
Prospecting for Marine Bacteria for Polyhydroxyalkanoate Production on Low-Cost Substrates

Abstract
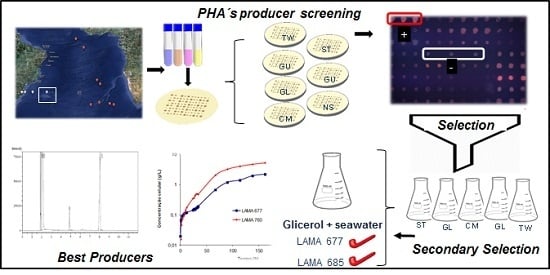
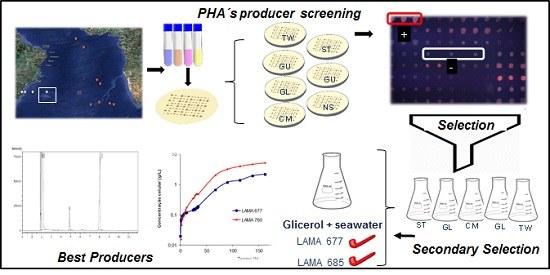
:
1. Introduction
2. Materials and Methods
2.1. Marine Bacteria
2.2. Qualitative Screening of PHA Producers
2.3. Production of PHA in Semi-Solid and Liquid Mediums
2.4. Quantification of PHA by Gas Chromatography
3. Results and Discussion
3.1. Screening for PHA-Producing Marine Bacteria
3.2. PHA Production in Different Substrates
3.3. Growth Kinetics and P(3HB) Production
3.4. Production of P(3HB) in Seawater and Residual Biodiesel Glycerol
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rehm, B.H.A.; Steinbüche, A. Biochemical and genetic analysis of PHA synthases and other proteins required for PHA synthesis. Int. J. Biol. Macromol. 1999, 25, 3–19. [Google Scholar] [CrossRef]
- Sudesh, K.; Abe, H.; Doi, Y. Synthesis, structure and properties of polyhydroxyalkanoates: Biological polyesters. Prog. Polym. Sci. 2000, 25, 1503–1555. [Google Scholar] [CrossRef]
- Sheu, D.S.; Chen, W.M.; Yang, J.Y.; Chang, R.C. Thermophilic bacterium Caldimonas taiwanensis produces poly(3-hydroxybutyrate-co-3-hydroxyvalerate) from starch and valerate as carbon sources. Enzym. Microb. Technol. 2009, 44, 289–294. [Google Scholar] [CrossRef]
- Tay, B.Y.; Lokesh, B.E.; Lee, C.Y.; Sudesh, K. Polyhydroxyalkanoate (PHA) accumulating bacteria from the gut of higher termite Macrotermes carbonarius (Blattodea: Termitidae). World J. Microbiol. Biotechnol. 2010, 26, 1015–1024. [Google Scholar] [CrossRef]
- Valentin, H.E.; Broyles, D.L.; Casagrande, L.A.; Colburn, S.M.; Creely, W.L.; Delaquil, P.A.; Felton, H.M.; Gon-zalez, K.A.; Houmiel, K.L.; Lutke, K.; et al. PHA production, from bacteria to plants. Int. J. Biol. Macromol. 1999, 25, 303–306. [Google Scholar] [CrossRef]
- Hermann-Krauss, C.; Koller, M.; Muhr, A.; Fasl, H.; Stelzer, F.; Braunergg, G. Archaeal production of polyhydroxyalkanoate (PHA) co-and terpolyesters from biodiesel industry-derived by-products. Archaea 2013. [Google Scholar] [CrossRef] [PubMed]
- Koller, M.; Maršálek, L.; Dias, M.M.S.; Braunegg, G. Producing microbial polyhydroxyalkanoate (PHA) biopolyesters in a sustainable manner. New Biotechnol. 2017, 37, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Quillaguamán, J.; Guzmán, H.; Van-Thuoc, D.; Hatti-Kaul, R. Synthesis and production of polyhydroxyalkanoates by halophiles: Current potential and future prospects. Appl. Microbiol. Biotechnol. 2010, 85, 1687–1696. [Google Scholar] [CrossRef] [PubMed]
- Shrivastav, A.; Mishra, S.K.; Shethia, B.; Pancha, I.; Jain, D.; Mishra, S. Isolation of promising bacterial strains from soil and marine environment for polyhydroxyalkanoates (PHAs) production utilizing Jatropha biodiesel byproduct. Int. J. Biol. Macromol. 2010, 47, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.; Xue, Y.S.; Aibaidula, G.; Chen, G.Q. Unsterile and continuous production of polyhydroxybutyrate by Halomonas TD01. Bioresour. Technol. 2011, 102, 8130–8136. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Ling, C.; Yang, T.; Chen, X.; Chen, Y.; Deng, H.; Wu, Q.; Chen, J.; Chen, G.-Q. A seawater-based open and continuous process for polyhydroxyalkanoates production by recombinant Halomonas campaniensis LS21 grown in mixed substrates. Biotechnol. Biofuels 2014, 7, 108. [Google Scholar] [CrossRef]
- Margesin, R.; Schinner, F. Potential of halotolerant and halophilic microorganisms for biotechnology. Extremophiles 2001, 5, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Kawata, Y.; Aiba, S. Poly(3-hydroxybutyrate) production by isolated Halomonas sp. KM-1 using waste glycerol. Biosci. Biotechnol. Biochem. 2010, 74, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Pandian, S.R.; Venkatraman, D.; Kalishwaralal, K.; Rameshkumar, N.; Jeraraj, M.; Gurunathan, S. Optimization and fed-batch production of PHB utilizing dairy waste and sea water as nutrient sources by Bacillus megaterium SRKP-3. Bioresour. Technol. 2010, 101, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Chen, J.C.; Wu, Q.; Chen, G.Q. Halophiles, coming stars for industrial biotechnology. Biotechnol. Adv. 2015, 33, 1433–1442. [Google Scholar] [CrossRef] [PubMed]
- Spiekermann, P.; Rehm, B.H.A.; Kalscheuer, R.; Baumeister, D.; Steinbüchel, A. A sensitive, viable-colony staining method using Nile red for direct screening of bacteria that accumulate polyhydroxyalkanoic acids and other lipid storage compounds. Arch. Microbiol. 1999, 171, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Baumann, P.; Baumann, L.; Mandel, M. Taxonomy of Marine Bacteria: The Genus Beneckea. J. Bacteriol. 1971, 107, 268–294. [Google Scholar] [PubMed]
- Alves, L.P.; Almeida, A.T.; Cruz, L.M.; Pedrosa, F.O.; De Souza, E.M.; Chubatsu, L.S.; Müller-Santos, M.; Valdameri, G. A simple and efficient method for poly-3-hydroxybutyrate quantification in diazotrophic bacteria within 5 minutes using flow cytometry. Braz. J. Med. Biol. Res. 2017, 50, e5492. [Google Scholar] [CrossRef] [PubMed]
- Zuriani, R.; Vigneswari, S.; Azizan, M.N.M.; Majid, M.I.A.; Amirul, A.A.A. High throughput Nile red fluorescence method for rapid quantification of intracellular bacterial polyhydroxyalkanoates. Biotechnol. Bioprocess Eng. 2013, 18, 472–478. [Google Scholar] [CrossRef]
- Arikawa, H.; Sato, S.; Fujiki, T.; Matsumoto, K. Simple and rapid method for isolation and quantitation of polyhydroxyalkanoate by SDS-sonication treatment. J. Biosci. Bioeng. 2017, S1389–S1723, 30664–30668. [Google Scholar] [CrossRef] [PubMed]
- Degelau, A.; Scheper, T.; Bailey, J.E.; Guske, C. Fluorometric measurement of poly-β hydroxybutyrate in Alcaligeneseutrophus by flow cytometry and spectrofluorometry. Appl. Microbiol. Biotechnol. 1995, 42, 653–657. [Google Scholar] [CrossRef]
- Reddy, C.S.K.; Ghai, R.; Rashmi; Kalia, V.C. Polyhydroxyalkanoates: An overview. Bioresour. Technol. 2003, 87, 137–146. [Google Scholar] [CrossRef]
- Hedgpeth, J. Classification of Marine Environments; Reseck, J., Jr., Ed.; Marine Biology: Englewood Cliffs, NJ, USA, 1957; pp. 18–27. [Google Scholar]
- Longhurst, R.A.; Harrison, G.W. Vertical nitrogen flux from the oceanic photic zone by diel migrant zooplankton and nekton. Deep Sea Res. Part A Oceanogr. Res. Pap. 1988, 35, 881–889. [Google Scholar] [CrossRef]
- Chien, C.C.; Chen, C.C.; Choi, M.H.; Kung, S.S.; Wei, Y.H. Production of poly-ß-hydroxybutyrate (PHB) by Vibrio spp. isolated from marine environment. J. Biotechnol. 2007, 132, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Mahishi, L.H.; Tripathi, G.; Rawal, S.K. Poly(3-hydroxybutyrate) (PHB) synthesis by recombinant Escherichia coli harbouring Streptomyces aureofaciens PHB biosynthesis genes: Effect of various carbon and nitrogen sources. Microbiol. Res. 2003, 158, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Mohandas, S.P.; Balan, L.; Lekshmi, N.; Cubelio, S.S.; Philip, R.; Sing, I.S.B. Production and characterization of Polyhydroxybutyrate from Vibrio Harveyi MCCB 284 utilizing glycerol as carbon source. J. Appl. Microbiol. 2016, 122, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Van-Thuoc, D.; Quillaguamán, J.; Mamo, G.; Matiason, B. Utilization of agricultural residues for poly(3-hydroxybutyrate) production by Halomonasboliviensis LC1. J. Appl. Microbiol. 2008, 104, 420–428. [Google Scholar] [PubMed]
- Bertrand, J.L.; Ramsay, B.A.; Ramsay, J.A.; Chavarie, C. Biosynthesis of poly-β-hydroxyalkanoates from pentoses by Pseudomonas pseudoflava. Appl. Environ. Microbiol. 1990, 56, 3133–3138. [Google Scholar] [PubMed]
- Silva, L.F.; Taciro, M.K.; Ramos, M.E.M.; Carter, J.M.; Pradella, J.G.C.; Gomez, J.G.C. Poly-3-hydroxybutyrate (P3HB) production by bacteria from xylose, glucose and sugarcane bagasse hydrolysate. J. Ind. Microbiol. Biotechnol. 2004, 31, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Fernández, D.; Rodríguez, E.; Bassas, M.; Viñas Solanas, A.M.; Liorens, J.; Marquéz, A.M.; Manresa, A. Agro-industrial oily wastes as substrates for PHA production by the new strain Pseudomonas aeruginosa NCIB 40045: Effect of culture conditions. Biochem. Eng. J. 2005, 26, 159–167. [Google Scholar] [CrossRef]
- He, W.; Tian, W.; Zhang, G.; Chen, G.-Q.; Zhang, Z. Production of novel polyhydroxyalkanoates by Pseudomonas stutzeri 1317 from glucose and soybean oil. FEMS Microbiol. Lett. 1998, 169, 45–49. [Google Scholar] [CrossRef]
- Cavalheiro, J.M.B.T.; Almeida, M.C.M.D.; Grandfils, C.; Fonseca, M.M.R. Poly(3-hydroxybutyrate) production by Cupriavidus necator using waste glycerol. Process Biochem. 2009, 44, 509–515. [Google Scholar] [CrossRef]
- Mothes, G.; Schnorpfeil, C.; Ackermann, J.U. Production of PHB from crude glycerol. Eng. Life Sci. 2007, 7, 475–479. [Google Scholar] [CrossRef]
- Zhu, C.; Nomura, C.T.; Perrotta, J.A.; Stipanovic, A.J.; Nakas, J.P. Production and characterization of poly-3-hydroxybutyrate from biodiesel-glycerol by Burkholderia cepacia ATCC 17759. Biotechnol. Prog. 2010, 26, 424–430. [Google Scholar] [PubMed]
- Kangsadan, T.; Swadchaipon, N.; Kongruang, S. Value-added utilization of crude glycerol from biodiesel production by microbial synthesis of polyhydroxybutyrate-valerate. Curr. Opin. Biotechnol. 2011, 22, S1–S35. [Google Scholar] [CrossRef]
- Piccoli, R.A.M.; Silva, E.S.; Taciro, M.K.; Maiorano, A.E.; Ribeiro, C.M.S.; Rodrigues, M.F.A. Produção de polihidroxibutirato a partir de glicerol resíduo da produção de biodiesel. In Simpósio Nacional de Bioprocessos; 17., 2011, Caxias do Sul. Anais...; Associação Brasileira de Engenharia Química: Caxias do Sul, Brazil, 2011; pp. 1–6. [Google Scholar]
- Rodrigues, R.C. Condições de Cultura Para a Produção de Poli(3-hidroxibutirato) por Ralstoniaeu tropha a partir de Resíduos de Indústrias de Alimento. Master’s Thesis, Universidade Federal de Santa Catarina, Trindade, Florianópolis, Brasil, 2005. [Google Scholar]
- Nascimento, V.M.; Silva, L.F.; Gomez, J.G.C.; Fonseca, G.G. Growth of Burkholderia sacchari LFM 101 cultivated in glucose, sucrose and glycerol at different temperatures. Sci. Agricola. 2016, 73, 429–433. [Google Scholar] [CrossRef]
- Yoneda, N.T. Área Temática: Plâncton, 1999. Centro de Estudos do Mar, Universidade Federal do Paraná. Available online: http://www.brasil-rounds.gov.br/round7/arquivos_r7/PERFURACAO_R7/refere/pl%E2ncton.pdf (accessed on 23 December 2011).
- Ray, S.; Prajapati, V.; Patel, K; Triedi, U. Optimization and characterization of PHA from isolate Pannonibacter phragmitetus ERC8 using glycerol waste. Int. J. Biol. Macromol. 2016, 86, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, J.M.; Posada, J.A.; Higuita, J.C.; Cardona, C.A. Valorization of glycerol through the production of biopolymers: The PHB case using Bacillus megaterium. Bioresour. Technol. 2013, 133, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Jincy, M.; Sindhu, R.; Pandey, A.; Binod, P. Bioprocess development for utilizing biodiesel industry generated crude glycerol for production of poly-3-hydroxybutyrate. J. Sci. Ind. Res. 2013, 72, 596–602. [Google Scholar]
- Rodríguez-Contreras, A.; Koller, M.; Dias, M.M.S.; Calaffel-Monfort, M.; Braunegg, G.; Marqués-Calvo, M.S. Influence of glycerol on poly(3-hydroxybutyrate) production by Cupriavidus necator and Burkholderia sacchari. Biochem. Eng. J. 2015, 94, 50–57. [Google Scholar] [CrossRef]
- Onwudili, J.A.; Williams, P.T. Hydrothermal reforming of bio-diesel plant waste: Products distribution and characterization. Fuel 2010, 89, 501–509. [Google Scholar] [CrossRef]
- Thompson, J.C.; He, B.B. Characterization of crude glycerol from biodiesel production from multiple feedstocks. Appl. Eng. Agric. 2006, 22, 261–265. [Google Scholar] [CrossRef]
- Posada, J.A.; Naranjo, J.M.; López, J.A.; Higuita, J.C.; Cardona, C.A. Design and analysis of poly-3-hydroxybutyrate production processes from crude glycerol. Process Biochem. 2011, 46, 310–317. [Google Scholar] [CrossRef]
- Andreeßen, B.; Lange, A.B.; Robenek, H.; Steinbüchel, A. Conversion of glycerol to poly(3-Hydroxypropionate) in recombinant Escherichia coli. Appl. Environ. Microbiol. 2010, 76, 622–626. [Google Scholar] [CrossRef] [PubMed]
- De Paula, F.C.; Kakazu, S.; de Paula, C.B.C.; Contiero, J. Polyhydroxyalkanoate production from crude glycerol by newly isolated Pandoraea sp. J. King Saud Univ. Sci. 2017, 29, 166–173. [Google Scholar] [CrossRef]
- Silva, A.M.X.P. Degradação de Efluentes Lácteo sem Reactores UASB Com Recirculação. Master’s Thesis, Universidade de Aveiro, Aveiro, Portugal, 2008. [Google Scholar]

{kind=link}
| Culture Media | Composition |
|---|---|
| 1 | 90% MM medium * + 5% (v/v) glycerol + 5% distilled water |
| 2 | 90% MM medium + 10% (v/v) glycerol |
| 3 | 90% seawater + 5% (v/v) glycerol + 5% distilled water |
| 4 | 90% seawater + 5% (v/v) residual glycerol + 5% distilled water |
| 5 | 90% seawater + 10% (v/v) residual glycerol |
| 6 | 90% MM medium + 5% (v/v) residual glycerol + 5% distilled water |
| Isolate LAMA 3 | MA 1 | MM 2 | Isolate LAMA | MA | MM | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NS | GU | ST | CM | GL | GU | TW | NS | GU | ST | CM | GL | GU | TW | ||
| 570 | − | − | wg | wg | wg | wg | wg | 671 | − | − | − | + | − | − | − |
| 571 | − | − | wg | wg | wg | − | wg | 672 | − | − | − | − | − | − | − |
| 572 | − | − | + | wg | − | − | − | 673 | − | − | + | + | + | + | − |
| 573 | − | − | wg | wg | wg | − | wg | 674 | + | − | + | + | + | + | + |
| 574 | − | − | wg | wg | wg | wg | − | 675 | − | − | wg | wg | wg | wg | − |
| 575 | − | − | wg | wg | wg | wg | wg | 677 | − | − | + | + | + | − | + |
| 576 | − | − | wg | wg | wg | wg | wg | 679 | + | + | + | + | + | + | + |
| 577 | − | − | wg | wg | − | − | wg | 680 | − | − | − | − | − | − | − |
| 580 | − | − | wg | − | − | − | wg | 681 | − | − | wg | − | − | − | wg |
| 582 | − | − | wg | wg | − | − | wg | 683 | − | − | wg | wg | wg | wg | + |
| 583 | − | − | − | wg | − | − | − | 684 | − | − | wg | wg | wg | wg | − |
| 584 | + | − | wg | wg | wg | wg | wg | 685 | − | − | − | + | + | + | − |
| 585 | − | − | wg | wg | − | − | wg | 687 | − | − | wg | wg | wg | wg | wg |
| 587 | − | − | − | − | − | − | − | 688 | − | − | wg | wg | wg | wg | wg |
| 592 | + | − | − | − | − | − | − | 689 | − | − | − | − | wg | − | − |
| 593 | − | − | wg | wg | wg | wg | wg | 690 | − | − | wg | − | wg | − | − |
| 594 | − | − | + | + | + | − | − | 691 | − | − | wg | wg | wg | wg | − |
| 595 | − | − | wg | − | wg | wg | wg | 692 | − | − | wg | wg | wg | wg | wg |
| 597 | − | − | wg | wg | − | wg | wg | 693 | − | − | − | − | − | − | − |
| 598 | − | − | wg | wg | wg | − | − | 694 | − | − | wg | wg | wg | wg | wg |
| 599 | − | − | + | + | − | − | − | 695 | − | − | − | − | − | − | − |
| 600 | − | − | − | − | − | − | − | 696 | − | − | wg | wg | wg | wg | wg |
| 601 | − | − | + | + | − | − | − | 697 | + | + | − | − | − | + | − |
| 604 | − | − | + | − | wg | − | − | 698 | − | − | wg | wg | wg | wg | wg |
| 606 | − | − | wg | wg | wg | wg | wg | 699 | − | − | wg | wg | wg | − | − |
| 607 | − | − | wg | wg | wg | − | − | 700 | − | − | − | wg | − | − | − |
| 608 | − | − | wg | wg | wg | wg | wg | 701 | − | − | wg | wg | − | wg | wg |
| 610 | − | − | wg | − | − | − | wg | 702 | − | − | + | + | + | + | − |
| 611 | − | − | wg | wg | wg | wg | − | 703 | − | − | + | − | − | − | − |
| 612 | − | − | + | + | + | + | − | 704 | − | − | + | + | − | − | − |
| 613 | − | − | wg | wg | wg | wg | wg | 705 | − | − | − | wg | − | − | − |
| 614 | − | − | wg | wg | + | − | wg | 706 | − | − | wg | wg | wg | wg | + |
| 615 | − | − | − | − | − | − | − | 707 | + | + | − | − | − | − | wg |
| 616 | − | − | wg | wg | wg | − | − | 708 | − | − | wg | − | − | − | wg |
| 617 | − | − | wg | wg | wg | wg | − | 709 | − | − | wg | wg | − | wg | wg |
| 618 | − | + | wg | wg | wg | wg | wg | 710 | − | − | + | wg | − | − | − |
| 619 | − | − | wg | wg | wg | − | − | 711 | + | + | + | + | + | + | − |
| 644 | − | + | − | − | − | − | − | 712 | − | − | wg | wg | wg | wg | − |
| 647 | − | − | − | − | − | − | − | 713 | − | − | wg | wg | wg | wg | wg |
| 650 | − | − | wg | − | − | + | − | 715 | − | − | wg | wg | wg | − | wg |
| 653 | + | − | − | − | − | − | + | 716 | − | − | wg | wg | wg | wg | wg |
| 659 | − | − | + | + | + | + | − | 717 | − | + | − | wg | wg | wg | wg |
| 667 | − | − | − | wg | − | − | − | 718 | − | − | wg | wg | − | − | wg |
| 669 | − | − | − | − | − | + | − | 719 | − | − | + | − | − | − | − |
| 720 | − | − | wg | wg | wg | wg | wg | 759 | − | − | wg | wg | wg | wg | wg |
| 722 | − | − | wg | + | − | + | − | 760 | − | − | + | + | + | − | − |
| 723 | − | − | − | − | − | − | − | 761 | + | + | − | + | + | + | + |
| 725 | − | − | − | wg | wg | − | wg | 762 | − | − | wg | wg | − | − | − |
| 726 | + | − | − | wg | − | + | + | 763 | + | + | wg | wg | wg | − | − |
| 727 | − | − | − | wg | wg | − | wg | 764 | − | − | wg | wg | wg | wg | wg |
| 728 | − | − | wg | wg | − | − | wg | 765 | − | − | + | + | + | + | − |
| 729 | − | + | + | + | + | + | − | 766 | − | − | + | wg | − | − | − |
| 730 | − | − | wg | wg | − | − | − | 767 | − | − | − | − | − | − | − |
| 731 | − | − | − | − | − | − | wg | 768 | − | − | wg | wg | wg | wg | wg |
| 732 | + | − | − | wg | wg | − | − | 769 | − | − | wg | wg | wg | wg | wg |
| 733 | − | − | − | − | − | − | − | 773 | − | − | + | + | + | + | − |
| 734 | − | + | − | wg | wg | − | − | 775 | − | − | wg | wg | − | − | wg |
| 735 | − | − | wg | wg | − | + | − | 778 | − | − | + | − | − | − | − |
| 736 | + | − | − | wg | + | + | − | 779 | − | − | − | − | wg | − | − |
| 737 | − | − | + | + | + | + | − | 781 | − | − | wg | − | − | − | − |
| 738 | − | − | wg | wg | − | − | − | 782 | − | − | − | wg | wg | wg | wg |
| 739 | − | − | wg | wg | wg | wg | wg | 786 | − | − | − | − | − | − | − |
| 741 | − | − | wg | wg | wg | − | − | 790 | − | − | + | − | − | − | − |
| 742 | − | − | wg | − | wg | − | − | 791 | − | − | + | − | − | − | − |
| 743 | − | − | wg | wg | − | − | wg | M 112 | − | − | wg | wg | wg | wg | wg |
| 744 | − | + | wg | wg | wg | − | − | M 135 | + | + | wg | wg | wg | wg | wg |
| 746 | − | − | wg | wg | − | − | − | M 151 | + | − | wg | wg | − | − | − |
| 747 | − | − | − | − | − | − | + | M 169 | − | − | wg | wg | wg | − | wg |
| 748 | − | − | + | + | + | − | − | M 171 | + | − | wg | wg | wg | − | − |
| 749 | − | − | wg | wg | wg | − | wg | M 173 | − | + | wg | wg | − | wg | wg |
| 750 | − | − | wg | − | + | + | − | M 180 | + | + | + | + | − | − | − |
| 751 | − | − | − | − | wg | − | wg | M 189 | − | − | − | − | − | − | − |
| 753 | − | − | wg | wg | wg | − | wg | M 198 | − | − | wg | wg | − | − | wg |
| 754 | − | − | + | + | + | − | − | M 199 | − | − | + | + | + | − | + |
| 755 | − | − | wg | wg | wg | wg | − | M 211 | − | − | − | + | − | − | − |
| 756 | − | − | wg | wg | wg | − | − | M 84 | + | − | wg | wg | wg | wg | wg |
| 757 | − | − | wg | − | − | − | wg | M 97 | − | − | wg | − | − | wg | wg |
| 758 | + | − | − | + | + | + | − | ||||||||
| Isolate | Carbon Source | Total Biomass (g·L−1) | P(3HB) Concentration (g·L−1) | P(3HB) Content in Total Biomass (%) | P(3HB) Productivity (g·L−1·h−1) |
|---|---|---|---|---|---|
| LAMA 748 | Starch | 16.80 ± 0.97 | 0 | 0 | 0 |
| LAMA 737 | Starch | 25.31 ± 1.09 | 0 | 0 | 0 |
| LAMA 748 | Carboxymethylcellulose | 1.19 ± 0.19 | 0 | 0 | 0 |
| LAMA 674 | Carboxymethylcellulose | 1.24 ± 0.20 | 0 | 0 | 0 |
| LAMA 677 | Glycerol | 1.41 ± 0.18 | 0.4 | 28.28 | 0.0058 |
| LAMA 685 | Glycerol | 2.03 ± 0.27 | 0.67 | 32.79 | 0.0098 |
| LAMA 685 | Glucose | 1.73 ± 0.22 | 0.17 | 9.62 | 0.0025 |
| LAMA 737 | Glucose | 0.68 ± 0.10 | 0.05 | 7.85 | 0.0008 |
| LAMA 726 | Tween 80 | 0.91 ± 0.08 | 0 | 0 | 0 |
| M 199 A | Tween 80 | 0.56 ± 0.05 | 0 | 0 | 0 |
| Isolate | Culture Medium Composition | Total Biomass (g·L−1) | P(3HB) Concentration (g·L−1) | P(3HB) Content in Total Biomass (%) | P(3HB) Productivity (g·L−1·h−1) |
|---|---|---|---|---|---|
| LAMA 677 | 90% mineral medium + 5% glycerol + 5% distilled water | 1.11 ± 0.04 | 0.71 | 64.28 | 0.0103 |
| 90% mineral medium + 10% glycerol | 0.85 ± 0.05 | 0.55 | 64.04 | 0.0079 | |
| 90% seawater + 5% glycerol + 5% distilled water | 0.10 ± 0.03 | 0.03 | 35.04 | 0.0005 | |
| 90% seawater + 5% residual glycerol + 5% distilled water | 0.06 ± 0.01 | 0.02 | 31.70 | 0.0003 | |
| 90% seawater + 10% residual glycerol | * | * | * | * | |
| 90% mineral medium + 5% residual glycerol + 5% distilled water | 1.39 ± 0.05 | 0.74 | 52.94 | 0.0107 | |
| LAMA 685 | 90% mineral medium + 5% glycerol + 5% distilled water | 0.95 ± 0.06 | 0.42 | 43.64 | 0.0060 |
| 90% mineral medium + 10% glycerol | 0.36 ± 0.07 | 0.10 | 28.08 | 0.0015 | |
| 90% seawater + 5% glycerol + 5% distilled water | 0.27 ± 0.03 | 0.13 | 48.26 | 0.0019 | |
| 90% seawater + 5% residual glycerol + 5% distilled water | 0.32 ± 0.01 | 0.17 | 53.60 | 0.0024 | |
| 90% seawater + 10% residual glycerol | 0.10 ± 0.02 | 0.01 | 10.97 | 0.0002 | |
| 90% mineral medium + 5% residual glycerol + 5% distilled water | 2.71 ± 0.96 | 1.22 | 44.95 | 0.0177 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takahashi, R.Y.U.; Castilho, N.A.S.; Silva, M.A.C.d.; Miotto, M.C.; Lima, A.O.d.S. Prospecting for Marine Bacteria for Polyhydroxyalkanoate Production on Low-Cost Substrates. Bioengineering 2017, 4, 60. https://doi.org/10.3390/bioengineering4030060
Takahashi RYU, Castilho NAS, Silva MACd, Miotto MC, Lima AOdS. Prospecting for Marine Bacteria for Polyhydroxyalkanoate Production on Low-Cost Substrates. Bioengineering. 2017; 4(3):60. https://doi.org/10.3390/bioengineering4030060
Chicago/Turabian StyleTakahashi, Rodrigo Yoji Uwamori, Nathalia Aparecida Santos Castilho, Marcus Adonai Castro da Silva, Maria Cecilia Miotto, and André Oliveira de Souza Lima. 2017. "Prospecting for Marine Bacteria for Polyhydroxyalkanoate Production on Low-Cost Substrates" Bioengineering 4, no. 3: 60. https://doi.org/10.3390/bioengineering4030060
APA StyleTakahashi, R. Y. U., Castilho, N. A. S., Silva, M. A. C. d., Miotto, M. C., & Lima, A. O. d. S. (2017). Prospecting for Marine Bacteria for Polyhydroxyalkanoate Production on Low-Cost Substrates. Bioengineering, 4(3), 60. https://doi.org/10.3390/bioengineering4030060