
Development and Verification of a Novel Three-Dimensional Aqueous Outflow Model for High-Throughput Drug Screening


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
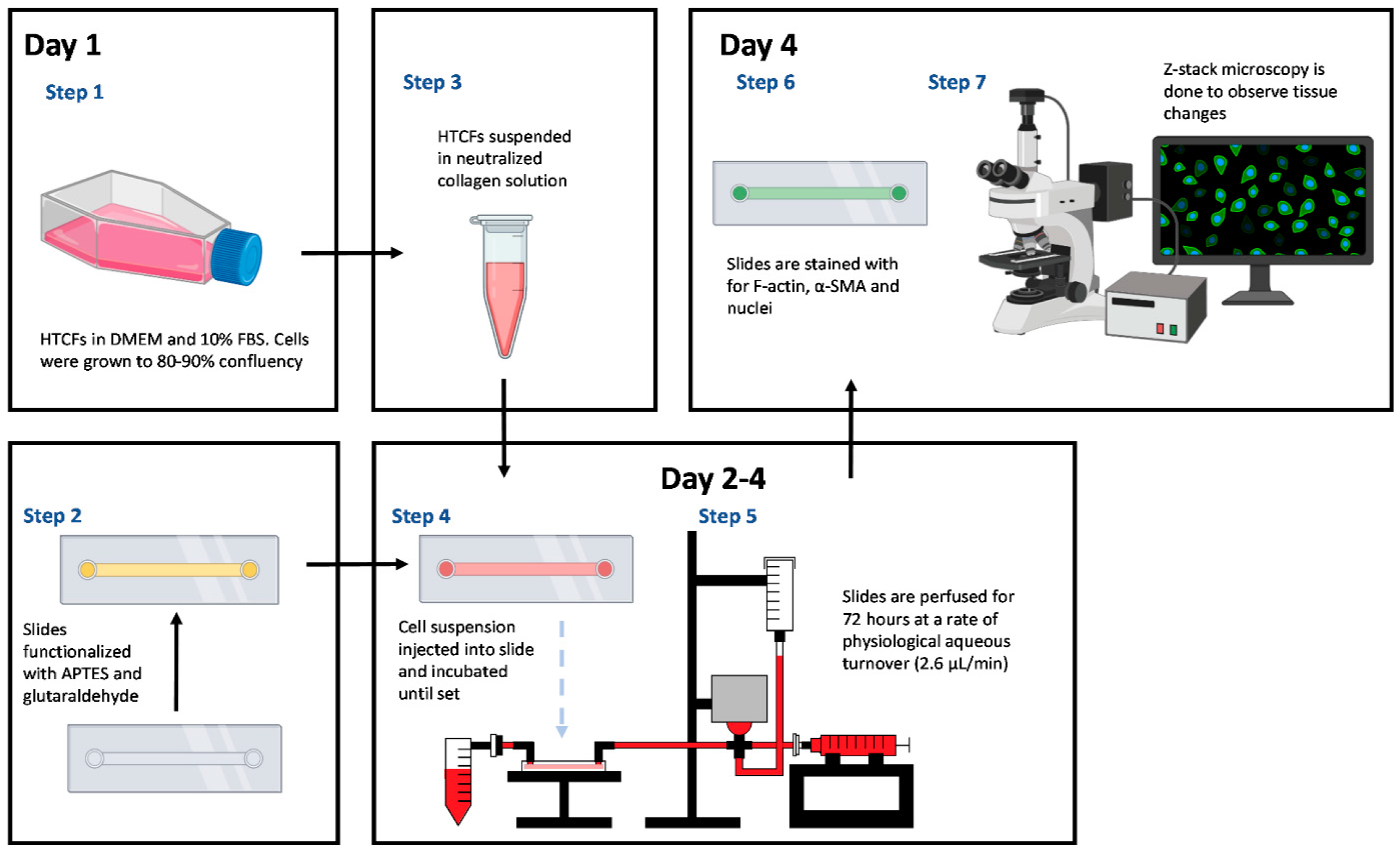
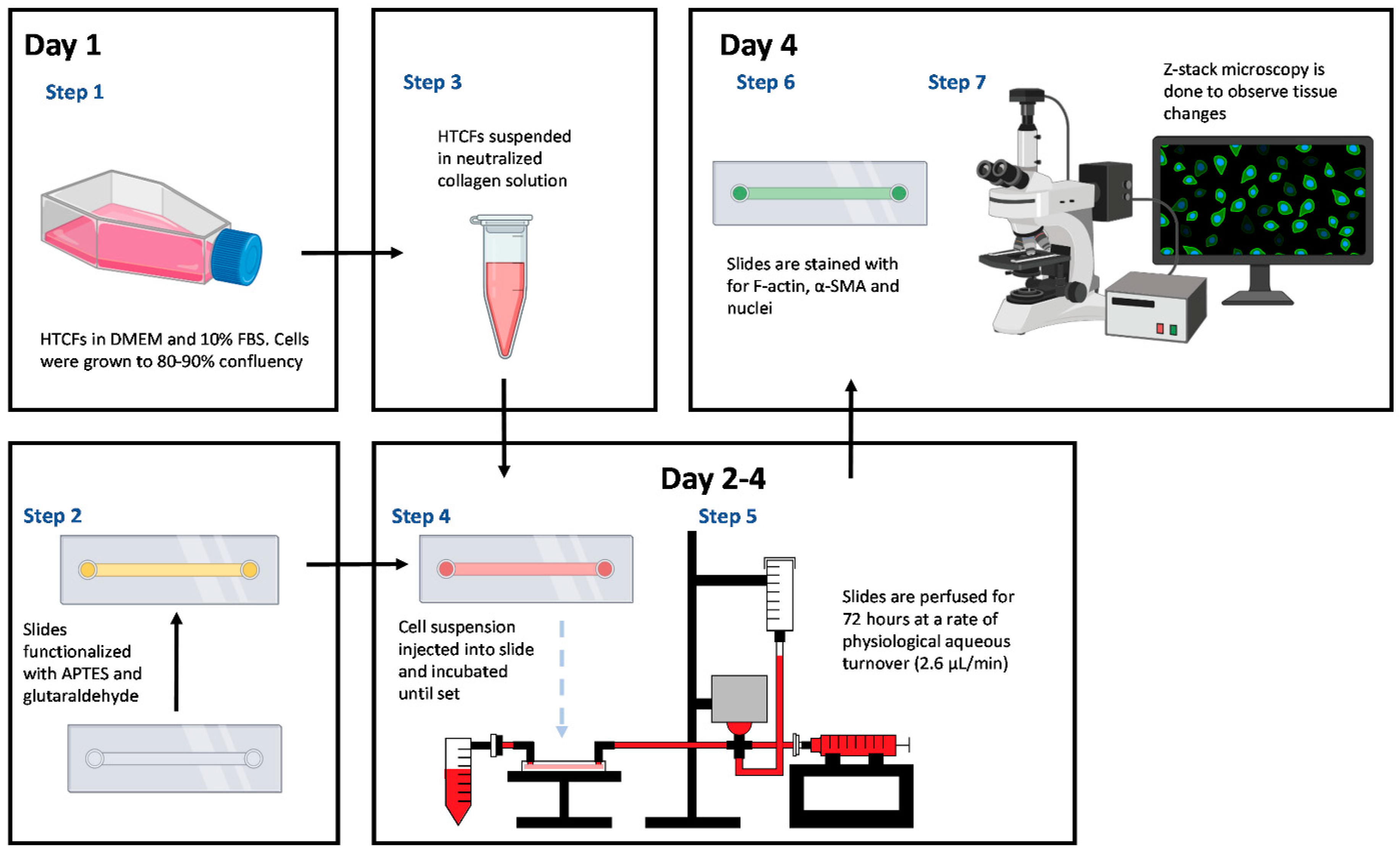
2.1. Primary Human Tenon’s Capsule Fibroblast (HTCF) Procurement and Culture
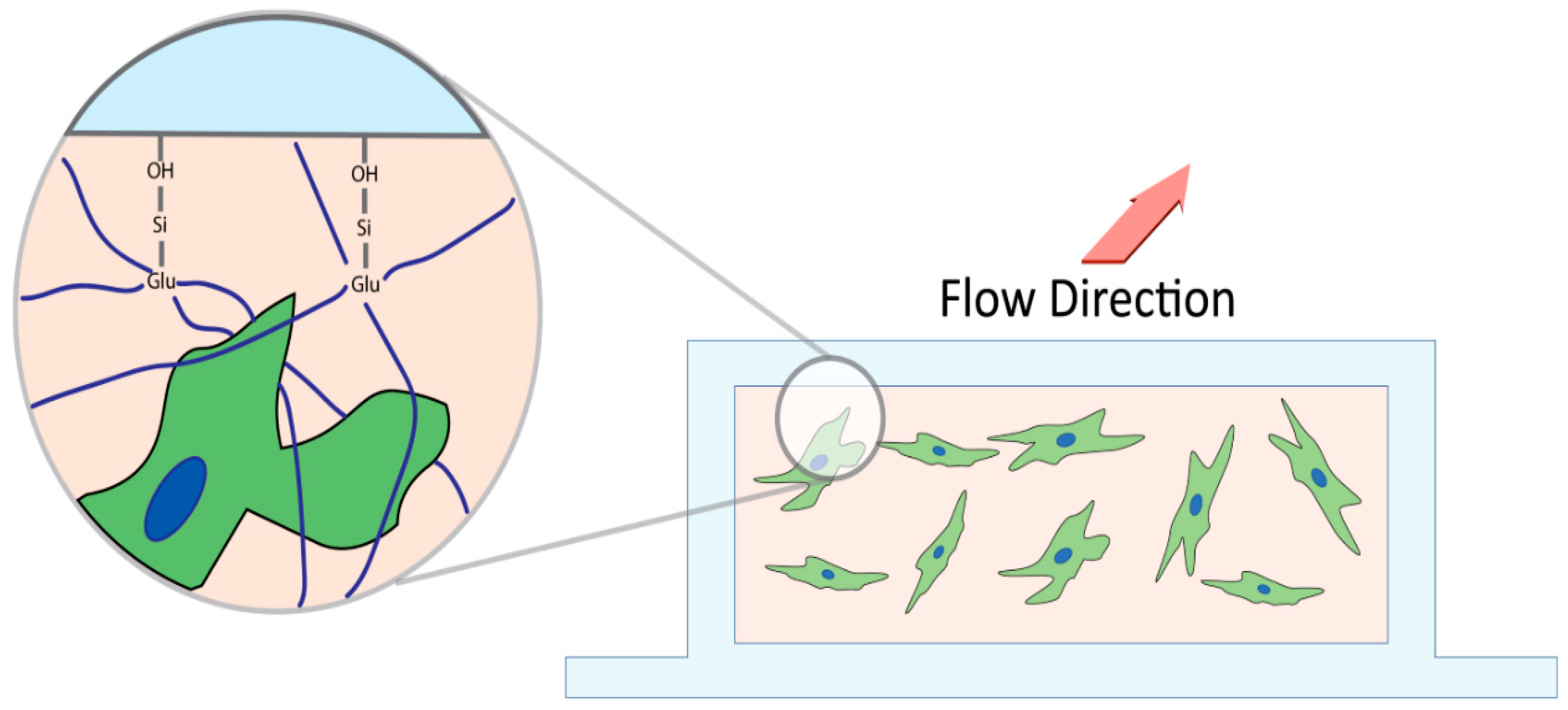
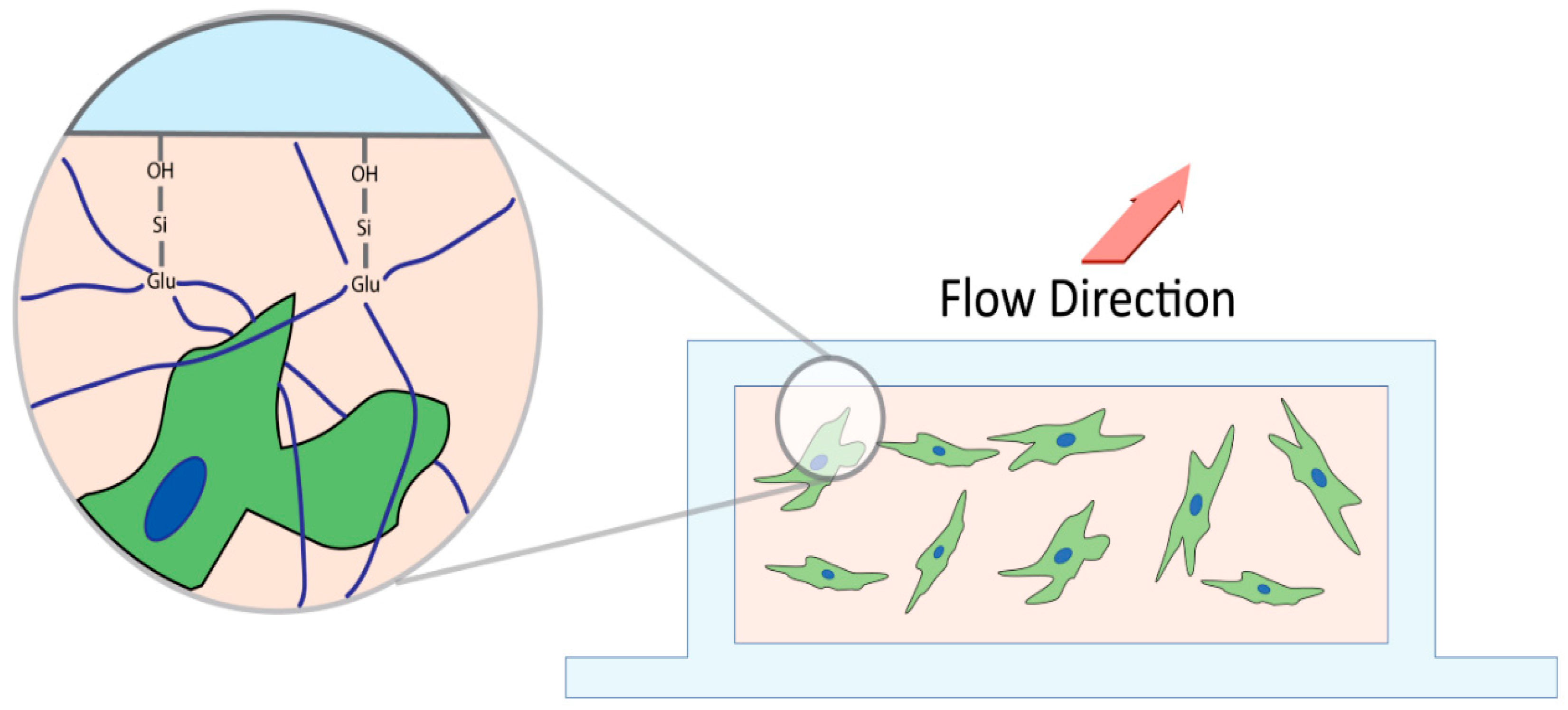
2.2. Microfluidic Slide Chamber Preparation
2.3. HTCF Preparation for Hydrogel Formation
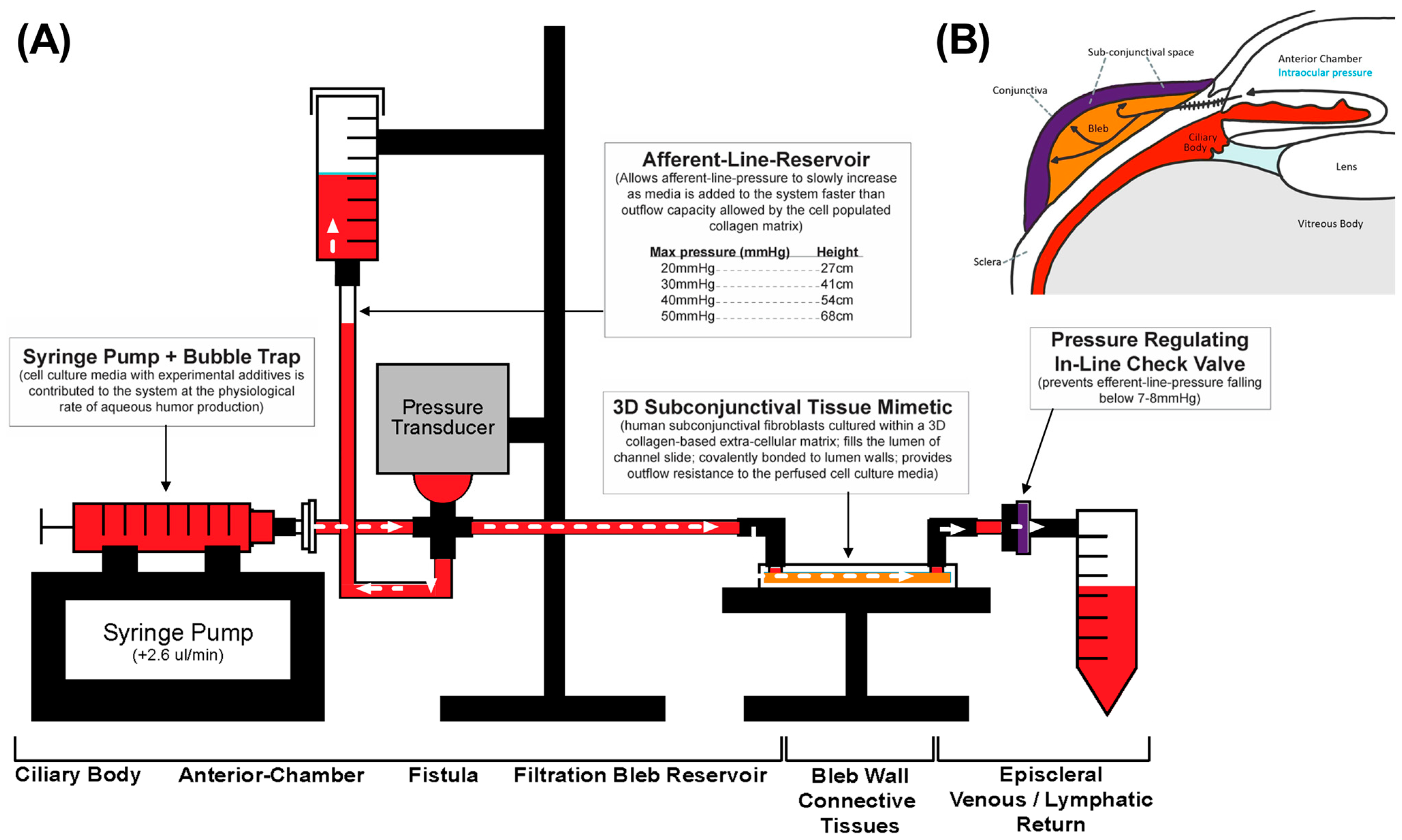
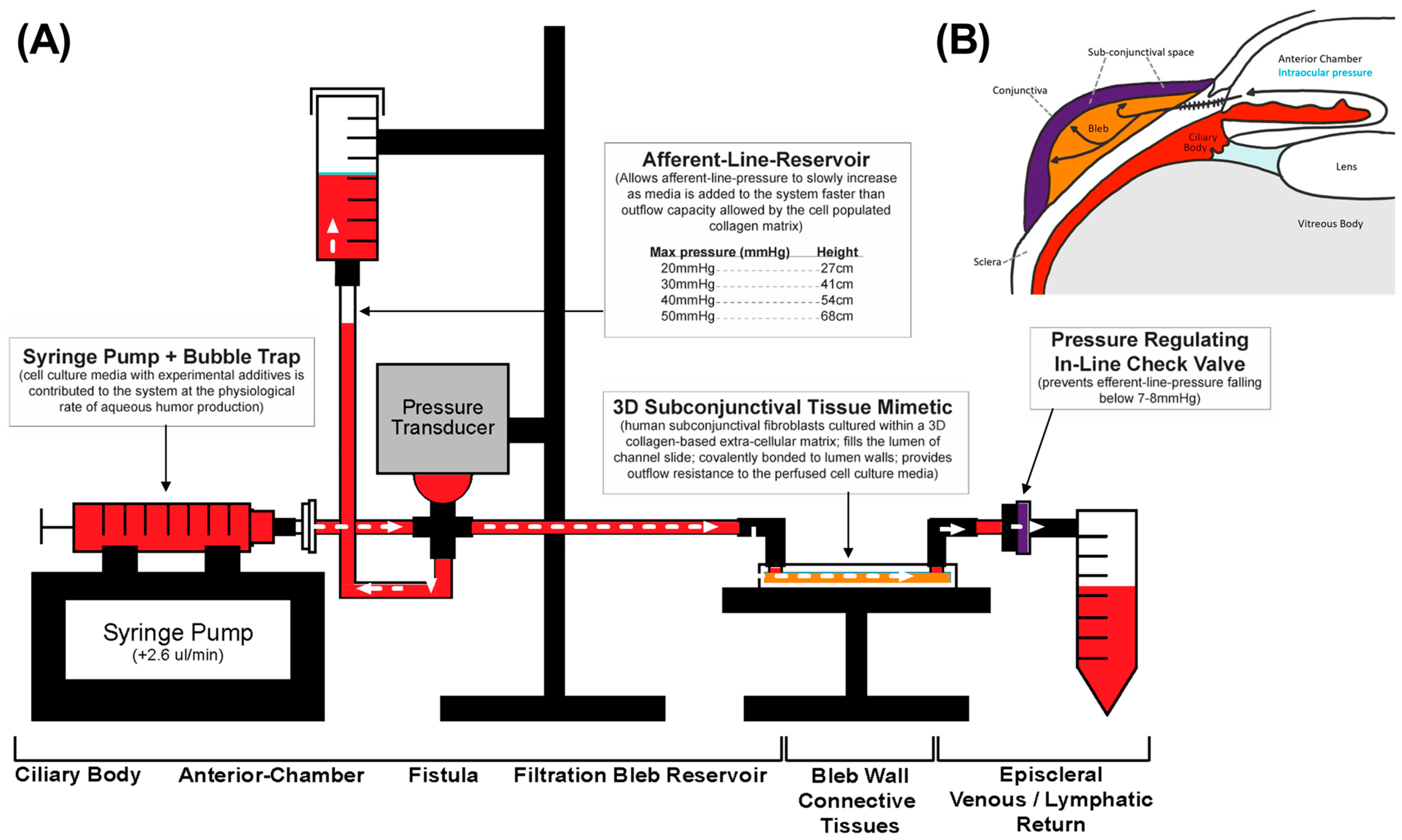
2.4. Perfusion Track Construction and Experimental Setup
2.5. Afferent Line Pressure Analysis
2.6. Tissue-Mimetic Fixation and Histological Staining
2.7. Confocal Immuno-Fluorescent Microscopy
2.8. Western Blot
3. Results
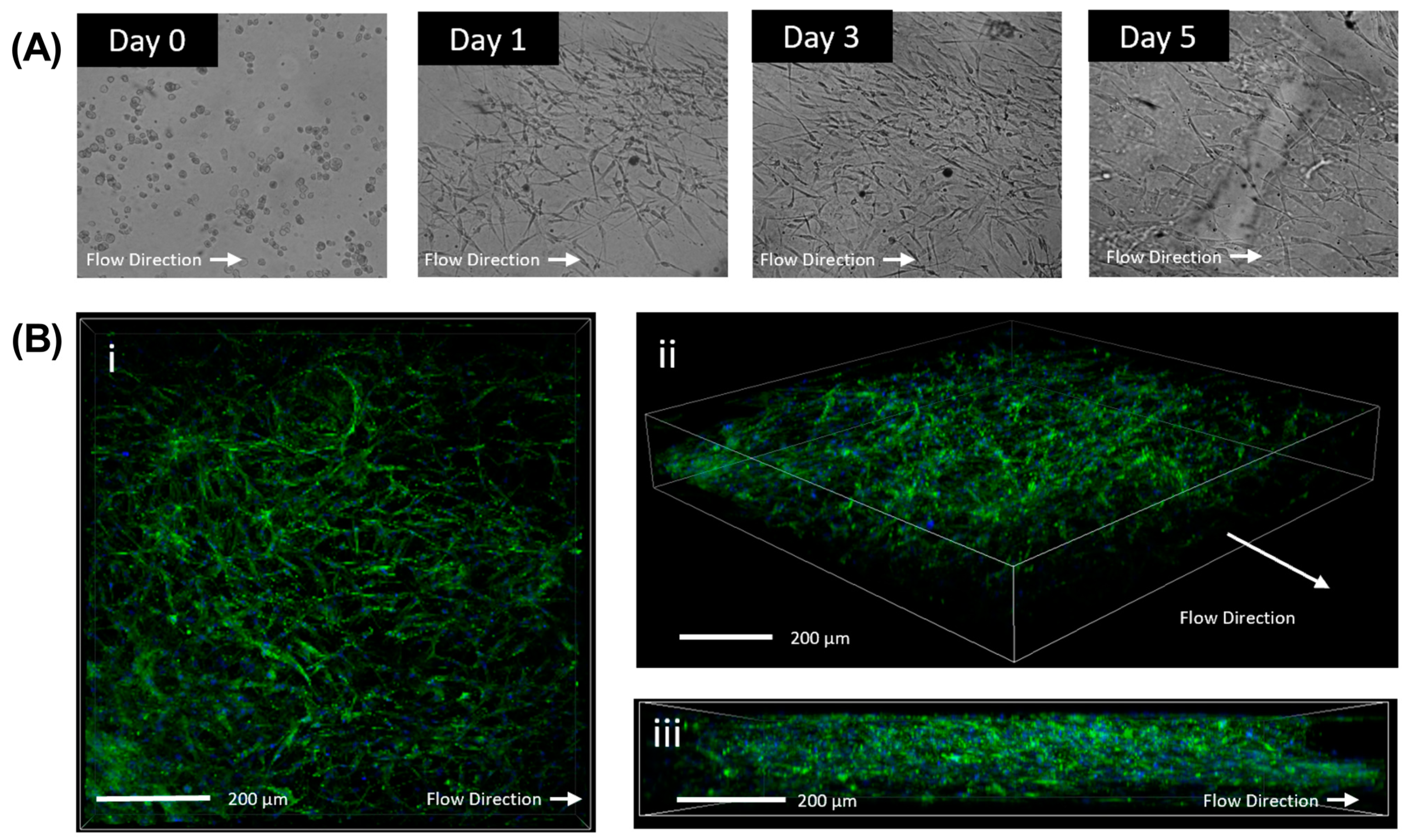
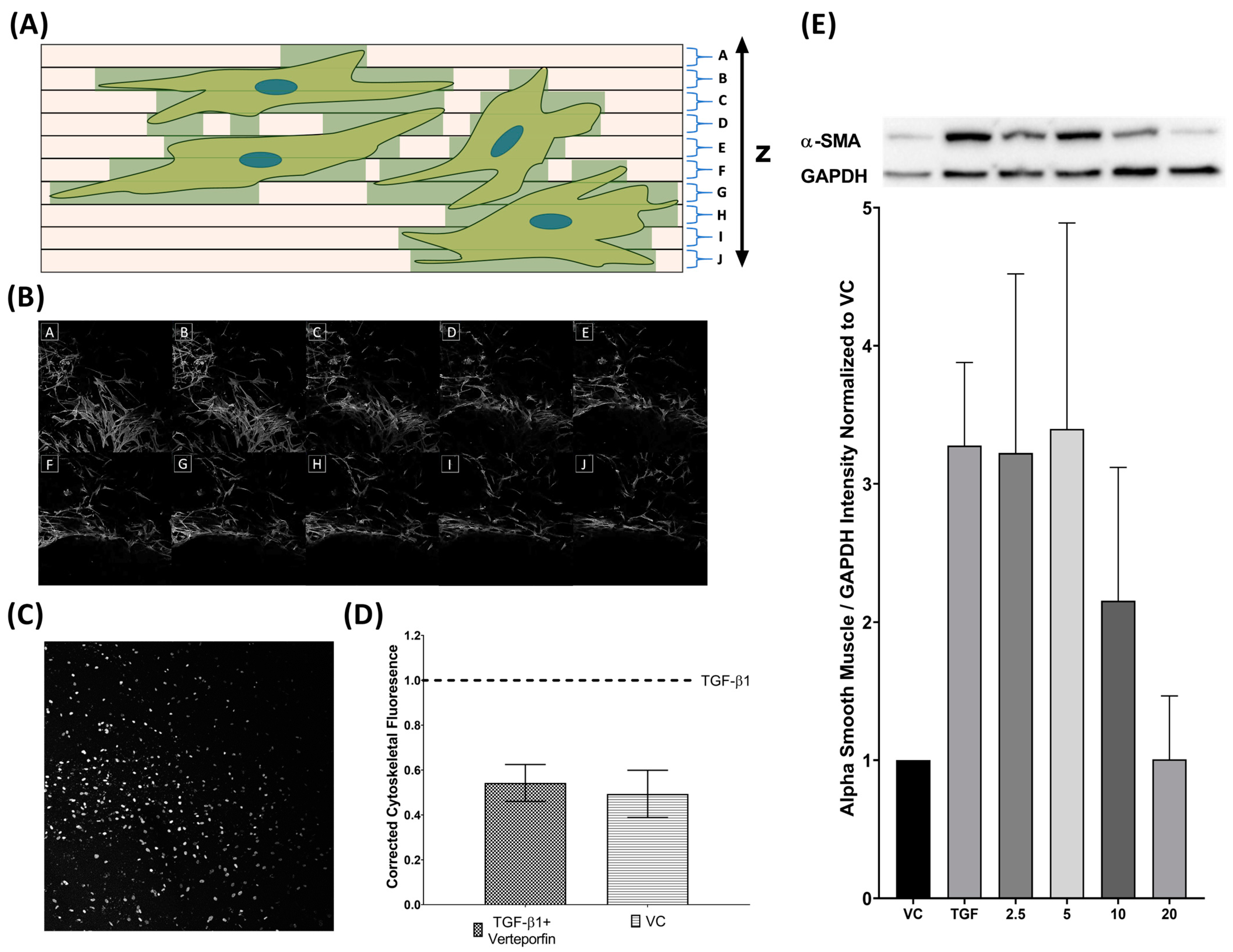
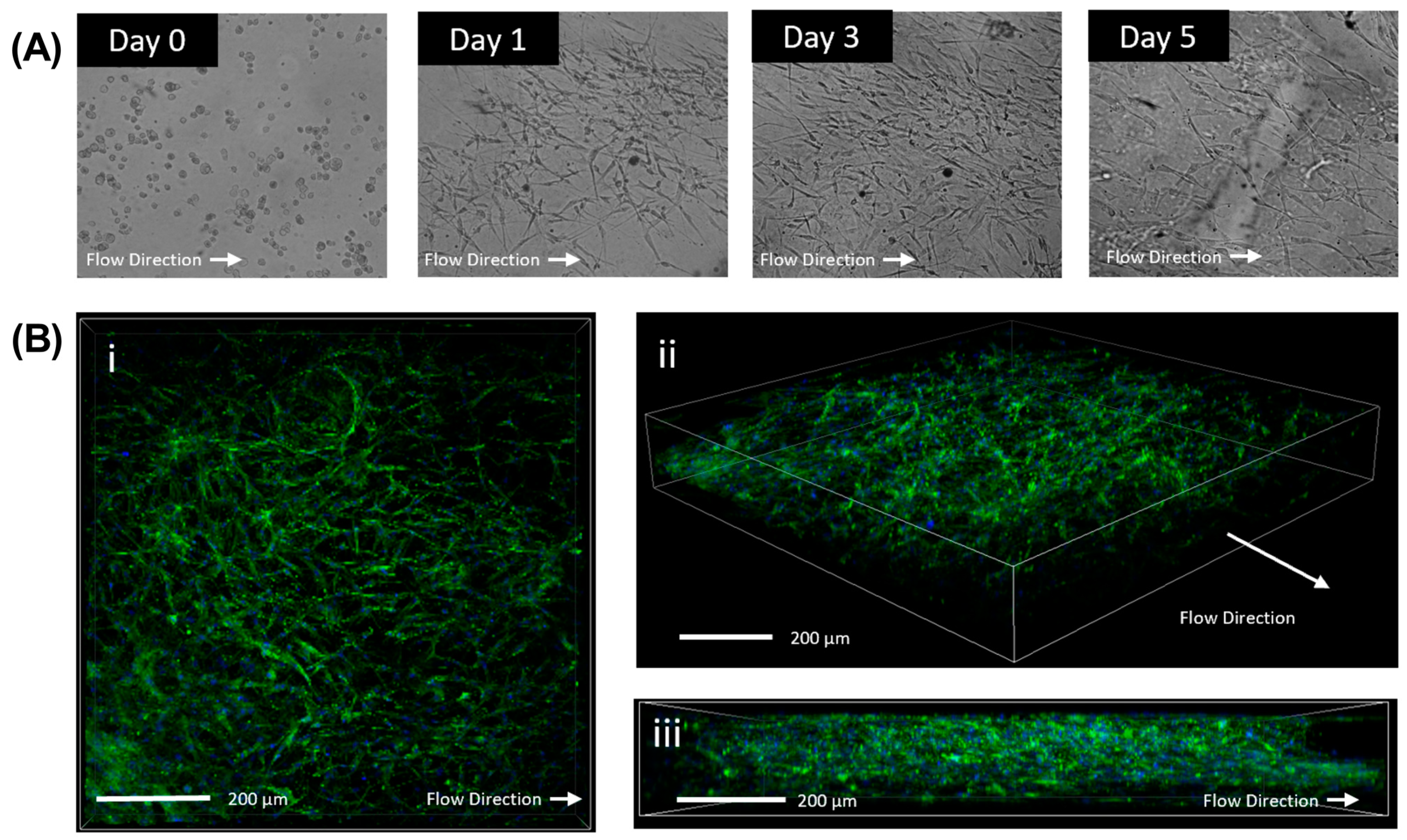
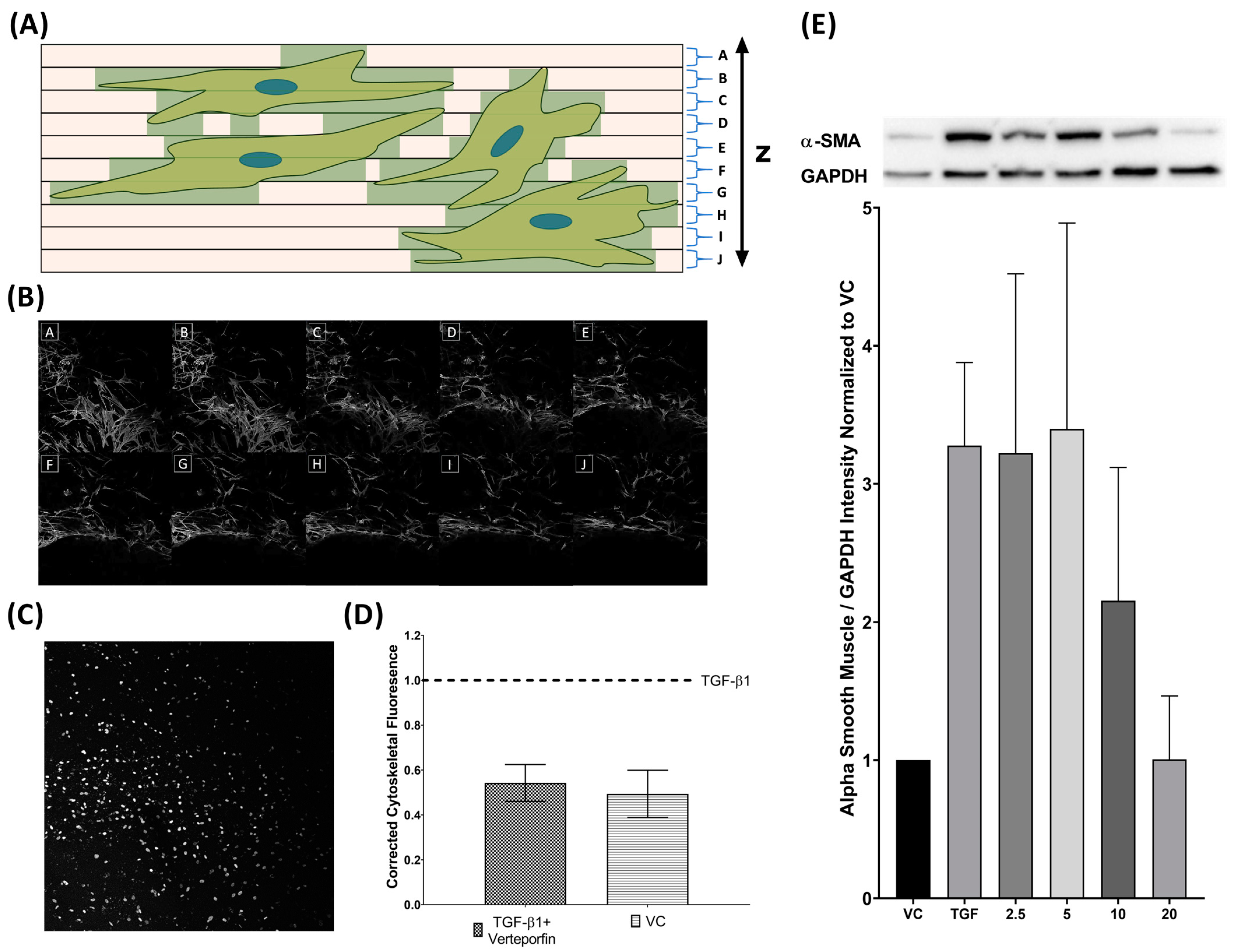
3.1. Analysis of Three-Dimensional Tissue Mimetic Structure
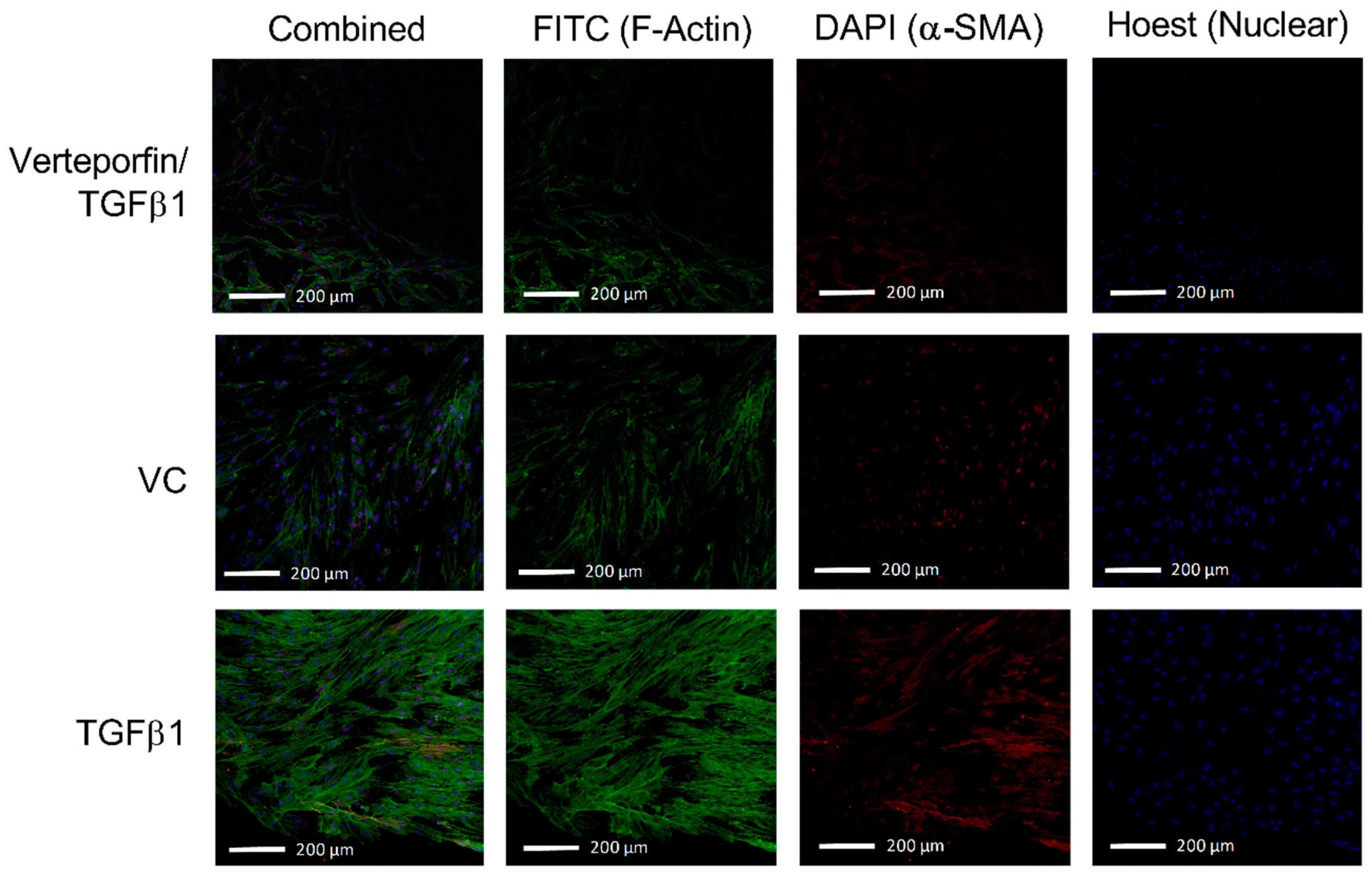
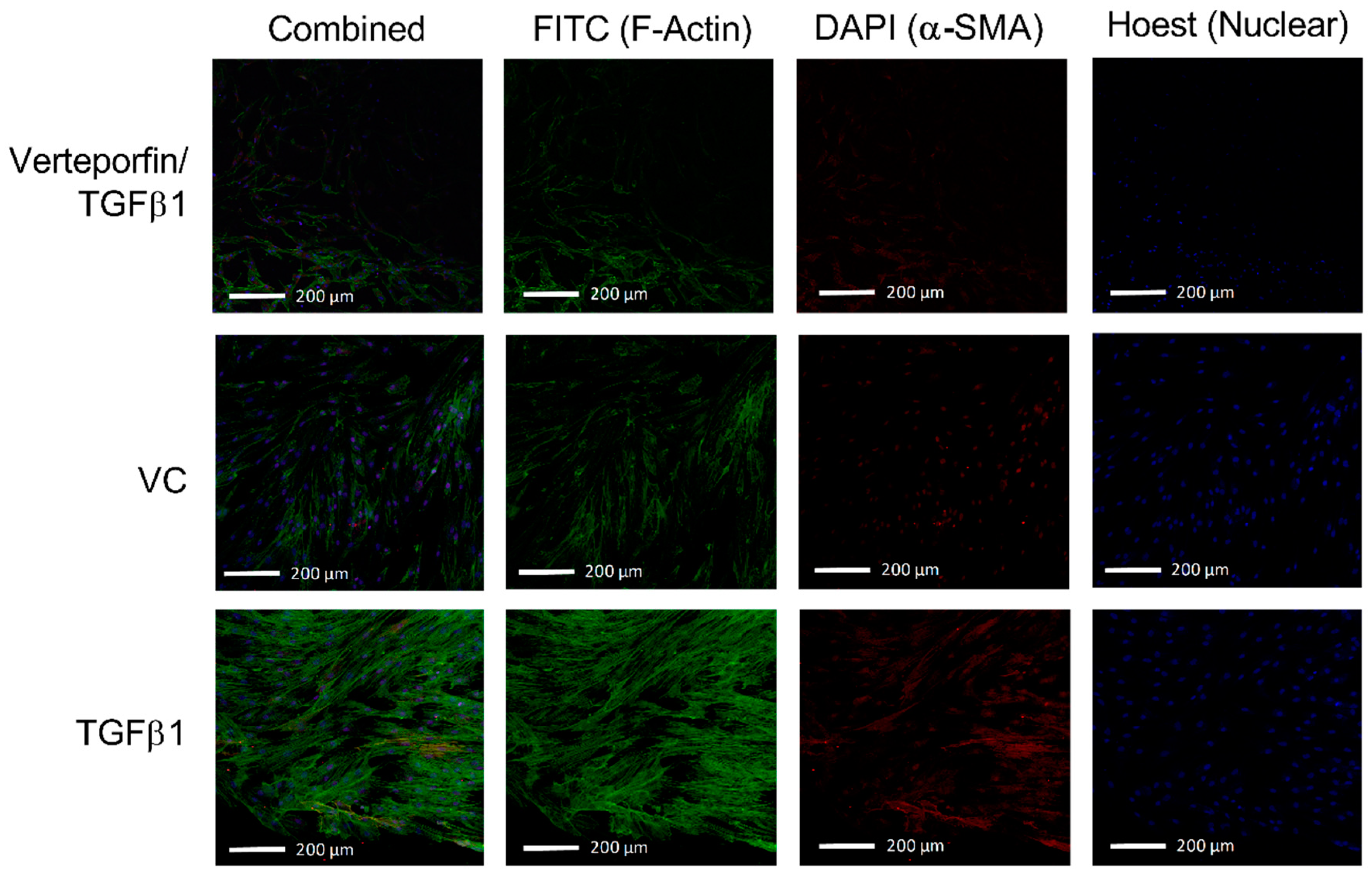
3.2. Morphological Changes Due to TGFβ1 and Verteporfin Co-Treatment
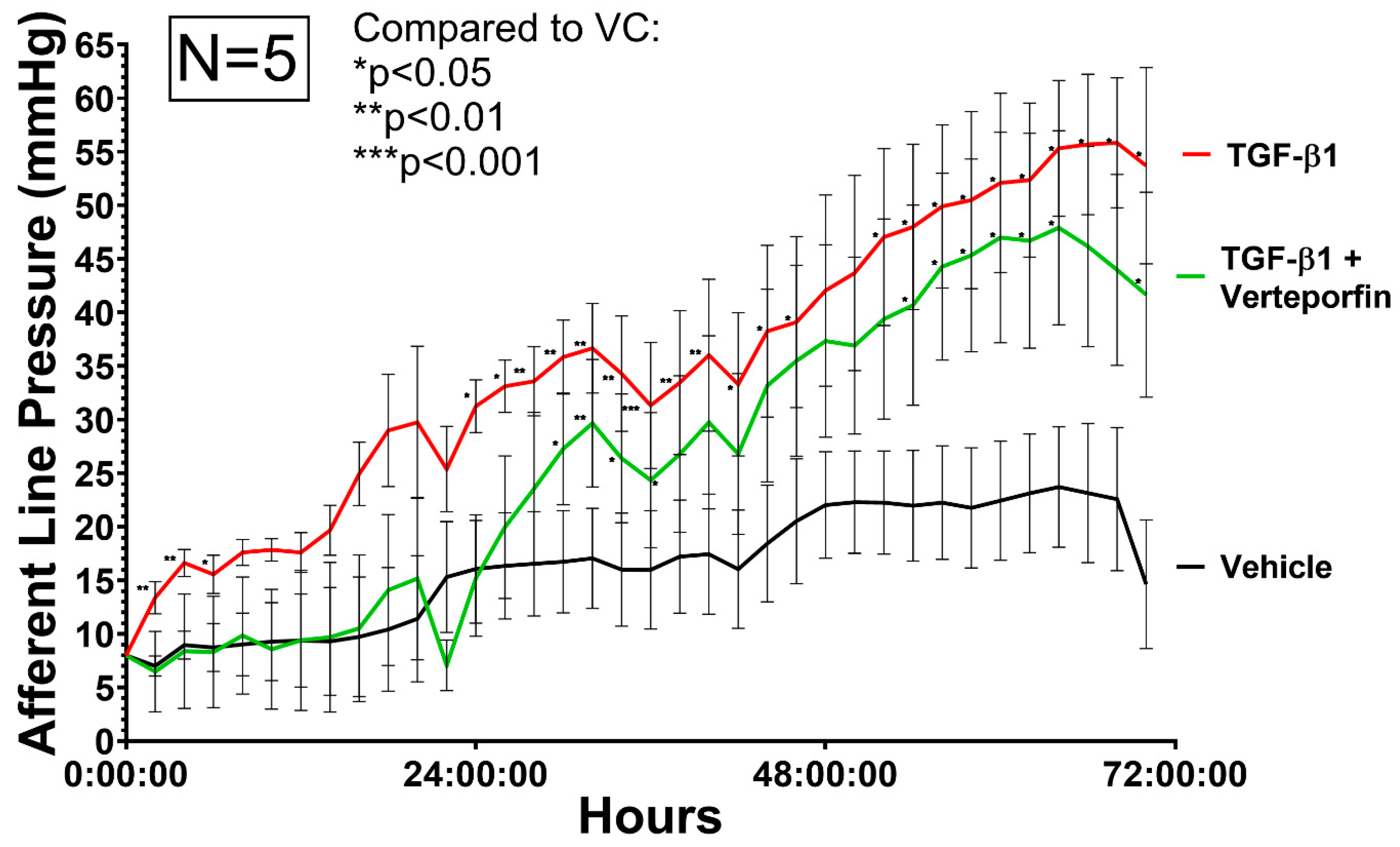
3.3. Afferent Line Pressure Changes
3.4. Semi-Quantification of TGFβ1 Mediated Phenotypic Changes
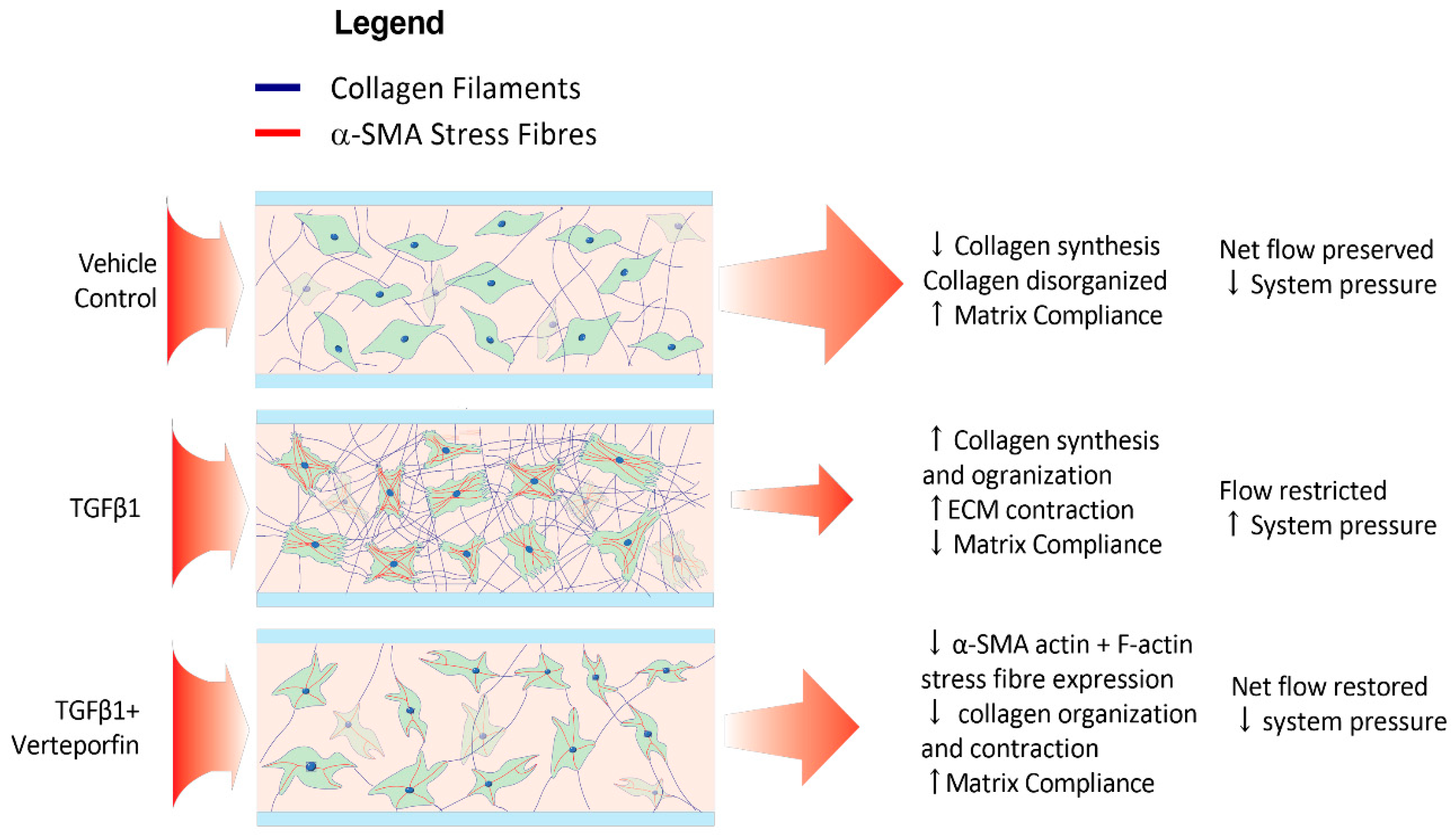
4. Discussion
4.1. Development of a Three-Dimensional Flow Model
4.2. Phenotypic Effects of TGF β1 Are Reflected within the Model
4.3. TGFβ1 Mediated SMAD2/3 Inhibition Leads to Attenuation of Fibroblast Transdifferentiation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, W.; Chen, S.; Gao, X.; Yang, M.; Zhang, J.; Li, X.; Wang, W.; Zhou, M.; Zhang, X.; Zhang, X. Inflammation-Related Cytokines of Aqueous Humor in Acute Primary Angle-Closure Eyes. Investig. Ophthalmol. Vis. Sci. 2014, 55, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, R.C.; Li, J.; Chan, W.F.A.; Tripathi, B.J. Aqueous Humor in Glaucomatous Eyes Contains an Increased Level of TGF-Β2. Exp. Eye Res. 1994, 59, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Tran, M.N.; Medveczki, T.; Besztercei, B.; Torok, G.; Szabo, A.J.; Gasull, X.; Kovacs, I.; Fekete, A.; Hodrea, J. Sigma-1 Receptor Activation Is Protective against TGFβ2-Induced Extracellular Matrix Changes in Human Trabecular Meshwork Cells. Life 2023, 13, 1581. [Google Scholar] [CrossRef] [PubMed]
- Prendes, M.A.; Harris, A.; Wirostko, B.M.; Gerber, A.L.; Siesky, B. The Role of Transforming Growth Factor β in Glaucoma and the Therapeutic Implications. Br. J. Ophthalmol. 2013, 97, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Shepard, A.R.; Cameron Millar, J.; Pang, I.H.; Jacobson, N.; Wang, W.H.; Clark, A.F. Adenoviral Gene Transfer of Active Human Transforming Growth Factor-Β2 Elevates Intraocular Pressure and Reduces Outflow Facility in Rodent Eyes. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2067–2076. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Fang, L.; Yan, C.; Wei, J.; Song, D.; Xu, C.; Luo, Y.; Fan, Y.; Guo, L.; Sun, H.; et al. MicroRNA-210-3p Mediates Trabecular Meshwork Extracellular Matrix Accumulation and Ocular Hypertension—Implication for Novel Glaucoma Therapy. Exp. Eye Res. 2023, 227, 109350. [Google Scholar] [CrossRef]
- Robertson, J.V.; Siwakoti, A.; West-Mays, J.A. Altered Expression of Transforming Growth Factor Beta 1 and Matrix Metalloproteinase-9 Results in Elevated Intraocular Pressure in Mice. Mol. Vis. 2013, 19, 684–695. [Google Scholar] [PubMed]
- Picht, G.; Welge-Luessen, U.; Grehn, F.; Lütjen-Drecoll, E. Transforming Growth Factor Beta 2 Levels in the Aqueous Humor in Different Types of Glaucoma and the Relation to Filtering Bleb Development. Graefes Arch. Clin. Exp. Ophthalmol. 2001, 239, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Desmouliere, A.; Geinoz, A.; Gabbiani, F.; Gabbiani, G. Transforming Growth Factor-Β1 Induces α-Smooth Muscle Actin Expression in Granulation Tissue Myofibroblasts and in Quiescent and Growing Cultured Fibroblasts. J. Cell Biol. 1993, 122, 103–111. [Google Scholar] [CrossRef]
- Lee, S.Y.; Chae, M.K.; Yoon, J.S.; Kim, C.Y. The Effect of CHIR 99021, a Glycogen Synthase Kinase-3β Inhibitor, on Transforming Growth Factor β-Induced Tenon Fibrosis. Investig. Ophthalmol. Vis. Sci. 2021, 62, 25. [Google Scholar] [CrossRef]
- Chen, J.; Li, H.; SundarRaj, N.; Wang, J.H.-C. Alpha-Smooth Muscle Actin Expression Enhances Cell Traction Force. Cell Motil. Cytoskelet. 2007, 64, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Hecker, L.; Jagirdar, R.; Jin, T.; Thannickal, V.J. Reversible Differentiation of Myofibroblasts by MyoD. Exp. Cell Res. 2011, 317, 1914–1921. [Google Scholar] [CrossRef] [PubMed]
- Talele, N.P.; Fradette, J.; Davies, J.E.; Kapus, A.; Hinz, B. Expression of α-Smooth Muscle Actin Determines the Fate of Mesenchymal Stromal Cells. Stem Cell Rep. 2015, 4, 1016–1030. [Google Scholar] [CrossRef]
- Desmoulière, A.; Badid, C.; Bochaton-Piallat, M.L.; Gabbiani, G. Apoptosis during Wound Healing, Fibrocontractive Diseases and Vascular Wall Injury. Int. J. Biochem. Cell Biol. 1997, 29, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Conlon, R.; Saheb, H.; Ahmed, I.I.K. Glaucoma Treatment Trends: A Review. Can. J. Ophthalmol. 2017, 52, 114–124. [Google Scholar] [CrossRef]
- Pillunat, L.E.; Erb, C.; Jünemann, A.G.; Kimmich, F. Micro-Invasive Glaucoma Surgery (MIGS): A Review of Surgical Procedures Using Stents. Clin. Ophthalmol. 2017, 11, 1583–1600. [Google Scholar] [CrossRef]
- Kessing, S.V.; Flesner, P.; Jensen, P.K. Determinants of Bleb Morphology in Minimally Invasive, Clear-Cornea Micropenetrating Glaucoma Surgery with Mitomycin C. J. Glaucoma 2006, 15, 84–90. [Google Scholar] [CrossRef]
- Kankainen, T.; Harju, M. Endophthalmitis and Blebitis Following Deep Sclerectomy and Trabeculectomy with Routine Use of Mitomycin C. Acta Ophthalmol. 2023, 101, 285–292. [Google Scholar] [CrossRef]
- Phulke, S.; Kaushik, S.; Kaur, S.; Pandav, S. Steroid-Induced Glaucoma: An Avoidable Irreversible Blindness. J. Curr. Glaucoma Pract. 2017, 11, 67. [Google Scholar]
- Kanta, J. Collagen Matrix as a Tool in Studying Fibroblastic Cell Behavior. Cell Adhes. Migr. 2015, 9, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B. The Myofibroblast: Paradigm for a Mechanically Active Cell. J. Biomech. 2010, 43, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.L.; Bloomer, S.A.; Chan, E.P.; Gaça, M.D.A.; Georges, P.C.; Sackey, B.; Uemura, M.; Janmey, P.A.; Wells, R.G. Hepatic Stellate Cells Require a Stiff Environment for Myofibroblastic Differentiation. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G110–G118. [Google Scholar] [CrossRef] [PubMed]
- Burgstaller, G.; Gerckens, M.; Eickelberg, O.; Königshoff, M. Decellularized Human Lung Scaffolds as Complex Three-Dimensional Tissue Culture Models to Study Functional Behavior of Fibroblasts. Methods Mol. Biol. 2021, 2299, 447–456. [Google Scholar] [CrossRef]
- Bracken, M.B. Why Animal Studies Are Often Poor Predictors of Human Reactions to Exposure. J. R. Soc. Med. 2009, 102, 120–122. [Google Scholar] [CrossRef]
- von Scheidt, M.; Zhao, Y.; Kurt, Z.; Pan, C.; Zeng, L.; Yang, X.; Schunkert, H.; Lusis, A.J. Applications and Limitations of Mouse Models for Understanding Human Atherosclerosis. Cell Metab. 2017, 25, 248–261. [Google Scholar] [CrossRef]
- Ng, C.P.; Hinz, B.; Swartz, M.A. Interstitial Fluid Flow Induces Myofibroblast Differentiation and Collagen Alignment in Vitro. J. Cell Sci. 2005, 118, 4731–4739. [Google Scholar] [CrossRef]
- Armstrong, J.; Denstedt, J.; Trelford, C.; Li, E.; Hutnik, C. Differential Effects of Dexamethasone and Indomethacin on Tenon’s Capsule Fibroblasts: Implications for Glaucoma Surgery. Exp. Eye Res. 2019, 182, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, E.; Elwing, H.; Askendal, A.; Lundström, I. Protein Immobilization of 3-Aminopropyl Triethoxy Silaneglutaraldehyde Surfaces: Characterization by Detergent Washing. J. Colloid. Interface Sci. 1991, 143, 327–335. [Google Scholar] [CrossRef]
- Gunda, N.S.K.; Singh, M.; Norman, L.; Kaur, K.; Mitra, S.K. Optimization and Characterization of Biomolecule Immobilization on Silicon Substrates Using (3-Aminopropyl)Triethoxysilane (APTES) and Glutaraldehyde Linker. Appl. Surf. Sci. 2014, 305, 522–530. [Google Scholar] [CrossRef]
- Leung, D.Y.L.; Tham, C.C.Y. Management of Bleb Complications after Trabeculectomy. Semin. Ophthalmol. 2013, 28, 144–156. [Google Scholar] [CrossRef]
- Yoshida, M.; Kokubun, T.; Sato, K.; Tsuda, S.; Yokoyama, Y.; Himori, N.; Nakazawa, T. DPP-4 Inhibitors Attenuate Fibrosis after Glaucoma Filtering Surgery by Suppressing the TGF-β/Smad Signaling Pathway. Investig. Ophthalmol. Vis. Sci. 2023, 64, 2. [Google Scholar] [CrossRef]
- Saika, S. TGFbeta Pathobiology in the Eye. Lab. Investig. 2006, 86, 106–115. [Google Scholar] [CrossRef] [PubMed]
- McDowell, C.M.; Tebow, H.E.; Wordinger, R.J.; Clark, A.F. Smad3 Is Necessary for Transforming Growth Factor-Beta2 Induced Ocular Hypertension in Mice. Exp. Eye Res. 2013, 116, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Chong, D.L.W.; Trinder, S.; Labelle, M.; Rodriguez-Justo, M.; Hughes, S.; Holmes, A.M.; Scotton, C.J.; Porter, J.C. Platelet-Derived Transforming Growth Factor-Β1 Promotes Keratinocyte Proliferation in Cutaneous Wound Healing. J. Tissue Eng. Regen. Med. 2020, 14, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Kane, C.J.M.; Hebda, P.A.; Mansbridge, J.N.; Hanawalt, P.C. Direct Evidence for Spatial and Temporal Regulation of Transforming Growth Factor Β1 Expression during Cutaneous Wound Healing. J. Cell Physiol. 1991, 148, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.B.; Sun, X.H.; Dahan, E.; Guo, W.Y.; Qian, S.H.; Meng, F.R.; Song, Y.L.; Simon, G.J.B. Increased Levels of Transforming Growth Factor-Beta1 and -Beta2 in the Aqueous Humor of Patients with Neovascular Glaucoma. Ophthalmic Surg. Lasers Imaging 2007, 38, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tripathi, B.J.; Tripathi, R.C. Modulation of Pre-MRNA Splicing and Protein Production of Fibronectin by TGF-2 in Porcine Trabecular Cells. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3437–3443. [Google Scholar]
- Zhao, X.; Ramsey, K.E.; Stephan, D.A.; Russell, P. Gene and Protein Expression Changes in Human Trabecular Meshwork Cells Treated with Transforming Growth Factor-Beta. Investig. Ophthalmol. Vis. Sci. 2004, 45, 4023–4034. [Google Scholar] [CrossRef]
- Flanders, K.C. Smad3 as a Mediator of the Fibrotic Response. Int. J. Exp. Pathol. 2004, 85, 47–64. [Google Scholar] [CrossRef]
- Meng, X.M.; Chung, A.C.K.; Lan, H.Y. Role of the TGF-β/BMP-7/Smad Pathways in Renal Diseases. Clin. Sci. 2013, 124, 243–254. [Google Scholar] [CrossRef]
- Chen, J.; Xia, Y.; Lin, X.; Feng, X.H.; Wang, Y. Smad3 Signaling Activates Bone Marrow-Derived Fibroblasts in Renal Fibrosis. Lab. Investig. 2014, 94, 545–556. [Google Scholar] [CrossRef]
- Tojkander, S.; Gateva, G.; Lappalainen, P. Actin Stress Fibers—Assembly, Dynamics and Biological Roles. J. Cell Sci. 2012, 125, 1855–1864. [Google Scholar] [CrossRef]
- Bressler, N.M. Photodynamic Therapy of Subfoveal Choroidal Neovascularization in Age- Related Macular Degeneration with Verteporfin: One-Year Results of 2 Randomized Clinical Trials—TAP Report 1. Arch. Ophthalmol. 1999, 117, 1329–1345. [Google Scholar] [CrossRef]
- Szeto, S.G.; Narimatsu, M.; Lu, M.; He, X.; Sidiqi, A.M.; Tolosa, M.F.; Chan, L.; De Freitas, K.; Bialik, J.F.; Majumder, S.; et al. YAP/TAZ Are Mechanoregulators of TGF-b-Smad Signaling and Renal Fibrogenesis. J. Am. Soc. Nephrol. 2016, 27, 3117–3128. [Google Scholar] [CrossRef] [PubMed]
- Liu-Chittenden, Y.; Huang, B.; Shim, J.S.; Chen, Q.; Lee, S.J.; Anders, R.A.; Liu, J.O.; Pan, D. Genetic and Pharmacological Disruption of the TEAD-YAP Complex Suppresses the Oncogenic Activity of YAP. Genes. Dev. 2012, 26, 1300–1305. [Google Scholar] [CrossRef]
- Piersma, B.; Bank, R.A.; Boersema, M. Signaling in Fibrosis: TGF-β, WNT, and YAP/TAZ Converge. Front. Med. 2015, 2, 59. [Google Scholar] [CrossRef]
- Miyazawa, K.; Miyazono, K. Regulation of TGF-β Family Signaling by Inhibitory Smads. Cold Spring Harb. Perspect. Biol. 2017, 9, a022095. [Google Scholar] [CrossRef]
- Wang, B.; Omar, A.; Angelovska, T.; Drobic, V.; Rattan, S.G.; Jones, S.C.; Dixon, I.M.C. Regulation of Collagen Synthesis by Inhibitory Smad7 in Cardiac Myofibroblasts. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1282–H1290. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Huang, N.; Lin, J.-Q.; Yan, L.-Y.; Jiang, Q.-G.; Yang, W.-Z. Effect and Mechanism of Pirfenidone Combined with 2-Methoxy-Estradiol Perfusion through Portal Vein on Hepatic Artery Hypoxia-Induced Hepatic Fibrosis. Adv. Med. Sci. 2023, 68, 46–53. [Google Scholar] [CrossRef]
- Chen, W.S.; Cao, Z.; Krishnan, C.; Panjwani, N. Verteporfin without Light Stimulation Inhibits YAP Activation in Trabecular Meshwork Cells: Implications for Glaucoma Treatment. Biochem. Biophys. Res. Commun. 2015, 466, 221–225. [Google Scholar] [CrossRef]
- Jin, J.; Wang, T.; Park, W.; Li, W.; Kim, W.; Park, S.K.; Kang, K.P. Inhibition of Yes-Associated Protein by Verteporfin Ameliorates Unilateral Ureteral Obstruction-Induced Renal Tubulointerstitial Inflammation and Fibrosis. Int. J. Mol. Sci. 2020, 21, 8184. [Google Scholar] [CrossRef] [PubMed]
- Houle, J.M.; Strong, A. Clinical Pharmacokinetics of Verteporfin. J. Clin. Pharmacol. 2002, 42, 547–557. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fung, M.; Armstrong, J.J.; Zhang, R.; Vinokurtseva, A.; Liu, H.; Hutnik, C. Development and Verification of a Novel Three-Dimensional Aqueous Outflow Model for High-Throughput Drug Screening. Bioengineering 2024, 11, 142. https://doi.org/10.3390/bioengineering11020142
Fung M, Armstrong JJ, Zhang R, Vinokurtseva A, Liu H, Hutnik C. Development and Verification of a Novel Three-Dimensional Aqueous Outflow Model for High-Throughput Drug Screening. Bioengineering. 2024; 11(2):142. https://doi.org/10.3390/bioengineering11020142
Chicago/Turabian StyleFung, Matthew, James J. Armstrong, Richard Zhang, Anastasiya Vinokurtseva, Hong Liu, and Cindy Hutnik. 2024. "Development and Verification of a Novel Three-Dimensional Aqueous Outflow Model for High-Throughput Drug Screening" Bioengineering 11, no. 2: 142. https://doi.org/10.3390/bioengineering11020142
APA StyleFung, M., Armstrong, J. J., Zhang, R., Vinokurtseva, A., Liu, H., & Hutnik, C. (2024). Development and Verification of a Novel Three-Dimensional Aqueous Outflow Model for High-Throughput Drug Screening. Bioengineering, 11(2), 142. https://doi.org/10.3390/bioengineering11020142