

Model-Based Characterization of E. coli Strains with Impaired Glucose Uptake

 , ,
, ,  , ,
, ,  and
and 
Abstract
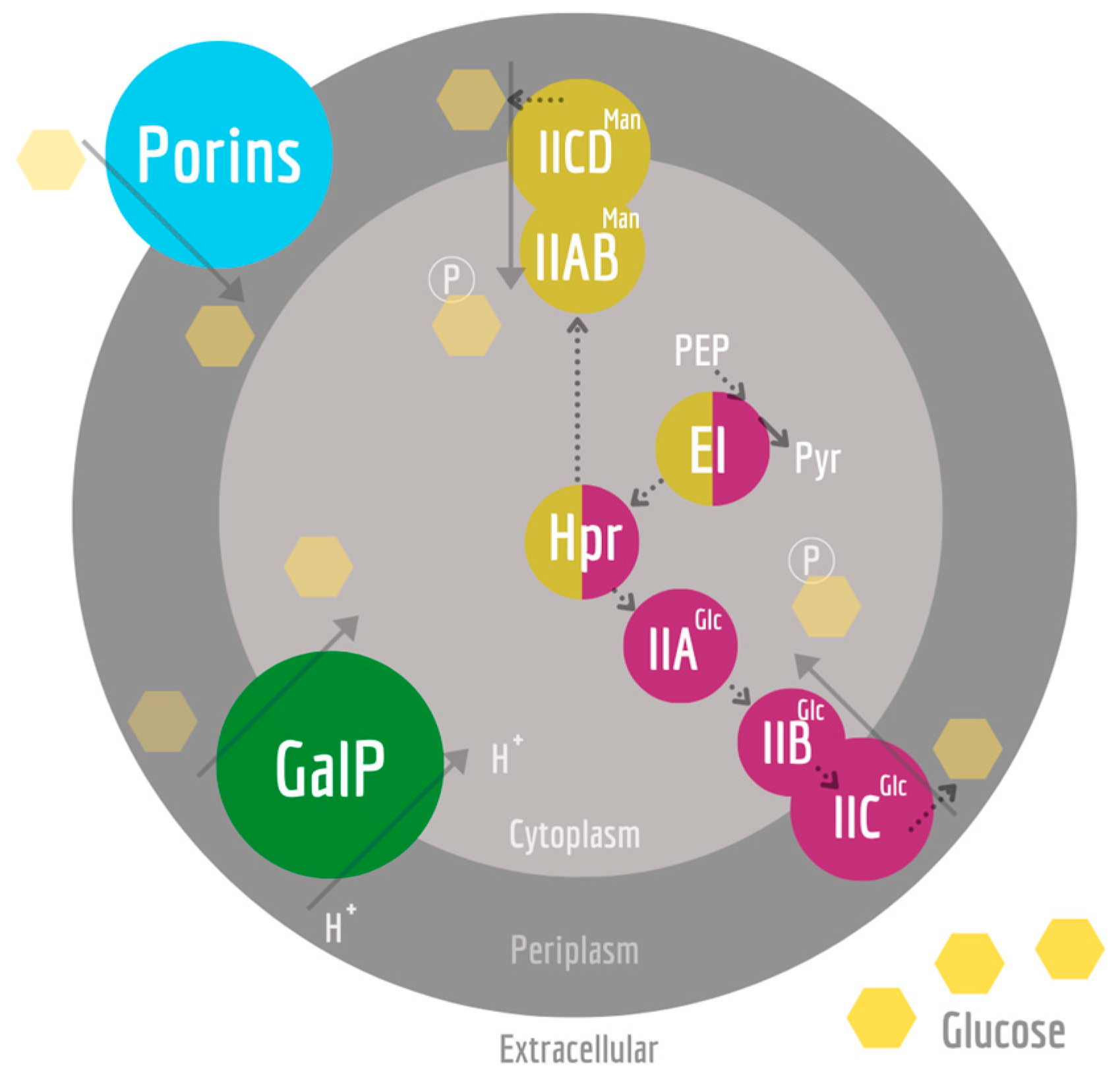
1. Introduction
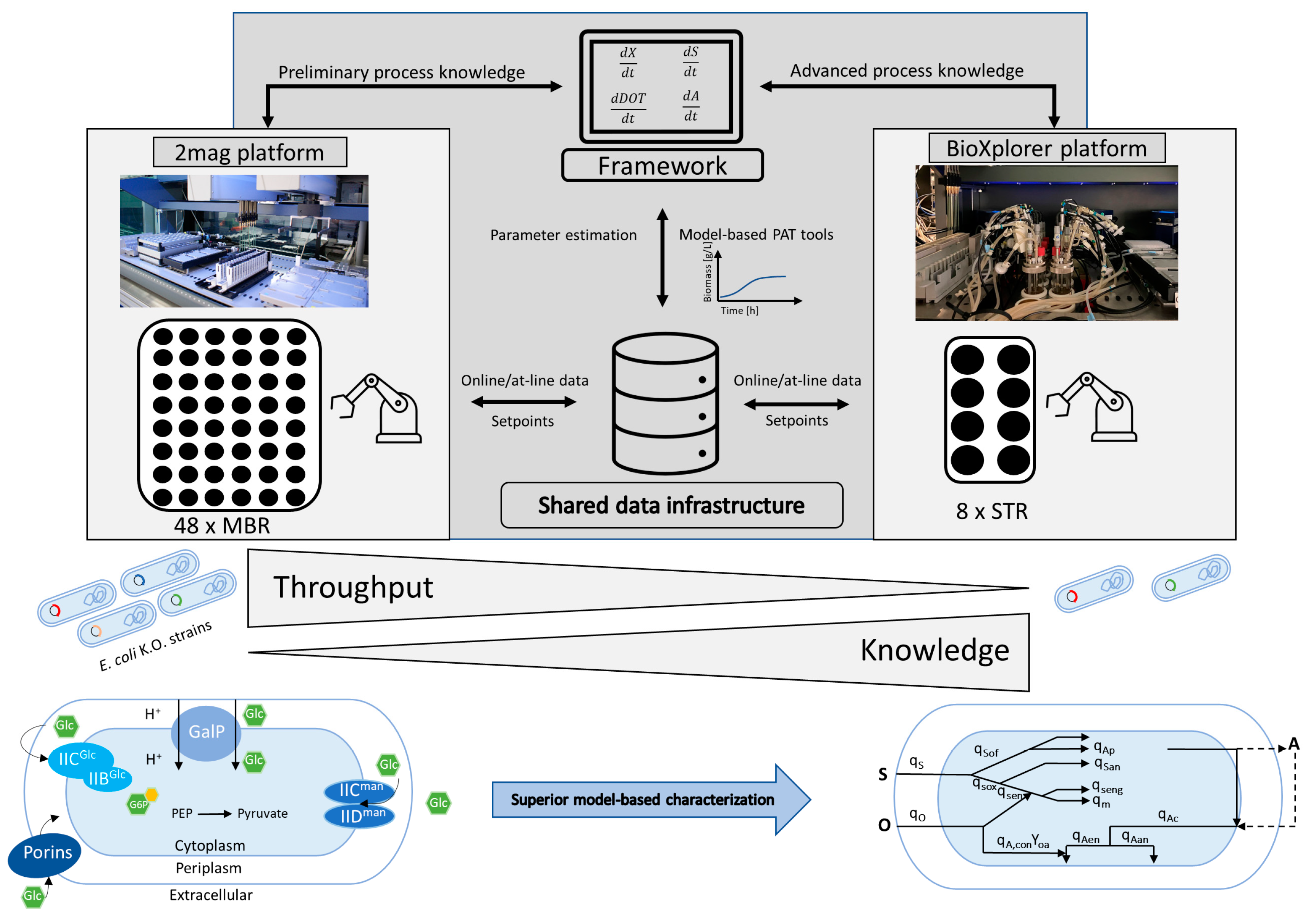
2. Materials and Methods
2.1. Strain and Cultivation Conditions
2.2. HT Bioprocess Development Facility
2.2.1. MTP Cultivation
2.2.2. 2mag Cultivation Platform
2.2.3. BioXplorer Cultivation Platform
2.2.4. Mobile Robotic Lab Assistant
2.2.5. High-Throughput Metabolite Analysis
2.3. Data Handling
2.4. Mathematical Modeling Tools
2.5. Off-Gas Analysis
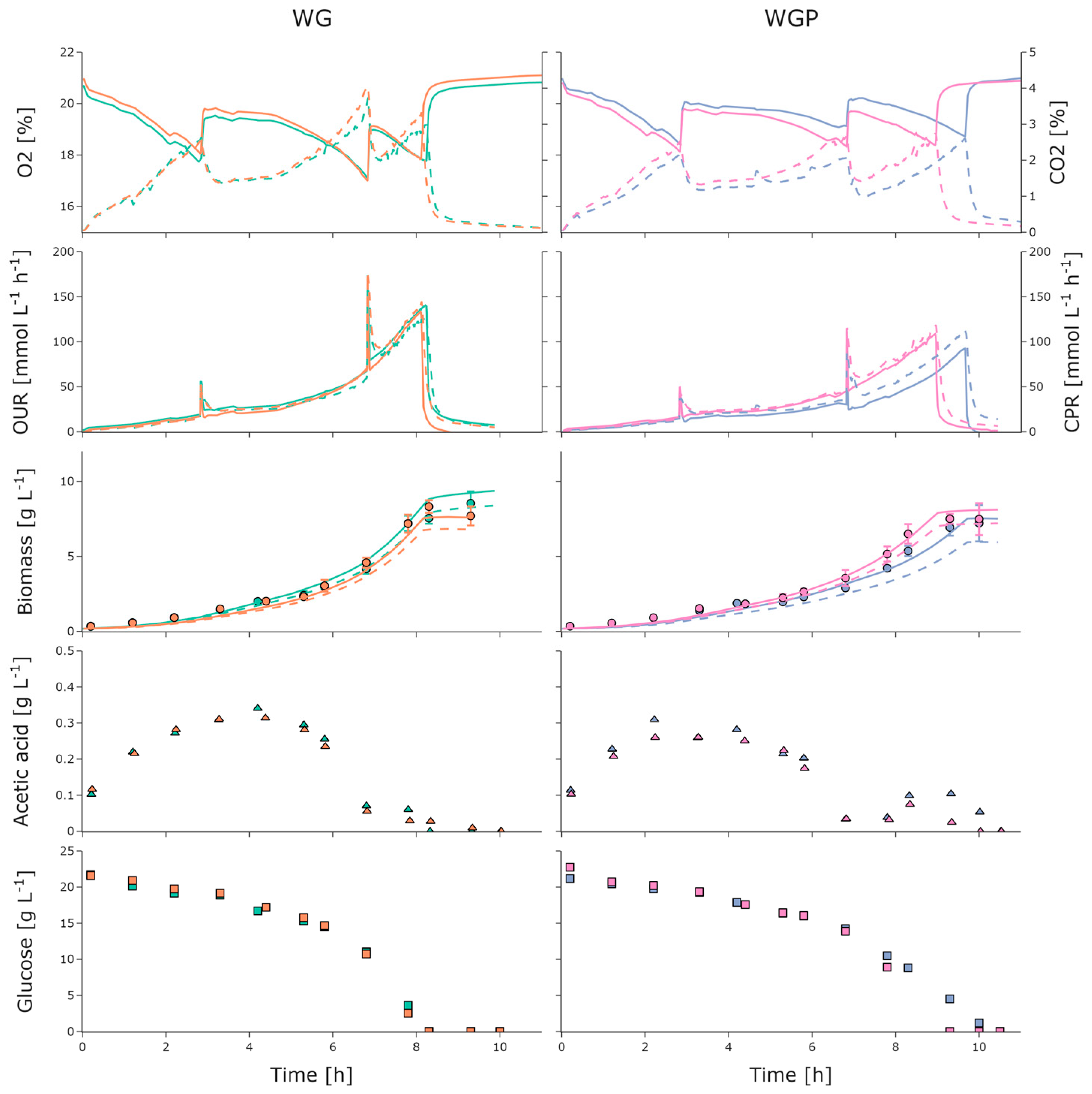
3. Results
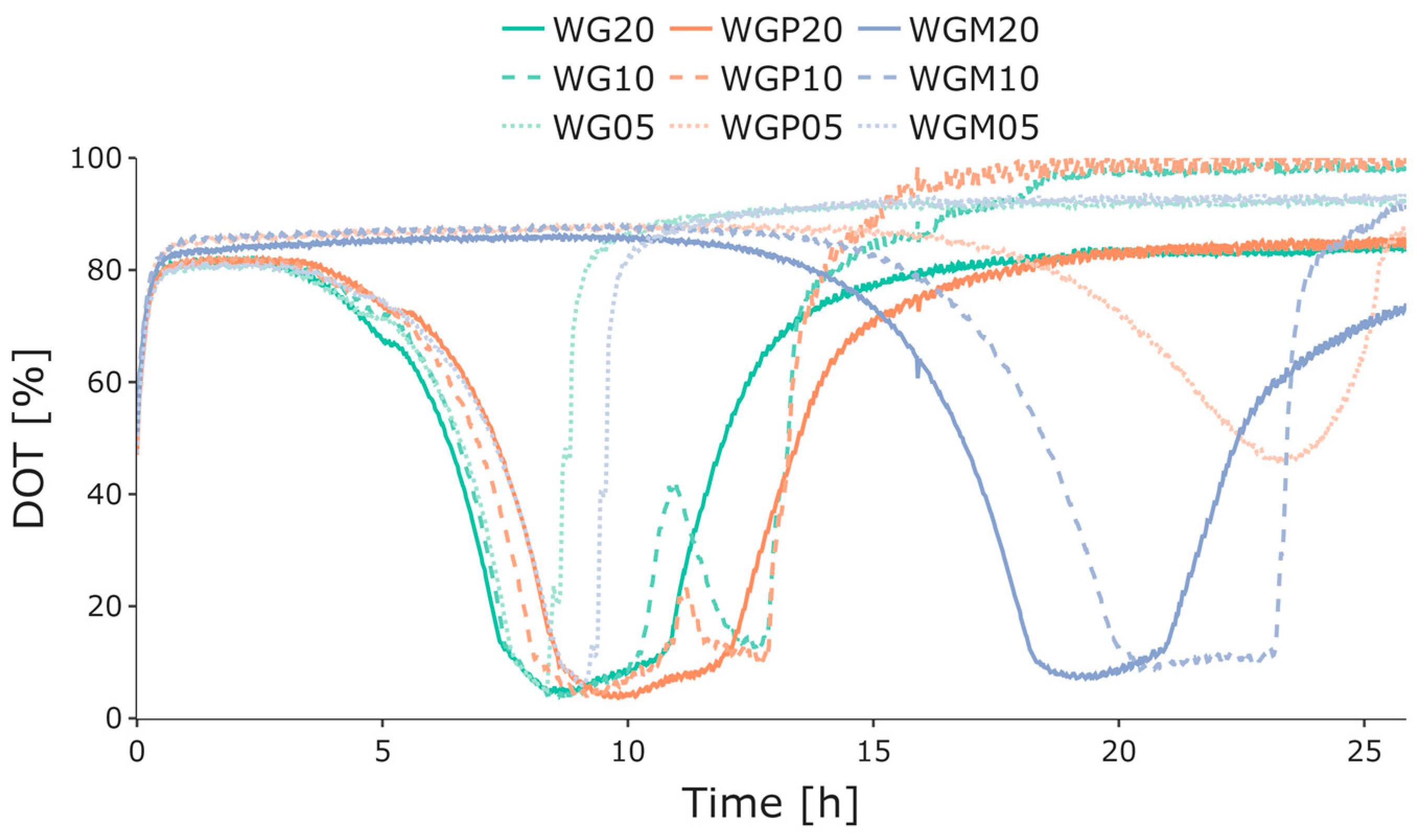
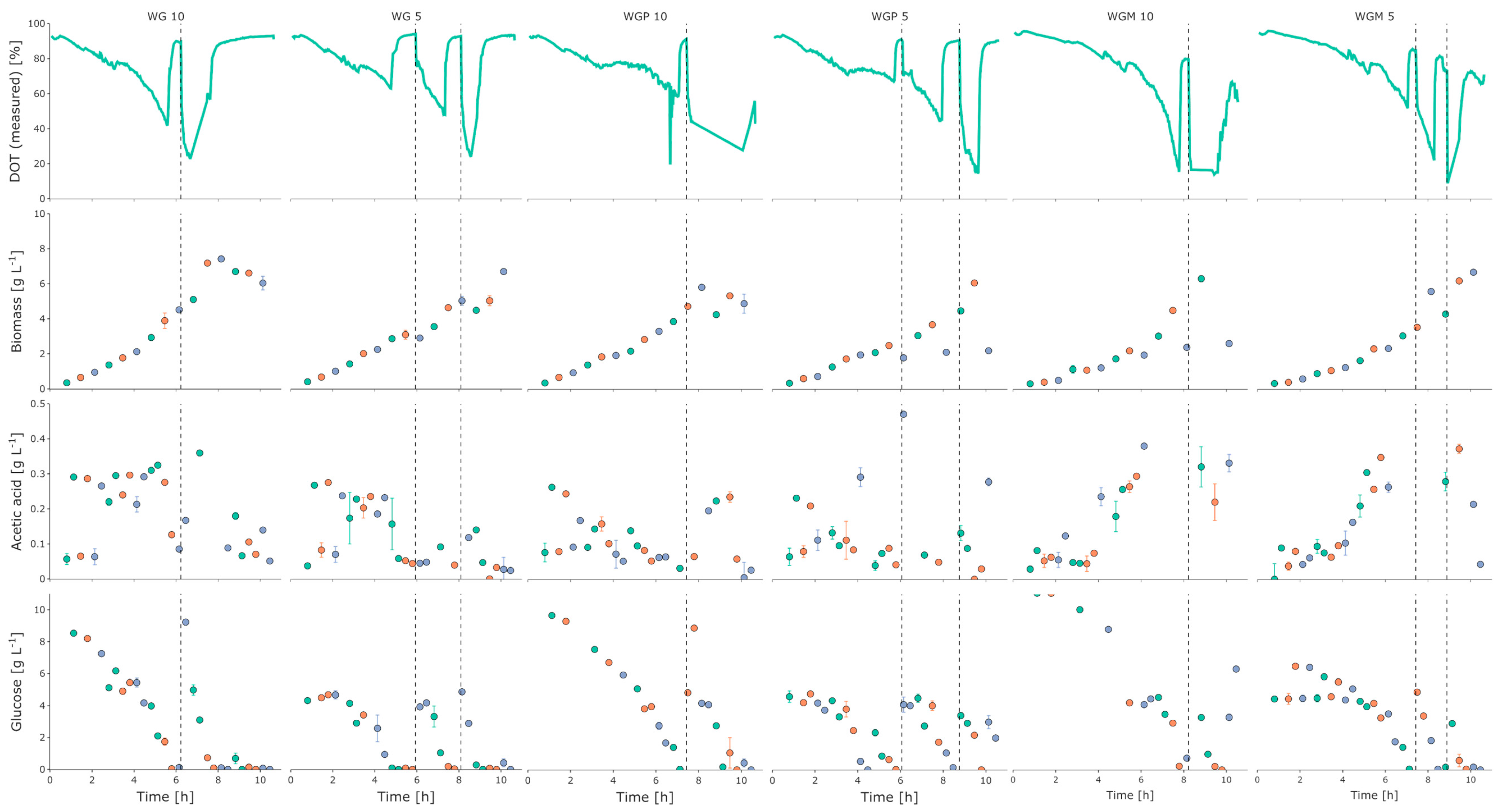
3.1. Initial Growth Characterization
3.2. BioXplorer Cultivation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, C.-J.; Lin, H.; Yang, X. Industrial production of recombinant therapeutics in Escherichia coli and its recent advancements. J. Ind. Microbiol. Biotechnol. 2012, 39, 383–399. [Google Scholar] [CrossRef] [PubMed]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, P.; Cruz Bournazou, M.N.; Glauche, F.; Junne, S.; Knepper, A.; Raven, M. Consistent development of bioprocesses from microliter cultures to the industrial scale. Eng. Life Sci. 2013, 13, 224–238. [Google Scholar] [CrossRef]
- Pan, J.G.; Rhee, J.S.; Lebeault, J.M. Physiological constraints in increasing biomass concentration of Escherichia coli B in fed-batch culture. Biotechnol. Lett. 1987, 9, 89–94. [Google Scholar] [CrossRef]
- Xu, B.; Jahic, M.; Enfors, S.O. Modeling of overflow metabolism in batch and fed-batch cultures of Escherichia coli. Biotechnol. Prog. 1999, 15, 81–90. [Google Scholar] [CrossRef]
- Bernal, V.; Castaño-Cerezo, S.; Cánovas, M. Acetate metabolism regulation in Escherichia coli: Carbon overflow, pathogenicity, and beyond. Appl. Microbiol. Biotechnol. 2016, 100, 8985–9001. [Google Scholar] [CrossRef]
- Dittrich, C.R.; Vadali, R.V.; Bennett, G.N.; San, K.-Y. Redistribution of metabolic fluxes in the central aerobic metabolic pathway of E. coli mutant strains with deletion of the ackA-pta and poxB pathways for the synthesis of isoamyl acetate. Biotechnol. Prog. 2005, 21, 627–631. [Google Scholar] [CrossRef]
- Lara, A.R.; Caspeta, L.; Gosset, G.; Bolívar, F.; Ramírez, O.T. Utility of an Escherichia coli strain engineered in the substrate uptake system for improved culture performance at high glucose and cell concentrations: An alternative to fed-batch cultures. Biotechnol. Bioeng. 2008, 99, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, L.G.; Lara, A.R.; Martínez, L.M.; Ramírez, O.T.; Martínez, A.; Bolívar, F.; Gosset, G. Modification of glucose import capacity in Escherichia coli: Physiologic consequences and utility for improving DNA vaccine production. Microb. Cell Fact. 2013, 12, 42. [Google Scholar] [CrossRef]
- Fragoso-Jiménez, J.C.; Baert, J.; Nguyen, T.M.; Liu, W.; Sassi, H.; Goormaghtigh, F.; van Melderen, L.; Gaytán, P.; Hernández-Chávez, G.; Martínez, A.; et al. Growth-dependent recombinant product formation kinetics can be reproduced through engineering of glucose transport and is prone to phenotypic heterogeneity. Microb. Cell Fact. 2019, 18, 26. [Google Scholar] [CrossRef]
- Velazquez, D.; Sigala, J.-C.; Martínez, L.M.; Gaytán, P.; Gosset, G.; Lara, A.R. Glucose transport engineering allows mimicking fed-batch performance in batch mode and selection of superior producer strains. Microb. Cell Fact. 2022, 21, 183. [Google Scholar] [CrossRef] [PubMed]
- Knabben, I.; Regestein, L.; Marquering, F.; Steinbusch, S.; Lara, A.R.; Büchs, J. High cell-density processes in batch mode of a genetically engineered Escherichia coli strain with minimized overflow metabolism using a pressurized bioreactor. J. Biotechnol. 2010, 150, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Hemmerich, J.; Noack, S.; Wiechert, W.; Oldiges, M. Microbioreactor Systems for Accelerated Bioprocess Development. Biotechnol. J. 2018, 13, e1700141. [Google Scholar] [CrossRef] [PubMed]
- Bareither, R.; Pollard, D. A review of advanced small-scale parallel bioreactor technology for accelerated process development: Current state and future need. Biotechnol. Prog. 2011, 27, 2–14. [Google Scholar] [CrossRef]
- Janzen, N.H.; Striedner, G.; Jarmer, J.; Voigtmann, M.; Abad, S.; Reinisch, D. Implementation of a Fully Automated Microbial Cultivation Platform for Strain and Process Screening. Biotechnol. J. 2019, 14, e1800625. [Google Scholar] [CrossRef]
- Haby, B.; Hans, S.; Anane, E.; Sawatzki, A.; Krausch, N.; Neubauer, P.; Cruz Bournazou, M.N. Integrated Robotic Mini Bioreactor Platform for Automated, Parallel Microbial Cultivation With Online Data Handling and Process Control. SLAS Technol. 2019, 24, 569–582. [Google Scholar] [CrossRef]
- Morschett, H.; Tenhaef, N.; Hemmerich, J.; Herbst, L.; Spiertz, M.; Dogan, D.; Wiechert, W.; Noack, S.; Oldiges, M. Robotic integration enables autonomous operation of laboratory scale stirred tank bioreactors with model-driven process analysis. Biotechnol. Bioeng. 2021, 118, 2759–2769. [Google Scholar] [CrossRef]
- Cruz Bournazou, M.N.; Barz, T.; Nickel, D.B.; Lopez Cárdenas, D.C.; Glauche, F.; Knepper, A.; Neubauer, P. Online optimal experimental re-design in robotic parallel fed-batch cultivation facilities. Biotechnol. Bioeng. 2017, 114, 610–619. [Google Scholar] [CrossRef]
- Anane, E.; García, Á.C.; Haby, B.; Hans, S.; Krausch, N.; Krewinkel, M.; Hauptmann, P.; Neubauer, P.; Cruz Bournazou, M.N. A model-based framework for parallel scale-down fed-batch cultivations in mini-bioreactors for accelerated phenotyping. Biotechnol. Bioeng. 2019, 116, 2906–2918. [Google Scholar] [CrossRef]
- Giese, H.; Azizan, A.; Kümmel, A.; Liao, A.; Peter, C.P.; Fonseca, J.A.; Hermann, R.; Duarte, T.M.; Büchs, J. Liquid films on shake flask walls explain increasing maximum oxygen transfer capacities with elevating viscosity. Biotechnol. Bioeng. 2014, 111, 295–308. [Google Scholar] [CrossRef]
- Aehle, M.; Kuprijanov, A.; Schaepe, S.; Simutis, R.; Lübbert, A. Simplified off-gas analyses in animal cell cultures for process monitoring and control purposes. Biotechnol. Lett. 2011, 33, 2103–2110. [Google Scholar] [CrossRef] [PubMed]
- Jenzsch, M.; Simutis, R.; Eisbrenner, G.; Stückrath, I.; Lübbert, A. Estimation of biomass concentrations in fermentation processes for recombinant protein production. Bioprocess Biosyst. Eng. 2006, 29, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Luedeking, R.; Piret, E.L. A kinetic study of the lactic acid fermentation. Batch process at controlled pH. J. Biochem. Microbiol. Technol. Eng. 1959, 1, 393–412. [Google Scholar] [CrossRef]
- Kaspersetz, L.; Waldburger, S.; Schermeyer, M.-T.; Riedel, S.L.; Groß, S.; Neubauer, P.; Cruz Bournazou, M.N. Automated Bioprocess Feedback Operation in a High-Throughput Facility via the Integration of a Mobile Robotic Lab Assistant. Front. Chem. Eng. 2022, 4, 196–208. [Google Scholar] [CrossRef]
- Krausch, N.; Kim, J.W.; Barz, T.; Lucia, S.; Groß, S.; Huber, M.C.; Schiller, S.M.; Neubauer, P.; Cruz Bournazou, M.N. High-throughput screening of optimal process conditions using model predictive control. Biotechnol. Bioeng. 2022, 119, 3584–3595. [Google Scholar] [CrossRef]
- Anane, E.; Sawatzki, A.; Neubauer, P.; Cruz Bournazou, M.N. Modelling concentration gradients in fed-batch cultivations of E. coli-towards the flexible design of scale-down experiments. J. Chem. Technol. Biotechnol. 2019, 94, 516–526. [Google Scholar] [CrossRef]
- Kim, J.W.; Krausch, N.; Aizpuru, J.; Barz, T.; Lucia, S.; Neubauer, P.; Cruz Bournazou, M.N. Model predictive control and moving horizon estimation for adaptive optimal bolus feeding in high-throughput cultivation of E. coli. Comput. Chem. Eng. 2022, 172, 108158. [Google Scholar] [CrossRef]
- Wechselberger, P.; Sagmeister, P.; Herwig, C. Real-time estimation of biomass and specific growth rate in physiologically variable recombinant fed-batch processes. Bioprocess Biosyst. Eng. 2013, 36, 1205–1218. [Google Scholar] [CrossRef]
- Urniezius, R.; Survyla, A.; Paulauskas, D.; Bumelis, V.A.; Galvanauskas, V. Generic estimator of biomass concentration for Escherichia coli and Saccharomyces cerevisiae fed-batch cultures based on cumulative oxygen consumption rate. Microb. Cell Fact. 2019, 18, 190. [Google Scholar] [CrossRef]
- Puskeiler, R.; Kaufmann, K.; Weuster-Botz, D. Development, parallelization, and automation of a gas-inducing milliliter-scale bioreactor for high-throughput bioprocess design (HTBD). Biotechnol. Bioeng. 2005, 89, 512–523. [Google Scholar] [CrossRef]
- Liang, Q.; Zhang, F.; Li, Y.; Zhang, X.; Li, J.; Yang, P.; Qi, Q. Comparison of individual component deletions in a glucose-specific phosphotransferase system revealed their different applications. Sci. Rep. 2015, 5, 13200. [Google Scholar] [CrossRef] [PubMed]
- Steinsiek, S.; Bettenbrock, K. Glucose transport in Escherichia coli mutant strains with defects in sugar transport systems. J. Bacteriol. 2012, 194, 5897–5908. [Google Scholar] [CrossRef] [PubMed]
- Misset, O.; Blaauw, M.; Postma, P.W.; Robillard, G.T. Bacterial phosphoenolpyruvate-dependent phosphotransferase system. Mechanism of the transmembrane sugar translocation and phosphorylation. Biochemistry 1983, 22, 6163–6170. [Google Scholar] [CrossRef] [PubMed]
- Smaluch, K.; Wollenhaupt, B.; Steinhoff, H.; Kohlheyer, D.; Grünberger, A.; Dusny, C. Assessing the growth kinetics and stoichiometry of Escherichia coli at the single-cell level. Eng. Life Sci. 2023, 23, e2100157. [Google Scholar] [CrossRef] [PubMed]
- Valgepea, K.; Adamberg, K.; Nahku, R.; Lahtvee, P.-J.; Arike, L.; Vilu, R. Systems biology approach reveals that overflow metabolism of acetate in Escherichia coli is triggered by carbon catabolite repression of acetyl-CoA synthetase. BMC Syst. Biol. 2010, 4, 166. [Google Scholar] [CrossRef] [PubMed]
- Borja, G.M.; Meza Mora, E.; Barrón, B.; Gosset, G.; Ramírez, O.T.; Lara, A.R. Engineering Escherichia coli to increase plasmid DNA production in high cell-density cultivations in batch mode. Microb. Cell Fact. 2012, 11, 132. [Google Scholar] [CrossRef]
- Garcia-Ochoa, F.; Gomez, E.; Santos, V.E.; Merchuk, J.C. Oxygen uptake rate in microbial processes: An overview. Biochem. Eng. J. 2010, 49, 289–307. [Google Scholar] [CrossRef]
- Lin, H.Y.; Mathiszik, B.; Xu, B.; Enfors, S.O.; Neubauer, P. Determination of the maximum specific uptake capacities for glucose and oxygen in glucose-limited fed-batch cultivations of Escherichia coli. Biotechnol. Bioeng. 2001, 73, 347–357. [Google Scholar] [CrossRef]
- Wolfe, A.J. The acetate switch. Microbiol. Mol. Biol. Rev. 2005, 69, 12–50. [Google Scholar] [CrossRef]
- O’Beirne, D.; Hamer, G. The utilisation of glucose/acetate mixtures by Escherichia coli W3110 under aerobic growth conditions. Bioprocess Biosyst. Eng. 2000, 23, 375–380. [Google Scholar] [CrossRef]
- Enjalbert, B.; Millard, P.; Dinclaux, M.; Portais, J.-C.; Létisse, F. Acetate fluxes in Escherichia coli are determined by the thermodynamic control of the Pta-AckA pathway. Sci. Rep. 2017, 7, 42135. [Google Scholar] [CrossRef] [PubMed]
- Enjalbert, B.; Cocaign-Bousquet, M.; Portais, J.-C.; Letisse, F. Acetate Exposure Determines the Diauxic Behavior of Escherichia coli during the Glucose-Acetate Transition. J. Bacteriol. 2015, 197, 3173–3181. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.X. Pharmaceutical quality by design: Product and process development, understanding, and control. Pharm. Res. 2008, 25, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Hemmerich, J.; Adelantado, N.; Barrigón, J.M.; Ponte, X.; Hörmann, A.; Ferrer, P.; Kensy, F.; Valero, F. Comprehensive clone screening and evaluation of fed-batch strategies in a microbioreactor and lab scale stirred tank bioreactor system: Application on Pichia pastoris producing Rhizopus oryzae lipase. Microb. Cell Fact. 2014, 13, 36. [Google Scholar] [CrossRef]
- Batchu, S.P.; Hernandez, B.; Malhotra, A.; Fang, H.; Ierapetritou, M.; Vlachos, D.G. Accelerating manufacturing for biomass conversion via integrated process and bench digitalization: A perspective. React. Chem. Eng. 2022, 7, 813–832. [Google Scholar] [CrossRef]
- King, R.D.; Rowland, J.; Oliver, S.G.; Young, M.; Aubrey, W.; Byrne, E.; Liakata, M.; Markham, M.; Pir, P.; Soldatova, L.N.; et al. The automation of science. Science 2009, 324, 85–89. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype |
|---|---|
| WG | W3110 ΔptsG |
| WGM | W3110 ΔptsG, ΔmanX |
| WGP | W3110 ΔptsG, ΔgalP |
| Parameter | WG (5) | WG (10) | WGP (5) | WGP (10) | WGM (5) | WGM (10) | WG (20) | WGP (20) |
|---|---|---|---|---|---|---|---|---|
| µmax [h−1] | 0.55 ± 0.06 | 0.62 ± 0.09 | 0.46 ± 0.04 | 0.45 ± 0.04 | 0.41 ± 0.03 | 0.47 ± 0.05 | 0.44 ± 0.03 | 0.36 ± 0.02 |
| KS [g L−1] | 0.010 ± 0.002 | 0.009 ± 0.002 | 0.011 ± 0.002 | 0.010 ± 0.002 | 0.010 ± 0.002 | 0.011 ± 0.002 | 0.010 ± 0.002 | 0.010 ± 0.002 |
| qSmax [g g−1 h−1] | 1.12 ± 0.01 | 1.13 ± 0.01 | 0.98 ± 0.01 | 0.97 ± 0.01 | 0.87 ± 0.01 | 0.98 ± 0.01 | 1.03 ± 0.01 | 0.98 ± 0.01 |
| qApmax [g g−1 h−1] | 0.10 ± 0.01 | 0.10 ± 0.01 | 0.05 ± 0.00 | 0.04 ± 0.00 | 0.13 ± 0.01 | 0.08 ± 0.01 | 0.06 ± 0.01 | 0.06 ± 0.01 |
| qAcmax [g g−1 h−1] | 0.07 ± 0.00 | 0.07 ± 0.01 | 0.06 ± 0.00 | 0.06 ± 0.01 | 0.06 ± 0.01 | 0.07 ± 0.00 | 0.08 ± 0.00 | 0.0708 ± 0.00 |
| qOmax [g g−1 h−1] | 0.79 ± 0.01 | 0.91 ± 0.01 | 0.67 ± 0.01 | 0.66 ± 0.01 | 0.62 ± 0.01 | 0.68 ± 0.01 | 0.55 ± 0.01 | 0.41 ± 0.01 |
| qm [g g−1 h−1] | 0.047 ± 0.002 | 0.056 ± 0.004 | 0.044 ± 0.003 | 0.047 ± 0.004 | 0.044 ± 0.004 | 0.044 ± 0.003 | 0.044 ± 0.002 | 0.044 ± 0.003 |
| YX/S,em [g g−1] | 0.53 ± 0.06 | 0.56 ± 0.08 | 0.49 ± 0.04 | 0.49 ± 0.05 | 0.50 ± 0.04 | 0.51 ± 0.05 | 0.45 ± 0.03 | 0.39 ± 0.02 |
| YOX [g g−1] | 1.43 ± 0.01 | 1.49 ± 0.01 | 1.45 ± 0.01 | 1.47 ± 0.01 | 1.46 ± 0.01 | 1.45 ± 0.01 | 1.25 ± 0.01 | 1.2 4 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krausch, N.; Kaspersetz, L.; Gaytán-Castro, R.D.; Schermeyer, M.-T.; Lara, A.R.; Gosset, G.; Cruz Bournazou, M.N.; Neubauer, P. Model-Based Characterization of E. coli Strains with Impaired Glucose Uptake. Bioengineering 2023, 10, 808. https://doi.org/10.3390/bioengineering10070808
Krausch N, Kaspersetz L, Gaytán-Castro RD, Schermeyer M-T, Lara AR, Gosset G, Cruz Bournazou MN, Neubauer P. Model-Based Characterization of E. coli Strains with Impaired Glucose Uptake. Bioengineering. 2023; 10(7):808. https://doi.org/10.3390/bioengineering10070808
Chicago/Turabian StyleKrausch, Niels, Lucas Kaspersetz, Rogelio Diego Gaytán-Castro, Marie-Therese Schermeyer, Alvaro R. Lara, Guillermo Gosset, Mariano Nicolas Cruz Bournazou, and Peter Neubauer. 2023. "Model-Based Characterization of E. coli Strains with Impaired Glucose Uptake" Bioengineering 10, no. 7: 808. https://doi.org/10.3390/bioengineering10070808
APA StyleKrausch, N., Kaspersetz, L., Gaytán-Castro, R. D., Schermeyer, M.-T., Lara, A. R., Gosset, G., Cruz Bournazou, M. N., & Neubauer, P. (2023). Model-Based Characterization of E. coli Strains with Impaired Glucose Uptake. Bioengineering, 10(7), 808. https://doi.org/10.3390/bioengineering10070808