
Dynamics of Nocturnal Evapotranspiration in a Dry Region of the Chinese Loess Plateau: A Multi-Timescale Analysis

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
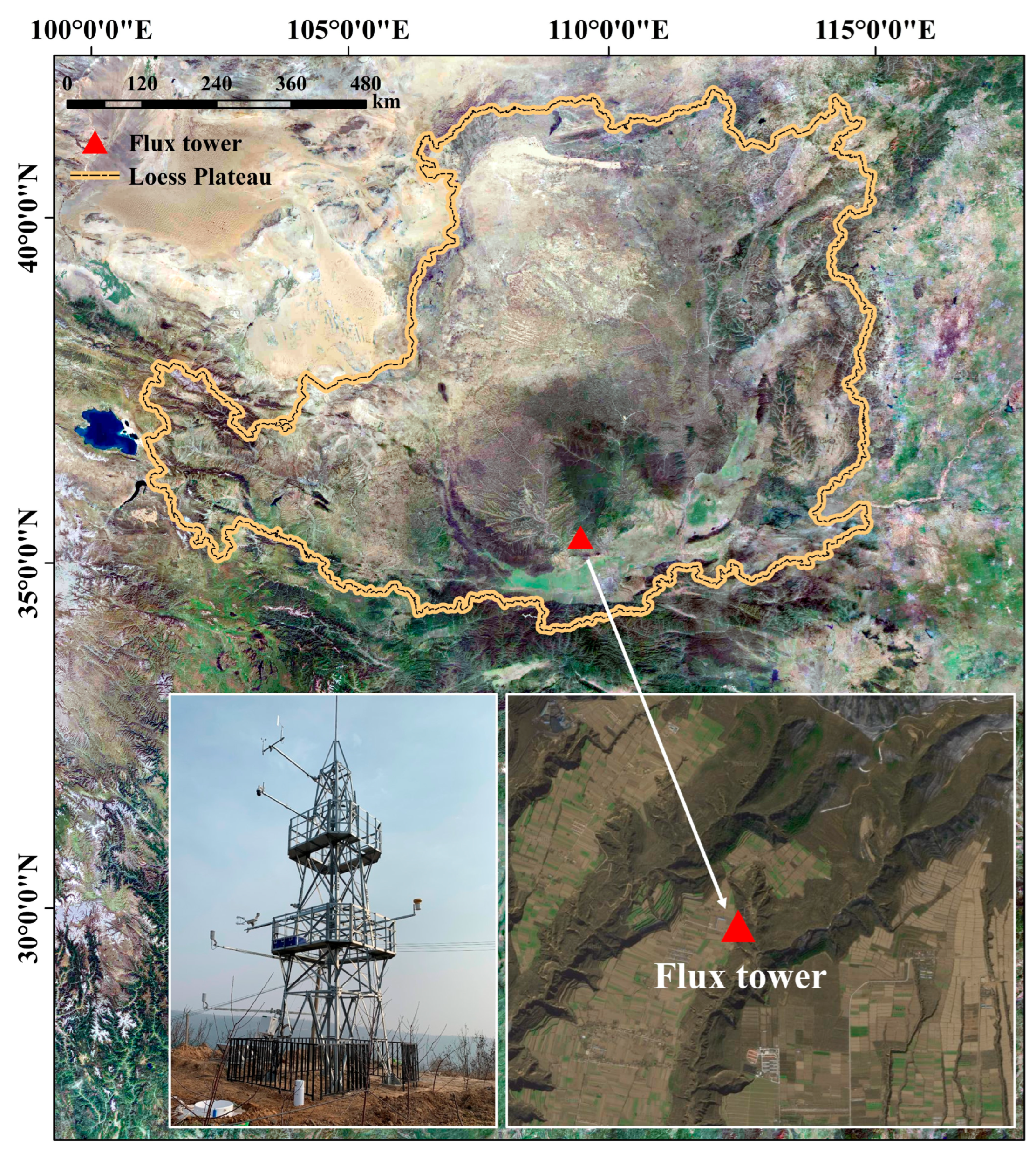
2.1. Site Description
2.2. Flux and Environmental Measurements
2.3. Data Processing
2.4. Methods
2.5. Statistical Analysis
3. Results
3.1. Variations in Environmental Variables
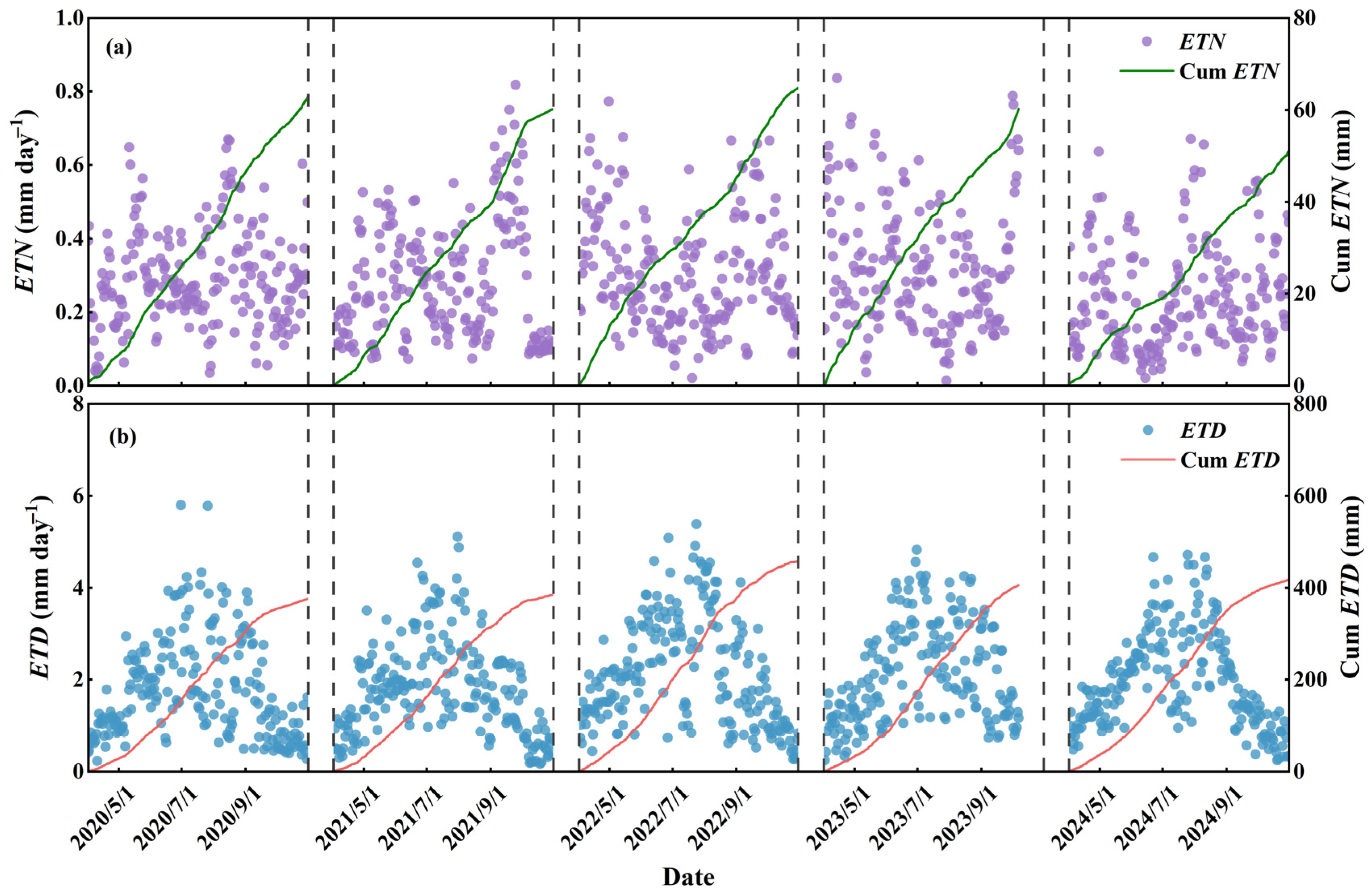
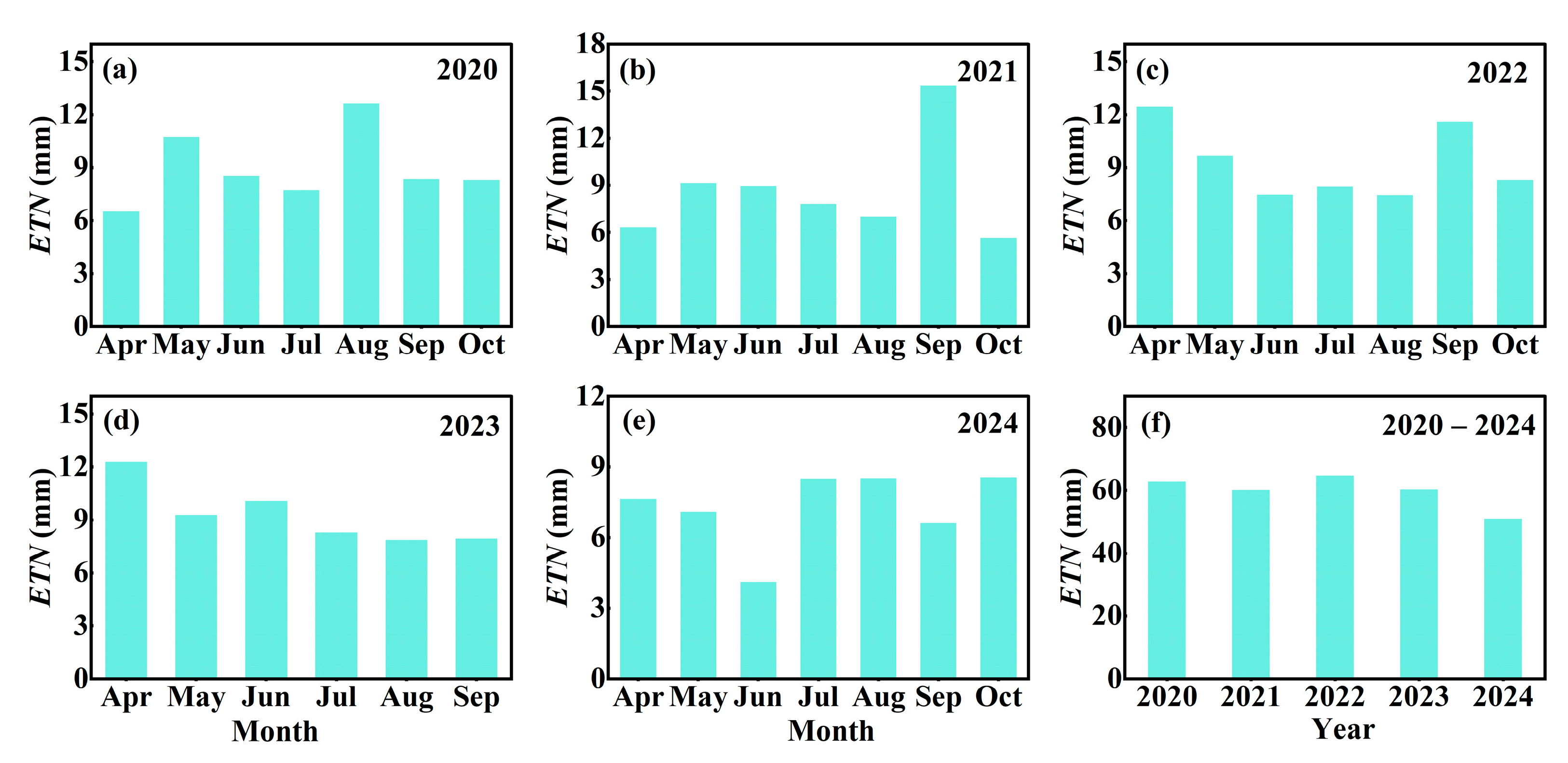
3.2. Variations in ETN on Multiple Time Scales
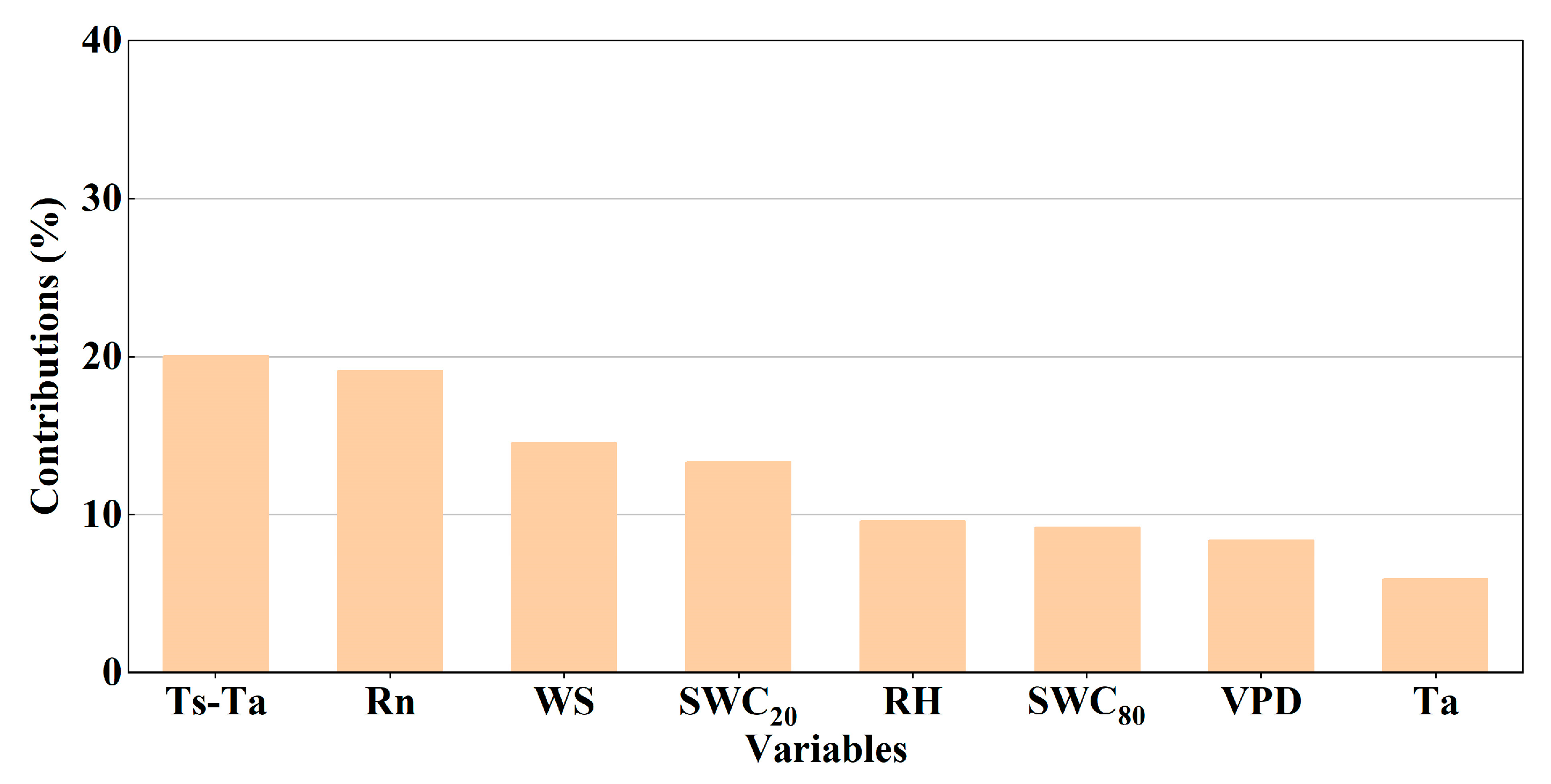
3.3. Environmental Factors Affecting ETN
4. Discussion
4.1. Uncertainties in Observations
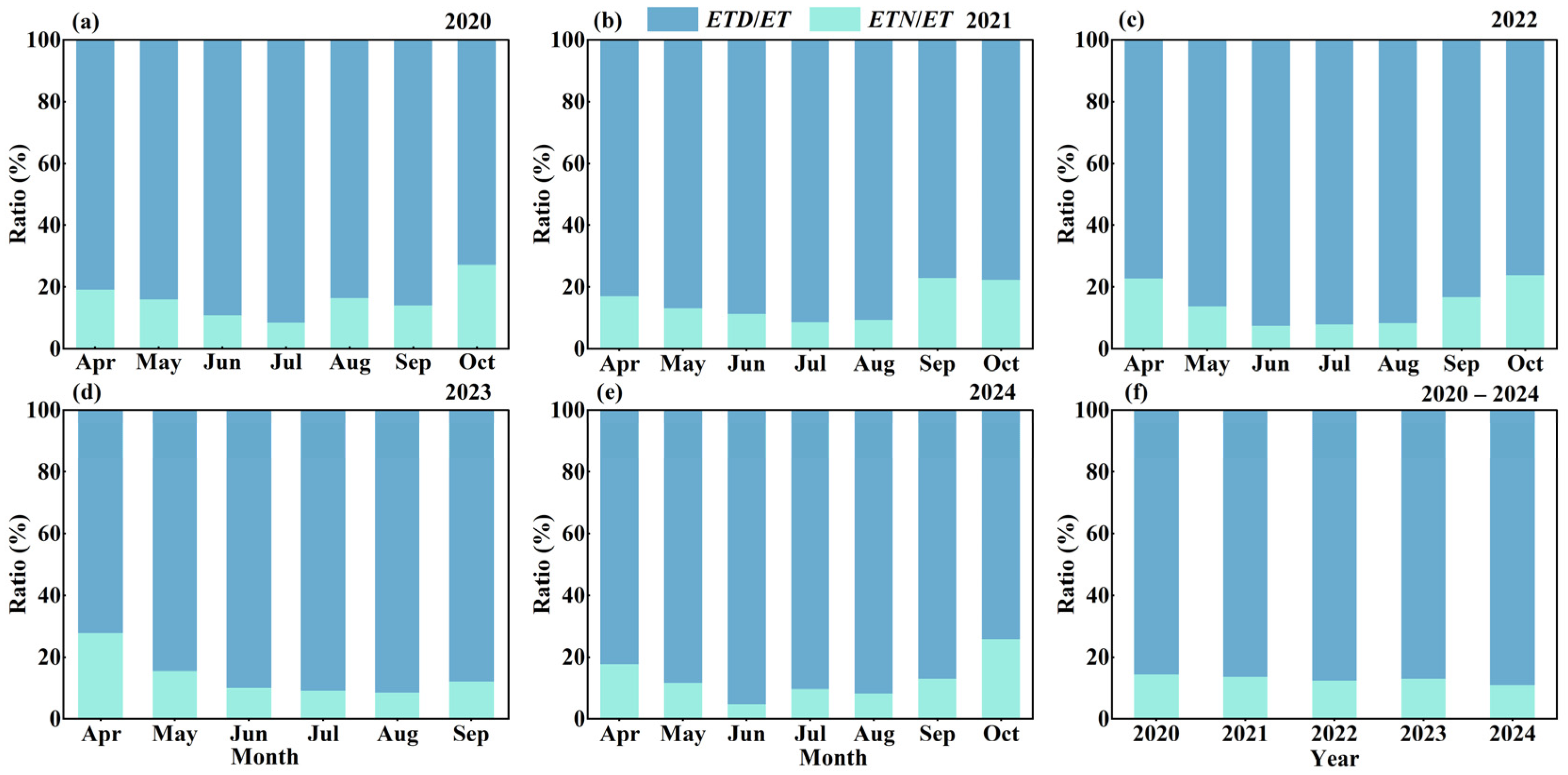
4.2. Changes in ETN and Its Proportion to ET (or ETD)
4.3. Effects of Environmental Factors on ETN
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2020 | 2021 | 2022 | 2023 | 2024 | Total | |
|---|---|---|---|---|---|---|
| Nighttime (%) | 36.0 | 31.2 | 51.6 | 33.1 | 47.2 | 40.0 |
| Daytime (%) | 52.7 | 51.8 | 72.8 | 49.1 | 69.1 | 59.3 |
| Total (%) | 45.3 | 42.6 | 63.4 | 42.1 | 59.4 | 50.8 |
| Variable | Year | Growing Season | Mean or Summation | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Apr. | May | Jun. | Jul. | Aug. | Sep. | Oct. | |||
| Ta (°C) | 2020 | 10.2 | 16.0 | 18.6 | 18.9 | 18.5 | 15.0 | 8.1 | 15.0 ± 4.9 |
| 2021 | 8.4 | 15.2 | 19.5 | 20.3 | 18.8 | 16.2 | 8.1 | 15.2 ± 5.6 | |
| 2022 | 11.4 | 14.3 | 20.3 | 20.5 | 20.7 | 15.3 | 9.5 | 15.9 ± 5.4 | |
| 2023 | 9.6 | 13.5 | 19.4 | 21.1 | 20.7 | / | / | 16.2 ± 5.9 | |
| 2024 | 12.6 | 16.4 | / | 20.8 | 20.8 | 17.5 | 10.5 | 16.3 ± 4.8 | |
| Ts-Ta (°C) | 2020 | −3.5 | −3.1 | −1.8 | −1.3 | −1.0 | −1.7 | −1.3 | −1.9 ± 1.6 |
| 2021 | −1.7 | −3.2 | −2.1 | −1.6 | −1.5 | −1.6 | −1.2 | −1.8 ± 1.5 | |
| 2022 | −2.7 | −2.3 | −2.0 | −1.0 | −0.7 | −1.5 | −1.3 | −1.6 ± 1.2 | |
| 2023 | −1.6 | −1.4 | −1.5 | −1.3 | −1.0 | / | / | −1.4 ± 1.0 | |
| 2024 | −1.8 | −1.7 | / | −0.6 | −0.8 | −0.5 | −0.9 | −1.1 ± 0.8 | |
| RH (%) | 2020 | 44.4 | 49.7 | 68.0 | 82.4 | 86.5 | 75.2 | 72.3 | 68.4 ± 23.9 |
| 2021 | 68.3 | 55.8 | 63.0 | 79.0 | 76.1 | 79.9 | 83.2 | 72.2 ± 20.6 | |
| 2022 | 48.9 | 59.9 | 57.0 | 76.9 | 81.3 | 71.8 | 69.8 | 66.6 ± 21.1 | |
| 2023 | 59.1 | 67.1 | 60.6 | 65.7 | 77.7 | / | / | 65.0 ± 20.3 | |
| 2024 | 60.5 | 58.6 | / | 85.6 | 81.6 | 85.1 | 73.8 | 73.8 ± 18.5 | |
| VPD (hPa) | 2020 | 10.5 | 14.1 | 13.4 | 10.0 | 9.5 | 10.5 | 8.5 | 10.9 ± 4.6 |
| 2021 | 8.5 | 14.9 | 16.8 | 13.3 | 13.6 | / | / | 13.2 ± 5.2 | |
| 2022 | 11.7 | 11.4 | 16.7 | 13.8 | 13.5 | 12.4 | 10.0 | 12.8 ± 4.8 | |
| 2023 | 12.3 | 13.8 | 18.5 | 19.6 | 14.4 | / | / | 15.7 ± 5.8 | |
| 2024 | 10.8 | 14.3 | / | 9.2 | 11.0 | 9.6 | 8.1 | 10.6 ± 3.4 | |
| WS (m s−1) | 2020 | 2.7 | 3.1 | 2.8 | 2.9 | 3.2 | 3.0 | 2.4 | 2.8 ± 1.4 |
| 2021 | 3.2 | 2.8 | 2.5 | 2.8 | 2.8 | 2.3 | 2.4 | 2.7 ± 1.4 | |
| 2022 | 3.1 | 2.7 | 2.9 | 3.3 | 3.6 | 2.8 | 2.7 | 3.0 ± 1.4 | |
| 2023 | 3.1 | 2.8 | 2.6 | 3.0 | 2.7 | / | / | 2.9 ± 1.3 | |
| 2024 | 2.8 | 2.7 | / | 2.0 | 2.7 | 4.1 | 2.9 | 2.9 ± 1.5 | |
| SWC20 (cm3 cm−3) | 2020 | 0.21 | 0.24 | 0.25 | 0.23 | 0.28 | 0.19 | 0.26 | 0.24 ± 0.05 |
| 2021 | 0.28 | 0.31 | 0.28 | 0.28 | 0.18 | 0.31 | 0.32 | 0.28 ± 0.05 | |
| 2022 | 0.22 | 0.21 | 0.12 | 0.22 | 0.13 | 0.20 | 0.22 | 0.19 ± 0.06 | |
| 2023 | 0.23 | 0.19 | 0.14 | 0.12 | 0.24 | / | / | 0.18 ± 0.06 | |
| 2024 | 0.18 | 0.10 | / | 0.26 | 0.16 | 0.09 | 0.10 | 0.15 ± 0.07 | |
| SWC40 (cm3 cm−3) | 2020 | 0.32 | 0.33 | 0.34 | 0.34 | 0.35 | 0.30 | 0.30 | 0.33 ± 0.02 |
| 2021 | 0.35 | 0.35 | 0.31 | 0.33 | 0.28 | 0.34 | 0.40 | 0.34 ± 0.04 | |
| 2022 | 0.37 | 0.36 | 0.27 | 0.31 | 0.26 | 0.27 | 0.29 | 0.30 ± 0.06 | |
| 2023 | 0.34 | 0.33 | 0.30 | 0.25 | 0.35 | / | / | 0.31 ± 0.04 | |
| 2024 | 0.33 | 0.25 | / | 0.33 | 0.28 | 0.21 | 0.19 | 0.26 ± 0.06 | |
| SWC80 (cm3 cm−3) | 2020 | 0.26 | 0.26 | 0.27 | 0.27 | 0.29 | 0.26 | 0.24 | 0.27 ± 0.02 |
| 2021 | 0.26 | 0.30 | 0.30 | 0.30 | 0.28 | 0.27 | 0.32 | 0.29 ± 0.03 | |
| 2022 | 0.31 | 0.30 | 0.24 | 0.19 | 0.19 | 0.18 | 0.18 | 0.23 ± 0.05 | |
| 2023 | 0.21 | 0.24 | 0.24 | 0.20 | 0.24 | / | / | 0.22 ± 0.02 | |
| 2024 | 0.27 | 0.23 | / | 0.23 | 0.27 | 0.20 | 0.16 | 0.23 ± 0.05 | |
| P (mm) | 2020 | 1.5 | 36.0 | 18.9 | 16.4 | 81.7 | 21.3 | 28.5 | 204.3 ± 4.2 |
| 2021 | 17.7 | 67.8 | 20.6 | 19.4 | 16.9 | 84.7 | 62.3 | 289.4 ± 4.0 | |
| 2022 | 24.3 | 26.0 | 14.2 | 91.6 | 24.6 | 15.0 | 15.7 | 211.4 ± 3.3 | |
| 2023 | 24.5 | 13.0 | 47.8 | 15.4 | / | / | / | 100.7 ± 2.3 | |
| 2024 | 17.6 | 1.2 | / | 70.4 | 42.3 | 10.4 | 18.4 | 160.3 ± 4.2 | |
| Rn (W m−2) | 2020 | −62.4 | −62.8 | −44.8 | −41.1 | −34.2 | −45.1 | −32.4 | −46.0 ± 25.8 |
| 2021 | −39.9 | −59.1 | −44.6 | −42.6 | −44.1 | −39.9 | −31.3 | −43.1 ± 23.3 | |
| 2022 | −54.9 | −58.6 | −57.4 | −46.2 | −36.2 | −48.4 | −42.3 | −49.1 ± 23.5 | |
| 2023 | −46.7 | −40.4 | −53.3 | −50.4 | −44.0 | / | / | −47.3 ± 24.2 | |
| 2024 | −51.8 | −58.3 | / | −28.8 | −43.0 | −39.7 | −45.4 | −45.0 ± 19.9 | |
| GCC | 2020 | 0.337 | 0.352 | 0.349 | 0.340 | 0.337 | 0.341 | 0.342 | 0.343 ± 0.006 |
| 2021 | 0.337 | 0.347 | 0.351 | 0.349 | 0.346 | 0.345 | 0.342 | 0.345 ± 0.005 | |
| 2022 | 0.336 | 0.346 | 0.352 | 0.348 | 0.344 | 0.344 | 0.340 | 0.344 ± 0.005 | |
| 2023 | 0.337 | 0.345 | 0.349 | 0.348 | 0.346 | 0.344 | / | 0.345 ± 0.005 | |
| 2024 | 0.338 | 0.347 | / | 0.342 | 0.341 | 0.342 | 0.341 | 0.343 ± 0.004 | |
| Variable | ETN | Ta | RH | VPD | WS | Ts-Ta | SWC20 | SWC40 | SWC80 | Rn |
|---|---|---|---|---|---|---|---|---|---|---|
| ETN | 1 | |||||||||
| Ta | 0.02 | 1 | ||||||||
| RH | −0.26 a | −0.03 b | 1 | |||||||
| VPD | 0.18 a | 0.59 a | 0.76 a | 1 | ||||||
| WS | 0.11 a | 0.04 a | 0.04 a | −0.03 a | 1 | |||||
| Ts-Ta | −0.14 a | −0.06 a | 0.77 a | 0.23 a | 0.23 a | 1 | ||||
| SWC20 | 0.16 a | −0.22 a | −0.06 a | 0.09 a | −0.04 a | −0.19 a | 1 | |||
| SWC40 | 0.13 a | −0.1 a | −0.10 a | 0.03 b | 0.02 | −0.23 a | 0.82 a | 1 | ||
| SWC80 | 0.08 a | 0.07 a | −0.06 a | 0.02 b | 0.05 a | −0.22 a | 0.50 a | 0.65 a | 1 | |
| Rn | 0.08 a | −0.16 a | 0.56 a | −0.52 a | −0.05 a | 0.76 a | 0.06 a | −0.04 a | −0.04 a | 1 |
References
- Xue, Y.; Zhang, Z.; Li, X.; Liang, H.; Yin, L. A Review of Evapotranspiration Estimation Models: Advances and Future Development. Water Resour. Manag. 2025, 39, 3641–3657. [Google Scholar] [CrossRef]
- Meidner, H.; Mansfield, T.A. Stomatal Responses to Illumination. Biol. Rev. 1965, 40, 483–508. [Google Scholar] [CrossRef]
- Groh, J.; Puetz, T.; Gerke, H.H.; Vanderborght, J.; Vereecken, H. Quantification and Prediction of Nighttime Evapotranspiration for Two Distinct Grassland Ecosystems. Water Resour. Res. 2019, 55, 2961–2975. [Google Scholar] [CrossRef]
- Padron, R.S.; Gudmundsson, L.; Michel, D.; Seneviratne, S. Terrestrial Water Loss at Night: Global Relevance from Observations and Climate Models. Hydrol. Earth Syst. Sci. 2020, 24, 793–807. [Google Scholar] [CrossRef]
- Liu, Z.; Yu, S.; Xu, L.; Wang, Y.; Yu, P.; Chao, Y. Differentiated Responses of Daytime and Nighttime Sap Flow to Soil Water Deficit in a Larch Plantation in Northwest China. Agric. Water Manag. 2023, 289, 108540. [Google Scholar] [CrossRef]
- de Dios, V.R.; Roy, J.; Ferrio, J.P.; Alday, J.G.; Landais, D.; Milcu, A.; Gessler, A. Processes Driving Nocturnal Transpiration and Implications for Estimating Land Evapotranspiration. Sci. Rep. 2015, 5, 10975. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Costa, J.M.; Zarrouk, O.; Pinheiro, C.; Lopes, C.M.; Pereira, J.S. Controlling Stomatal Aperture in Semi-Arid Regions-The Dilemma of Saving Water or Being Cool? Plant Sci. 2016, 251, 54–64. [Google Scholar] [CrossRef]
- Han, Q.; Wang, T.; Wang, L.; Smettem, K.; Mai, M.; Chen, X. Comparison of Nighttime With Daytime Evapotranspiration Responses to Environmental Controls Across Temporal Scales Along a Climate Gradient. Water Resour. Res. 2021, 57, e2021WR029638. [Google Scholar] [CrossRef]
- Yu, S.; Guo, J.; Liu, Z.; Wang, Y.; Xu, L.; Yu, P.; He, L. Impacts of Environmental and Canopy Conditions on the Nighttime Sap Flow of Larch Plantations in the Liupan Mountains, China. J. For. Res. 2023, 34, 1927–1940. [Google Scholar] [CrossRef]
- Forster, M.A. How Significant Is Nocturnal Sap Flow? Tree Physiol. 2014, 34, 757–765. [Google Scholar] [CrossRef]
- Kupper, P.; Rohula, G.; Saksing, L.; Sellin, A.; Lõhmus, K.; Ostonen, I.; Helmisaari, H.-S.; Sõber, A. Does Soil Nutrient Availability Influence Night-Time Water Flux of Aspen Saplings? Environ. Exp. Bot. 2012, 82, 37–42. [Google Scholar] [CrossRef]
- Zeppel, M.J.B.; Lewis, J.D.; Medlyn, B.; Barton, C.V.M.; Duursma, R.A.; Eamus, D.; Adams, M.A.; Phillips, N.; Ellsworth, D.S.; Forster, M.A.; et al. Interactive Effects of Elevated CO2 and Drought on Nocturnal Water Fluxes in Eucalyptus Saligna. Tree Physiol. 2011, 31, 932–944. [Google Scholar] [CrossRef]
- Karpul, R.H.; West, A.G. Wind Drives Nocturnal, but Not Diurnal, Transpiration in Leucospermum Conocarpodendron Trees: Implications for Stilling on the Cape Peninsula. Tree Physiol. 2016, 36, 954–966. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Yano, K. Nocturnal versus Diurnal Transpiration in Rice Plants: Analysis of Five Genotypes Grown under Different Atmospheric CO2 and Soil Moisture Conditions. Agric. Water Manag. 2023, 286, 108397. [Google Scholar] [CrossRef]
- Vega, C.; Chi, C.-J.E.; Fernandez, V.; Burkhardt, J. Nocturnal Transpiration May Be Associated with Foliar Nutrient Uptake. Plants 2023, 12, 531. [Google Scholar] [CrossRef]
- Susana Alvarado-Barrientos, M.; Holwerda, F.; Geissert, D.R.; Munoz-Villers, L.E.; Gotsch, S.G.; Asbjornsen, H.; Dawson, T.E. Nighttime Transpiration in a Seasonally Dry Tropical Montane Cloud Forest Environment. Trees-Struct. Funct. 2015, 29, 259–274. [Google Scholar] [CrossRef]
- Yu, T.; Feng, Q.; Si, J.; Zhang, X.; Alec, D.; Zhao, C. Evidences and Magnitude of Nighttime Transpiration Derived from Populus Euphratica in the Extreme Arid Region of China. J. Plant Biol. 2016, 59, 648–657. [Google Scholar] [CrossRef]
- Chen, L.X.; Zhang, Z.Q.; Li, Z.D.; Zhang, W.J.; Zhang, X.F.; Dong, K.Y.; Wang, G.Y. Nocturnal Sap Flow of Four Urban Greening Tree Species in Dalian, Liaoning Province, China. Acta Phytoecol. Sin. 2010, 34, 535–546. [Google Scholar] [CrossRef]
- O’Keefe, K.; Nippert, J.B. Drivers of Nocturnal Water Flux in a Tallgrass Prairie. Funct. Ecol. 2018, 32, 1155–1167. [Google Scholar] [CrossRef]
- Zhao, C.; Si, J.; Feng, Q.; Yu, T.; Li, P.; Forster, M.A. Nighttime Transpiration of Populus Euphratica during Different Phenophases. J. For. Res. 2019, 30, 435–444. [Google Scholar] [CrossRef]
- Novick, K.A.; Oren, R.; Stoy, P.C.; Siqueira, M.B.S.; Katul, G.G. Nocturnal Evapotranspiration in Eddy-Covariance Records from Three Co-Located Ecosystems in the Southeastern US: Implications for Annual Fluxes. Agric. For. Meteorol. 2009, 149, 1491–1504. [Google Scholar] [CrossRef]
- Guo, X.; Shang, G.; Tian, Y.; Jia, X.; Zha, T.; Li, C.; Yang, H.; Zhang, X. Dynamics of Nocturnal Evapotranspiration and Its Biophysical Controls over a Desert Shrubland of Northwest China. Forests 2021, 12, 1296. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, Q.; Wang, S.; Liang, Y.; Zhang, L.; Yue, P.; Zhao, F. Preliminary study on the nocturnal surface energy imbalance on the Loess Plateau. Chin. J. Geophys. 2025, 68, 385–398. [Google Scholar] [CrossRef]
- Liang, J.; Zhang, L.; Cao, X.; Wen, J.; Wang, J.; Wang, G. Energy Balance in the Semiarid Area of the Loess Plateau, China. J. Geophys. Res. Atmos. 2017, 122, 2155–2168. [Google Scholar] [CrossRef]
- Montoro, A.; Torija, I.; Manas, F.; Lopez-Urrea, R. Lysimeter Measurements of Nocturnal and Diurnal Grapevine Transpiration: Effect of Soil Water Content, and Phenology. Agric. Water Manag. 2020, 229, 105882. [Google Scholar] [CrossRef]
- Liao, Q.; Li, X.; Shi, F.; Deng, Y.; Wang, P.; Wu, T.; Wei, J.; Zuo, F. Diurnal Evapotranspiration and Its Controlling Factors of Alpine Ecosystems during the Growing Season in Northeast Qinghai-Tibet Plateau. Water 2022, 14, 700. [Google Scholar] [CrossRef]
- Skaggs, K.E.; Irmak, S. Characterization of Nighttime Evapotranspiration and Other Surface Energy Fluxes and Interactions with Microclimatic Variables in Subsurface Drip and Center-Pivot Irrigated Soybean Fields. Trans. ASABE 2011, 54, 941–952. [Google Scholar] [CrossRef]
- Guo, X.; Xiao, J.; Zha, T.; Shang, G.; Liu, P.; Jin, C.; Zhang, Y. Dynamics and Biophysical Controls of Nocturnal Water Loss in a Winter Wheat-Summer Maize Rotation Cropland: A Multi-Temporal Scale Analysis. Agric. For. Meteorol. 2023, 342, 109701. [Google Scholar] [CrossRef]
- Chowdhury, F.I.; Arteaga, C.; Alam, M.S.; Alam, I.; Resco de Dios, V. Drivers of Nocturnal Stomatal Conductance in C3 and C4 Plants. Sci. Total Environ. 2022, 814, 151952. [Google Scholar] [CrossRef]
- Sun, M.; Dong, Q.; Jiao, M.; Zhao, X.; Gao, X.; Wu, P.; Wang, A. Estimation of Actual Evapotranspiration in a Semiarid Region Based on GRACE Gravity Satellite Data—A Case Study in Loess Plateau. Remote Sens. 2018, 10, 2032. [Google Scholar] [CrossRef]
- Zhao, G.; Mu, X.; Wen, Z.; Wang, F.; Gao, P. Soil Erosion, Conservation, and Eco-Environment Changes in the Loess Plateau of China. Land Degrad. Dev. 2013, 24, 499–510. [Google Scholar] [CrossRef]
- Sun, W.; Song, X.; Mu, X.; Gao, P.; Wang, F.; Zhao, G. Spatiotemporal Vegetation Cover Variations Associated with Climate Change and Ecological Restoration in the Loess Plateau. Agric. For. Meteorol. 2015, 209, 87–99. [Google Scholar] [CrossRef]
- Li, Z.; Liu, W.; Zhang, X.; Zheng, F. Impacts of Land Use Change and Climate Variability on Hydrology in an Agricultural Catchment on the Loess Plateau of China. J. Hydrol. 2009, 377, 35–42. [Google Scholar] [CrossRef]
- Zhang, S.; Sadras, V.; Chen, X.; Zhang, F. Water Use Efficiency of Dryland Wheat in the Loess Plateau in Response to Soil and Crop Management. Field Crops Res. 2013, 151, 9–18. [Google Scholar] [CrossRef]
- Ge, J.; Pitman, A.J.; Guo, W.; Zan, B.; Fu, C. Impact of Revegetation of the Loess Plateau of China on the Regional Growing Season Water Balance. Hydrol. Earth Syst. Sci. 2020, 24, 515–533. [Google Scholar] [CrossRef]
- Zhao, F.; Ma, S.; Wu, Y.; Qiu, L.; Wang, W.; Lian, Y.; Chen, J.; Sivakumar, B. The Role of Climate Change and Vegetation Greening on Evapotranspiration Variation in the Yellow River Basin, China. Agric. For. Meteorol. 2022, 316, 108842. [Google Scholar] [CrossRef]
- Liu, Y.; Li, S.; Chen, F.; Yang, S.; Chen, X. Soil Water Dynamics and Water Use Efficiency in Spring Maize (Zea Mays L.) Fields Subjected to Different Water Management Practices on the Loess Plateau, China. Agric. Water Manag. 2010, 97, 769–775. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, D.; Liu, H.; Lei, H.; Guo, F.; Xie, S.; Meng, X.; Huang, Q. Energy Flux Observation in a Shrub Ecosystem of a Gully Region of the Chinese Loess Plateau. Ecohydrol. Hydrobiol. 2022, 22, 323–336. [Google Scholar] [CrossRef]
- Guo, F.; Liu, D.; Mo, S.; Li, Q.; Meng, J.; Huang, Q. Assessment of Phenological Dynamics of Different Vegetation Types and Their Environmental Drivers with Near-Surface Remote Sensing: A Case Study on the Loess Plateau of China. Plants 2024, 13, 1826. [Google Scholar] [CrossRef]
- Guo, F.; Liu, D.; Mo, S.; Huang, Q.; Ma, L.; Xie, S.; Deng, W.; Ming, G.; Fan, J. Estimation of Daily Evapotranspiration in Gully Area Scrub Ecosystems on Loess Plateau of China Based on Multisource Observation Data. Ecol. Indic. 2023, 154, 110671. [Google Scholar] [CrossRef]
- Richardson, A.D.; Hufkens, K.; Milliman, T.; Aubrecht, D.M.; Chen, M.; Gray, J.M.; Johnston, M.R.; Keenan, T.F.; Klosterman, S.T.; Kosmala, M.; et al. Tracking Vegetation Phenology across Diverse North American Biomes Using PhenoCam Imagery. Sci. Data 2018, 5, 180028. [Google Scholar] [CrossRef] [PubMed]
- Sonnentag, O.; Hufkens, K.; Teshera-Sterne, C.; Young, A.M.; Friedl, M.; Braswell, B.H.; Milliman, T.; O’Keefe, J.; Richardson, A.D. Digital Repeat Photography for Phenological Research in Forest Ecosystems. Agric. For. Meteorol. 2012, 152, 159–177. [Google Scholar] [CrossRef]
- Richardson, A.D.; Braswell, B.H.; Hollinger, D.Y.; Jenkins, J.P.; Ollinger, S.V. Near-Surface Remote Sensing of Spatial and Temporal Variation in Canopy Phenology. Ecol. Appl. 2009, 19, 1417–1428. [Google Scholar] [CrossRef] [PubMed]
- Webb, E.K.; Pearman, G.I.; Leuning, R. Correction of Flux Measurements for Density Effects Due to Heat and Water Vapour Transfer. Q. J. R. Meteorol. Soc. 1980, 106, 85–100. [Google Scholar] [CrossRef]
- Wilczak, J.M.; Oncley, S.P.; Stage, S.A. Sonic Anemometer Tilt Correction Algorithms. Bound.-Layer Meteorol. 2001, 99, 127–150. [Google Scholar] [CrossRef]
- Mauder, M.; Cuntz, M.; Drüe, C.; Graf, A.; Rebmann, C.; Schmid, H.P.; Schmidt, M.; Steinbrecher, R. A Strategy for Quality and Uncertainty Assessment of Long-Term Eddy-Covariance Measurements. Agric. For. Meteorol. 2013, 169, 122–135. [Google Scholar] [CrossRef]
- Wilson, K.; Goldstein, A.; Falge, E.; Aubinet, M.; Baldocchi, D.; Berbigier, P.; Bernhofer, C.; Ceulemans, R.; Dolman, H.; Field, C.; et al. Energy Balance Closure at FLUXNET Sites. Agric. For. Meteorol. 2002, 113, 223–243. [Google Scholar] [CrossRef]
- Sun, L.; Zhu, W.; Xie, Z.; Zhan, P.; Li, X. Multi-Dimension Evaluation of Remote Sensing Indices for Land Surface Phenology Monitoring. Natl. Remote Sens. Bull. 2023, 27, 2653–2669. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R.; Hastie, T. A Working Guide to Boosted Regression Trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef]
- Sun, S.; Che, T.; Li, H.; Wang, T.; Ma, C.; Liu, B.; Wu, Y.; Song, Z. Water and Carbon Dioxide Exchange of an Alpine Meadow Ecosystem in the Northeastern Tibetan Plateau Is Energy-Limited. Agric. For. Meteorol. 2019, 275, 283–295. [Google Scholar] [CrossRef]
- Gao, G.; Guo, X.; Feng, Q.; Xu, E.; Hao, Y.; Wang, R.; Jing, W.; Ren, X.; Liu, S.; Shi, J.; et al. Environmental Controls on Evapotranspiration and Its Components in a Qinghai Spruce Forest in the Qilian Mountains. Plants 2024, 13, 801. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zheng, N.; Zhang, J.; Meng, P. Advances in the Study of Regional-Averaged Evapotranspiration Using the Scintillation Method. Acta Ecol. Sin. 2018, 38, 2625–2635. [Google Scholar] [CrossRef]
- Fratini, G.; McDermitt, D.K.; Papale, D. Eddy-Covariance Flux Errors Due to Biases in Gas Concentration Measurements: Origins, Quantification and Correction. Biogeosciences 2014, 11, 1037–1051. [Google Scholar] [CrossRef]
- Heusinkveld, B.G.; Jacobs, A.F.G.; Holtslag, A.A.M. Effect of Open-Path Gas Analyzer Wetness on Eddy Covariance Flux Measurements: A Proposed Solution. Agric. For. Meteorol. 2008, 148, 1563–1573. [Google Scholar] [CrossRef]
- van Gorsel, E.; Leuning, R.; Cleugh, H.A.; Keith, H.; Kirschbaum, M.U.F.; Suni, T. Application of an Alternative Method to Derive Reliable Estimates of Nighttime Respiration from Eddy Covariance Measurements in Moderately Complex Topography. Agric. For. Meteorol. 2008, 148, 1174–1180. [Google Scholar] [CrossRef]
- Sabbatini, S.; Mammarella, I.; Arriga, N.; Fratini, G.; Graf, A.; Hoertriagl, L.; Ibrom, A.; Longdoz, B.; Mauder, M.; Merbold, L.; et al. Eddy Covariance Raw Data Processing for CO2 and Energy Fluxes Calculation at ICOS Ecosystem Stations. Int. Agrophys. 2018, 32, 495–515. [Google Scholar] [CrossRef]
- Guo, X.; Zhang, Y.; Zha, T.; Shang, G.; Jin, C.; Wang, Y.; Yang, H. Biophysical Controls of Dew Formation in a Typical Cropland and Its Relationship to Drought in the North China Plain. J. Hydrol. 2023, 617, 128945. [Google Scholar] [CrossRef]
- Paulus, S.J.; Orth, R.; Lee, S.-C.; Hildebrandt, A.; Jung, M.; Nelson, J.A.; El-Madany, T.S.; Carrara, A.; Moreno, G.; Mauder, M.; et al. Interpretability of Negative Latent Heat Fluxes from Eddy Covariance Measurements in Dry Conditions. Biogeosciences 2024, 21, 2051–2085. [Google Scholar] [CrossRef]
- Liu, M.; Shi, H.; Li, X.; Yan, J.; Sun, W.; Dou, X. Path Analysis of Evapotranspiration Dynamic Variation and Its Influencing Factors in Hetao Irrigation District. J. Drain. Irrig. Mach. Eng. 2018, 36, 1081–1086. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, S.; Jin, L.; Wang, J.; Yang, J. Evapotranspiration of a Film-Mulched Cotton Field under Drip Irrigation in North Xinjiang. Arid Zone Res. 2012, 29, 360–368. [Google Scholar] [CrossRef]
- Ritchie, J.T. Atmospheric and Soil Water Influences on the Plant Water Balance. Agric. Meteorol. 1974, 14, 183–198. [Google Scholar] [CrossRef]
- Fang, W.; Lv, N.; Fu, B. Research Advances in Nighttime Sap Flow Density, Its Physiological Implications, and Influencing Factors in Plants. Acta Ecol. Sin. 2018, 38, 7521–7529. [Google Scholar] [CrossRef]
- Fisher, J.B.; Baldocchi, D.D.; Misson, L.; Dawson, T.E.; Goldstein, A.H. What the Towers Don’t See at Night: Nocturnal Sap Flow in Trees and Shrubs at Two AmeriFlux Sites in California. Tree Physiol. 2007, 27, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Feng, Q.; Si, J.; Mitchell, P.J.; Forster, M.A.; Zhang, X.; Zhao, C. Depressed Hydraulic Redistribution of Roots More by Stem Refilling than by Nocturnal Transpiration for Populus Euphratica Oliv. in Situ Measurement. Ecol. Evol. 2018, 8, 2607–2616. [Google Scholar] [CrossRef]
- Di, N.; Yang, S.; Liu, Y.; Fan, Y.; Duan, J.; Nadezhdina, N.; Li, X.; Xi, B. Soil-Moisture-Dependent Nocturnal Water Use Strategy and Its Responses to Meteorological Factors in a Seasonal-Arid Poplar Plantation. Agric. Water Manag. 2022, 274, 107984. [Google Scholar] [CrossRef]
- Dawson, T.E.; Burgess, S.S.O.; Tu, K.P.; Oliveira, R.S.; Santiago, L.S.; Fisher, J.B.; Simonin, K.A.; Ambrose, A.R. Nighttime Transpiration in Woody Plants from Contrasting Ecosystems. Tree Physiol. 2007, 27, 561–575. [Google Scholar] [CrossRef]
- Si, J.; Feng, Q.; Yu, T.; Zhao, C. Research Advances in Nighttime Transpiration and Its Eco-Hydrological Implications. Adv. Water Sci. 2014, 25, 907–914. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, Z.; Sun, G.; Chen, L.; Xu, H.; Chen, S. Biophysical Controls on Nocturnal Sap Flow in Plantation Forests in a Semi-Arid Region of Northern China. Agric. For. Meteorol. 2020, 284, 107904. [Google Scholar] [CrossRef]
- Bo, Y.; Wang, L.; Jian, S. Variations of Sap Flow of Caragana Korshinskii and Hippophae Rhamnoides in Hilly and Gully Region of the Loess Plateau. Acta Ecol. Sin. 2023, 43, 1553–1562. [Google Scholar] [CrossRef]


| Relationship Between ETN/ET and Environmental Variables | Slope | Intercept | R2 | p-Value |
|---|---|---|---|---|
| ETN/ET–Ta | −0.012 | 0.35 | 0.77 | <0.001 |
| ETN/ET–RH | −0.001 | 0.22 | 0.04 | >0.05 |
| ETN/ET–VPD | −0.011 | 0.28 | 0.27 | <0.01 |
| ETN/ET–WS | −0.02 | 0.21 | 0.02 | >0.05 |
| ETN/ET–(Ts-Ta) | −0.013 | 0.13 | 0.03 | >0.05 |
| ETN/ET–SWC20 | 0.225 | 0.10 | 0.06 | >0.05 |
| ETN/ET–SWC40 | 0.185 | 0.09 | 0.02 | >0.05 |
| ETN/ET–SWC80 | −0.101 | 0.17 | 0.01 | >0.05 |
| ETN/ET–Rn | 0.001 | 0.18 | 0.01 | >0.05 |
| ETN/ET–GCC | −7.51 | 2.73 | 0.31 | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, F.; Liu, D.; Mo, S.; Li, Q.; Zhao, F.; Li, M.; Hussain, F. Dynamics of Nocturnal Evapotranspiration in a Dry Region of the Chinese Loess Plateau: A Multi-Timescale Analysis. Hydrology 2025, 12, 188. https://doi.org/10.3390/hydrology12070188
Guo F, Liu D, Mo S, Li Q, Zhao F, Li M, Hussain F. Dynamics of Nocturnal Evapotranspiration in a Dry Region of the Chinese Loess Plateau: A Multi-Timescale Analysis. Hydrology. 2025; 12(7):188. https://doi.org/10.3390/hydrology12070188
Chicago/Turabian StyleGuo, Fengnian, Dengfeng Liu, Shuhong Mo, Qiang Li, Fubo Zhao, Mingliang Li, and Fiaz Hussain. 2025. "Dynamics of Nocturnal Evapotranspiration in a Dry Region of the Chinese Loess Plateau: A Multi-Timescale Analysis" Hydrology 12, no. 7: 188. https://doi.org/10.3390/hydrology12070188
APA StyleGuo, F., Liu, D., Mo, S., Li, Q., Zhao, F., Li, M., & Hussain, F. (2025). Dynamics of Nocturnal Evapotranspiration in a Dry Region of the Chinese Loess Plateau: A Multi-Timescale Analysis. Hydrology, 12(7), 188. https://doi.org/10.3390/hydrology12070188