Pleiotropic Immune Functions of Chemokine Receptor 6 in Health and Disease


Abstract
1. Introduction
Chemokines
2. Chemokine Receptor CCR6 and Its Ligand CCL20
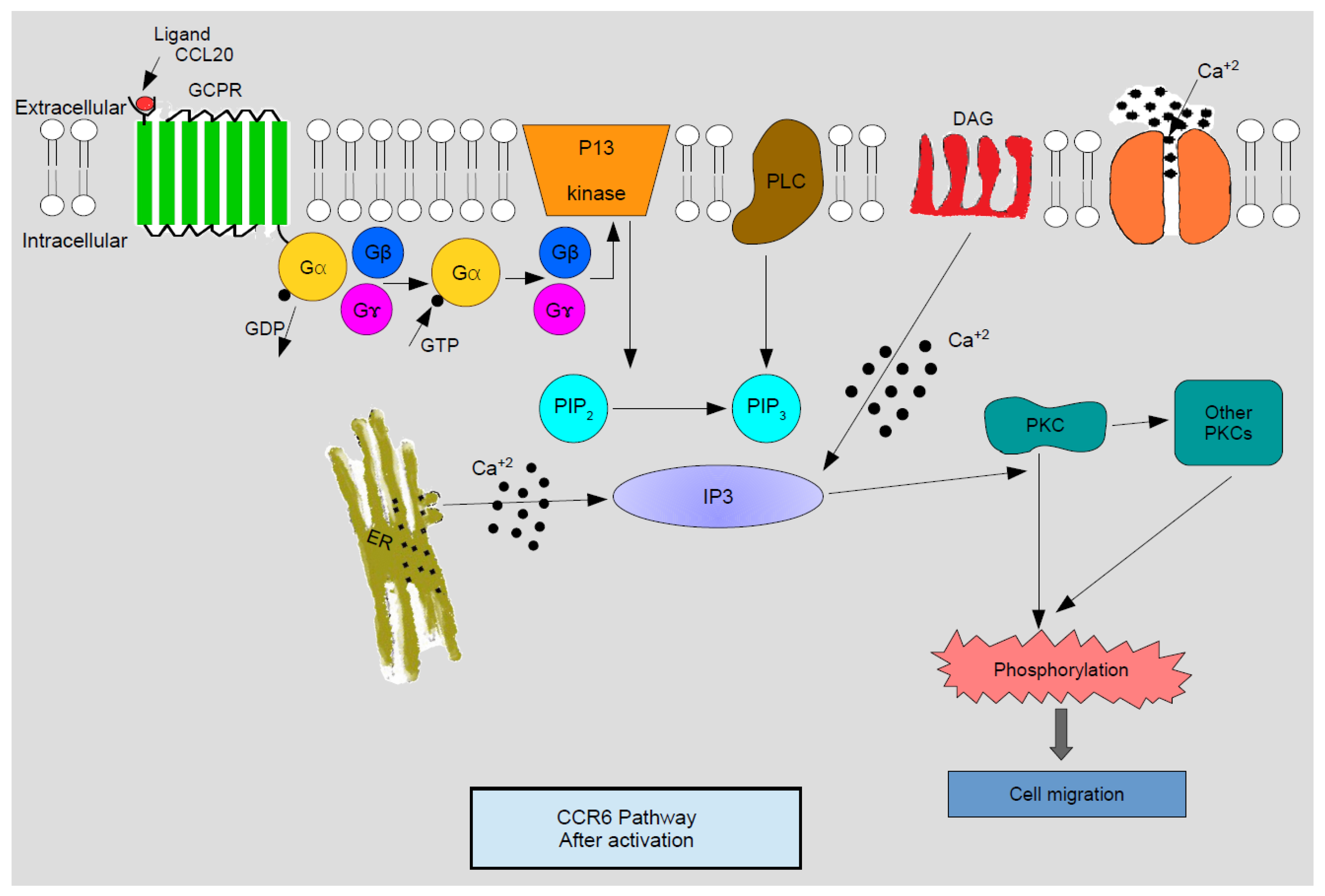
3. CCR6 Signaling Pathway
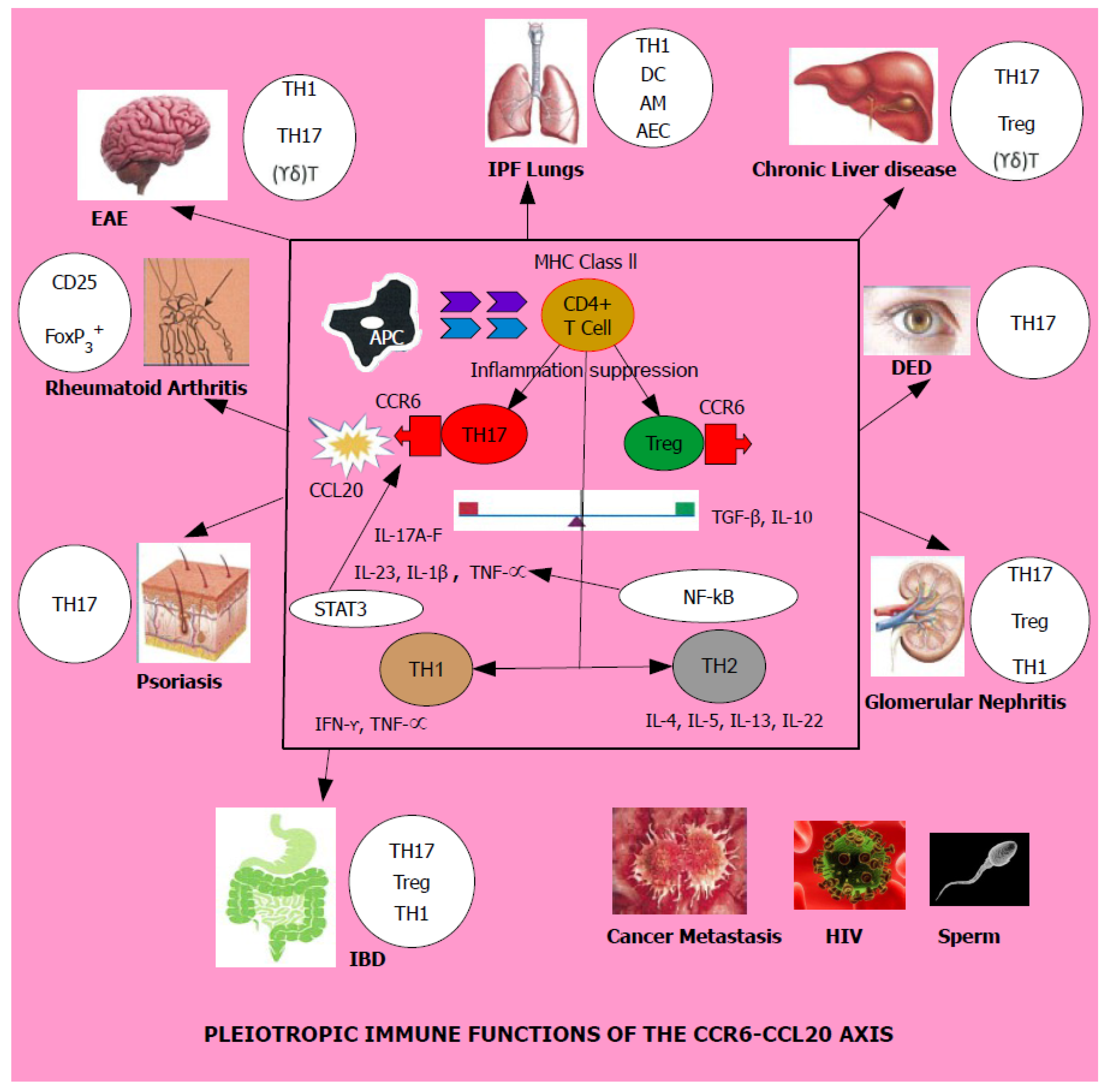
4. CCR6 and CCL20 in Health and Disease
4.1. Lung
4.2. Kidney
4.3. Liver
4.4. Brain
4.5. Eye
4.6. Skin
4.7. Joints
4.8. Gonad
4.9. Gut
4.10. Cancer
4.11. HIV
4.12. Obesity and Diabetes
5. Conclusions
Funding
Conflicts of Interest
References
- Lee, A.Y.S.; Eri, R.; Lyons, A.B.; Grimm, M.C.; Korner, H. CC chemokine ligand CCL20 and its cognate receptor CCR6 in mucosal T cell immunology and inflammatory bowel disease: Odd couple or axis of evil? Front. Immunol. 2013, 4, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Griffith, J.W.; Sokol, C.L.; Luster, A.D. Chemokines and chemokine receptors: Positioning cells for host defense and immunity. Annu. Rev. Immunol. 2014, 32, 659–702. [Google Scholar] [CrossRef] [PubMed]
- Basheer, W.; Kunde, D.; Eri, R. Role of chemokine ligand CCL20 and its receptor CCR6 in intestinal inflammation. Immunol. Infect. Dis. 2013, 1, 30–37. [Google Scholar] [CrossRef]
- Strieter, R.M.; Kunkel, S.L.; Standiford, T. Chemokines in the Lung; Lenfant, C., Ed.; Lung Biology in Health and Disease; CRC Press: Boca Raton, FL, USA, 2003; pp. 1–341. [Google Scholar]
- Lee, A.Y.; Phan, T.K.; Hulett, M.D.; Korner, H. The relationship between CCR6 and its binding partners: Does the CCR6-CCL20 axis have to be extended? Cytokine 2015, 72, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Frick, V.O.; Rubie, C.; Keilholz, U.; Ghadjar, P. Chemokine/chemokine receptor pair CCL20/CCR6 in human colorectal malignancy: An overview. World J. Gastroenterol. 2016, 22, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Williams, I.R. CCR6 and CCL20: Partners in intestinal immunity and lymphorganogenesis. Ann. N. Y. Acad. Sci. 2006, 1072, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Bannan, J. The Role of the CC-Chemokine Receptor 6 in B Cell Differentiation during the Humoral Immune Response. Ph.D. Thesis, University of Tasmania, Hobart, Australia, 2014. [Google Scholar]
- Zlotnik, A.; Yoshie, O.; Nomiyama, H. The chemokine and chemokine receptor super families and their molecular evolution. Genome Biol. 2006, 7, 243. [Google Scholar] [CrossRef] [PubMed]
- Proudfoot, A.E. Chemokine receptors: Multifaceted therapeutic targets. Nat. Rev. Immunol. 2002, 2, 106–115. [Google Scholar] [CrossRef] [PubMed]
- National Centre for Biotechnlogy Information. CCR6 C-C Motif Chemokine Receptor 6 (Human); US National Library of Medicine: Bethesda, MD, USA, 2018. [Google Scholar]
- Zissel, G.; Hohne, K.; Kilic, A.; Maier, C.; Ploenes, T.; Prasse, A.; Trepel, M.; Eibel, H.; Muller-Quernheim, J. CCR6 is a receptor for CCL18 expressed on human lung fibroblasts from IPF lungs. Eur. Respir. J. 2011, 38 (Suppl. 55), 3232. [Google Scholar]
- Lafferty, M.K.; Sun, L.; Christensen-Quick, A.; Lu, W.; Garzino-Demo, A. Human Beta Defensin 2 Selectively Inhibits HIV-1 in Highly Permissive CCR6+CD4+ T Cells. Viruses 2017, 9, 111. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, R.; Rusch, L.; Itano, A.; Jenkins, M.K.; McSorley, S.J. CCR6–dependent recruitment of blood phagocytes is necessary for rapid CD4 T cell responses to local bacterial infection. Proc. Natl. Acad. Sci. USA 2007, 104, 12075–12080. [Google Scholar] [CrossRef] [PubMed]
- Nandi, B.; Pai, C.; Huang, Q.; Prabhala, R.H.; Munshi, N.C.; Gold, J.S. CCR6 the sole receptor for the chemokine CCL20 promotes spontaneous intestinal tumorigenesis. PLoS ONE 2014, 9, e97566. [Google Scholar] [CrossRef] [PubMed]
- Wiede, F.; Fromm, P.D.; Comerford, I.; Kara, E.; Bannan, J.; Schuh, W.; Ranasinghe, C.; Tarlinton, D.; Winkler, T.; McColl, S.R.; et al. CCR6 is transiently upregulated on B cells after activation and modulates the germinal centre reaction in the mouse. Immunol. Cell Biol. 2013, 91, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Comerford, I.; Bunting, M.; Fenix, K.; Haylock-Jacobs, S.; Litchfield, W.; Harata-Lee, Y.; Turvey, M.; Brazzatti, J.; Gregor, C.; Nguyen, P.; et al. An immune paradox: How can the same chemokine axis regulate both immune tolerance and activation? CCR6/CCL20: A chemokine axis balancing immunological tolerance and activation in autoimmune disease. Bioessays 2010, 32, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Schutyser, E.; Struyf, S.; Van Damme, J. The CC chemokine CCL20 and its receptor CCR6. Cytokine Growth Factor Rev. 2003, 14, 409–426. [Google Scholar] [CrossRef]
- Varona, R.; Villares, R.; Carramolino, L.; Gova, I.; Gutierrez, J.; Torres, M.; Martinez-A, C.; Marquez, G. CCR6 deficient mice have impaired leukocyte homeostasis and altered contact hypersensitivity and delayed type hypersensitivity responses. J. Clin. Investig. 2001, 107, R37–R45. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, N.P.; Vongsa, R.A.; Wendt, M.K.; Dwinell, M.B. Chemokines and chemokine receptors in mucosal homeostasis at the intestinal epithelial barrier in inflammatory bowel disease. Inflamm. Bowel Dis. 2008, 14, 1000–1011. [Google Scholar] [CrossRef] [PubMed]
- Elgueta, R.; Marks, E.; Nowak, E.; Menezes, S.; Benson, M.; Raman, V.S.; Ortiz, C.; O’Connell, S.; Hess, H.; Lord, G.M.; et al. CCR6 dependent positioning of memory B cells is essential for their ability to mount a recall response to antigen. J. Immunol. 2015, 194, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Luegering, A.; Mead, J.; Hudson, J.T., III; Kucharzik, T.; Williams, I. CCR6 deficient mice with increased numbers of αβ TCR intestinal intraepithelial lymphocytes exhibit enhanced innate immunity to infection with the nematode Heligmosomoides polygyrus. Gastroenterology 2003, 124 (Suppl. 1), A339. [Google Scholar] [CrossRef]
- Jiang, S. (Ed.) Th17 Cells in Health and Disease; Springer Science and Business Media: New York, NY, USA, 2011; ISBN 978-1-4419-9371-7. [Google Scholar] [CrossRef]
- Lu, M.Y.; Lu, S.S.; Chang, S.L.; Liao, F. The Phosphorylation of CCR6 on Distinct Ser/Thr Residues in the Carboxyl Terminus Differentially Regulates Biological Function. Front. Immunol. 2018, 9, 415. [Google Scholar] [CrossRef] [PubMed]
- Facco, M.; Baesso, I.; Miorin, M.; Bortoli, M.; Cabrelle, A.; Boscaro, E.; Gurrieri, C.; Trentin, L.; Zambello, R.; Calabrese, F.; et al. Expression and role of CCR6/CCL20 chemokine axis in pulmonary sarcoidosis. J. Leukoc. Biol. 2007, 82, 946–955. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Carson, W.F., 4th; Cavassani, K.A.; Connett, J.M.; Kunkel, S.L. CCR6 as a mediator of immunity in the lung and gut. Exp. Cell Res. 2011, 317, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Luzina, I.G.; Atamas, S.P. CCR6 is not necessary for functional effects of human CCL18 in a mouse model. Fibrogenes. Tissue Repair 2012, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.; Paust, H.J.; Steinmetz, O.M.; Peters, A.; Riedel, J.H.; Erhardt, A.; Wegscheid, C.; Velden, J.; Fehr, S.; Mittrucker, H.W.; et al. CCR6 recruits regulatory T cells and TH17 cells to the kidney in glomerularnephritis. J. Am. Soc. Nephrol. 2010, 21, 974–985. [Google Scholar] [CrossRef] [PubMed]
- Welsh-Bacic, D.; Lindenmeyer, M.; Cohen, C.D.; Draganovici, D.; Mandelbaum, J.; Edenhofer, I.; Ziegler, U.; Regele, H.; Wuthrich, R.P.; Segerer, S. Expression of the chemokine receptor CCR6 in human renal inflammation. Nephrol. Dial. Transpl. 2011, 26, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Alikhan, M.A.; Huynh, M.; Kitching, A.R.; Ooi, J.D. Regulatory T cells in renal disease. Clin. Transl. Immunol. 2018, 7, e1004. [Google Scholar] [CrossRef] [PubMed]
- Chung, A.C.; Lan, H.Y. Chemokines in renal injury. J. Am. Soc. Nephrol. 2011, 22, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Hammerich, L.; Bangen, J.M.; Govaere, O.; Zimmermann, H.W.; Gassler, N.; Huss, S.; Liedtke, C.; Prinz, I.; Lira, S.A.; Luedde, T.; et al. Chemokine Receptor CCR6-Dependent Accumulation of gamma delta T Cells in Injured Liver Restricts Hepatic Inflammation and Fibrosis. Hepatology 2014, 59, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Dellacasagrande, J.; Schreurs, O.J.F.; Hofgaard, P.O.; Omholt, H.; Steinsvoll, S.; Schenck, K.; Bogen, B.; Dembic, Z. Liver metastasis of cancer facilitated by chemokine receptor CCR6. Scand J. Immunol. 2003, 57, 534. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M.; Trettel, F. Chemokines and chemokine receptors in brain homeostasis. Front. Cell. Neurosci. 2015, 9, 132. [Google Scholar] [CrossRef] [PubMed]
- Robert, R.; Ang, C.; Sun, G.; Juglair, L.; Lim, F.X.; Mason, L.J.; Payne, N.L.; Bernard, C.C.A.; Mackay, C.R. Essential role for CCR6 in certain inflammatory diseases demonstrated using specific antagonist and knockin mice. JCI Insight 2017, 2, e94821. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, P.; Ludewig, P.; Melich, P.; Arumugam, T.V.; Gerloff, C.; Prinz, I.; Magnus, T.; Gelderblom, M. CCR6 (CC Chemokine Receptor 6) is essential for the migration of detrimental natural interleukin-17—Producing γδ T cells in stroke. Stroke 2017, 48, 1957–1965. [Google Scholar] [CrossRef] [PubMed]
- Dohlman, T.H.; Chauhan, S.K.; Kodati, S.; Hua, J.; Chen, Y.; Omoto, M.; Sadrai, Z.; Dana, R. The CCR6/CCL20 axis mediates Th17 cell migration to the ocular surface in dry eye disease. Investig. Opthalmol. Vis. Sci. 2013, 54, 4081–4091. [Google Scholar] [CrossRef] [PubMed]
- Rafaels, N.M.; Beck, L.A.; Gao, P.S.; Hand, T.; Boguniewicz, M.; Hata, T.R.; Schneider, L.; Hanifin, J.M.; Gallo, R.L.; Gao, L.; et al. Variations in the CCL20 and CCR6 Genes are Associated with Atopic Dermatitis and Eczema Herpeticum in Populations of European and African descent. J. Allergy Clin. Immunol. 2009, 123, S70. [Google Scholar] [CrossRef]
- Hedrick, M.N.; Lonsdorf, A.S.; Hwang, S.T.; Farber, J.M. CCR6 as a possible therapeutic target in psoriasis. Expert Opin. Ther. Targets 2010, 14, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.T. Function and Targeting of CCR6/CCL20 in Autoimmune Psoriasiform; National Institute of Health: Bethesda, MD, USA, 2017; Available online: grantome.com/grant/NIH/R01-AR063091-01A1 (accessed on 25 May 2018).
- Bonelli, M.; Puchner, A.; Goeschl, L.; Hayer, S.; Smolen, J.; Scheinecker, C.; Bluml, S. CCR6 expression drives arthritis in a T cell dependent manner [abstract]. In Proceedings of the 2016 ACR/ARHP Annual Meeting, Washington, DC, USA, 11 November 2016; Available online: http://acrabstracts.org/abstract/ccr6-expression-regulates-arthritis-in-a-t-cell-dependent-manner/ (accessed on 25 May 2018).
- Bonelli, M.; Goeschl, L.; Puchner, A.; Bluml, S. CCR6 modulates severity of arthritis in T cell dependent manner. BMJ J. 2017, 76 (Suppl. 1). [Google Scholar] [CrossRef]
- Dairaghi, D.; Zhang, P.; Leleti, M.; Berahovich, R.; Ebsworth, K.; Ertl, L.; Miao, S.; Miao, Z.; Seitz, L.; Tan, J.; et al. Inhibition of Chemokine Receptors CCR1 and CCR6 as Promising Therapies for Autoimmune Diseases such as Rheumatoid Arthritis and Psoriasis. In Proceedings of the 2013 ACR/ARHP Annual Meeting in Annals of the Rheumatic Diseases, San Diego, CA, USA, 25–30 October 2013; Volume 72. [Google Scholar]
- Julian, B.; Gao, K.; Harwood, B.N.; Beinborn, M.; Kopin, A.S. Mutation-induced functional alterations of CCR6. J. Pharmacol. Exp. Ther. 2017, 360, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Campo, P.; Buffone, M.G.; Benencia, F.; Conejo-Garcia, J.R.; Rinaudo, P.F.; Gerton, G.L. A Role for the Chemokine Receptor CCR6 in mammalian sperm motility and chemotaxis. J. Cell. Physiol. 2013, 229, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Varona, R.; Cadenas, V.; Flores, J.; Martinez-A, C.; Marquez, G. CCR6 has a non redundant role in the development of inflammatory bowel disease. Eur. J. Immunol. 2003, 33, 2937–2946. [Google Scholar] [CrossRef] [PubMed]
- Skovdhal, H.K.; Granlund, A.V.; Ostvik, A.E.; Bruland, T.; Bakke, J.; Torp, S.H.; Damas, J.K.; Sandvik, A.K. Expression of CCL20 and its corresponding receptor CCR6 is enhanced in active inflammatory bowel disease and TLR3 mediates CCL20 expression in colonic epithelial cells. PLoS ONE 2015, 10, e0141710. [Google Scholar] [CrossRef] [PubMed]
- Shouval, D.S.; Rufo, P.A. The role of environmental factors in the pathogenesis of inflammatory bowel diseases: A review. JAMA Pediatr. 2017, 171, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Cader, M.Z.; Kaser, A. Recent advances in inflammatory bowel disease: Mucosal immune cells in intestinal inflammation. Gut 2013, 62, 1653–1664. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kwon, J.; Cho, M. Immunological pathogenesis of inflammatory bowel disease. Intest. Res. 2018, 16, 26–42. [Google Scholar] [CrossRef] [PubMed]
- Fakhoury, M.; Negruli, R.; Mooranian, A.; Al-Salami, H. Inflammatory bowel disease: Clinical aspects and treatments. J. Inflamm. Res. 2014, 7, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Pedros, C.; Duguet, F.; Saoudi, A.; Chabod, M. Disrupted regulatory T cell homeostasis in inflammatory bowel disease. World J. Gastroenterol. 2016, 22, 974–995. [Google Scholar] [CrossRef] [PubMed]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef] [PubMed]
- Muzes, G.; Molnar, B.; Tulassay, Z.; Sipos, F. Changes of the cytokine profile in inflammatory bowel diseases. World J. Gastroenterol. 2012, 18, 5848–5861. [Google Scholar] [CrossRef] [PubMed]
- Eri, R.; McGuckin, M.A.; Wadley, R. T cell transfer model of colitis: A great tool to assess the contribution of T cells in chronic intestinal inflammation. Methods Mol. Biol. 2012, 844, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Yousaf, W.; Giannella, R.; Shata, M.T. Th17 cells interactions with predisposing factors of immunopathogenesis of inflammatory bowel disease. Expert Rev. Clin. Immunol. 2012, 8, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Westphal, S.; Lugering, A.; von Wedel, J.; von Eiff, C.; Maaser, C.; Spahn, T.; Heusipp, G.; Schmidt, M.A.; Herbst, H.; Williams, I.R.; et al. Resistance of chemokine receptor 6-deficient mice to Yersinia enterocolitica infection: Evidence of defective M-cell formation in vivo. Am. J. Pathol. 2008, 172, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Rubie, C.; Frick, V.O.; Ghadjar, P.; Wagner, M.; Grimm, H.; Vicinus, B.; Justinger, C.; Graeber, S.; Schilling, M.K. CCL20/CCR6 expression profile in pancreatic cancer. J. Transl. Med. 2010, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Wei, J.W.; Tao, Y.L.; Ding, P.R.; Xia, Y.F.; Gao, Y.H.; Xiao, W.W. CCR6 is a predicting biomarker of radiosensitivity and potential target of radiosensitization in rectal cancer. Cancer Res. Treat 2017. [Google Scholar] [CrossRef] [PubMed]
- Martin-Garcia, D.; Lonsdorf, A.; Enk, A. Characterization and modulation of CC-chemokine receptor 6 (CCR6) and CC ligand 20 (CCL20)—Mediated immunosurveillance in malignant melanoma. J. Investig. Dermatol. 2017, 137, S289. [Google Scholar] [CrossRef]
- Ikeda, S.; Kitadate, A.; Ito, M.; Abe, F.; Nara, M.; Watanabe, A.; Takahashi, N.; Miyagaki, T.; Sugaya, M.; Tagawa, H. Disruption of CCL20-CCR6 interaction inhibits metastasis of advanced cutaneous T cell lymphoma. Oncotarget 2016, 7, 13563–13574. [Google Scholar] [CrossRef] [PubMed]
- Boyle, S.T.; Faulkner, J.W.; McColl, S.R.; Kochetkova, M. The chemokine receptor CCR6 facilitates the onset of mammary neoplasia in the MMTV-PyMT mouse model via recruitment of tumor-promoting macrophages. Mol. Cancer 2015, 14, 115. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, J.; Tie, H.; Huang, H.; Li, H.; Ren, G. Prognostic significance of CCL20 and its receptor CCR6 in cancers: A meta-analysis. Int. J. Clin. Exp. Med. 2016, 9, 1908–1919. [Google Scholar]
- Moody, S.; Thompson, J.S.; Chuang, S.S.; Liu, H.; Raderer, M.; Vassiliou, G.; Wlodarska, I.; Wu, F.; Cogliatti, S.; Robson, A.; et al. Novel GPR34 and CCR6 mutation and distinct genetic profiles in MALT lymphomas of different sites. Haematologica 2018. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.S.; Korner, H. CCR6/CCL20 chemokine axis in human immunodeficiency virus immunity and pathogenesis. J. Gen. Virol. 2017, 98, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, Y.; Tuan, M.; Nawaz, F.; Arthos, J.; Wolff, M.J.; Poles, M.A.; Hioe, C.E. Preferential HIV infection of CCR6+ Th17 cells is associated with higher levels of virus receptor expression and lack of CCR5 ligands. J. Virol. 2013, 87, 10843–10854. [Google Scholar] [CrossRef] [PubMed]
- Duffaut, C.; Zakaroff-Girard, A.; Bourlier, V.; Decaunes, P.; Maumus, M.; Chiotasso, P.; Sengenes, C.; Lafontan, M.; Galitzky, J.; Bouloumie, A. Interplay between human adipocytes and T lymphocytes in obesity: CCL20 as an adipochemokine and T lymphocytes as lipogenic modulators. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1608–1614. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.J.; Karlstad, M.D.; Regal, K.M.; Sparer, T.E.; Lu, D.; Elks, C.M.; Grant, R.W.; Stephens, J.M.; Burk, D.H.; Jason, C.J. CCL20 is elevated during obesity and differentially regulated by NF-kB subunits in pancreatic β-cells. Biochim. Biophys. Acta 2015, 1849, 637–652. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Yang, H.; Tartar, D.M.; Gao, B.; Luo, X.; Ye, S.Q.; Zaghouani, H.; Fang, D. Prevention and treatment of diabetes with resveratrol in a non-obese mouse model of type 1 diabetes. Diabetologia 2011, 54, 1136–1146. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Organ | CCR6/CCL20 Axis | Cell Types Involved | Disease Produced | References |
|---|---|---|---|---|
| Lung | Operative | Fibroblasts, AEC, DC, TH1, AM | IPFL, Sarcoidosis | [12,25,26] |
| Liver | Operative | TH17, Treg, (γδ) T cells | Chronic liver disease | [32,33] |
| Kidney | Operative | TH1, TH17, Treg | Glomerular nephritis | [28,29,30,31] |
| Brain | Operative | TH1, TH17, (γδ) T cells | EAE, Stroke | [34,35,36] |
| Eye | Operative | TH17 | Dry eye disease | [37] |
| Skin | Operative | TH17 | Psoriasis | [39,40] |
| Joints | Operative | CD25-FoxP3+ | Rheumatoid arthritis | [41,42,43] |
| Gut | Operative | TH17, Treg, TH1 | IBD | [1,50,51,52] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ranasinghe, R.; Eri, R. Pleiotropic Immune Functions of Chemokine Receptor 6 in Health and Disease. Medicines 2018, 5, 69. https://doi.org/10.3390/medicines5030069
Ranasinghe R, Eri R. Pleiotropic Immune Functions of Chemokine Receptor 6 in Health and Disease. Medicines. 2018; 5(3):69. https://doi.org/10.3390/medicines5030069
Chicago/Turabian StyleRanasinghe, Ranmali, and Rajaraman Eri. 2018. "Pleiotropic Immune Functions of Chemokine Receptor 6 in Health and Disease" Medicines 5, no. 3: 69. https://doi.org/10.3390/medicines5030069
APA StyleRanasinghe, R., & Eri, R. (2018). Pleiotropic Immune Functions of Chemokine Receptor 6 in Health and Disease. Medicines, 5(3), 69. https://doi.org/10.3390/medicines5030069