7) | 20 (1.7) | 19 (2.1) | 13 (1.7) | 11 (2.7) | - | - | - |
| P. aeruginosa ATCC 27853 | 14 (2.3) | 16 (7.8) | 11 (1.0) | 14 (6.9) | - | - | - | - |
| E. coli ATCC 35218 | 14 (1.5) | 15 (2.5) | 11 (1.7) | 12 (2.9) | - | - | - | - |
| Product Name | Company | Product Category |
|---|---|---|
| Medihoney® Paste | Derma Sciences | Paste, topical application |
| Manuka Honey (Paste) | Manuka Health | Paste, topical application |
| Activon Tube | Advancis Medical | Paste, topical application |
| Medihoney® Gel | Derma Sciences | Gel, topical application |
| ManukaApli | ManukaMed | Gel, topical application |
| Medihoney® Alginate Dressing | Derma Sciences | Composite Hydrogel |
| Manuka Health Wound Dressing | Manuka Health | Composite Hydrogel |
| Algivon, Algivon Plus | Advancis Medical | Composite Hydrogel |
| Medihoney® HCS Application | Derma Sciences | Honey-impregnated Dressing |
| Medihoney® Honeycolloid Application | Derma Sciences | Honey-impregnated Dressing |
| Manukahd | ManukaMed | Honey-impregnated Dressing |
| ManukaMed MedSaf | ManukaMed | Honey-impregnated Dressing |
| Manukahd Lite | ManukaMed | Honey-impregnated Dressing |
| Manukahd Lite rope | ManukaMed | Honey-impregnated Dressing |
| Actilite | Advancis Medical | Honey-impregnated Dressing |
| Algivon Tulle | Advancis Medical | Honey-impregnated Dressing |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minden-Birkenmaier, B.A.; Bowlin, G.L. Honey-Based Templates in Wound Healing and Tissue Engineering. Bioengineering 2018, 5, 46. https://doi.org/10.3390/bioengineering5020046
Minden-Birkenmaier BA, Bowlin GL. Honey-Based Templates in Wound Healing and Tissue Engineering. Bioengineering. 2018; 5(2):46. https://doi.org/10.3390/bioengineering5020046
Chicago/Turabian StyleMinden-Birkenmaier, Benjamin A., and Gary L. Bowlin. 2018. "Honey-Based Templates in Wound Healing and Tissue Engineering" Bioengineering 5, no. 2: 46. https://doi.org/10.3390/bioengineering5020046
APA StyleMinden-Birkenmaier, B. A., & Bowlin, G. L. (2018). Honey-Based Templates in Wound Healing and Tissue Engineering. Bioengineering, 5(2), 46. https://doi.org/10.3390/bioengineering5020046