

Aptasensor for the Detection of Moraxella catarrhalis Adhesin UspA2

 ,
,  ,
,  ,
,  , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Sedimentation Assay
2.3. Protein Analysis
2.4. ssDNA Library, Aptamer Sequences and Primers
2.5. In Vitro Selection of Aptamers by Cell-SELEX
2.6. Sequencing and Data Analysis
2.7. Binding Experiments
2.8. Electrochemical Apparatus, Electrodes and Reagents
2.9. Assembly of the Aptasensor
2.10. Evaluation of the Aptasensor in Standard Solutions
3. Results and Discussion
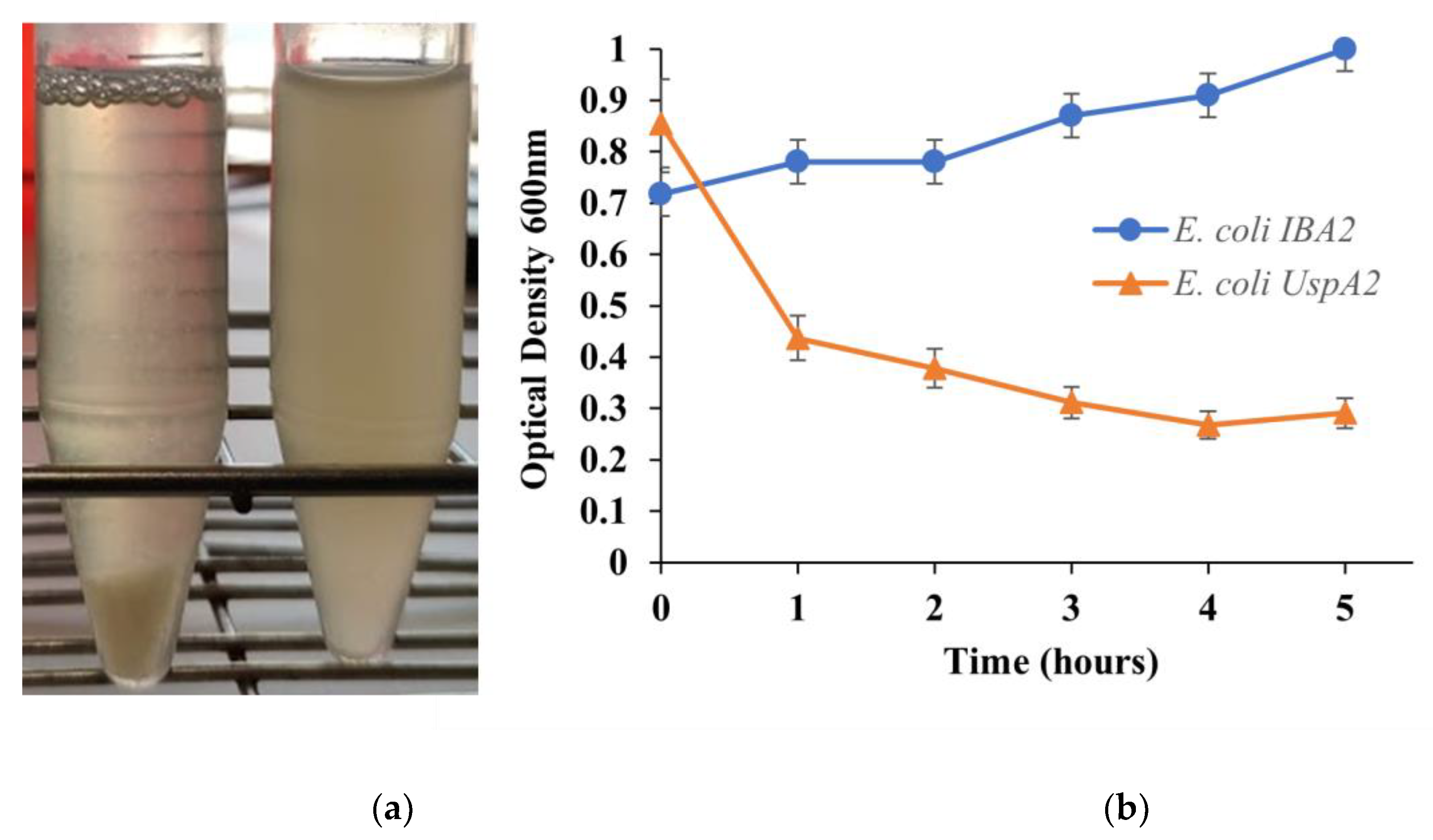
3.1. Characterization of UspA2
3.2. Selection of Aptamers with High Affinity for UspA2 by Cell-SELEX
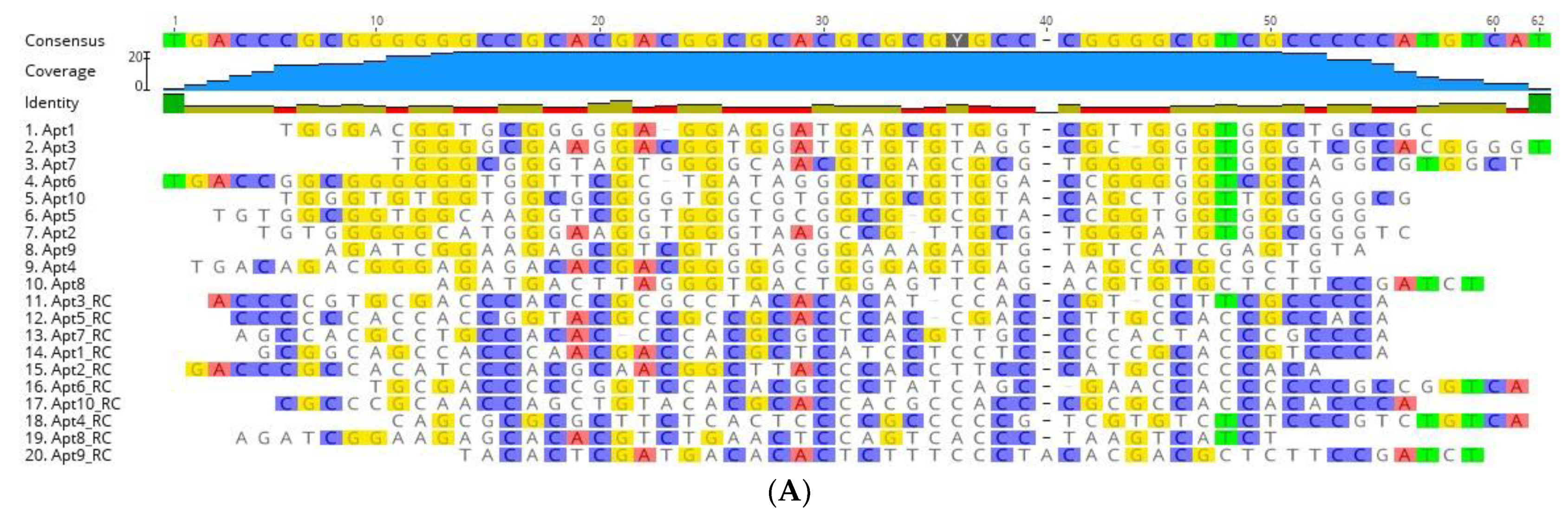
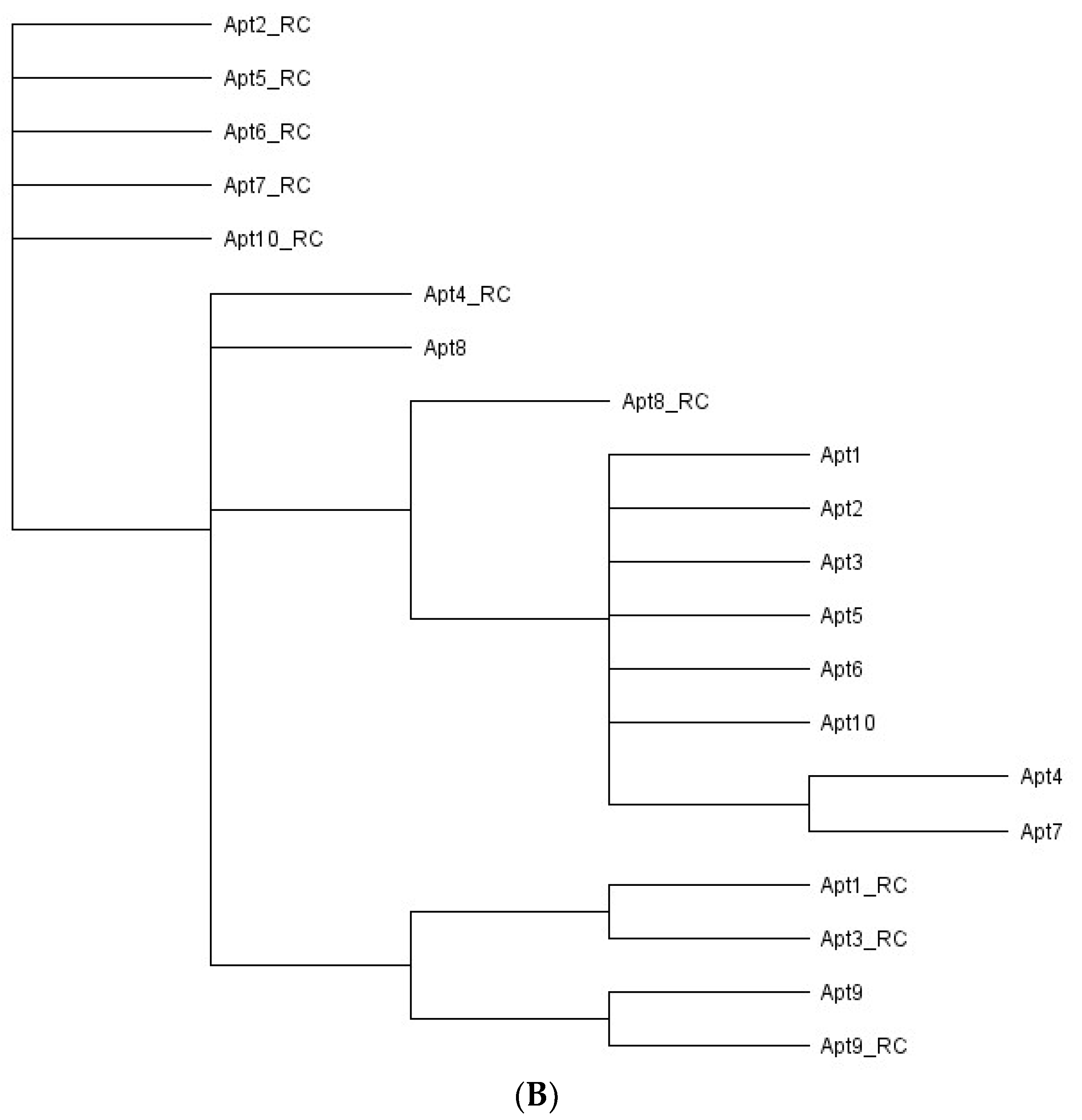
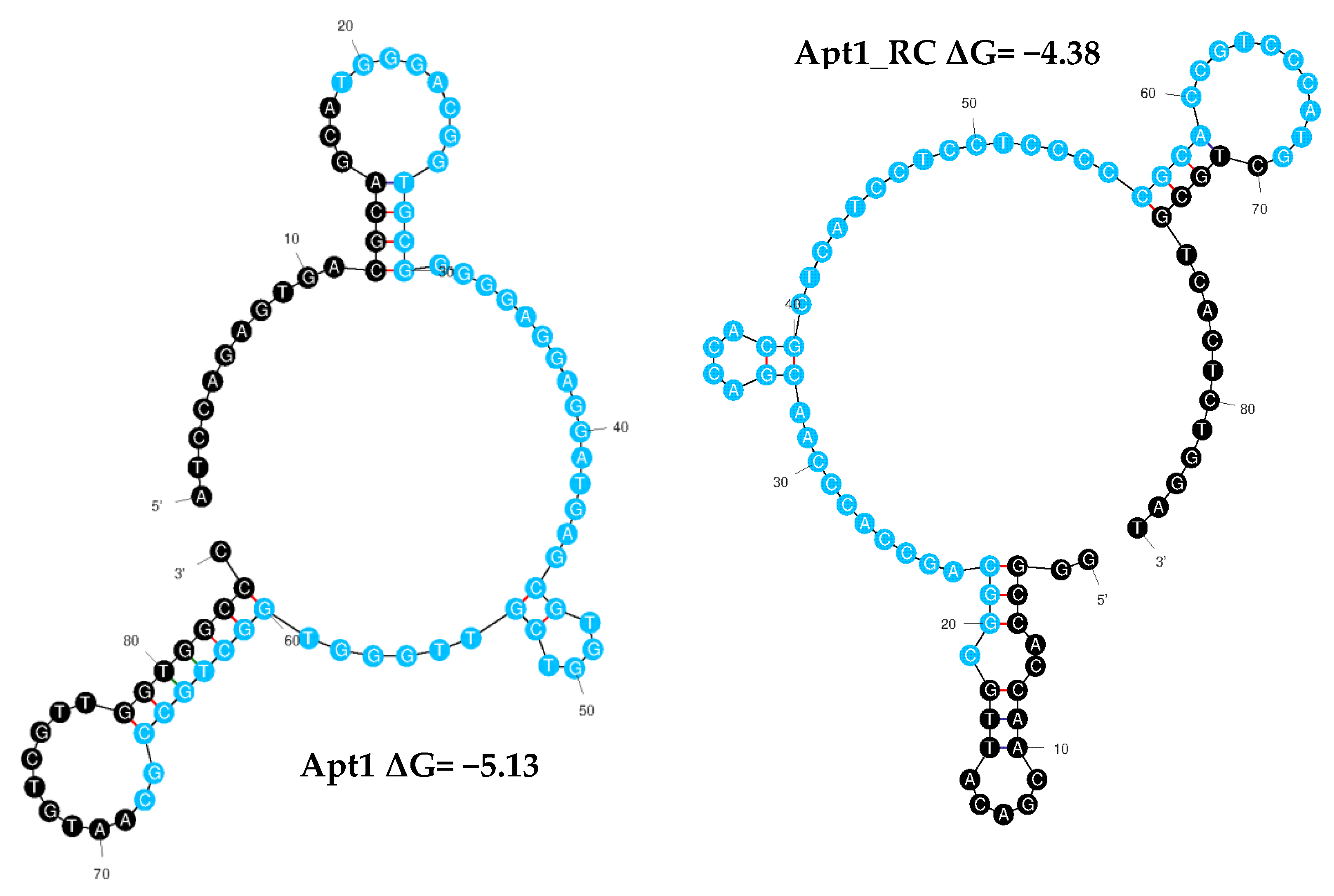
3.3. Identification of Aptamer Candidates Aided by Bioinformatics
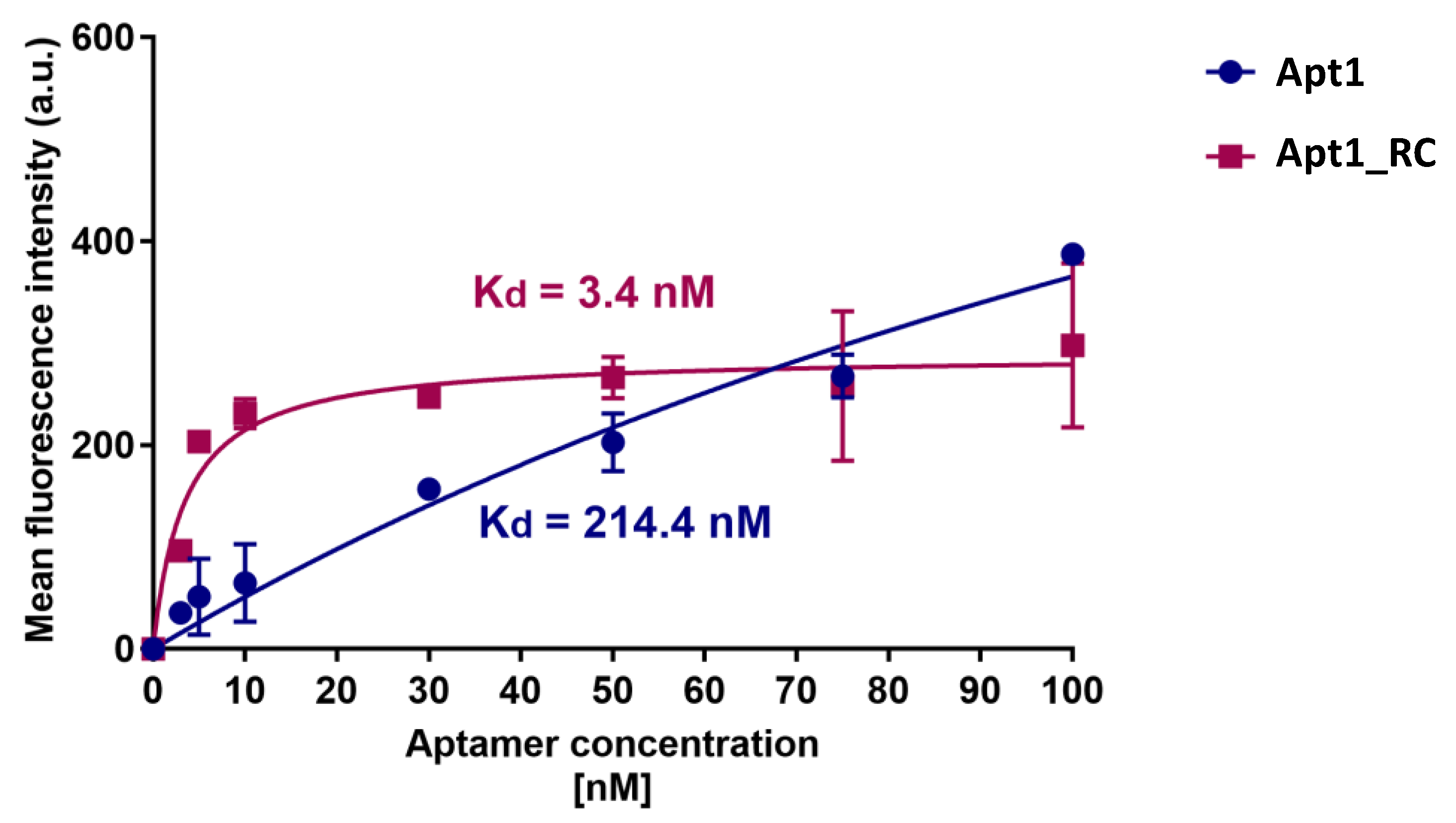
3.4. Dissociation Constants of Candidate Aptamers with UspA2
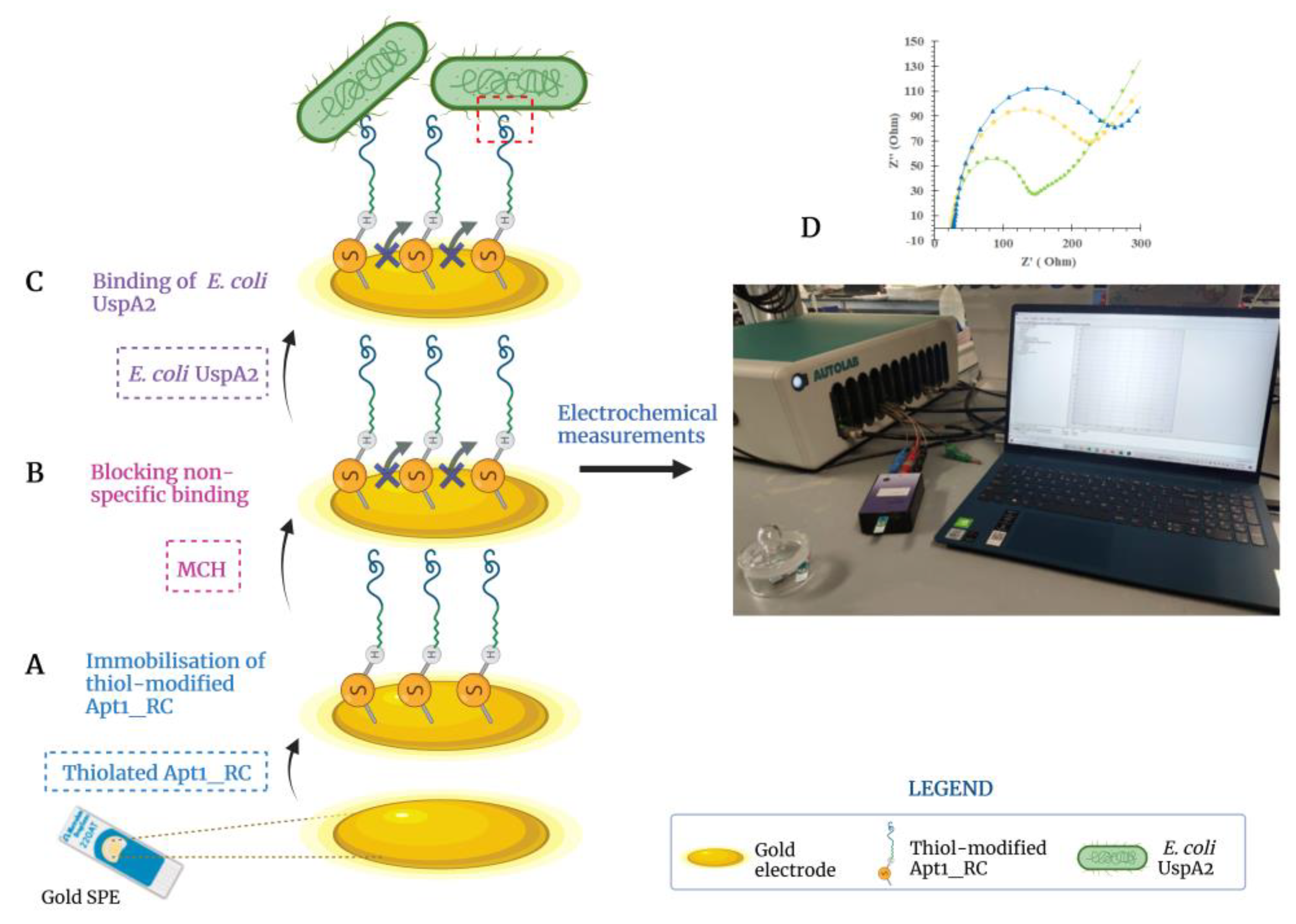
3.5. Assembly of the Aptasensor
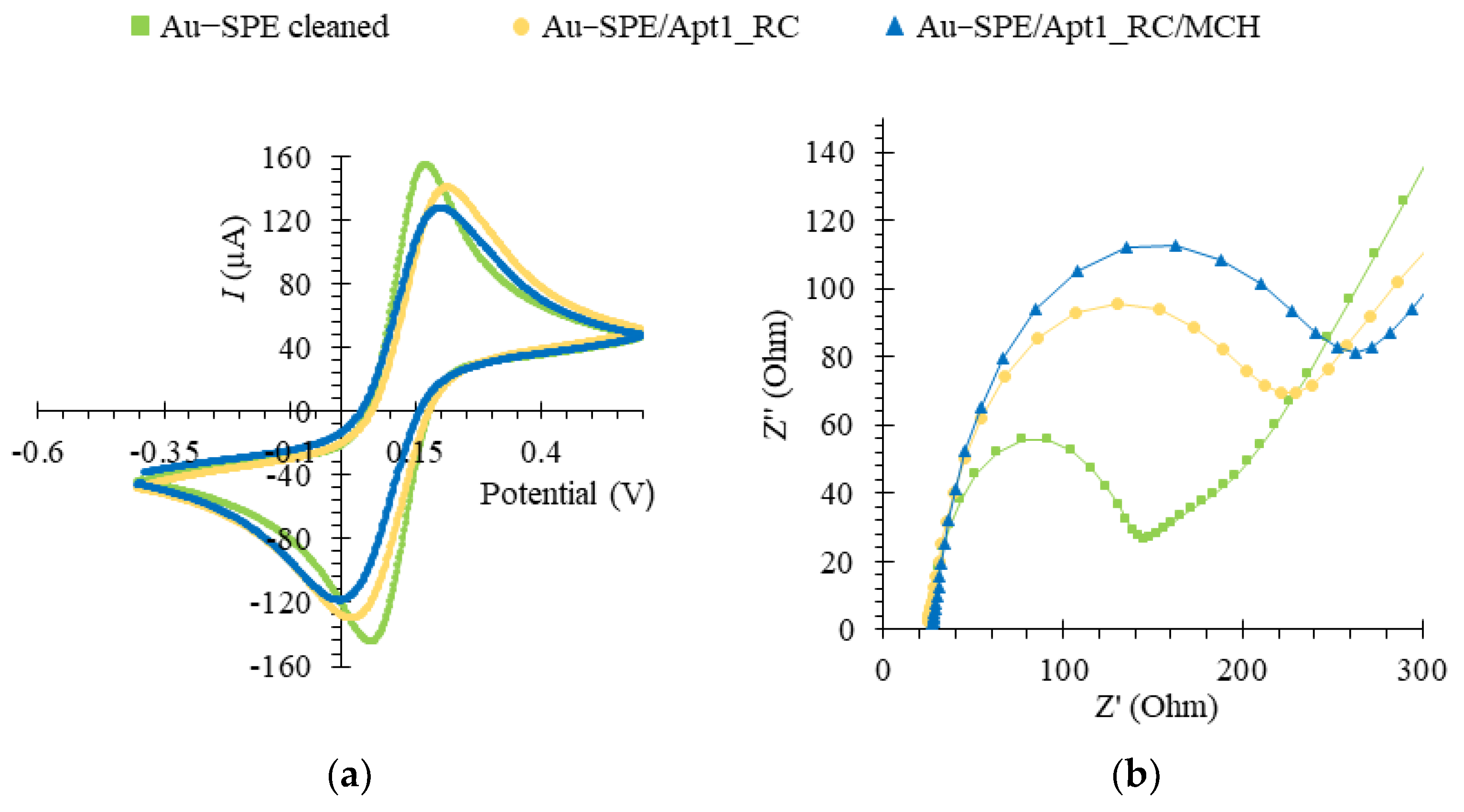
3.6. Electrochemical Characterization of the Assembled Aptasensor
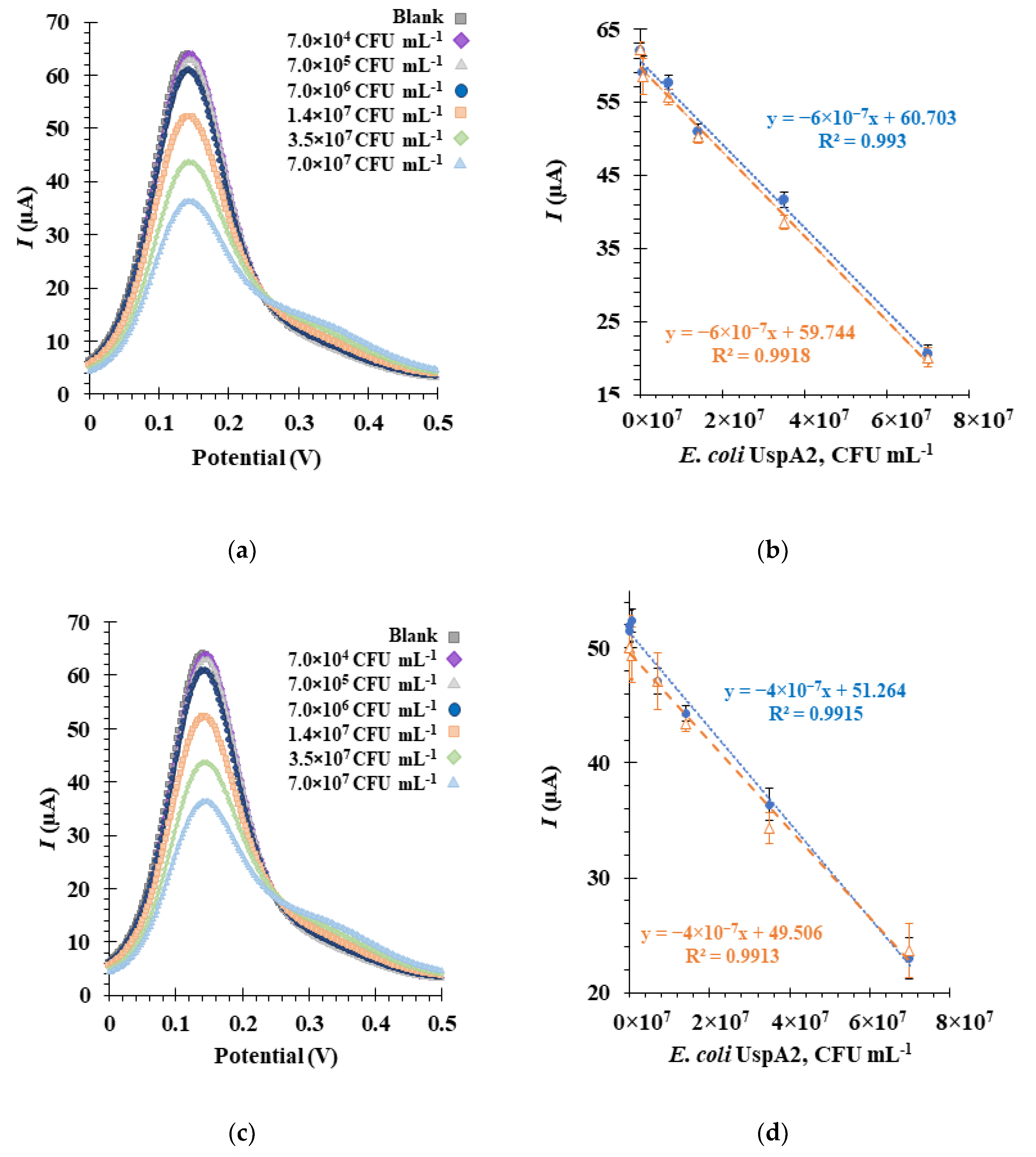
3.7. Detection of E. coli UspA2 by the Aptasensor
4. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Enright, M.C.; McKenzie, H. Moraxella (Branhamella) catarrhalis—Clinical and molecular aspects of a rediscovered pathogen. J. Med. Microbiol. 1997, 46, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.-C.; Hallström, B.M.; Bernhard, S.; Singh, B.; Riesbeck, K. Impact of sequence diversity in the Moraxella catarrhalis UspA2/UspA2H head domain on vitronectin binding and antigenic variation. Microbes Infect. 2013, 15, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Hays, J.P. Moraxella catarrhalis: A mini review. J. Pediatr. Infect. Dis. 2009, 4, 211–220. [Google Scholar] [CrossRef]
- Sillanpää, S.; Oikarinen, S.; Sipilä, M.; Kramna, L.; Rautiainen, M.; Huhtala, H.; Aittoniemi, J.; Laranne, J.; Hyöty, H.; Cinek, O. Moraxella catarrhalis Might Be More Common than Expected in Acute Otitis Media in Young Finnish Children. J. Clin. Microbiol. 2016, 54, 2373–2379. [Google Scholar] [CrossRef] [PubMed]
- Greiner, O.; Day, P.J.R.; Altwegg, M.; Nadal, D. Quantitative Detection of Moraxella catarrhalis in Nasopharyngeal Secretions by Real-Time PCR. J. Clin. Microbiol. 2003, 41, 1386–1390. [Google Scholar] [CrossRef] [PubMed]
- Kais, M.; Spindler, C.; Kalin, M.; Örtqvist, Å.; Giske, C.G. Quantitative detection of Streptococcus pneumoniae, Haemophilus influenzae, and Moraxella catarrhalis in lower respiratory tract samples by real-time PCR. Diagn. Microbiol. Infect. Dis. 2006, 55, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Schaller, A.; Troller, R.; Molina, D.; Gallati, S.; Aebi, C.; Meier, P.S. Rapid typing of Moraxella catarrhalis subpopulations based on outer membrane proteins using mass spectrometry. Proteomics 2006, 6, 172–180. [Google Scholar] [CrossRef]
- Public Health Investigation. Identification of Moraxella Species and Morphologically Similar Organisms. UK Stand Microbiology Investigation. 2015. Available online: https://www.gov.uk/government/uploads/system/uploads/attachment_data/file/401418/ID_13i3.pdf (accessed on 13 August 2020).
- Sande, M.G.; Rodrigues, J.L.; Ferreira, D.; Silva, C.J.; Rodrigues, L.R. Novel Biorecognition Elements against Pathogens in the Design of State-of-the-Art Diagnostics. Biosensors 2021, 11, 418. [Google Scholar] [CrossRef]
- Hashem, A.; Hossain, M.A.M.; Marlinda, A.R.; Al Mamun, M.; Sagadevan, S.; Shahnavaz, Z.; Simarani, K.; Johan, M.R. Nucleic acid-based electrochemical biosensors for rapid clinical diagnosis: Advances, challenges, and opportunities. Crit. Rev. Clin. Lab. Sci. 2021, 59, 156–177. [Google Scholar] [CrossRef]
- Li, H.; Liu, X.; Li, L.; Mu, X.; Genov, R.; Mason, A.J. CMOS Electrochemical Instrumentation for Biosensor Microsystems: A Review. Sensors 2017, 17, 74. [Google Scholar] [CrossRef]
- Cesewski, E.; Johnson, B.N. Electrochemical biosensors for pathogen detection. Biosens. Bioelectron. 2020, 159, 112214. [Google Scholar] [CrossRef]
- Cho, I.-H.; Kim, D.H.; Park, S. Electrochemical biosensors: Perspective on functional nanomaterials for on-site analysis. Biomater. Res. 2020, 24, 6. [Google Scholar] [CrossRef]
- Zamani, M.; Pourmadadi, M.; Ebrahimi, S.S.; Yazdian, F.; Shayeh, J.S. A novel labeled and label-free dual electrochemical detection of endotoxin based on aptamer-conjugated magnetic reduced graphene oxide-gold nanocomposite. J. Electroanal. Chem. 2022, 908, 116116. [Google Scholar] [CrossRef]
- Ramanujam, A.; Almodovar, S.; Botte, G. Ultra-Fast Electrochemical Sensor for Point-of-Care COVID-19 Diagnosis Using Non-Invasive Saliva Sampling. Processes 2021, 9, 1236. [Google Scholar] [CrossRef]
- Sande, M.G.; Ferreira, D.; Rodrigues, J.L.; Melo, L.D.R.; Linke, D.; Silva, C.J.; Moreira, F.T.C.; Sales, M.G.F.; Rodrigues, L.R. Electrochemical Aptasensor for the Detection of the Key Virulence Factor YadA of Yersinia enterocolitica. Biosensors 2022, 12, 614. [Google Scholar] [CrossRef]
- Jo, H.J.; Ryu, J.S.; Robby, A.I.; Kim, Y.S.; Chung, H.J.; Park, S.Y. Rapid and selective electrochemical sensing of bacterial pneumonia in human sputum based on conductive polymer dot electrodes. Sens. Actuators B Chem. 2022, 368, 132084. [Google Scholar] [CrossRef]
- Pandey, R.; Lu, Y.; Osman, E.; Saxena, S.; Zhang, Z.; Qian, S.; Pollinzi, A.; Smieja, M.; Li, Y.; Soleymani, L.; et al. DNAzyme-Immobilizing Microgel Magnetic Beads Enable Rapid, Specific, Culture-Free, and Wash-Free Electrochemical Quantification of Bacteria in Untreated Urine. ACS Sens. 2022, 7, 985–994. [Google Scholar] [CrossRef] [PubMed]
- Hoiczyk, E.; Roggenkamp, A.; Reichenbecher, M.; Lupas, A.; Heesemann, J. Structure and sequence analysis of Yersinia YadA and Moraxella UspAs reveal a novel class of adhesins. EMBO J. 2000, 19, 5989–5999. [Google Scholar] [CrossRef]
- Conners, R.; Hill, D.J.; Borodina, E.; Agnew, C.; Daniell, S.J.; Burton, N.M.; Sessions, R.B.; Clarke, A.R.; E Catto, L.; Lammie, D.; et al. The Moraxella adhesin UspA1 binds to its human CEACAM1 receptor by a deformable trimeric coiled-coil. EMBO J. 2008, 27, 1779–1789. [Google Scholar] [CrossRef] [PubMed]
- Linke, D.; Riess, T.; Autenrieth, I.B.; Lupas, A.; Kempf, V.A. Trimeric autotransporter adhesins: Variable structure, common function. Trends Microbiol. 2006, 14, 264–270. [Google Scholar] [CrossRef]
- Su, Y.-C.; Singh, B.; Riesbeck, K. Moraxella catarrhalis: From interactions with the host immune system to vaccine development. Futur. Microbiol. 2012, 7, 1073–1100. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.M.; Lafontaine, E.R.; Wagner, N.J.; Geme, J.W.S.; Hansen, E.J. A hag Mutant of Moraxella catarrhalis Strain O35E Is Deficient in Hemagglutination, Autoagglutination, and Immunoglobulin D-Binding Activities. Infect. Immun. 2002, 70, 4523–4533. [Google Scholar] [CrossRef] [PubMed]
- Meier, P.S.; Freiburghaus, S.; Martin, A.; Heiniger, N.; Troller, R.; Aebi, C. Mucosal immune response to specific outer membrane proteins of Moraxella catarrhalis in young children. Pediatr. Infect. Dis. J. 2003, 22, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Meier, P.S.; Heiniger, N.; Troller, R.; Aebi, C. SalivaryAntibodies Directed against Outer Membrane Proteins of Moraxella catarrhalis in Healthy Adults. Infect. Immun. 2003, 71, 6793–6798. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.L.; Ferreira, D.; Rodrigues, L.R. Synthetic biology strategies towards the development of new bioinspired technologies for medical applications. In Bioinspired Materials for Medical Applications; Rodrigues, L., Mota, M., Eds.; Elsevier: Oxford, UK, 2017; pp. 451–497. [Google Scholar] [CrossRef]
- Kaur, H.; Bruno, J.G.; Kumar, A.; Sharma, T.K. Aptamers in the Therapeutics and Diagnostics Pipelines. Theranostics 2018, 8, 4016–4032. [Google Scholar] [CrossRef] [PubMed]
- Sola, M.; Menon, A.P.; Moreno, B.; Meraviglia-Crivelli, D.; Soldevilla, M.M.; Cartón-García, F.; Pastor, F. Aptamers Against Live Targets: Is In Vivo SELEX Finally Coming to the Edge? Mol. Ther. Nucleic Acids 2020, 21, 192–204. [Google Scholar] [CrossRef]
- Liu, L.; Han, Z.; An, F.; Gong, X.; Zhao, C.; Zheng, W.; Mei, L.; Zhou, Q. Aptamer-based biosensors for the diagnosis of sepsis. J. Nanobiotechnology 2021, 19, 216. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, L.; Huang, Q.; Tong, T.; Zhou, Y.; Li, Z.; Bai, Q.; Liang, H.; Chen, L. Recent advances on aptamer-based biosensors for detection of pathogenic bacteria. World J. Microbiol. Biotechnol. 2021, 37, 45. [Google Scholar] [CrossRef]
- Beyer, H.; Gonschorek, P.; Samodelov, S.L.; Meier, M.; Weber, W.; Zurbriggen, M.D. AQUA Cloning: A Versatile and Simple Enzyme-Free Cloning Approach. PLoS ONE 2015, 10, e0137652. [Google Scholar] [CrossRef]
- Cardoso, A.R.; Moreira, F.T.; Fernandes, R.; Sales, M.G.F. Novel and simple electrochemical biosensor monitoring attomolar levels of miRNA-155 in breast cancer. Biosens. Bioelectron. 2016, 80, 621–630. [Google Scholar] [CrossRef]
- Harvey, D. Modern Analytic Chemistry; McGraw-Hill Co.: New York, NY, USA, 2000; pp. 1–798. [Google Scholar]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial autoaggregation. AIMS Microbiol. 2018, 4, 140–164. [Google Scholar] [CrossRef] [PubMed]
- Aebi, C.; Lafontaine, E.R.; Cope, L.D.; Latimer, J.L.; Lumbley, S.L.; McCracken, G.H.; Hansen, E.J. Phenotypic Effect of Isogenic uspA1 and uspA2 Mutations on Moraxella catarrhalis 035E. Infect. Immun. 1998, 66, 3113–3119. [Google Scholar] [CrossRef] [PubMed]
- Cope, L.D.; Lafontaine, E.R.; Slaughter, C.A.; Hasemann, C.A.; Aebi, C.; Henderson, F.W.; McCracken, G.H.; Hansen, E.J. Characterization of the Moraxella catarrhalis uspA1 and uspA2 Genes and Their Encoded Products. J. Bacteriol. 1999, 181, 4026–4034. [Google Scholar] [CrossRef] [PubMed]
- Grosskinsky, U.; Schütz, M.; Fritz, M.; Schmid, Y.; Lamparter, M.C.; Szczesny, P.; Lupas, A.N.; Autenrieth, I.B.; Linke, D. A Conserved Glycine Residue of Trimeric Autotransporter Domains Plays a Key Role in Yersinia Adhesin A Autotransport. J. Bacteriol. 2007, 189, 9011–9019. [Google Scholar] [CrossRef] [PubMed]
- Riess, T.; Raddatz, G.; Linke, D.; Schäfer, A.; Kempf, V.A.J. Analysis of Bartonella Adhesin A Expression Reveals Differences between Various B. henselae Strains. Infect. Immun. 2007, 75, 35–43. [Google Scholar] [CrossRef]
- Kim, H.R.; Song, M.Y.; Kim, B.C. Rapid isolation of bacteria-specific aptamers with a non-SELEX-based method. Anal. Biochem. 2019, 591, 113542. [Google Scholar] [CrossRef]
- Song, S.; Wang, X.; Xu, K.; Li, Q.; Ning, L.; Yang, X. Selection of highly specific aptamers to Vibrio parahaemolyticus using cell-SELEX powered by functionalized graphene oxide and rolling circle amplification. Anal. Chim. Acta 2019, 1052, 153–162. [Google Scholar] [CrossRef]
- Boel, E.; Bootsma, H.; de Kruif, J.; Jansze, M.; Klingman, K.L.; van Dijk, H.; Logtenberg, T. Phage Antibodies Obtained by Competitive Selection on Complement-Resistant Moraxella (Branhamella) catarrhalis Recognize the High-Molecular-Weight Outer Membrane Protein. Infect. Immun. 1998, 66, 83–88. [Google Scholar] [CrossRef]
- Ferreira, D.; Barbosa, J.; Sousa, D.A.; Silva, C.; Melo, L.D.R.; Avci-Adali, M.; Wendel, H.P.; Rodrigues, L.R. Selection of aptamers against triple negative breast cancer cells using high throughput sequencing. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Kolm, C.; Cervenka, I.; Aschl, U.J.; Baumann, N.; Jakwerth, S.; Krska, R.; Mach, R.L.; Sommer, R.; DeRosa, M.C.; Kirschner, A.K.T.; et al. DNA aptamers against bacterial cells can be efficiently selected by a SELEX process using state-of-the art qPCR and ultra-deep sequencing. Sci. Rep. 2020, 10, 8614. [Google Scholar] [CrossRef]
- Hoinka, J.; Berezhnoy, A.; Dao, P.; Sauna, Z.E.; Gilboa, E.; Przytycka, T.M. Large scale analysis of the mutational landscape in HT-SELEX improves aptamer discovery. Nucleic Acids Res. 2015, 43, 5699–5707. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, H.; Savory, N.; Abe, K.; Ikebukuro, K. Methods for Improving Aptamer Binding Affinity. Molecules 2016, 21, 421. [Google Scholar] [CrossRef] [PubMed]
- Lavu, P.S.; Mondal, B.; Ramlal, S. Selection and Characterization of Cell Surface Specific Aptamer and Development of Fluorescence Assay for Detection of Shigella flexneri from Water Samples. J. Fluoresc. 2021, 31, 685–693. [Google Scholar] [CrossRef]
- Nguyen, T.T.-Q.; Kim, E.R.; Gu, M.B. A new cognate aptamer pair-based sandwich-type electrochemical biosensor for sensitive detection of Staphylococcus aureus. Biosens. Bioelectron. 2022, 198, 113835. [Google Scholar] [CrossRef]
- Gupta, R.; Kumar, A.; Kumar, S.; Pinnaka, A.K.; Singhal, N.K. Naked eye colorimetric detection of Escherichia coli using aptamer conjugated graphene oxide enclosed Gold nanoparticles. Sens. Actuators B Chem. 2020, 329, 129100. [Google Scholar] [CrossRef]
- Wu, H.; Gu, L.; Ma, X.; Tian, X.; Fan, S.; Qin, M.; Lu, J.; Lyu, M.; Wang, S. Rapid Detection of Helicobacter pylori by the Naked Eye Using DNA Aptamers. ACS Omega 2021, 6, 3771–3779. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Cao, C.; Bi, W.; Lin, J.; Tan, L.; Gan, N. On-site enrichment and detection of live Salmonella typhimurium using a bioluminescence sensor coupled with a hyperbranched aptamer probe-labelled stir-bars array. Sens. Actuators B Chem. 2022, 364, 131862. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, S.; Zhao, M.; Zhang, P.; Xin, Z.; Tao, J.; Bai, L. Amperometric DNA biosensor for Mycobacterium tuberculosis detection using flower-like carbon nanotubes-polyaniline nanohybrid and enzyme-assisted signal amplification strategy. Biosens. Bioelectron. 2018, 119, 215–220. [Google Scholar] [CrossRef]
- Alatraktchi, F.A.; Dimaki, M.; Støvring, N.; Johansen, H.K.; Molin, S.; Svendsen, W.E. Nanograss sensor for selective detection of Pseudomonas aeruginosa by pyocyanin identification in airway samples. Anal. Biochem. 2020, 593, 113586. [Google Scholar] [CrossRef]
- Güner, A.; Çevik, E.; Şenel, M.; Alpsoy, L. An electrochemical immunosensor for sensitive detection of Escherichia coli O157:H7 by using chitosan, MWCNT, polypyrrole with gold nanoparticles hybrid sensing platform. Food Chem. 2017, 229, 358–365. [Google Scholar] [CrossRef]
- Kaya, H.O.; Cetin, A.E.; Azimzadeh, M.; Topkaya, S.N. Pathogen detection with electrochemical biosensors: Advantages, challenges and future perspectives. J. Electroanal. Chem. 2021, 882, 114989. [Google Scholar] [CrossRef]
- Angell, D.; Dickinson, T. The kinetics of the ferrous/ferric and ferro/ferricyanide reactions at platinum and gold electrodes. J. Electroanal. Chem. Interfacial Electrochem. 1972, 35, 55–72. [Google Scholar] [CrossRef]
- Cheng, A.K.H.; Ge, B.; Yu, H.-Z. Aptamer-Based Biosensors for Label-Free Voltammetric Detection of Lysozyme. Anal. Chem. 2007, 79, 5158–5164. [Google Scholar] [CrossRef] [PubMed]
- Bard, A.J.; Faulkner, L. Electrochemical Methods: Fundamentals and Applications, 2nd ed.; Wiley: New York, NY, USA, 2000. [Google Scholar]
- Mirceski, V.; Gulaboski, R.; Lovric, M.; Bogeski, I.; Kappl, R.; Hoth, M. Square-Wave Voltammetry: A Review on the Recent Progress. Electroanalysis 2013, 25, 2411–2422. [Google Scholar] [CrossRef]
- Figueroa-Miranda, G.; Feng, L.; Shiu, S.C.-C.; Dirkzwager, R.M.; Cheung, Y.-W.; Tanner, J.A.; Schöning, M.J.; Offenhäusser, A.; Mayer, D. Aptamer-based electrochemical biosensor for highly sensitive and selective malaria detection with adjustable dynamic response range and reusability. Sens. Actuators B Chem. 2018, 255, 235–243. [Google Scholar] [CrossRef]
- Lee, U.; Kim, Y.H.; Yoon, K.S.; Kim, Y. Selective Butyrate Esterase Probe for the Rapid Colorimetric and Fluorogenic Identification of Moraxella catarrhalis. Anal. Chem. 2020, 92, 16051–16057. [Google Scholar] [CrossRef] [PubMed]
- Sande, M.G.; Roque, L.; Braga, A.; Marques, M.; Ferreira, D.; Saragliadis, A.; Rodrigues, J.L.; Linke, D.; Ramada, D.; Silva, C.; et al. Design of new hydrolyzed collagen-modified magnetic nanoparticles to capture pathogens. J. Biomed. Mater. Res. Part B Appl. Biomater. 2022, 111, 354–365. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cycles | E. coli UspA2 (µL) | Incubation (Minutes) | Washes (after Selection) | PCR (Cycles) |
|---|---|---|---|---|
| 1 | 500 | 60 | 2 | 30 |
| 2 | 500 | 50 | 2 | 25 |
| 3 | 400 | 45 | 3 | 25 |
| 4 | 400 | 40 | 3 | 18 |
| 5 | 400 (counter-selection) | 40 | - | 20 |
| 6 | 350 | 35 | 4 | 22 |
| 7 | 350 (counter-selection) | 35 | - | 13 |
| 8 | 350 | 30 | 4 | 15 |
| Aptamer | Sequences (5′–3′) | Copies |
|---|---|---|
| Apt1 | TGGGACGGTGCGGGGGAGGAGGATGAGCGTGGTCGTTGGGTGGCTGCCGC | 16,130 |
| Apt1_RC | GCGGCAGCCACCCAACGACCACGCTCATCCTCCTCCCCCGCACCGTCCCA | |
| Apt2 | TGTGGGGGCATGGGAAGGTGGGTAAGCCGTTGCGTGGGATGTGGCGGGTC | 13,652 |
| Apt2_RC | GACCCGCCACATCCCACGCAACGGCTTACCCACCTTCCCATGCCCCCACA | |
| Apt3 | TGGGGCGAAGGACGGTGGATGTGTGTAGGCGCGGGTGGGTCGCACGGGGT | 12,992 |
| Apt3_RC | ACCCCGTGCGACCCACCCGCGCCTACACACATCCACCGTCCTTCGCCCCA | |
| Apt4 | TGACAGACGGGAGAGACACGACGGGGGCGGGGAGTGAGAAGCGCGCGCTG | 12,694 |
| Apt4_RC | CAGCGCGCGCTTCTCACTCCCCGCCCCCGTCGTGTCTCTCCCGTCTGTCA | |
| Apt5 | TGTGGCGGTGGCAAGGTCGGTGGGTGCGGCGGCGTACCGGTGGTGGGGGG | 9598 |
| Apt5_RC | CCCCCCACCACCGGTACGCCGCCGCACCCACCGACCTTGCCACCGCCACA | |
| Apt6 | TGACCGGCGGGGGGTGGTTCGCTGATAGGGCGTGTGGACCGGGGGTCGCA | 9107 |
| Apt6_RC | TGCGACCCCCGGTCCACACGCCCTATCAGCGAACCACCCCCCGCCGGTCA | |
| Apt7 | TGGGCGGGTAGTGGGGCAACGTGAGCGCGTGGGGTGTGGCAGGCGTGGCT | 8004 |
| Apt7_RC | AGCCACGCCTGCCACACCCCACGCGCTCACGTTGCCCCACTACCCGCCCA | |
| Apt8 | AGATGACTTAGGGTGACTGGAGTTCAGACGTGTGCTCTTCCGATCT | 7558 |
| Apt8_RC | AGATCGGAAGAGCACACGTCTGAACTCCAGTCACCCTAAGTCATCT | |
| Apt9 | AGATCGGAAGAGCGTCGTGTAGGGAAAGAGTGTGTCATCGAGTGTA | 7229 |
| Apt9_RC | TACACTCGATGACACACTCTTTCCCTACACGACGCTCTTCCGATCT | |
| Apt10 | TGGGTGTGGTGGCGCGGGTGGCGTGGTGCGTGTACAGCTGGTTGCGGGCG | 7129 |
| Apt10_RC | CGCCCGCAACCAGCTGTACACGCACCACGCCACCCGCGCCACCACACCCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sande, M.G.; Ferreira, D.; Rodrigues, J.L.; Melo, L.D.R.; Saragliadis, A.; Linke, D.; Moreira, F.T.C.; Sales, M.G.F.; Rodrigues, L.R. Aptasensor for the Detection of Moraxella catarrhalis Adhesin UspA2. Bioengineering 2023, 10, 178. https://doi.org/10.3390/bioengineering10020178
Sande MG, Ferreira D, Rodrigues JL, Melo LDR, Saragliadis A, Linke D, Moreira FTC, Sales MGF, Rodrigues LR. Aptasensor for the Detection of Moraxella catarrhalis Adhesin UspA2. Bioengineering. 2023; 10(2):178. https://doi.org/10.3390/bioengineering10020178
Chicago/Turabian StyleSande, Maria G., Débora Ferreira, Joana L. Rodrigues, Luís D. R. Melo, Athanasios Saragliadis, Dirk Linke, Felismina T. C. Moreira, Maria Goreti F. Sales, and Ligia R. Rodrigues. 2023. "Aptasensor for the Detection of Moraxella catarrhalis Adhesin UspA2" Bioengineering 10, no. 2: 178. https://doi.org/10.3390/bioengineering10020178
APA StyleSande, M. G., Ferreira, D., Rodrigues, J. L., Melo, L. D. R., Saragliadis, A., Linke, D., Moreira, F. T. C., Sales, M. G. F., & Rodrigues, L. R. (2023). Aptasensor for the Detection of Moraxella catarrhalis Adhesin UspA2. Bioengineering, 10(2), 178. https://doi.org/10.3390/bioengineering10020178