
Improved Anaerobic Fermentation of Wheat Straw by Alkaline Pre-Treatment and Addition of Alkali-Tolerant Microorganisms


 ,
, 
Abstract
:1. Introduction
2. Experimental Section
2.1. Substrate Pre-Treatment and Analysis
2.2. Inocula
2.3. BMP Assay
2.4. Reactor Operation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Substrate type | Inoculum | OD700 of the inoculum |
|---|---|---|---|
| R (Reference) | Untreated | StI | 0.163 |
| P1 | Pre-treated | StI | 0.163 |
| P2 | Pre-treated | S37°C | 0.160 |
| P3 | Pre-treated | S37°C | 0.293 |
2.5. Analysis of Process Parameters
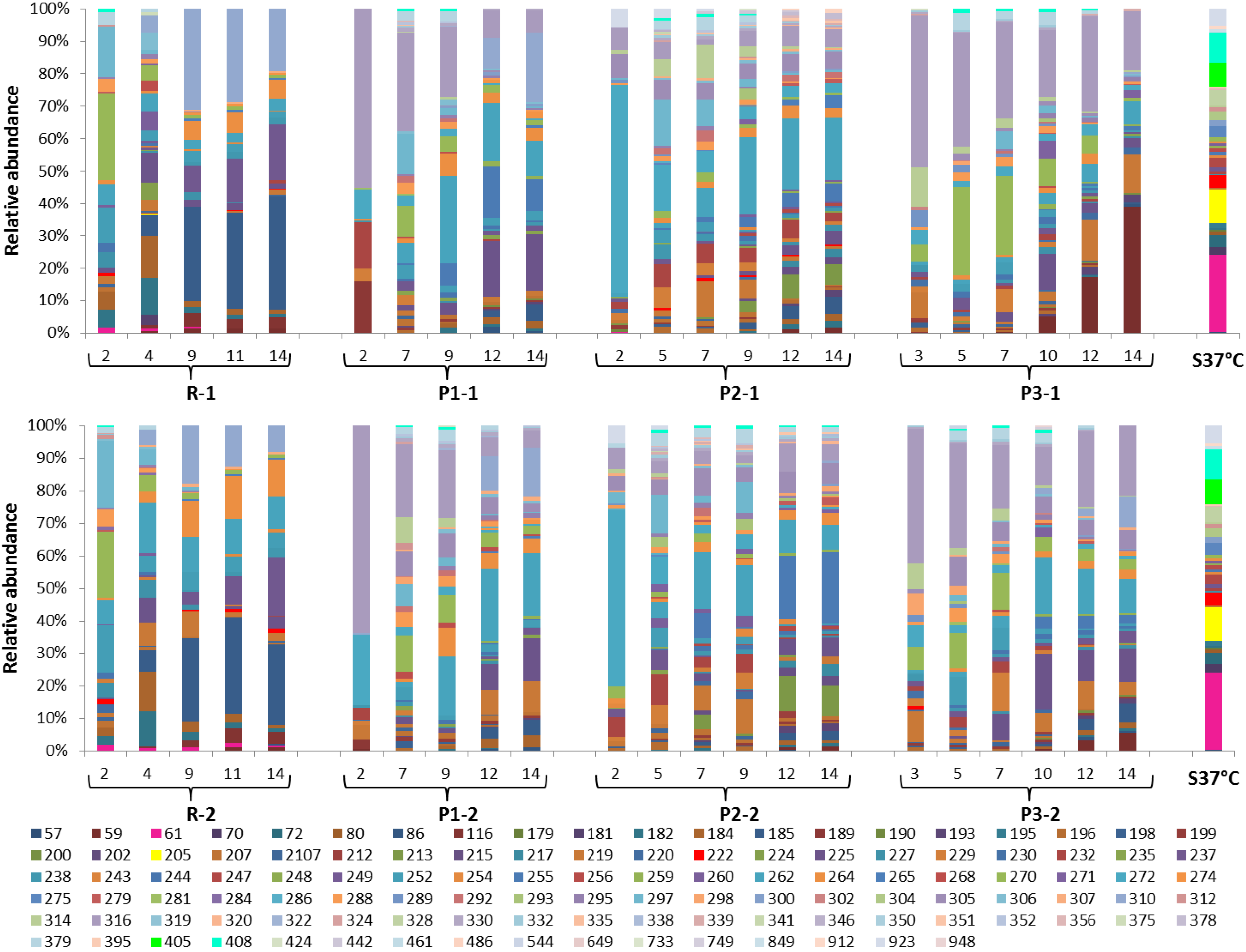
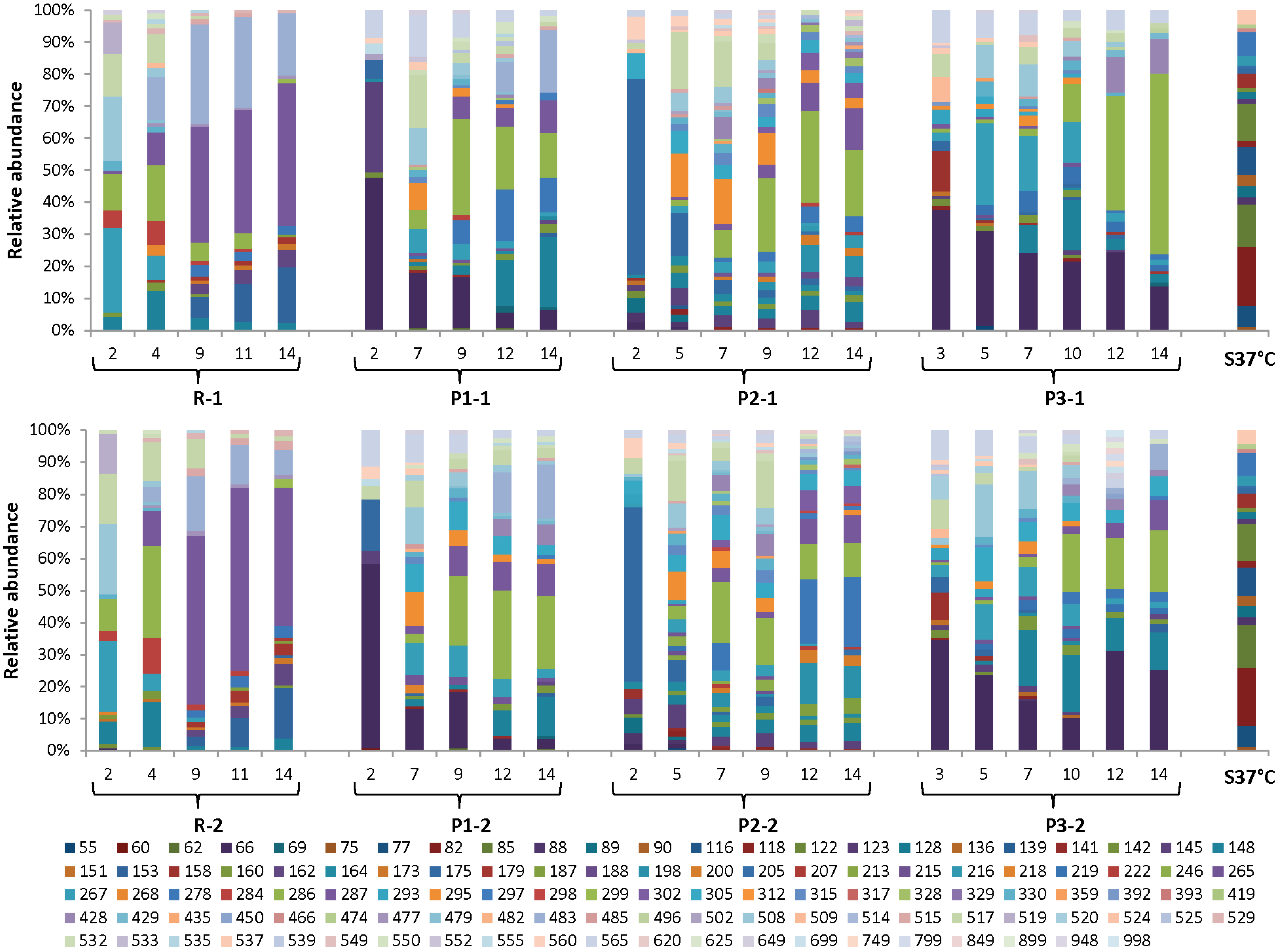
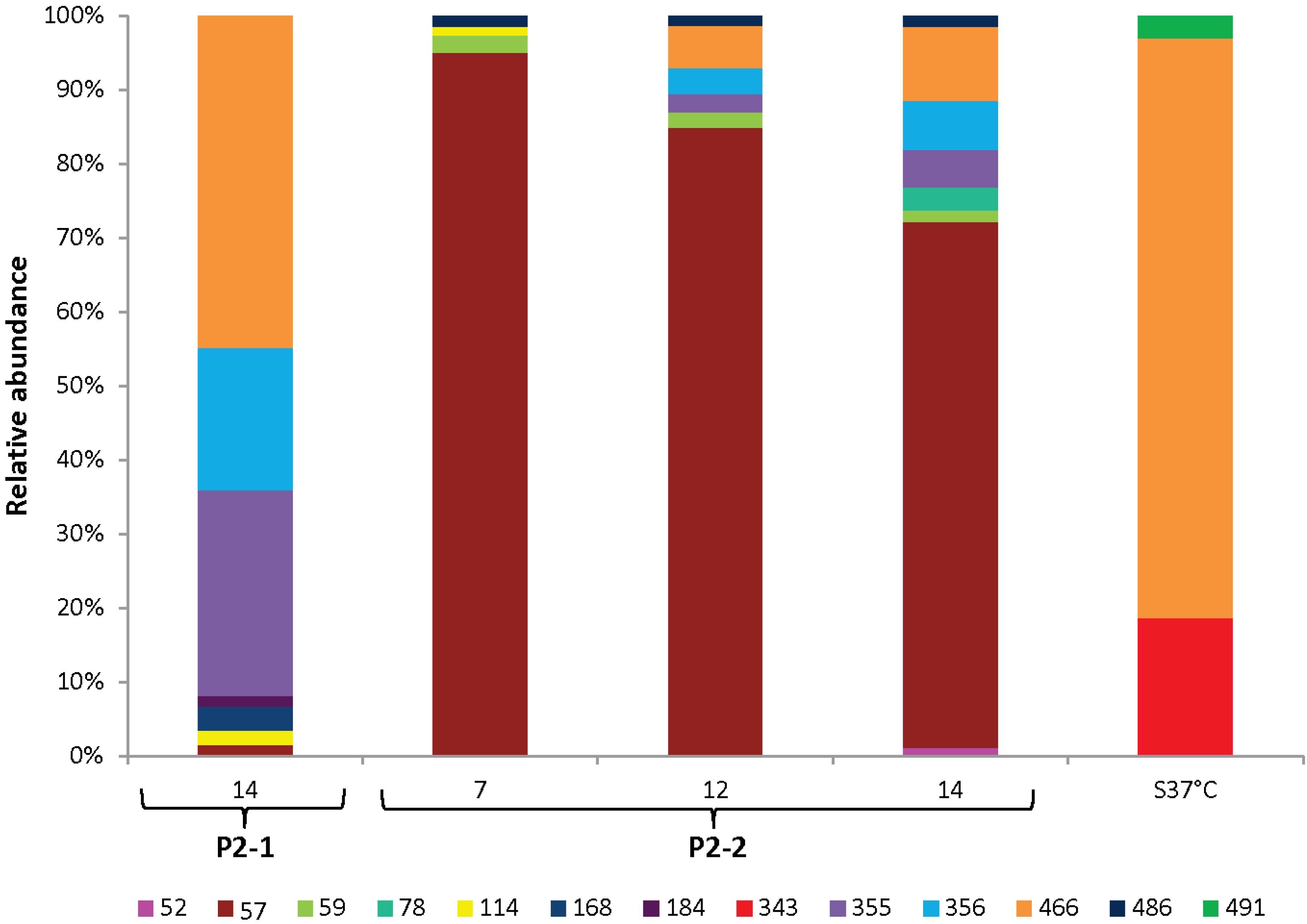
2.6. Microbial Community Analysis
2.7. Statistical Analysis
3. Results
3.1. Effect of Alkaline Pre-Treatment on Substrate Characteristics
| Parameter | Untreated wheat straw | Alkaline pre-treated wheat straw |
|---|---|---|
| TS (%Fresh·Mass) | 90.4 * | 14.0 (13.2–14.8) |
| VS (%TS) | 91.1 * | 85.2 (85.0–85.4) |
| Ash (g·kgTS−1) | 88.6 * | 148.4 (146.0–150.2) |
| Crude protein (g·kgTS−1) | 63.0 (±0.5) | 54.7 (±0.3) |
| Crude lipid (g·kgTS−1) | 16.8 (±0.4) | 7.2 (±0.2) |
| NFC (g·kgTS−1) | 33.1 (±2.6) | 38.6 (±1.7) |
| Cellulose (g·kgTS−1) | 466.1 (±0.2) | 460.5 (±2.6) |
| Hemicellulose (g·kgTS−1) | 237.7 (±4.7) | 194.4 (±4.5) |
| Lignin (g·kgTS−1) | 94.7 (±3.2) | 96.3 (±0.6) |
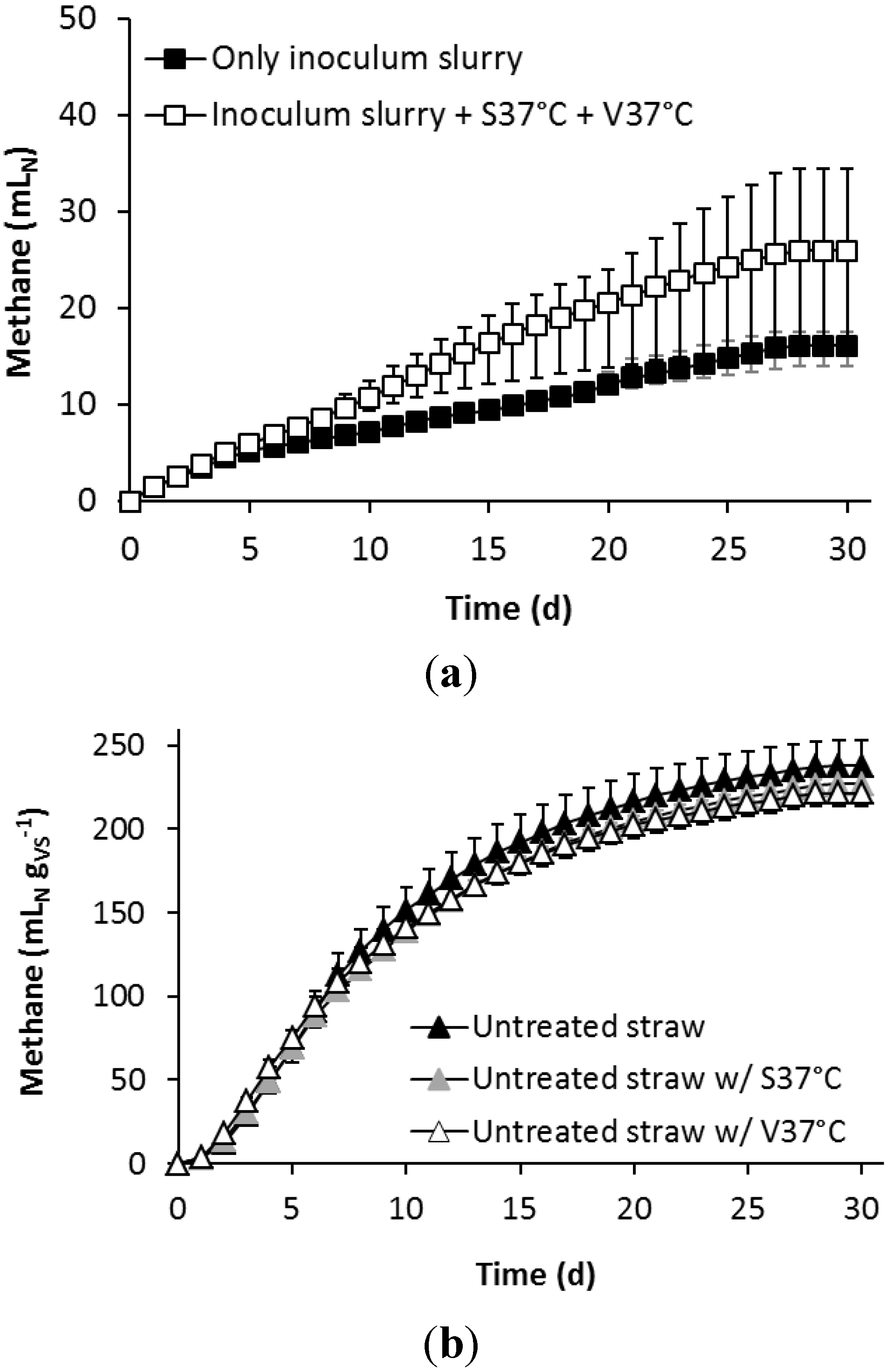
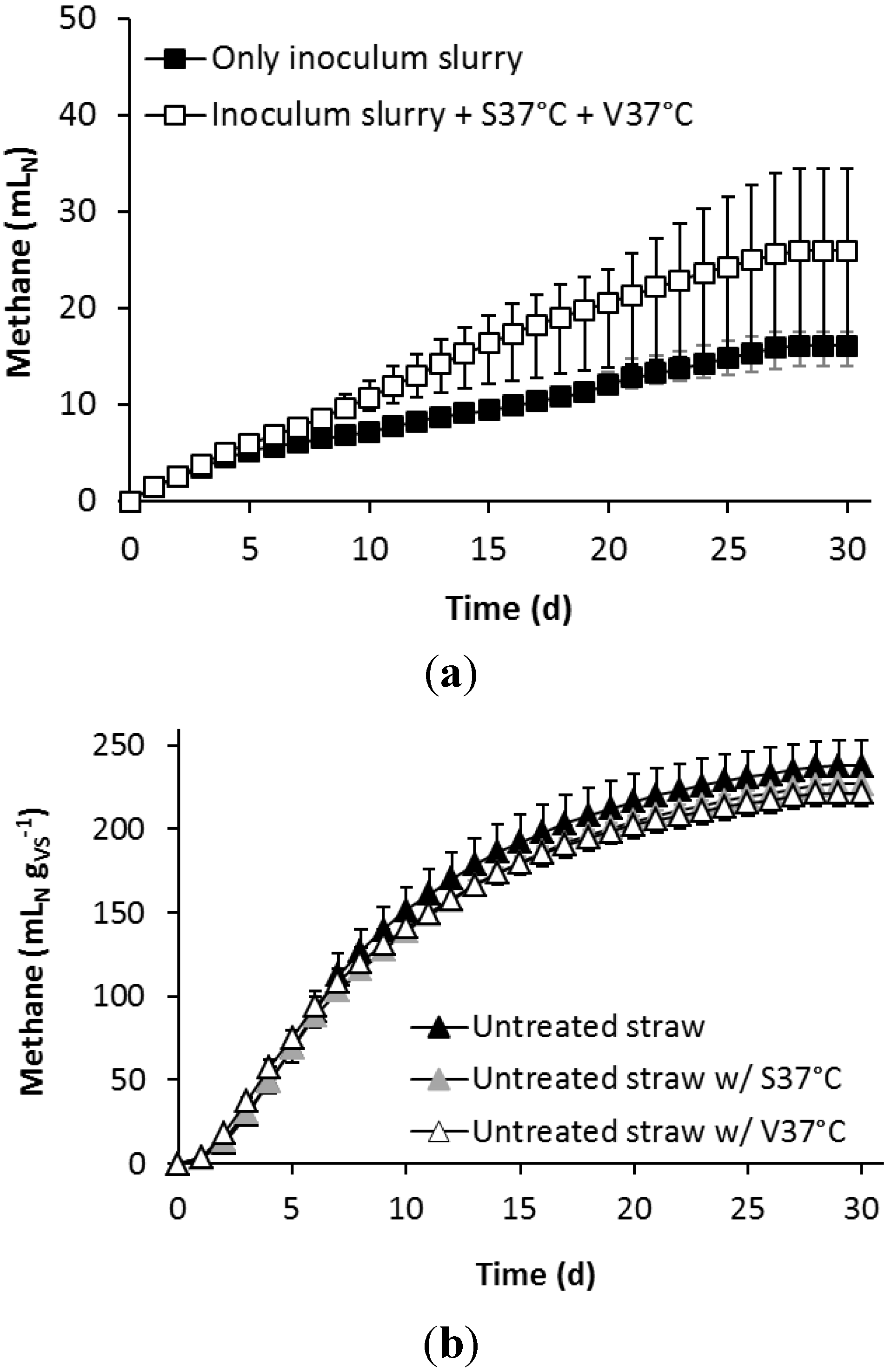
3.2. Effect of Alkaline Pre-Treatment and Bioaugmentation on the Biomethane Potential
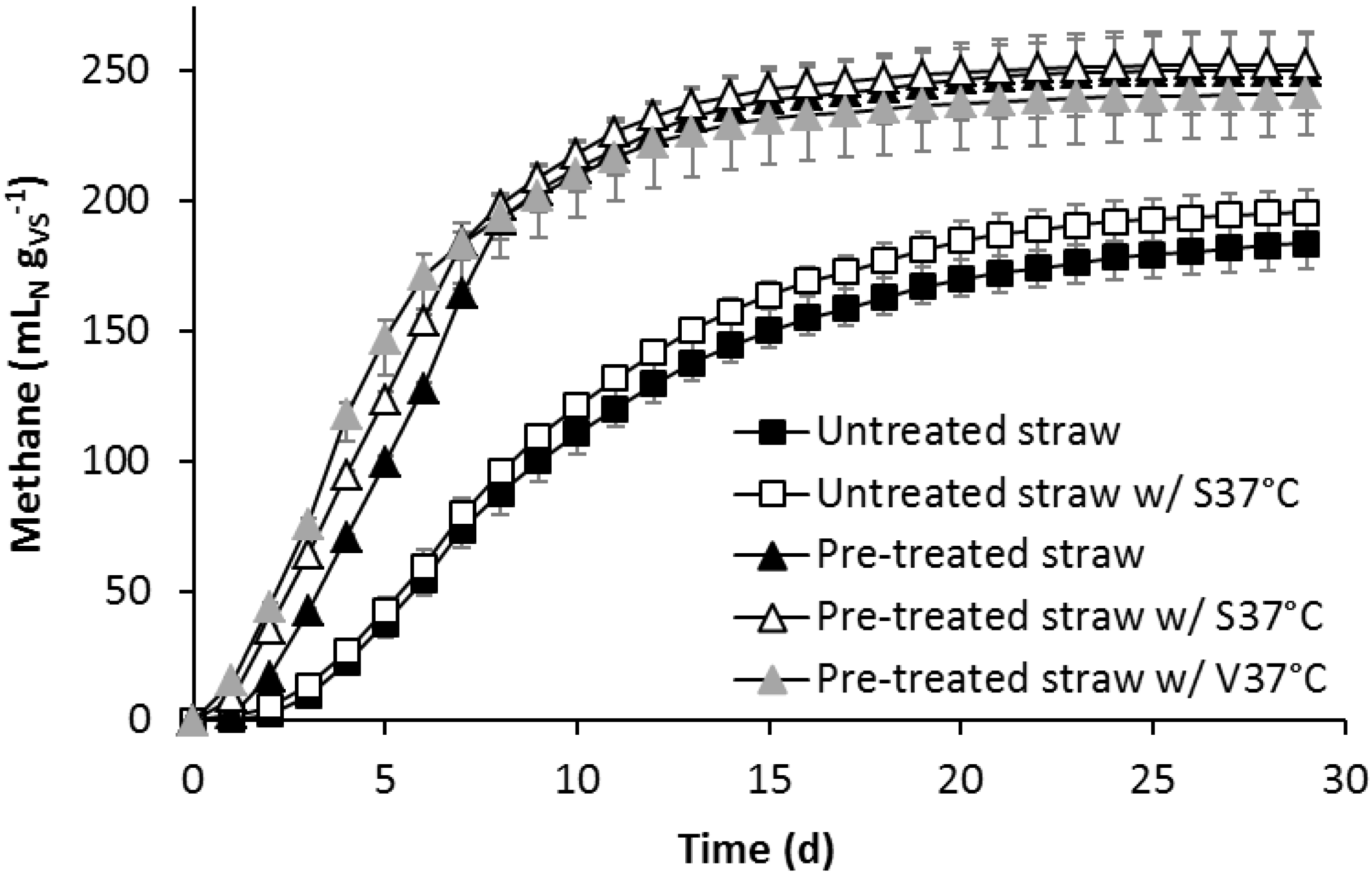
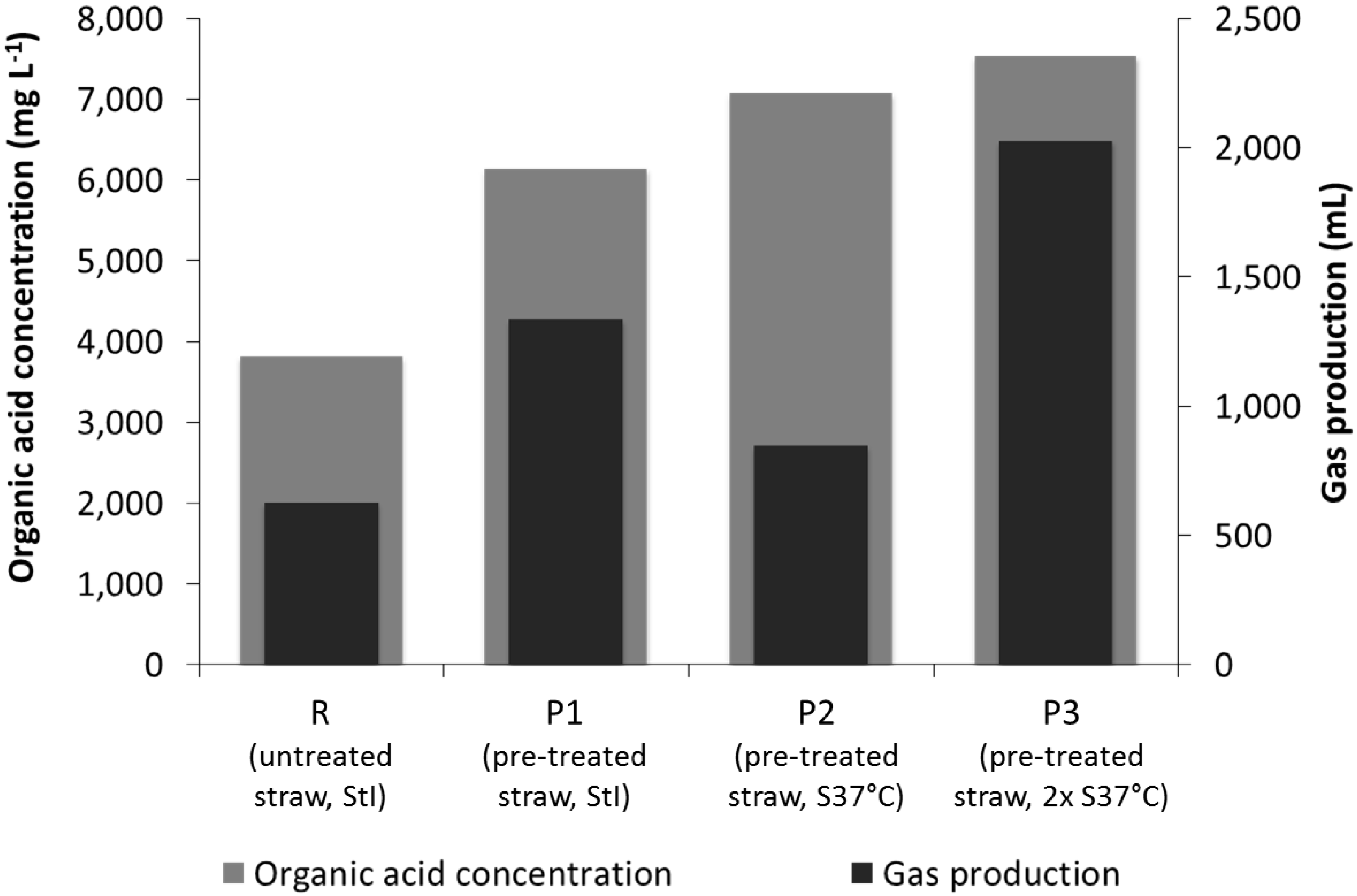
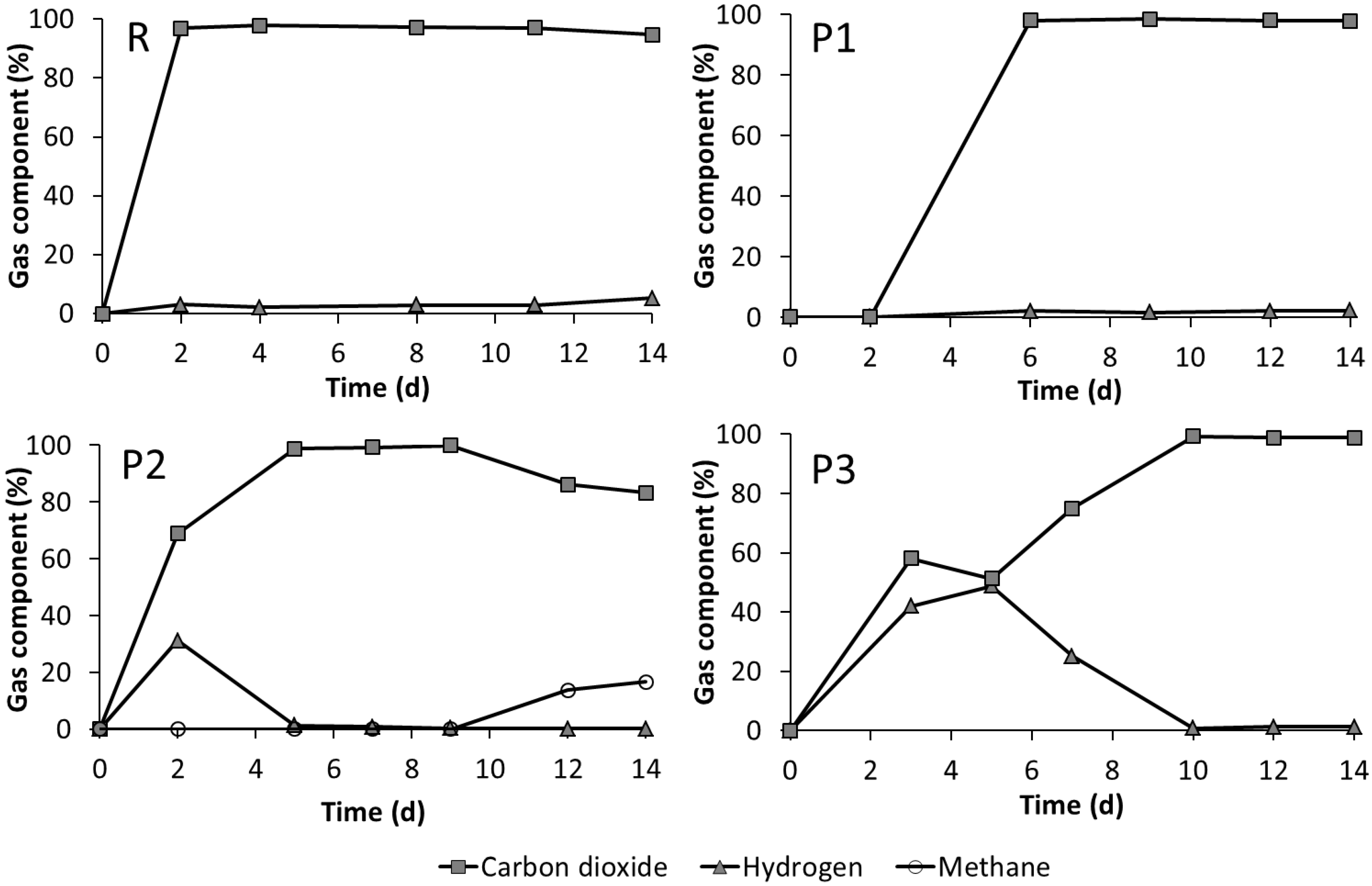
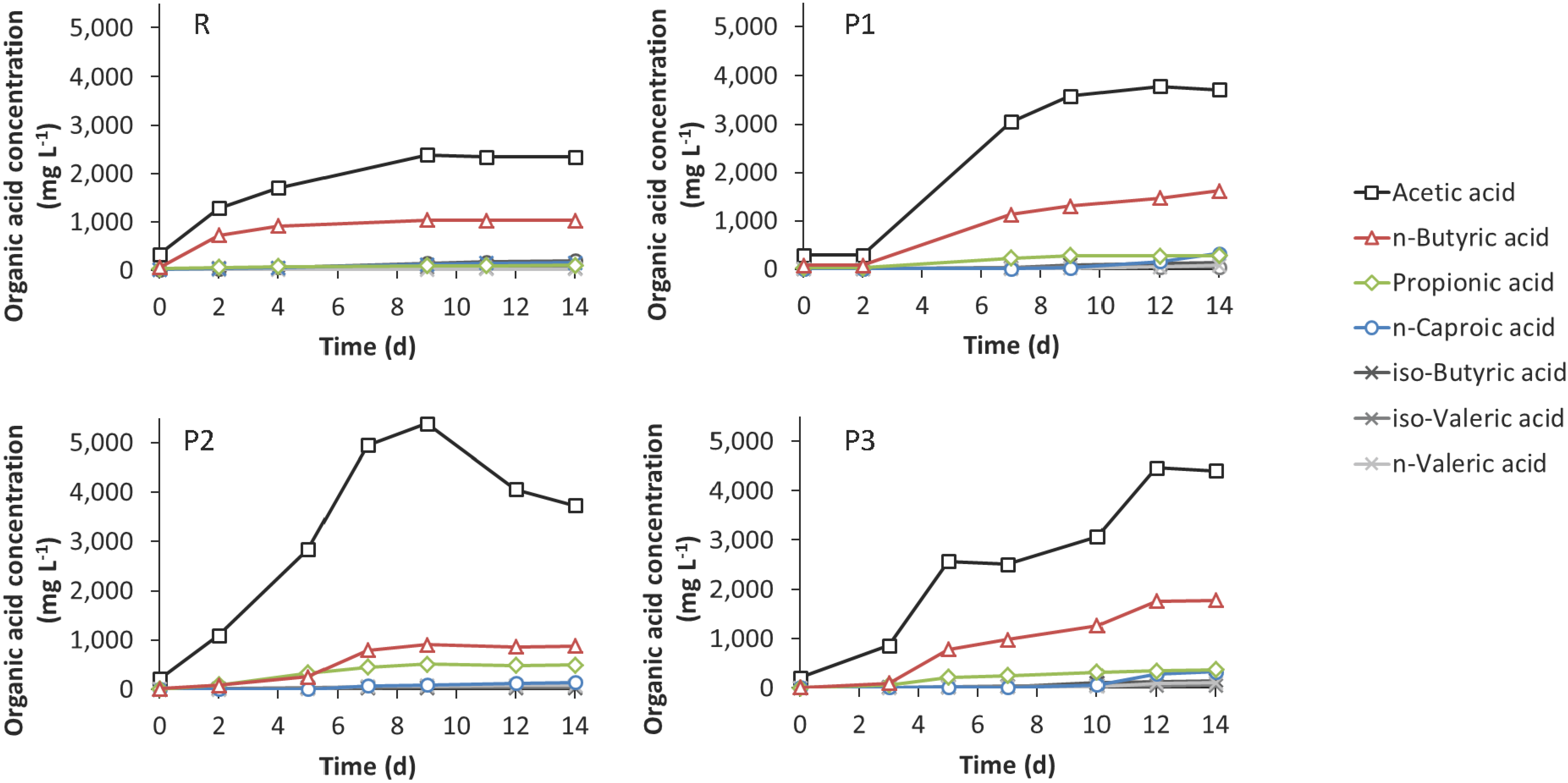
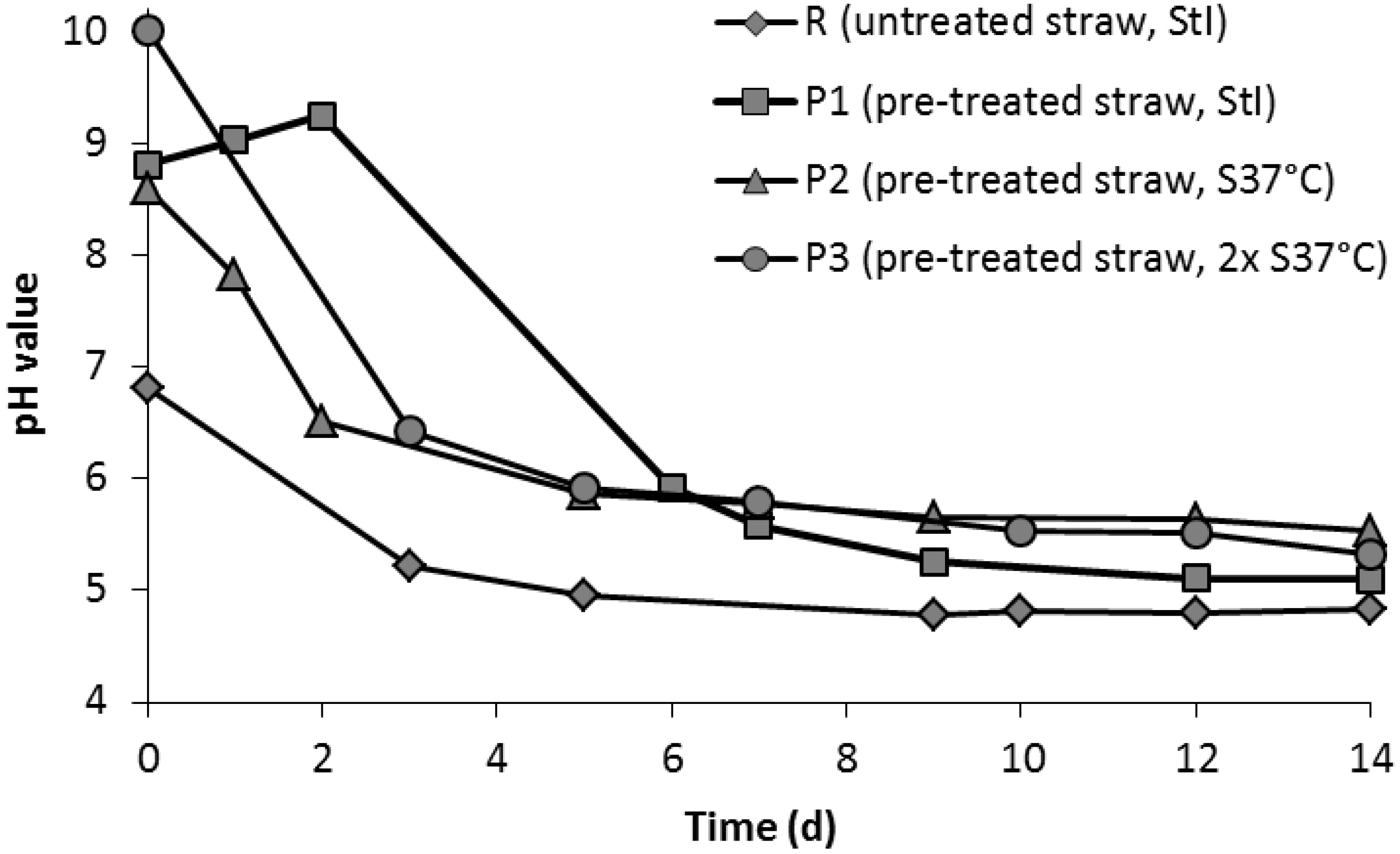
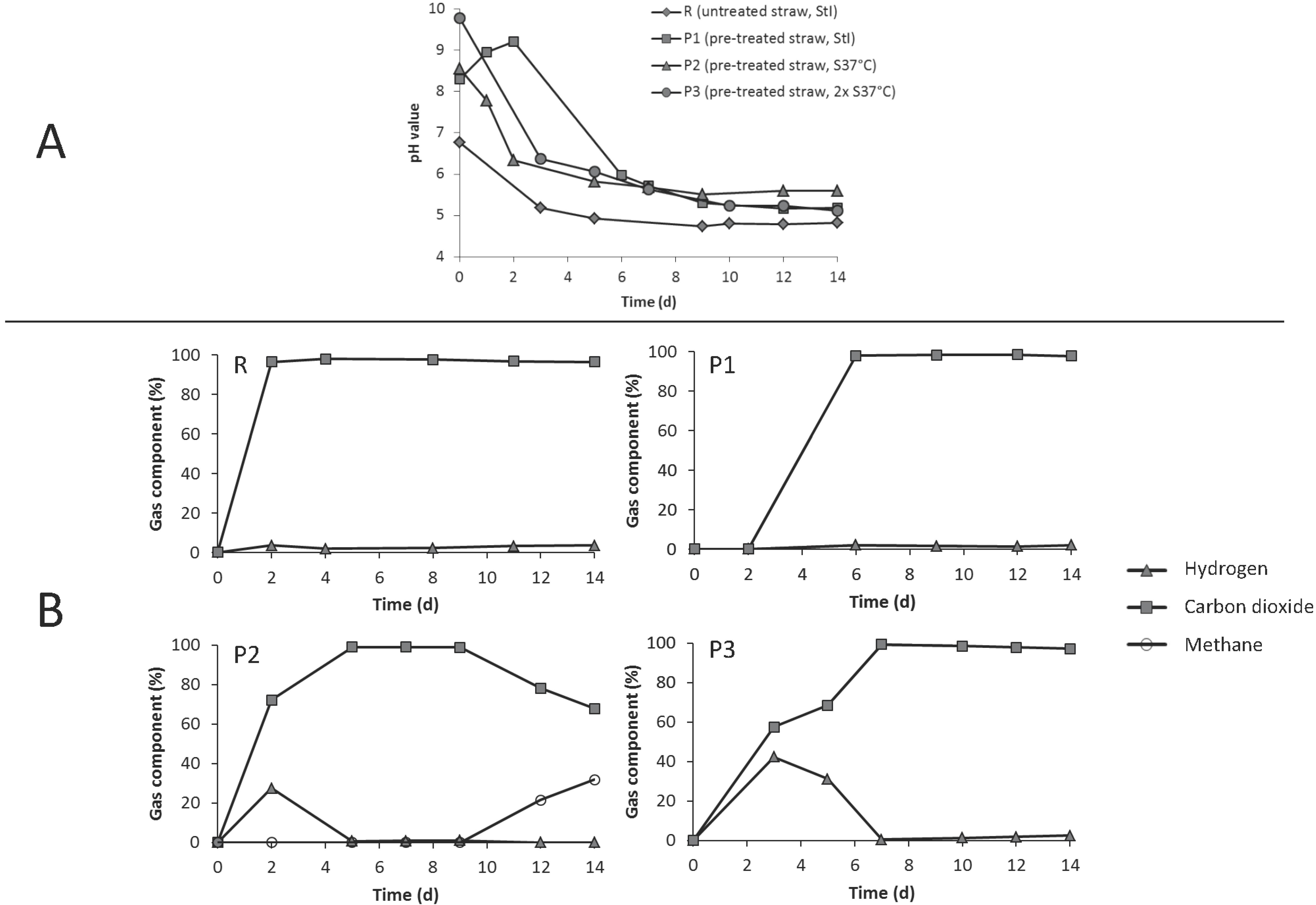
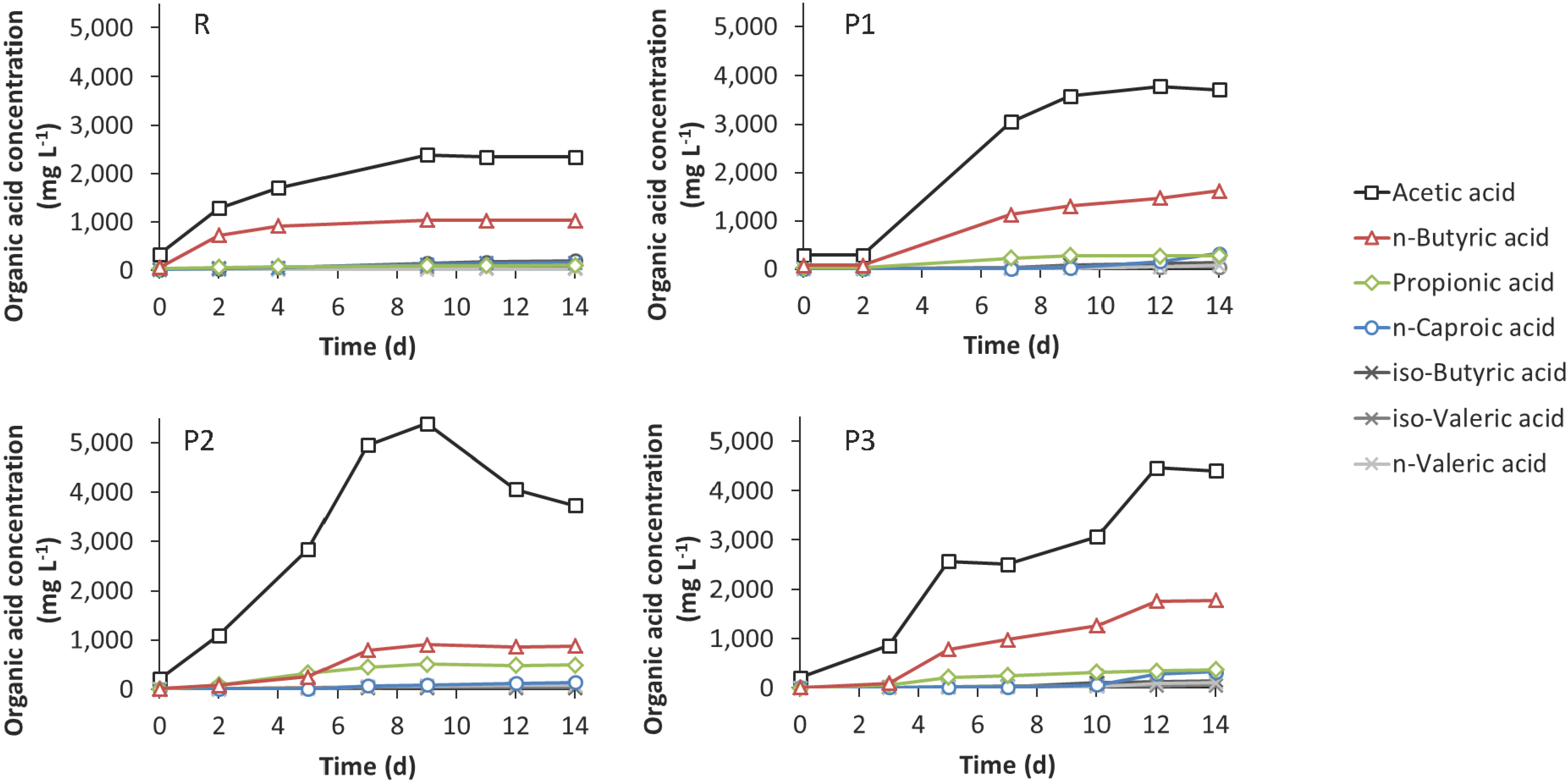
3.3. Leach-Bed Fermentation

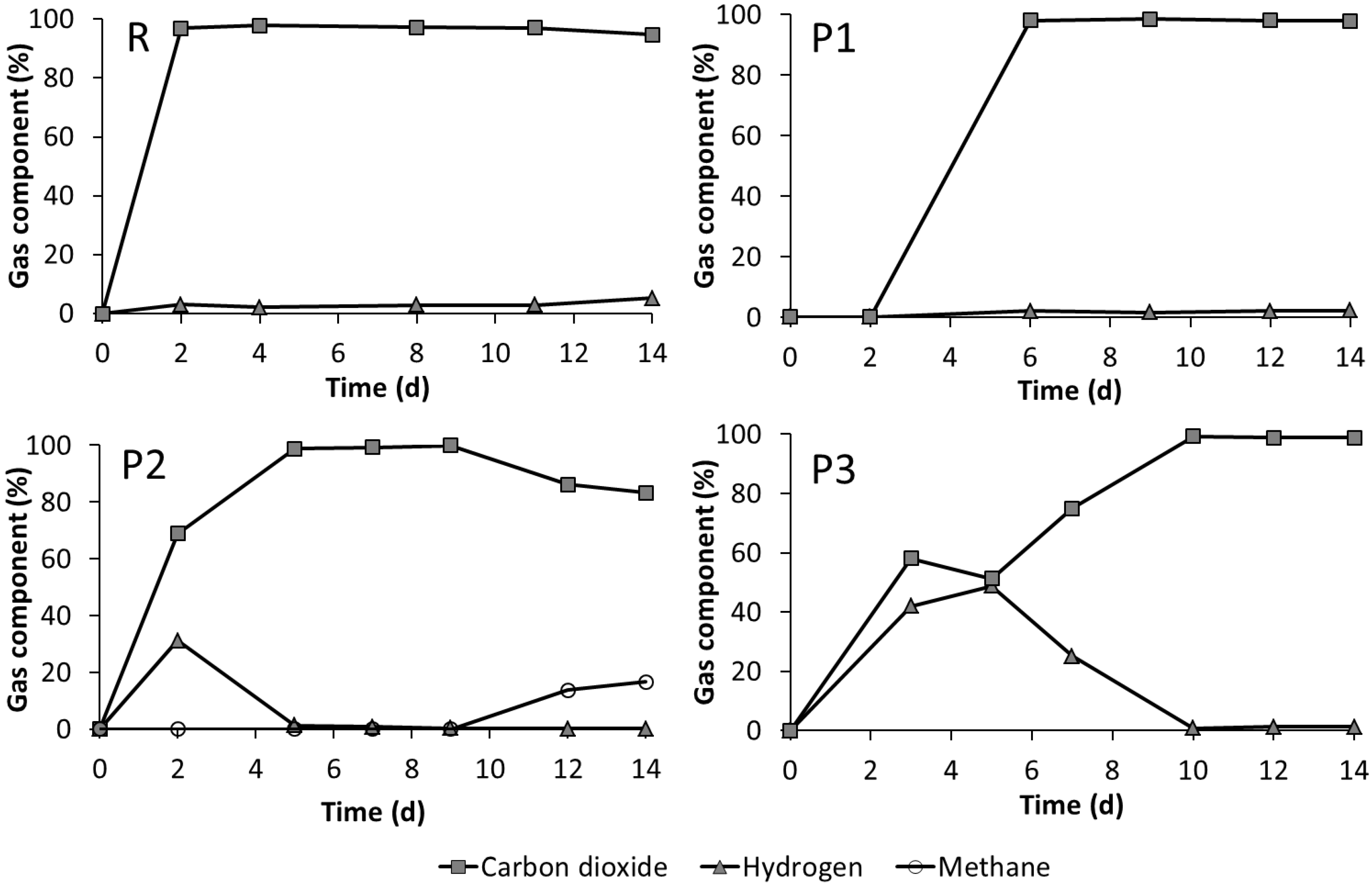
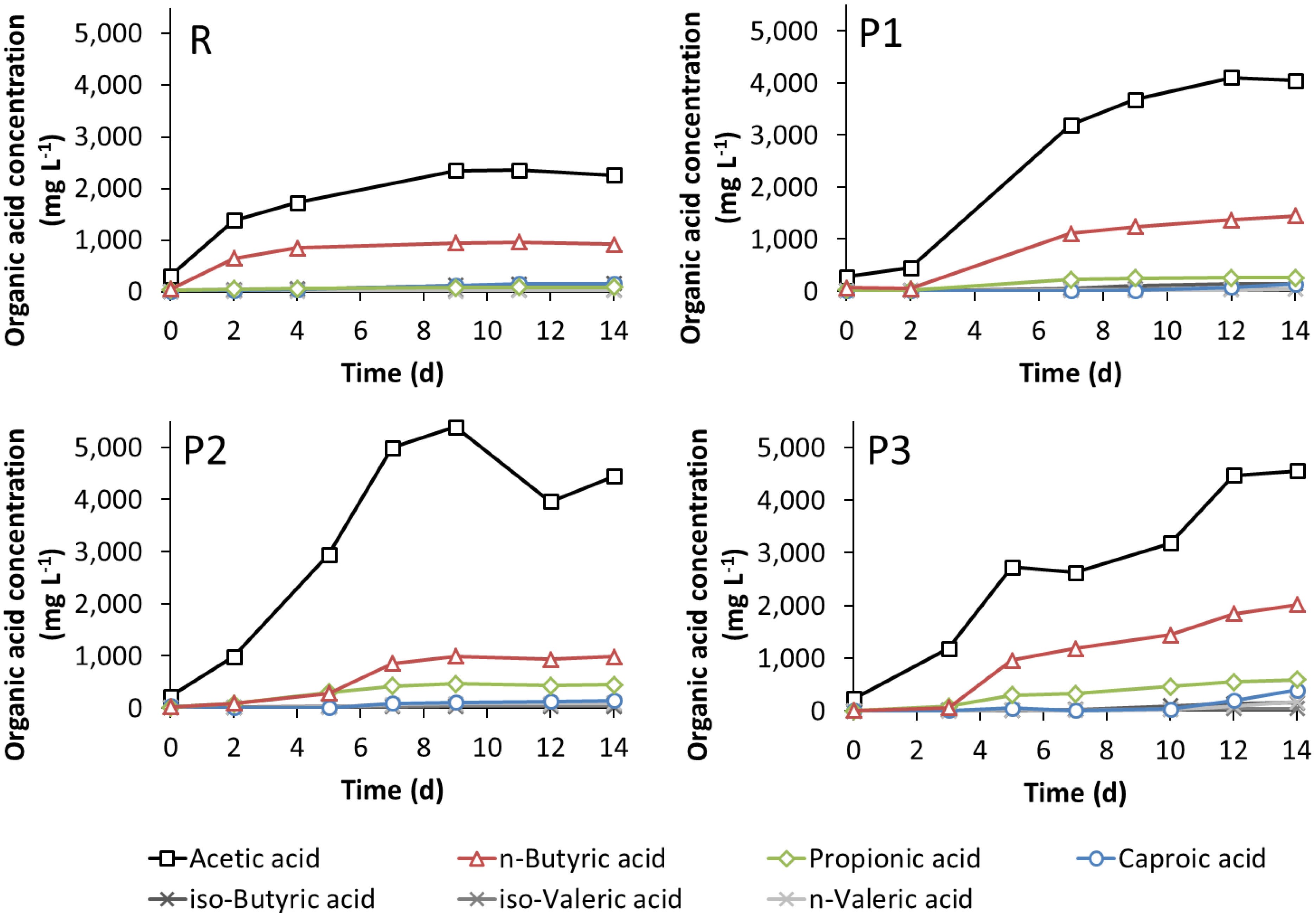
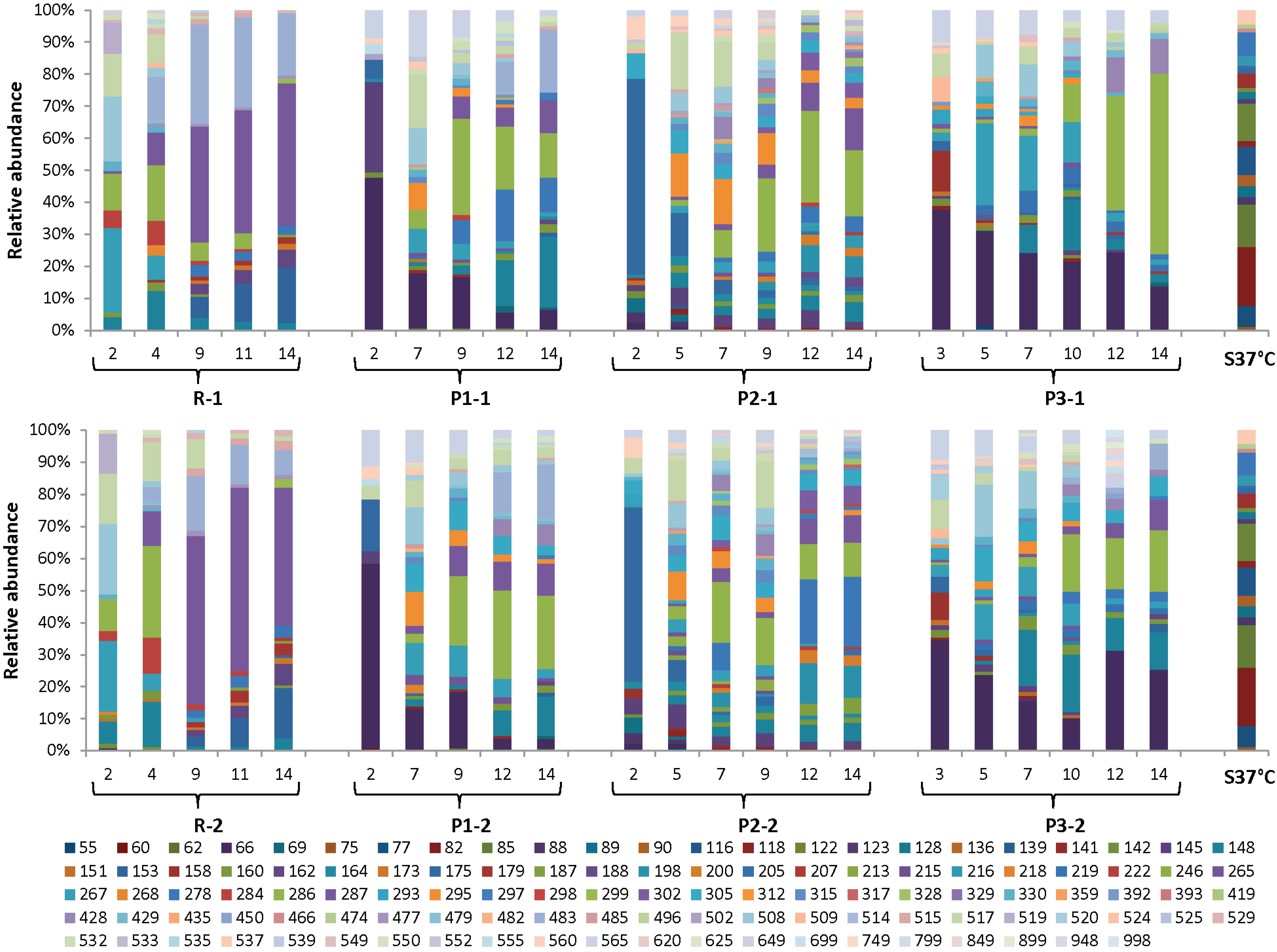
3.4. Dynamics of the Microbial Communities and Correlation with Abiotic Data
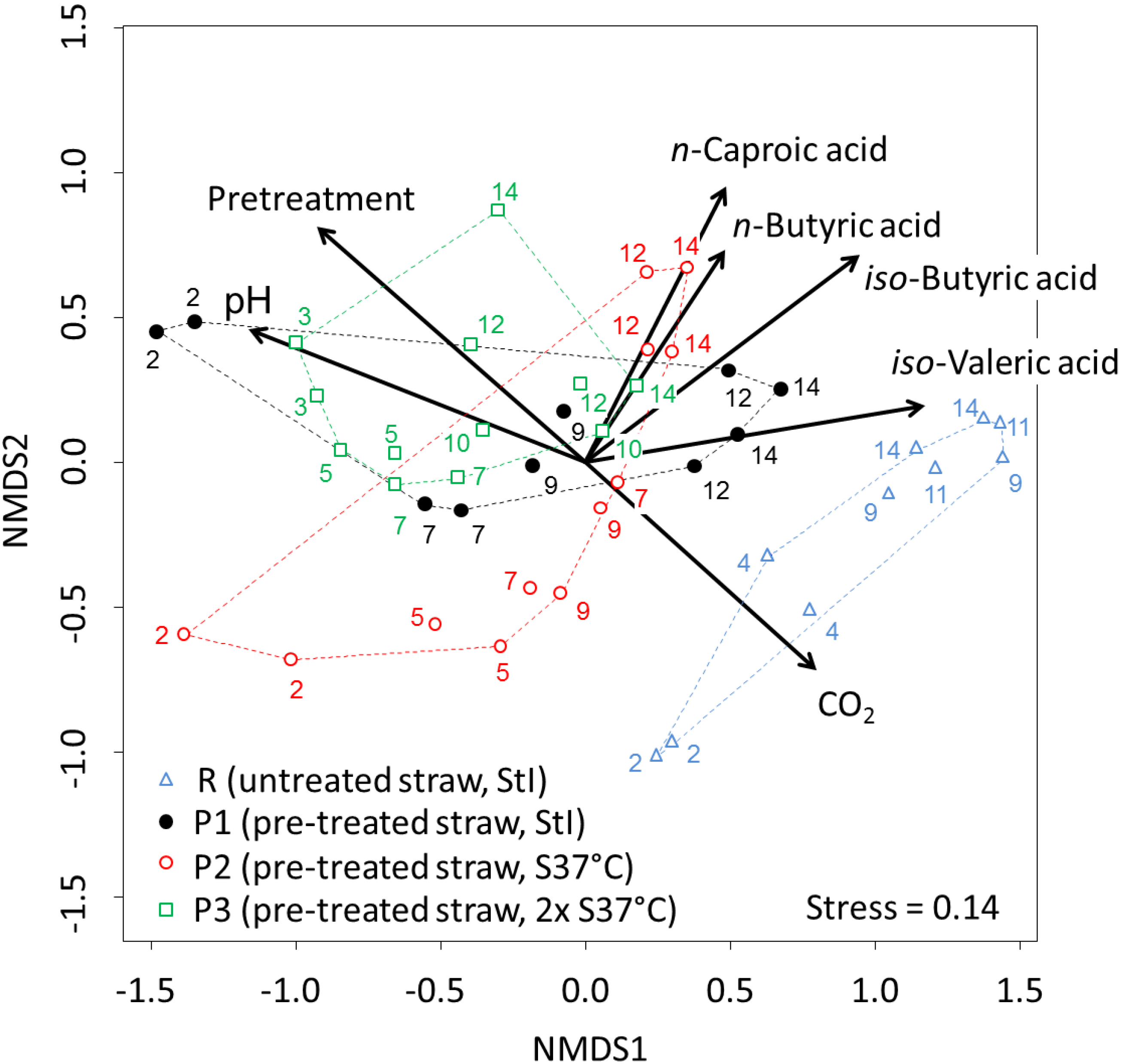
4. Discussion
4.1. Effect of Alkaline Pre-Treatment on the AD of Wheat Straw
4.2. Effect of Bioaugmentation on the AD of Alkaline Pre-Treated Wheat Straw
4.3. Effect of Bioaugmentation on Anaerobic Fermentation in a Leach-Bed Fermenter
5. Conclusions
Acknowledgements
Author Contributions
Conflict of Interests
References
- Chandra, R.; Takeuchi, H.; Hasegawa, T. Methane production from lignocellulosic agricultural crop wastes: A review in context to second generation of biofuel production. Renew. Sust. Energ. Rev. 2012, 16, 1462–1476. [Google Scholar] [CrossRef]
- Thomsen, S.T.; Spliid, H.; Østergård, H. Statistical prediction of biomethane potentials based on the composition of lignocellulosic biomass. Bioresource Technol. 2014, 154, 80–86. [Google Scholar] [CrossRef]
- Thanakoses, P.; Black, A.S.; Holtzapple, M.T. Fermentation of corn stover to carboxylic acids. Biotechnol. Bioeng. 2003, 83, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.J.; Kim, B.J.; Jeong, C.M.; Choi, J.D.; Ahn, Y.H.; Chang, H.N. Anaerobic organic acid production of food waste in once-a-day feeding and drawing-off bioreactor. Bioresource Technol. 2008, 99, 7866–7874. [Google Scholar] [CrossRef]
- Hendriks, A.T.W.M.; Zeeman, G. Pretreatments to enhance the digestibility of lignocellulosic biomass. Bioresource Technol. 2009, 100, 10–18. [Google Scholar] [CrossRef]
- Weiser, C.; Zeller, V.; Reinicke, F.; Wagner, B.; Majer, S.; Vetter, A.; Thraen, D. Integrated assessment of sustainable cereal straw potential and different straw-based energy applications in Germany. Appl. Energ. 2014, 114, 749–762. [Google Scholar] [CrossRef]
- Saha, B.C.; Cotta, M.A. Ethanol production from alkaline peroxide pretreated enzymatically saccharified wheat straw. Biotechnol. Prog. 2006, 22, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Margeot, A.; Hahn-Hagerdal, B.; Edlund, M.; Slade, R.; Monot, F. New improvements for lignocellulosic ethanol. Curr. Opin. Biotechnol. 2009, 20, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, B.; Wedwitschka, H.; Hofmann, J.; Denysenko, V.; Lorenz, H.; Liebetrau, J. Disintegration in the biogas sector—Technologies and effects. Bioresource Technol. 2014, 168, 2–6. [Google Scholar] [CrossRef]
- Palmowski, L.M.; Müller, J.A. Influence of the size reduction of organic waste on their anaerobic digestion. Water Sci. Technol. 1999, 41, 155–162. [Google Scholar]
- Kelsey, R.G.; Shafizadeh, F. Enhancement of cellulose accessibility and enzymatic hydrolysis by simultaneous wet milling. Biotechnol. Bioeng. 1980, 22, 1025–1036. [Google Scholar] [CrossRef]
- Fan, L.T.; Lee, Y.-H.; Gharpuray, M.M. The Nature of Lignocellulosics and Their Pretreatments for Enzymatic Hydrolysis. In Microbial Reactions; Springer: Berlin/Heidelberg, Germany, 1982; Volume 23, pp. 157–187. [Google Scholar]
- Sun, Y.; Cheng, J. Hydrolysis of lignocellulosic materials for ethanol production: A review. Bioresource Technol. 2002, 83, 1–11. [Google Scholar] [CrossRef]
- Taherzadeh, M.J.; Karimi, K. Pretreatment of lignocellulosic wastes to improve ethanol and biogas production: A review. Int. J. Mol. Sci. 2008, 9, 1621–1651. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C.; Whitman, W.B. Metabolic, phylogenetic, and ecological diversity of the methanogenic archaea. Ann. NY Acad. Sci. 2008, 1125, 171–189. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.J.; Hobbs, P.J.; Holliman, P.J.; Jones, D.L. Optimisation of the anaerobic digestion of agricultural resources. Bioresource Technol. 2008, 99, 7928–7940. [Google Scholar] [CrossRef]
- Weiland, P. Biogas production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2010, 85, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Lu, F.; Li, L.; Wu, Q.; Shao, L.; He, P. Self-adaption of methane-producing communities to pH disturbance at different acetate concentrations by shifting pathways and population interaction. Bioresource Technol. 2013, 140, 319–327. [Google Scholar] [CrossRef]
- Lin, Y.; Lu, F.; Shao, L.; He, P. Influence of bicarbonate buffer on the methanogenetic pathway during thermophilic anaerobic digestion. Bioresource Technol. 2013, 137, 245–253. [Google Scholar] [CrossRef]
- Gao, W.J.J.; Lin, H.J.; Leung, K.T.; Liao, B.Q. Influence of elevated pH shocks on the performance of a submerged anaerobic membrane bioreactor. Process Biochem. 2010, 45, 1279–1287. [Google Scholar] [CrossRef]
- Semrany, S.; Favier, L.; Djelal, H.; Taha, S.; Amrane, A. Bioaugmentation: Possible solution in the treatment of bio-refractory organic compounds (bio-ROCs). Biochem. Eng. J. 2012, 69, 75–86. [Google Scholar] [CrossRef]
- Akila, G.; Chandra, T.S. Stimulation of biomethanation by Clostridium sp. PXYL1 in coculture with a Methanosarcina strain PMET1 at psychrophilic temperatures. J. Appl. Microbiol. 2010, 108, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.B.; Mladenovska, Z.; Ahring, B.K. Bioaugmentation of a two-stage thermophilic (68 °C/55 °C) anaerobic digestion concept for improvement of the methane yield from cattle manure. Biotechnol. Bioeng. 2007, 97, 1638–1643. [Google Scholar] [CrossRef] [PubMed]
- Ács, N.; Bagi, Z.; Rakhely, G.; Kovacs, E.; Wirth, R.; Kovacs, K.L. Improvement of Biogas Production by Biotechnological Manipulation of the Microbial Population. In Proceeding of the 3rd IEEE International Symposium on Exploitation of Renewable Energy Sources, Subotica, Serbia, 11–12 March 2011.
- Bagi, Z.; Acs, N.; Balint, B.; Horvath, L.; Dobo, K.; Perei, K.R.; Rakhely, G.; Kovacs, K.L. Biotechnological intensification of biogas production. Appl. Microbiol. Biotechnol. 2007, 76, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Weiß, S.; Tauber, M.; Somitsch, W.; Meincke, R.; Müller, H.; Berg, G.; Guebitz, G.M. Enhancement of biogas production by addition of hemicellulolytic bacteria immobilised on activated zeolite. Water Res. 2010, 44, 1970–1980. [Google Scholar] [CrossRef]
- Sträuber, H.; Schröder, M.; Kleinsteuber, S. Metabolic and microbial community dynamics during the hydrolytic and acidogenic fermentation in a leach-bed process. Energ. Sustain. Soc. 2012, 2. [Google Scholar] [CrossRef]
- Naumann, C.; Bassler, R. Handbuch der landwirtschaftlichen Versuchs- und Untersuchungsmethodik (VDLUFA-Methodenbuch); VDLUFA-Verlag: Darmstadt, Germany, 2006; Volume Bd. III. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar]
- Porsch, K.; Wirth, B.; Tóth, E.M.; Schattenberg, F.; Nikolausz, M. Characterization of wheat straw-degrading anaerobic alkali-tolerant mixed cultures from soda lake sediments by molecular and cultivation techniques. Microb. Biotechnol. 2015. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Steinberg, L.M.; Regan, J.M. Phylogenetic comparison of the methanogenic communities from an acidic, oligotrophic fen and an anaerobic digester treating municipal wastewater sludge. Appl. Environ. Microbiol. 2008, 74, 6663–6671. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J. Multivariate Analysis of Ecological Communities in R: Vegan Tutorial, 2.0-6. 2011. Available online: http://cc.oulu.fi/~jarioksa/opetus/metodi/vegantutor.pdf (accessed on 19 December 2014).
- Pavlostathis, S.G.; Gossett, J.M. Alkaline treatment of wheat straw for increasing anaerobic biodegradability. Biotechnol. Bioeng. 1985, 27, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.; Bösch, P.; Friedl, A.; Amon, T. Analysis of methane potentials of steam-exploded wheat straw and estimation of energy yields of combined ethanol and methane production. J. Biotechnol. 2009, 142, 50–55. [Google Scholar] [CrossRef]
- Krishania, M.; Vijay, V.K.; Chandra, R. Methane fermentation and kinetics of wheat straw pretreated substrates co-digested with cattle manure in batch assay. Energy 2013, 57, 359–367. [Google Scholar] [CrossRef]
- Kaparaju, P.; Serrano, M.; Thomsen, A.B.; Kongjan, P.; Angelidaki, I. Bioethanol, biohydrogen and biogas production from wheat straw in a biorefinery concept. Bioresource Technol. 2009, 100, 2562–2568. [Google Scholar] [CrossRef]
- Sambusiti, C.; Monlau, F.; Ficara, E.; Carrere, H.; Malpei, F. A comparison of different pre-treatments to increase methane production from two agricultural substrates. Appl. Energ. 2013, 104, 62–70. [Google Scholar] [CrossRef]
- Kumar, P.; Barrett, D.M.; Delwiche, M.J.; Stroeve, P. Methods for pretreatment of lignocellulosic biomass for efficient hydrolysis and biofuel production. Ind. Eng. Chem. Res. 2009, 48, 3713–3729. [Google Scholar] [CrossRef]
- Xu, J.; Cheng, J.J.; Sharma-Shivappa, R.R.; Burns, J.C. Lime pretreatment of switchgrass at mild temperatures for ethanol production. Bioresource Technol. 2010, 101, 2900–2903. [Google Scholar] [CrossRef]
- Thompson, I.P.; van der Gast, C.J.; Ciric, L.; Singer, A.C. Bioaugmentation for bioremediation: The challenge of strain selection. Environ. Microbiol. 2005, 7, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Park, S.Y.; Zhu, J. Solid-state anaerobic digestion for methane production from organic waste. Renew. Sust. Energ. Rev. 2011, 15, 821–826. [Google Scholar] [CrossRef]
- Karthikeyan, O.P.; Visvanathan, C. Bio-energy recovery from high-solid organic substrates by dry anaerobic bio-conversion processes: A review. Rev. Environ. Sci. Biotechnol. 2012, 12, 257–284. [Google Scholar] [CrossRef]
- He, Y.; Pang, Y.; Liu, Y.; Li, X.; Wang, K. Physicochemical characterization of rice straw pretreated with sodium hydroxide in the solid state for enhancing biogas production. Energ. Fuel. 2008, 22, 2775–2781. [Google Scholar] [CrossRef]
- Pang, Y.Z.; Liu, Y.P.; Li, X.J.; Wang, K.S.; Yuan, H.R. Improving biodegradability and biogas production of corn stover through sodium hydroxide solid state pretreatment. Energ. Fuel. 2008, 22, 2761–2766. [Google Scholar] [CrossRef]
- Abbassi-Guendouz, A.; Trably, E.; Hamelin, J.; Dumas, C.; Steyer, J.P.; Delgenes, J.P.; Escudie, R. Microbial community signature of high-solid content methanogenic ecosystems. Bioresource Technol. 2013, 133, 256–262. [Google Scholar] [CrossRef]
- Martin, D.J. The site of reaction in solid-state digestion: A new hypothesis. Process Saf. Environ. 2001, 79, 29–37. [Google Scholar] [CrossRef]
- Karakashev, D.; Batstone, D.J.; Angelidaki, I. Influence of environmental conditions on methanogenic compositions in anaerobic biogas reactors. Appl. Environ. Microbiol. 2005, 71, 331–338. [Google Scholar] [CrossRef] [PubMed]
- De Vrieze, J.; Hennebel, T.; Boon, N.; Verstraete, W. Methanosarcina: The rediscovered methanogen for heavy duty biomethanation. Bioresource Technol. 2012, 112, 1–9. [Google Scholar]
- Staley, B.F.; de los Reyes, F.L.; Barlaz, M.A. Effect of spatial differences in microbial activity, pH, and substrate levels on methanogenesis initiation in refuse. Appl. Environ. Microbiol. 2011, 77, 2381–2391. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.-H.; Yu, H.-Q. Application of rumen microorganisms for enhanced anaerobic fermentation of corn stover. Process Biochem. 2005, 40, 2371–2377. [Google Scholar] [CrossRef]
- Li, L.; Yang, X.; Li, X.; Zheng, M.; Chen, J.; Zhang, Z. The influence of inoculum sources on anaerobic biogasification of NaOH-treated corn stover. Energ. Source Part A 2010, 33, 138–144. [Google Scholar] [CrossRef]
- Lopes, W.S.; Leite, V.D.; Prasad, S. Influence of inoculum on performance of anaerobic reactors for treating municipal solid waste. Bioresource Technol. 2004, 94, 261–266. [Google Scholar] [CrossRef]
- Ali Shah, F.; Mahmood, Q.; Maroof Shah, M.; Pervez, A.; Ahmad Asad, S. Microbial ecology of anaerobic digesters: The key players of anaerobiosis. Sci. World J. 2014. [Google Scholar] [CrossRef]
- Forster-Carneiro, T.; Perez, M.; Romero, L.I.; Sales, D. Dry-thermophilic anaerobic digestion of organic fraction of the municipal solid waste: Focusing on the inoculum sources. Bioresource Technol. 2007, 98, 3195–3203. [Google Scholar] [CrossRef]
Appendix
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sträuber, H.; Bühligen, F.; Kleinsteuber, S.; Nikolausz, M.; Porsch, K. Improved Anaerobic Fermentation of Wheat Straw by Alkaline Pre-Treatment and Addition of Alkali-Tolerant Microorganisms. Bioengineering 2015, 2, 66-93. https://doi.org/10.3390/bioengineering2020066
Sträuber H, Bühligen F, Kleinsteuber S, Nikolausz M, Porsch K. Improved Anaerobic Fermentation of Wheat Straw by Alkaline Pre-Treatment and Addition of Alkali-Tolerant Microorganisms. Bioengineering. 2015; 2(2):66-93. https://doi.org/10.3390/bioengineering2020066
Chicago/Turabian StyleSträuber, Heike, Franziska Bühligen, Sabine Kleinsteuber, Marcell Nikolausz, and Katharina Porsch. 2015. "Improved Anaerobic Fermentation of Wheat Straw by Alkaline Pre-Treatment and Addition of Alkali-Tolerant Microorganisms" Bioengineering 2, no. 2: 66-93. https://doi.org/10.3390/bioengineering2020066
APA StyleSträuber, H., Bühligen, F., Kleinsteuber, S., Nikolausz, M., & Porsch, K. (2015). Improved Anaerobic Fermentation of Wheat Straw by Alkaline Pre-Treatment and Addition of Alkali-Tolerant Microorganisms. Bioengineering, 2(2), 66-93. https://doi.org/10.3390/bioengineering2020066