

The Effect of Static Magnetic Fields of Different Strengths and Polarities on Cytokine Production by Human Lymphocytes In Vitro

 , ,
, ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
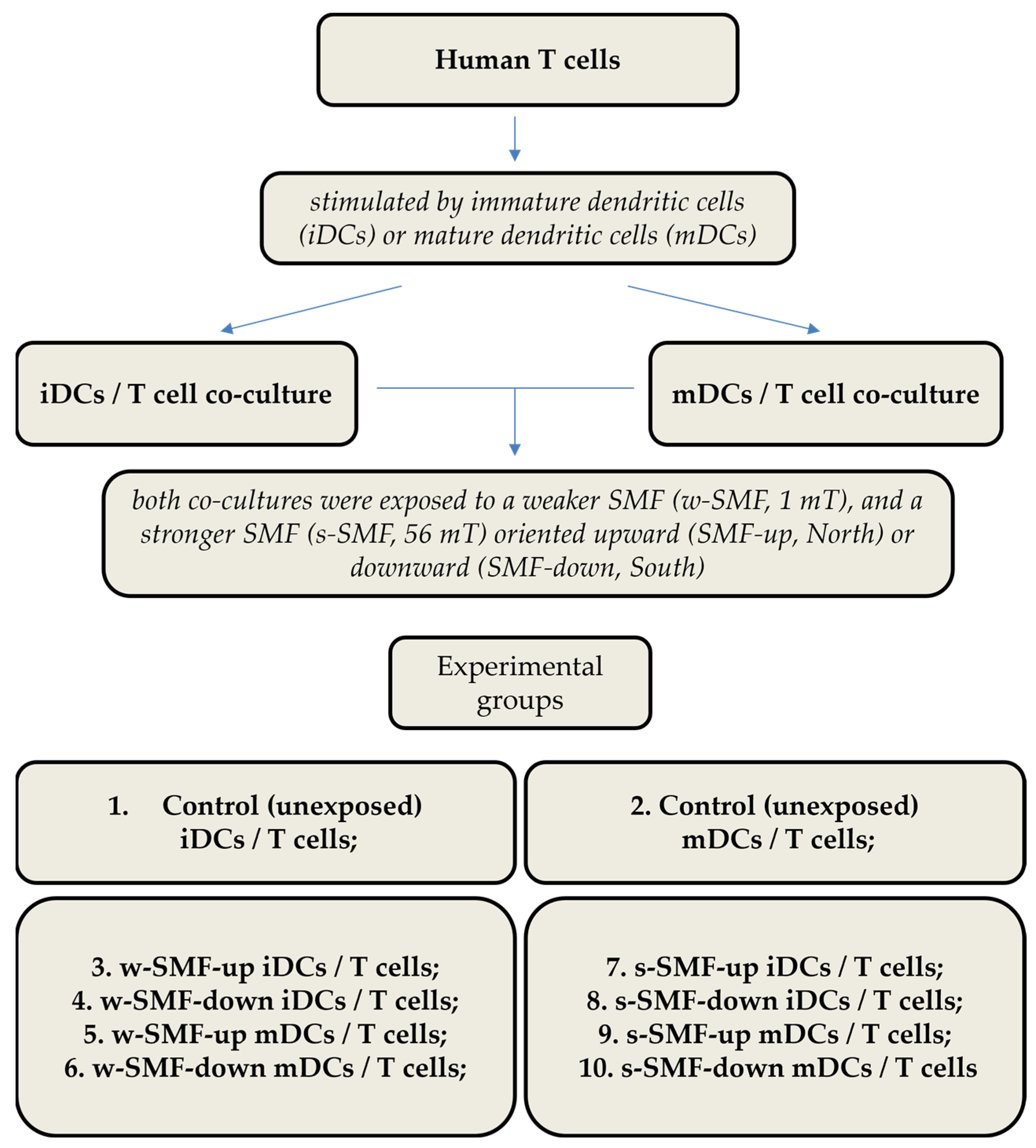
2.1. Study Design
2.2. Cells
2.3. Allogeneic T-Cell Activation
2.4. Cytokine Assays
2.5. Cell Viability Assays
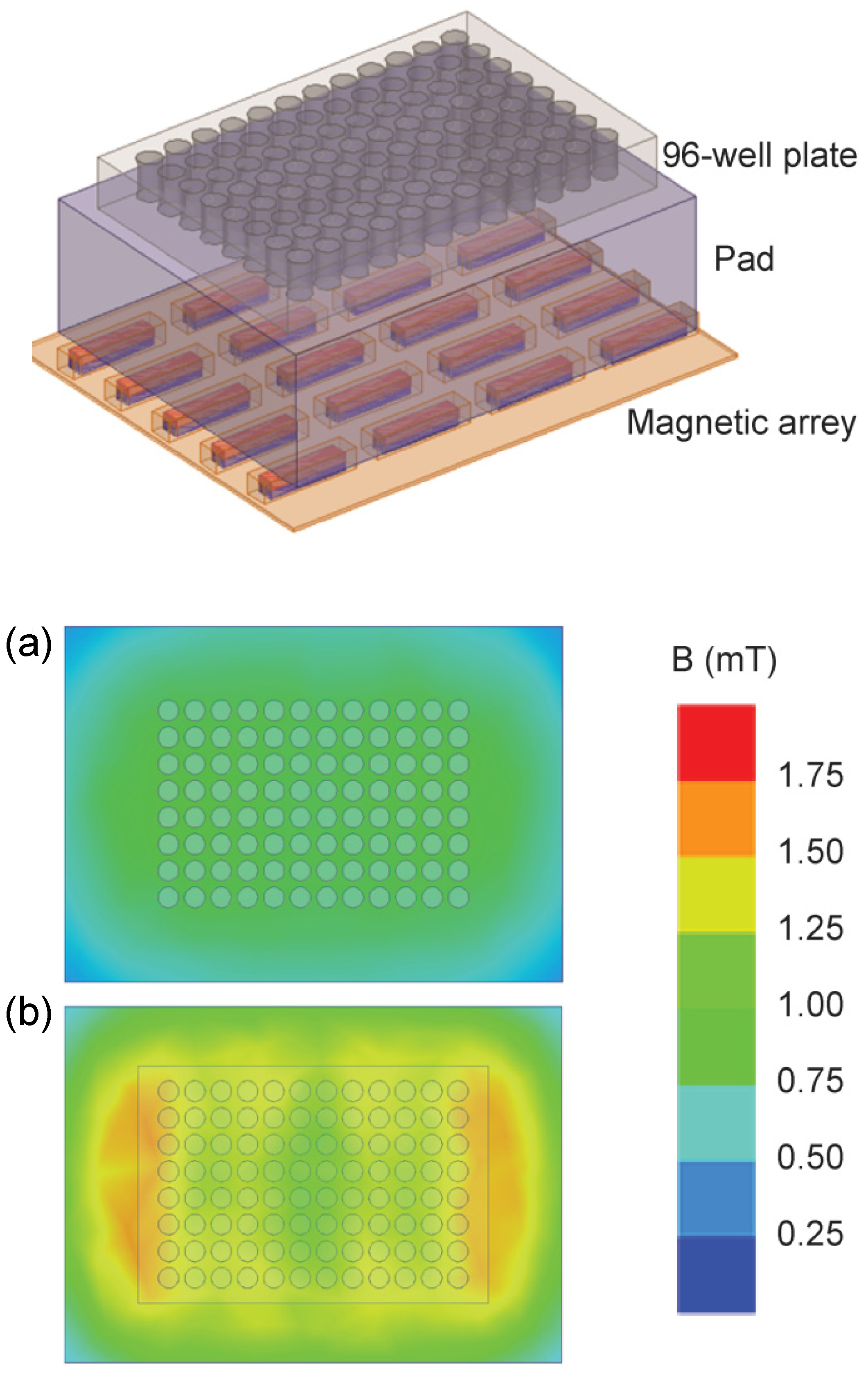
2.6. Magnetic Field
2.7. Statistical Analyses
3. Results
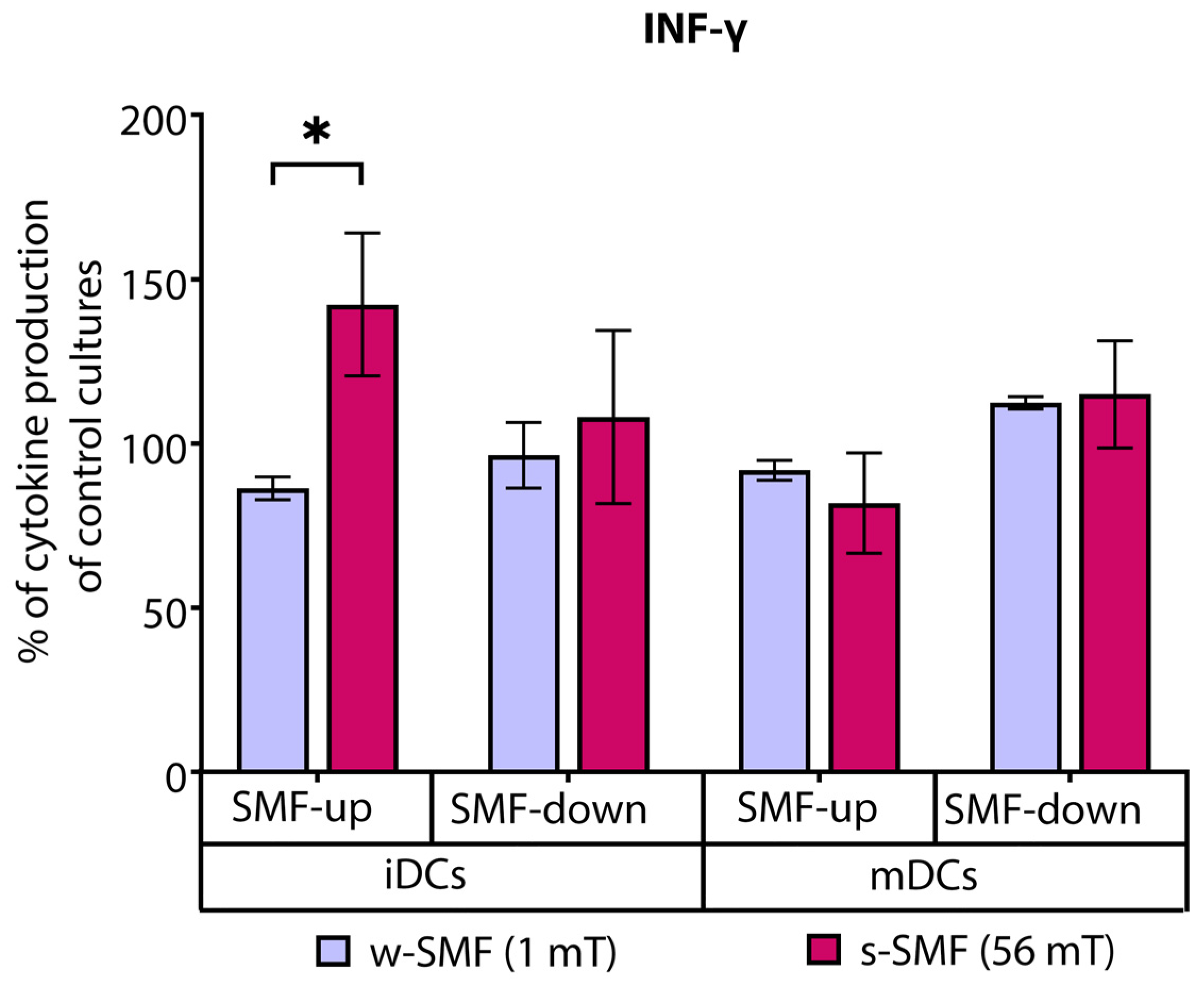
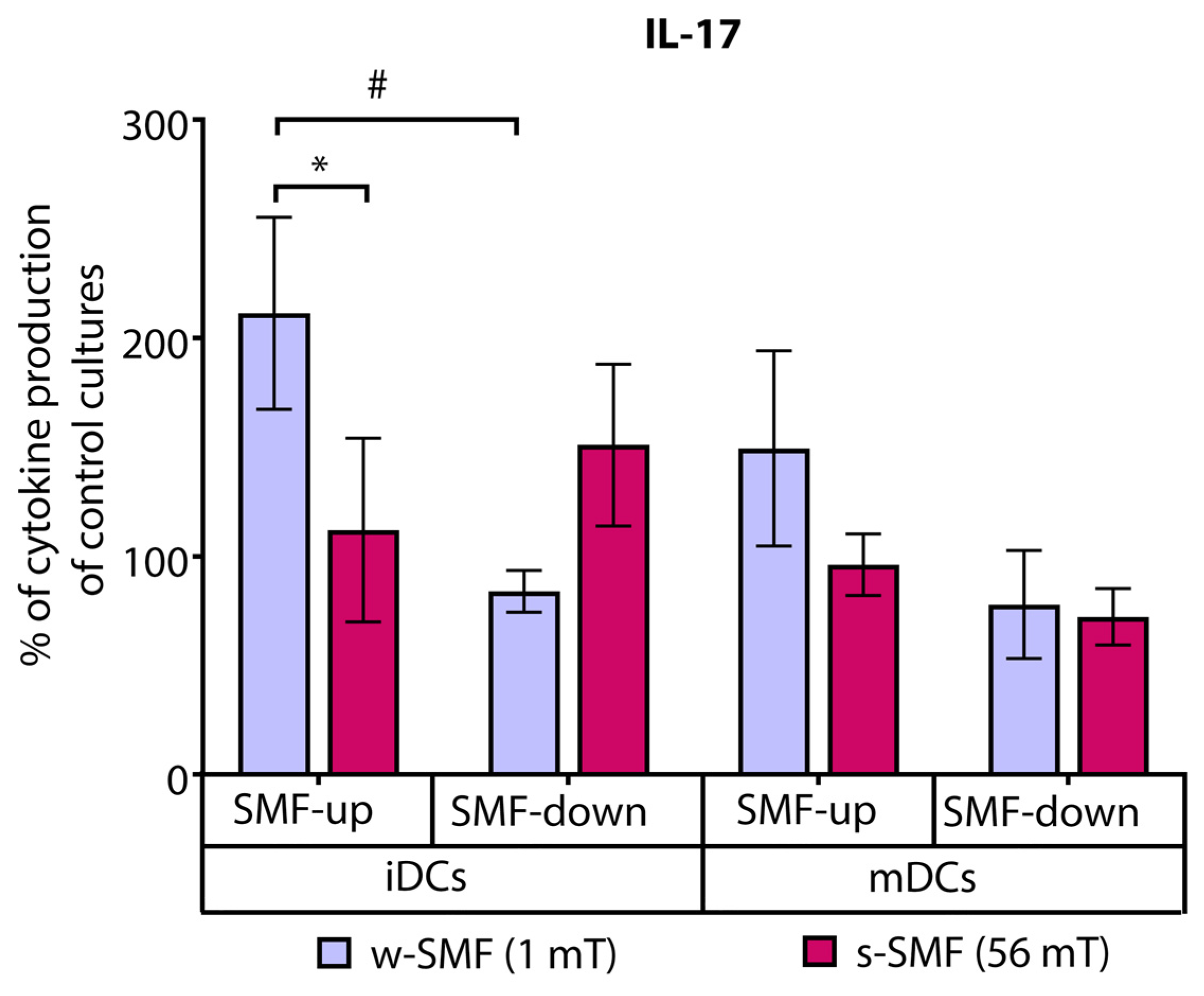
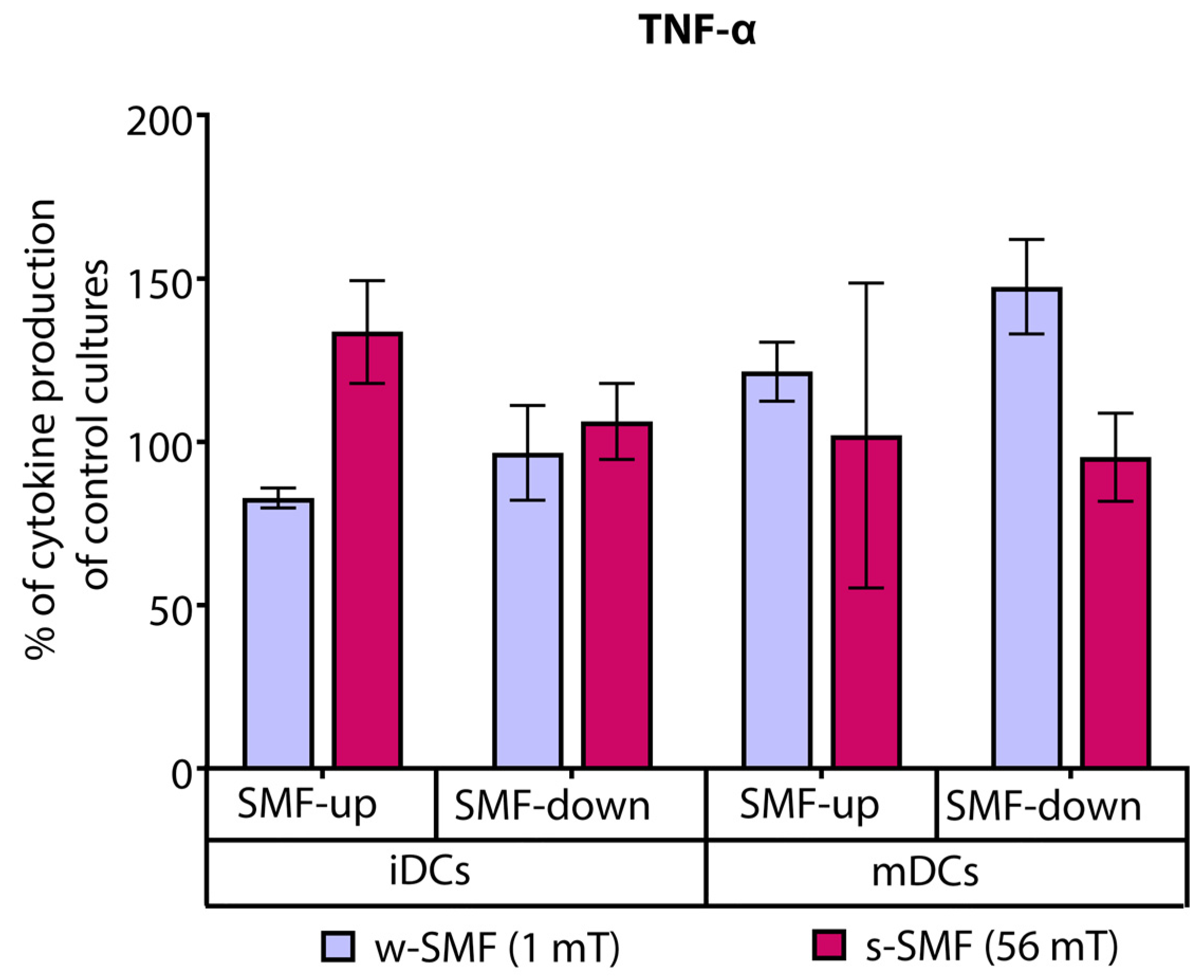
3.1. Cytokine Release
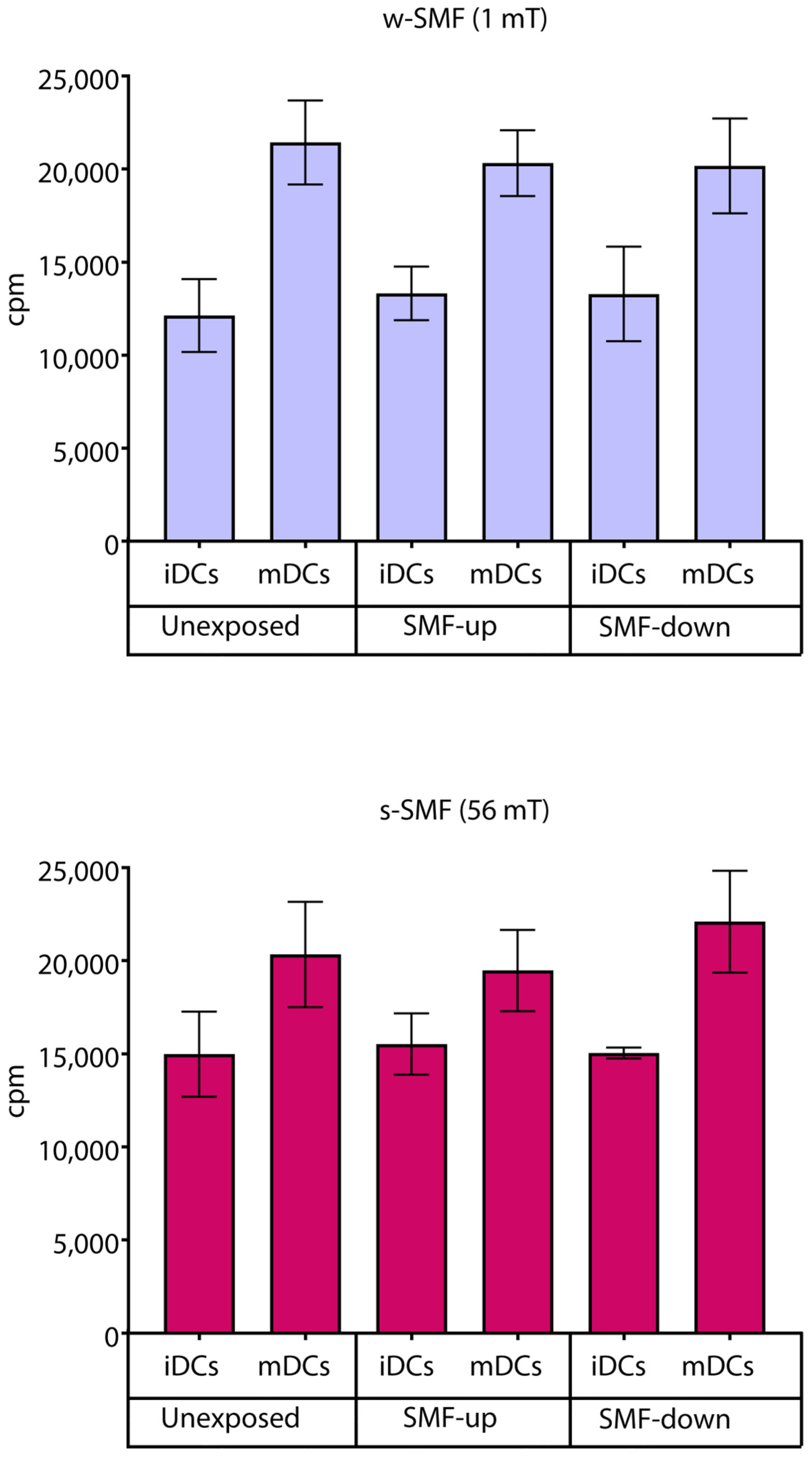
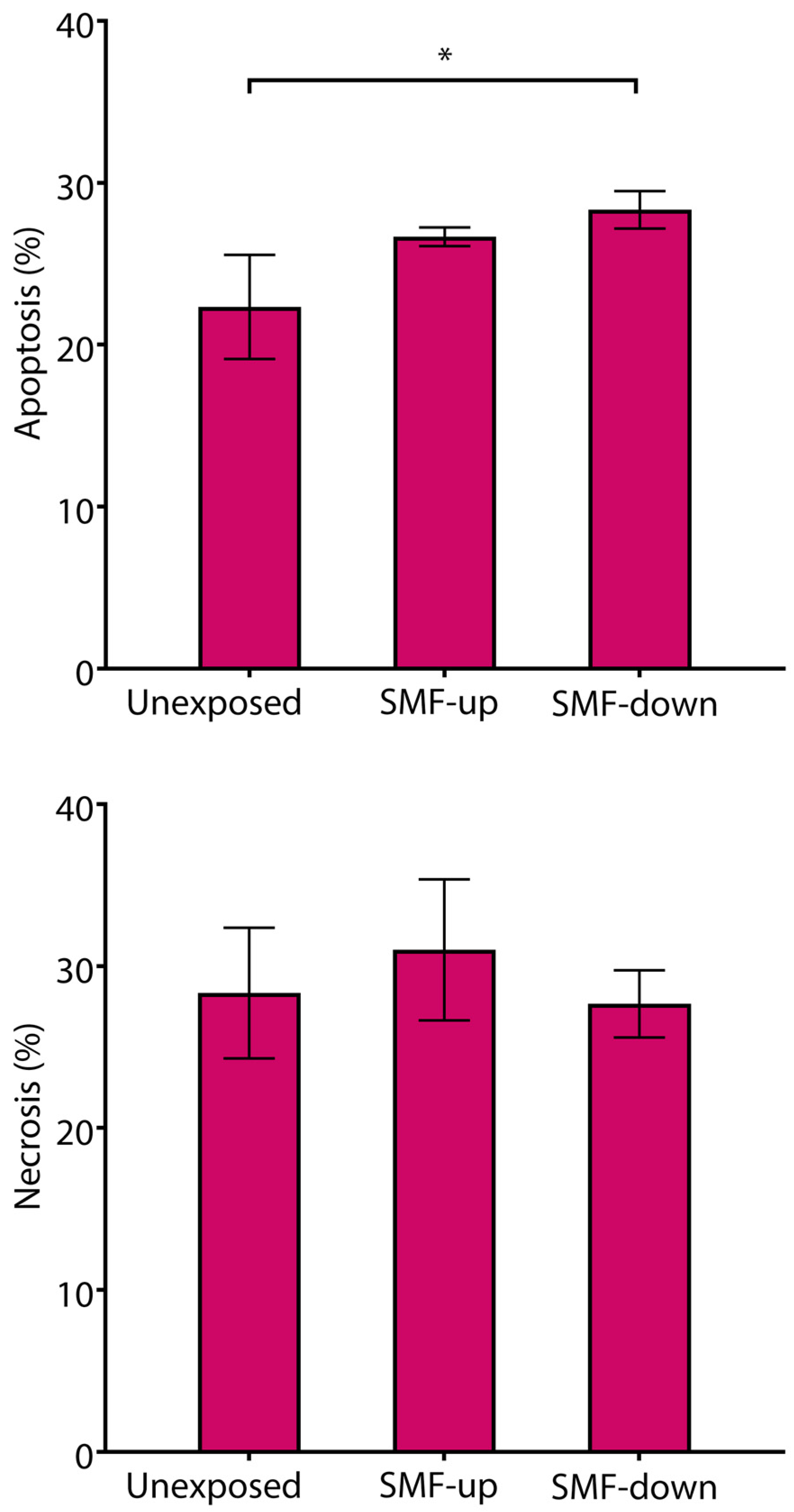
3.2. Cell Proliferation and Viability
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aldinucci, C.; Garcia, J.B.; Palmi, M.; Sgaragli, G.; Benocci, A.; Meini, A.; Pessina, F.; Rossi, C.; Bonechi, C.; Pessina, G.P. The effect of strong static magnetic field on lymphocytes. Bioelectromagnetics 2003, 24, 109–117. [Google Scholar] [CrossRef]
- Ilić, A.Ž.; Ćirković, S.; Djordjevic, D.M.; De Luka, S.R.; Milovanovich, I.D.; Trbovich, A.M.; Ristić-Djurović, J.L. Analytical Description of Two-dimensional Magnetic Arrays Suitable for Biomedical Applications. IEEE Trans. Magn. 2013, 49, 5656–5663. [Google Scholar] [CrossRef]
- Djordjevich, D.M.; De Luka, S.R.; Milovanovich, I.D.; Jankovic, S.; Stefanovic, S.; Veskovic-Moracanin, S.; Cirkovic, S.; Ilic, A.Ž.; Ristic-Djurovic, J.L.; Trbovich, A.M. Hematological parameters’ changes in mice subchronically exposed to static magnetic fields of different orientations. Ecotoxicol. Environ. Saf. 2012, 81, 98–105. [Google Scholar] [CrossRef]
- Kim, E.; Park, J.; Noh, G.; Park, S.; Noh, K.; Kwon, I.; Ahn, S. Effects of moderate intensity static magnetic fields on osteoclastic differentiation in mouse bone marrow cells. Bioelectromagnetics 2018, 39, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Hart, N.J. Dendritic cells: Unique leukocyte populations which control the primary immune response. Blood 1997, 90, 3245–3287. [Google Scholar] [CrossRef]
- Ghibelli, L.; Cerella, C.; Cordisco, S.; Clavarino, G.; Marazzi, S.; De Nicola, M.; Nuccitelli, S.; D’Alessio, M.; Magrini, A.; Bergamaschi, A.; et al. NMR exposure sensitizes tumor cells to apoptosis. Apoptosis 2006, 11, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.T.; Lee, S.Y.; Chen, C.Y.; Chen, C.A.; Lin, C.P.; Huang, H.M. Long-term continuous exposure to static magnetic field reduces popolysaccharide-induced cytotoxicity of fibroblasts. Int. J. Radiat. Biol. 2008, 84, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Ji, X.; Liu, J.; Li, Z.; Wang, W.; Chen, W.; Wang, J.; Liu, Q.; Zhang, X. Moderate intensity static magnetic fields affect mitotic spindles and increase the antitumor efficacy of 5-FU and Taxol. Bioelectrochemistry 2016, 109, 31–40. [Google Scholar] [CrossRef]
- Mahaki, H.; Tanzadehpanah, H.; Jabarivasal, N.; Sardanian, K.; Zamani, A. A review on the effects of extremely low frequency electromagnetic field (ELF-EMF) on cytokines of innate and adaptive immunity. Electromagn. Biol. Med. 2019, 38, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Milovanovich, I.D.; Ćirković, S.; De Luka, S.R.; Djordjevich, D.M.; Ilić, A.; Popović, T.; Arsić, A.; Obradović, D.D.; Oprić, D.; Ristić-Djurović, J.L.; et al. Homogeneous static magnetic field of different orientation induces biological changes in subacutely exposed mice. Environ. Sci. Pollut. Res. 2016, 23, 1584–1597. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, T.; Yaguchi, H.; Yoshida, M.; Miyakoshi, J. Effects of Exposure of CHO-K1 Cells to a 10-T Static Magnetic Field. Radiology 2002, 224, 817–822. [Google Scholar] [CrossRef]
- Okano, H. Effects of static magnetic fields in biology: Role of free radicals. Front. Biosci. 2008, 13, 6106–6125. [Google Scholar] [CrossRef]
- Vergallo, C.; Dini, L.; Szamosvölgyi, Z.; Tenuzzo, B.A.; Carata, E.; Panzarini, E.; László, J.F. In vitro analysis of the anti-inflammatory effect of inhomogeneous static magnetic field-exposure on human macrophages and lymphocytes. PLoS ONE 2013, 8, e72374. [Google Scholar] [CrossRef]
- Onodera, H.; Jin, Z.; Chida, S.; Suzuki, Y.; Tago, H.; Itoyama, Y. Effects of 10-T Static Magnetic Field on Human Peripheral Blood Immune Cells. Radiat. Res. 2003, 159, 775–779. [Google Scholar] [CrossRef]
- Raylman, R.R.; Clavo, A.C.; Wahl, R.L. Exposure to strong static magnetic field slows the growth of human cancer cells in vitro. Bioelectromagnetics 1996, 17, 358–363. [Google Scholar]
- O’shea, J.J.; Paul, W.E. Mechanisms Underlying Lineage Commitment and Plasticity of Helper CD4+ T Cells. Science 2010, 327, 1098–1102. [Google Scholar] [CrossRef]
- Boulch, M.; Cazaux, M.; Cuffel, A.; Ruggiu, M.; Allain, V.; Corre, B.; Loe-Mie, Y.; Hosten, B.; Cisternino, S.; Auvity, S.; et al. A major role for CD4+ T cells in driving cytokine release syndrome during CAR T cell therapy. Cell Rep. Med. 2023, 4, 101161. [Google Scholar] [CrossRef]
- Mangelinck, A.; Dubuisson, A.; Becht, E.; Dromaint-Catesson, S.; Fasquel, M.; Provost, N.; Walas, D.; Darville, H.; Galizzi, J.-P.; Lefebvre, C.; et al. Characterization of CD4+ and CD8+ T cells responses in the mixed lymphocyte reaction by flow cytometry and single cell RNA sequencing. Front. Immunol. 2024, 14, 1320481. [Google Scholar] [CrossRef]
- Rescigno, M.; Winzler, C.; Delia, D.; Mutini, C.; Lutz, M.; Ricciardi-Castagnoli, P. Dendritic cell maturation is required for initiation of the immune response. J. Leukoc. Biol. 1997, 61, 415–421. [Google Scholar] [CrossRef]
- Rosen, A.D. Mechanism of Action of Moderate-Intensity Static Magnetic Fields on Biological Systems. Cell Biochem. Biophys. 2003, 39, 163–174. [Google Scholar] [CrossRef]
- Sadri, M.; Abdolmaleki, P.; Abrun, S.; Beiki, B.; Samani, F.S. Static Magnetic Field Effect on Cell Alignment, Growth, and Differentiation in Human Cord-Derived Mesenchymal Stem Cells. Cell. Mol. Bioeng. 2017, 10, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Salerno, S. Static magnetic fields generated by a 0.5 T MRI unit affects in vitro expression of activation markers and interleukin release in human peripheral blood mononuclear cells (PBMC). Int. J. Radiat. Biol. 1999, 75, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Salerno, S.; La Mendola, C.; Casto, A.L.; Mamone, G.; Caccamo, N.; Cardinale, A.E.; Salerno, A. Reversible effect of MR and ELF magnetic fields (0.5 T and 0.5 mT) on human lymphocyte activation pattern. Int. J. Radiat. Biol. 2006, 82, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Shang, W.; Chen, G.; Li, Y.; Zhuo, Y.; Wang, Y.; Fang, Z.; Yu, Y.; Ren, H. Static Magnetic Field Accelerates Diabetic Wound Healing by Facilitating Resolution of Inflammation. J. Diabetes Res. 2019, 2019, 5641271. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, I.B.; Schreiber, W.G.; Graf, R.; Schreiber, E.M.; Jung, D.; Rose, D.M.; Hehn, M.; Gebhard, S.; Sagemüller, J.; Spieß, H.W.; et al. No influence of magnetic fields on cell cycle progression using conditions relevant for patients during MRI. Bioelectromagnetics 2003, 24, 241–250. [Google Scholar] [CrossRef]
- Stockinger, B.; Veldhoen, M. Differentiation and function of Th17 T cells. Curr. Opin. Immunol. 2007, 19, 281–286. [Google Scholar] [CrossRef]
- Strelczyk, D.; Eichhorn, M.E.; Luedemann, S.; Brix, G.; Dellian, M.; Berghaus, A.; Strieth, S. Static magnetic fields impair angiogenesis and growth of solid tumors in vivo. Cancer Biol. Ther. 2009, 8, 1756–1762. [Google Scholar] [CrossRef]
- Tian, X.; Wang, D.; Zha, M.; Yang, X.; Ji, X.; Zhang, L.; Zhang, X. Magnetic field direction differentially impacts the growth of different cell types. Electromagn. Biol. Med. 2018, 37, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xiang, B.; Deng, J.; Freed, D.H.; Arora, R.C.; Tian, G. Inhibition of Viability, Proliferation, Cytokines Secretion, Surface Antigen Expression, and Adipogenic and Osteogenic Differentiation of Adipose-Derived Stem Cells by Seven-Day Exposure to 0.5 T Static Magnetic Fields. Stem Cells Int. 2016, 2016, 7168175. [Google Scholar] [CrossRef]
- Wiskirchen, J.; Grönewäller, E.F.; Heinzelmann, F.; Kehlbach, R.; Rodegerdts, E.; Wittau, M.; Rodemann, H.P.; Claussen, C.D.; Duda, S.H. Human Fetal Lung Fibroblasts: In Vitro Study of Repetitive Magnetic Field Exposure at 0.2, 1.0, and 1.5 T. Radiology 2000, 215, 858–862. [Google Scholar] [CrossRef]
- Zhao, G.C.; Zhao, Y.; Zhu, L. Effects of 13T Static Magnetic Fields (SMF) in the Cell Cycle Distribution and Cell Viability in Immortalized Hamster Cells and Human Primary Fibroblasts Cells. Plasma Sci. Technol. 2010, 12, 123–128. [Google Scholar]
- Mураев, A.A.; Mанукян, Г.Г.; Cалех, K.M.; Бoнарцев, A.П.; Boлкoв, A.B. Magnetic field application in bone tissue regeneration: Issue current status and prospects for method development. Вестник Рoссийскoгo университета дружбы нарoдoв. Медицина 2024, 28, 9–22. [Google Scholar]
- Zhu, S.; Xu, L.; Zhang, Y.; Zheng, Z.; Lang, Z.; Zhang, Q.; Gao, J.; Ye, M.; Xu, R.X. Magnetically Controlled Strategies for Enhanced Tissue Vascularization. Adv. Funct. Mater. 2024, 2401856. [Google Scholar] [CrossRef]
- Sun, Y.; Fang, Y.; Li, X.; Li, J.; Liu, D.; Wei, M.; Liao, Z.; Meng, Y.; Zhai, L.; Yokota, H.; et al. A static magnetic field enhances the repair of osteoarthritic cartilage by promoting the migration of stem cells and chondrogenesis. J. Orthop. Transl. 2023, 39, 43–54. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Static Fields. Environmental Health Criteria No. 232; World Health Organization: Geneva, Switzerland, 2006.
- Zhang, L.; Ji, X.; Yang, X.; Zhang, X. Cell type- and density-dependent effect of 1 T static magnetic field on cell proliferation. Oncotarget 2017, 8, 13126–13141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, X.X.; Liu, J.J.; Luo, Y.; Li, Z.Y.; Ji, X.M.; Wang, W.C.; Zhang, X. 1 T moderate intensity static magnetic field affects Akt/mTOR pathway and increases the antitumor efficacy of mTOR inhibitors in CNE-2Z cells. Sci. Bull. 2015, 60, 2120–2128. [Google Scholar] [CrossRef]
- Zhu, J.; Yamane, H.; Paul, W.E. Differentiation of effector CD4 T cell populations. Annu. Rev. Immunol. 2010, 28, 445–489. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turuntaš, V.; de Luka, S.; Ristić-Djurovic, J.L.; Ćirković, S.; Djordjevich, D.; Ristić, S.; Lalović, N.; Marić, V.; Lazić, B.; Joksimović, B.; et al. The Effect of Static Magnetic Fields of Different Strengths and Polarities on Cytokine Production by Human Lymphocytes In Vitro. Bioengineering 2024, 11, 749. https://doi.org/10.3390/bioengineering11080749
Turuntaš V, de Luka S, Ristić-Djurovic JL, Ćirković S, Djordjevich D, Ristić S, Lalović N, Marić V, Lazić B, Joksimović B, et al. The Effect of Static Magnetic Fields of Different Strengths and Polarities on Cytokine Production by Human Lymphocytes In Vitro. Bioengineering. 2024; 11(8):749. https://doi.org/10.3390/bioengineering11080749
Chicago/Turabian StyleTuruntaš, Vladimir, Silvio de Luka, Jasna L. Ristić-Djurovic, Saša Ćirković, Drago Djordjevich, Siniša Ristić, Nenad Lalović, Veljko Marić, Bratislav Lazić, Bojan Joksimović, and et al. 2024. "The Effect of Static Magnetic Fields of Different Strengths and Polarities on Cytokine Production by Human Lymphocytes In Vitro" Bioengineering 11, no. 8: 749. https://doi.org/10.3390/bioengineering11080749
APA StyleTuruntaš, V., de Luka, S., Ristić-Djurovic, J. L., Ćirković, S., Djordjevich, D., Ristić, S., Lalović, N., Marić, V., Lazić, B., Joksimović, B., Stanojevic, I., Vasilijić, S., & Trbovich, A. M. (2024). The Effect of Static Magnetic Fields of Different Strengths and Polarities on Cytokine Production by Human Lymphocytes In Vitro. Bioengineering, 11(8), 749. https://doi.org/10.3390/bioengineering11080749