Do Anti-Oxidants Vitamin D3, Melatonin, and Alpha-Lipoic Acid Have Synergistic Effects with Temozolomide on Cultured Glioblastoma Cells?


Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture and Treatments
2.3. MTT Cytotoxicity Assay
2.4. Measurement of ROS Production
2.5. Glutathione Peroxidase Activity Assay
2.6. Statistical Analysis
3. Results
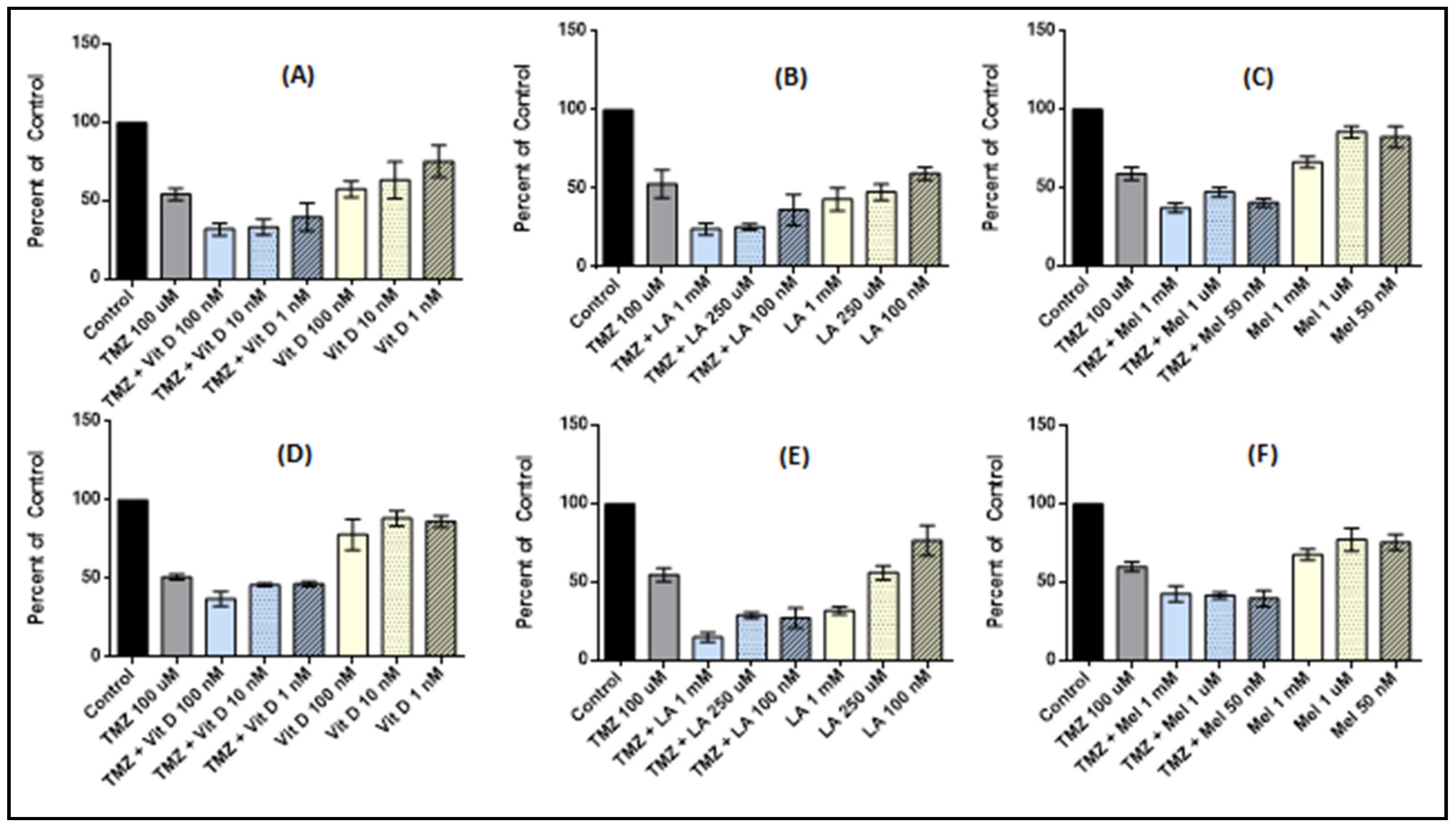
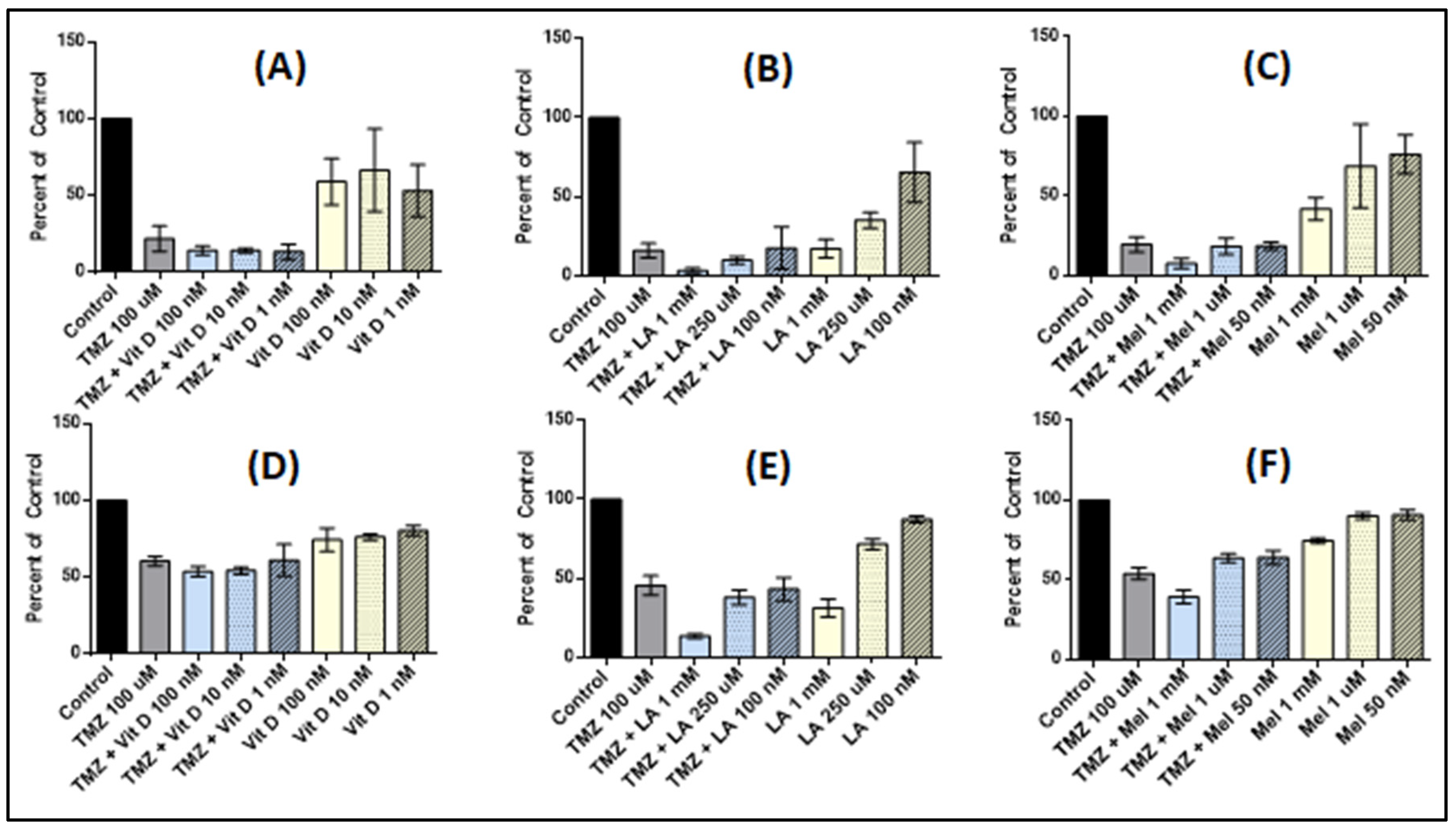
3.1. Vitamin D3 Effect on Cell Proliferation
3.2. Lipoic Acid Effects on Cell Proliferation
3.3. Melatonin Effects on Cell Proliferation
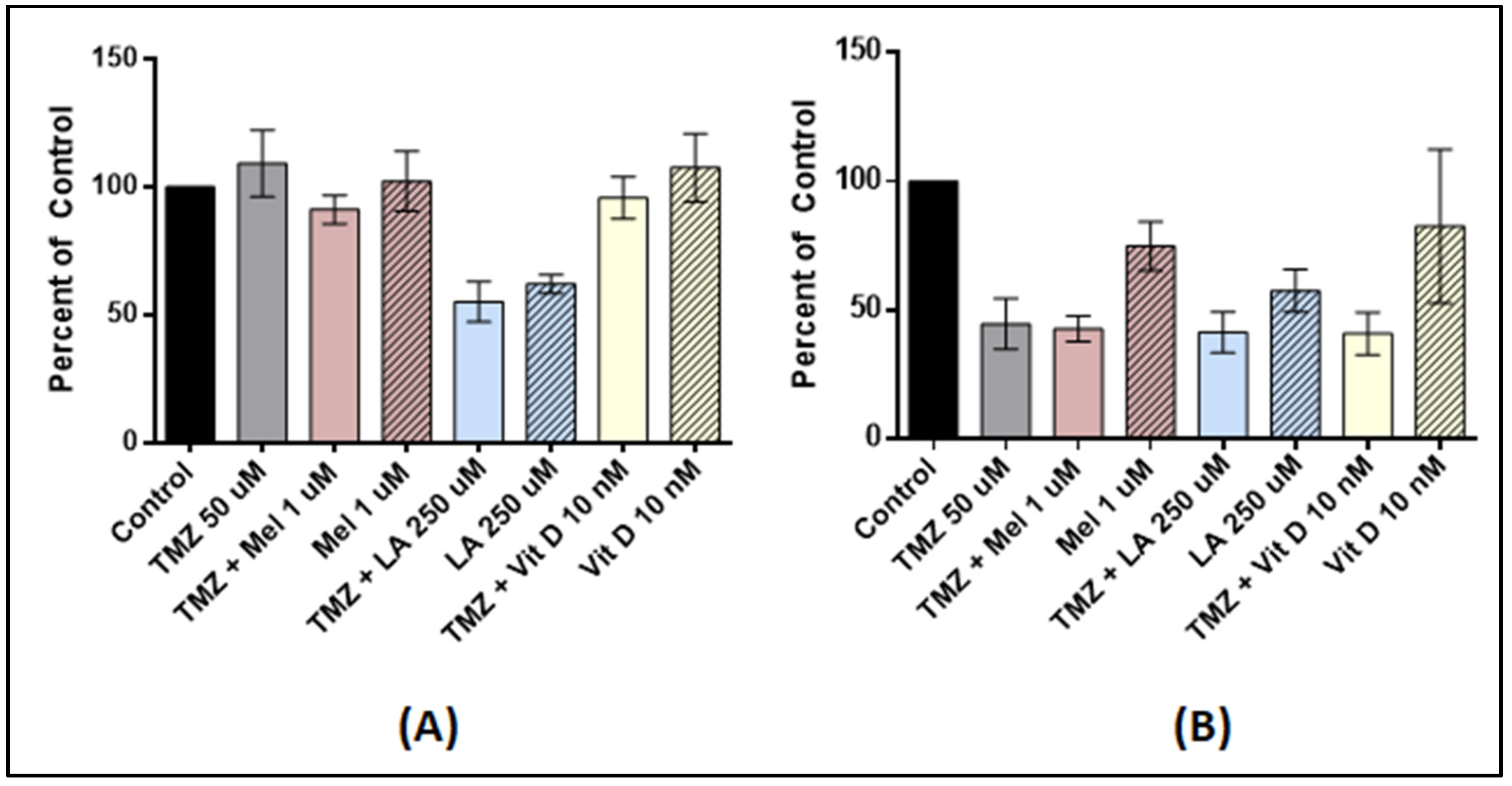
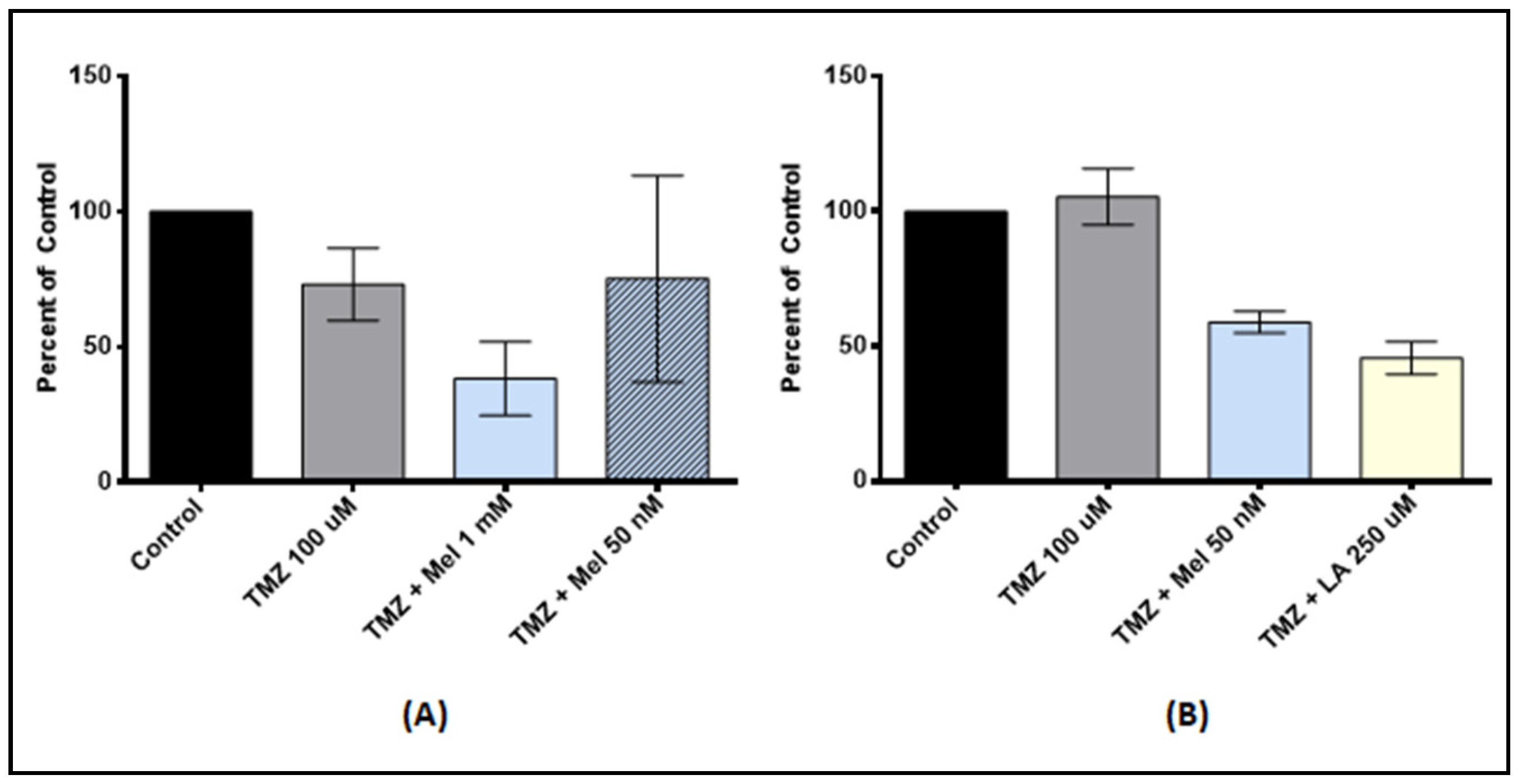
3.4. Effect of Anti-Oxidant and TMZ Co-Treatment on Proliferation
3.5. Antioxidant Pre-Treatment on Tumor Stem-Like Cells
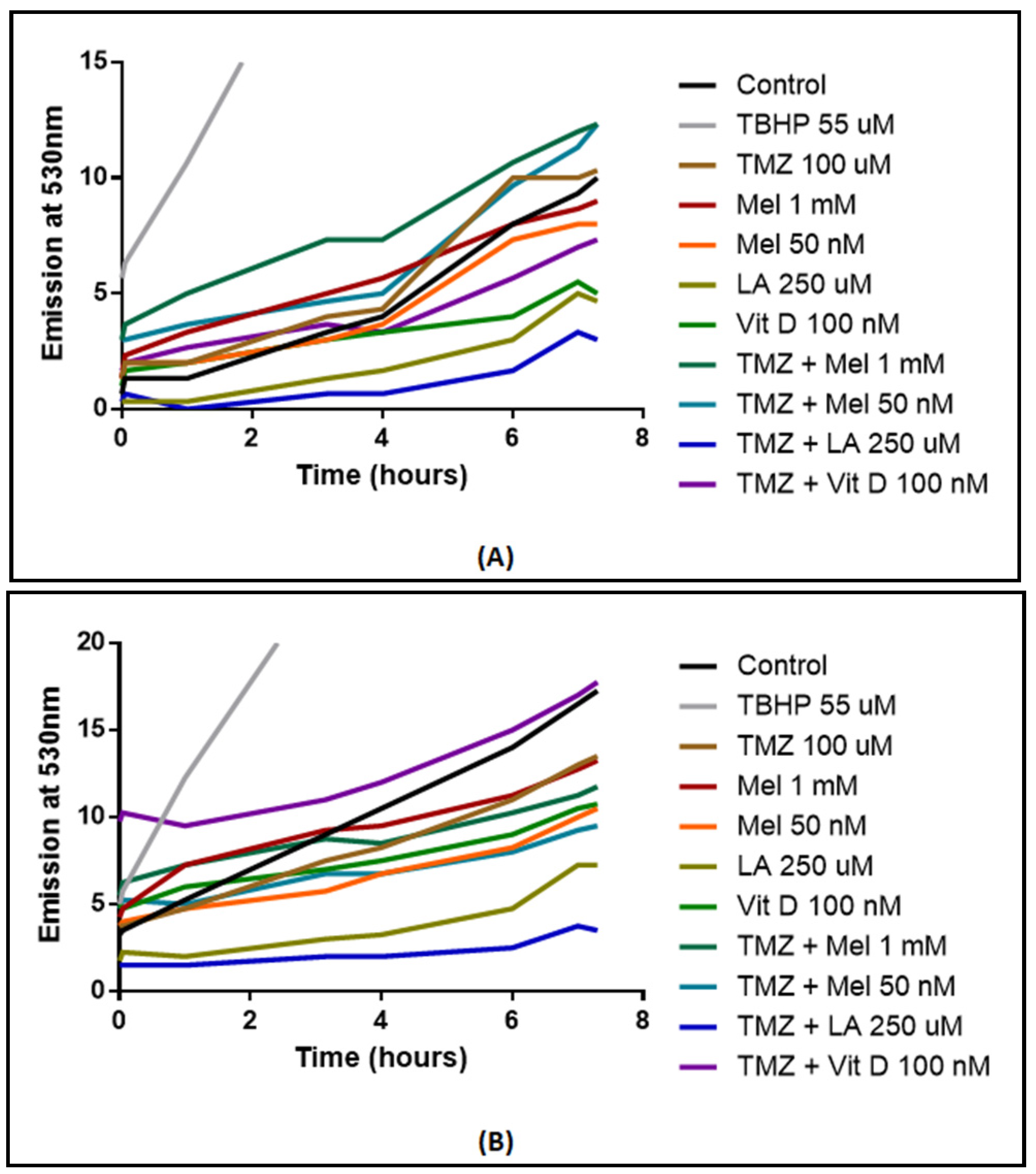
3.6. ROS Production
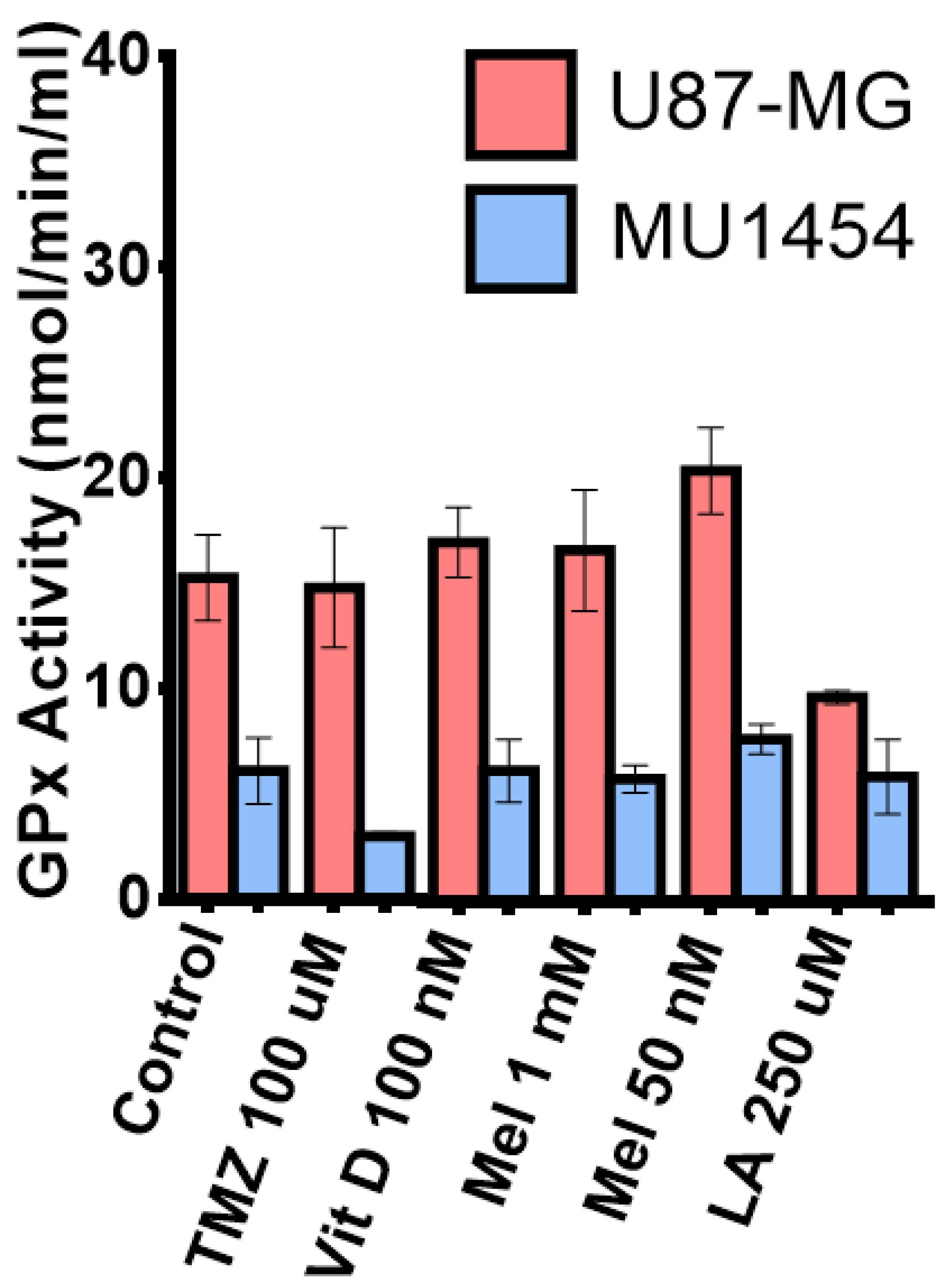
3.7. Glutathione Peroxidase Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gramatzki, D.; Dehler, S.; Rushing, E.J.; Zaugg, K.; Hofer, S.; Yonekawa, Y.; Bertalanffy, H.; Valavanis, A.; Korol, D.; Rohrmann, S.; et al. Glioblastoma in the canton of Zurich, Switzerland revisited: 2005 to 2009. Cancer 2016, 122, 2206–2215. [Google Scholar] [CrossRef] [PubMed]
- Affronti, M.L.; Heery, C.R.; Herndon, J.E., 2nd; Rich, J.N.; Reardon, D.A.; Desjardins, A.; Vredenburgh, J.J.; Friedman, A.H.; Bigner, D.D.; Friedman, H.S. Overall survival of newly diagnosed glioblastoma patients receiving carmustine wafers followed by radiation and concurrent temozolomide plus rotational multiagent chemotherapy. Cancer 2009, 115, 3501–3511. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Hegi, M.E.; Mason, W.P.; van den Bent, M.J.; Taphoorn, M.J.; Janzer, R.C.; Ludwin, S.K.; Allgeier, A.; Fisher, B.; Belanger, K.; et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase iii study: 5-year analysis of the eortc-ncic trial. Lancet Oncol. 2009, 10, 459–466. [Google Scholar] [CrossRef]
- deSouza, R.M.; Shaweis, H.; Han, C.; Sivasubramaniam, V.; Brazil, L.; Beaney, R.; Sadler, G.; Al-Sarraj, S.; Hampton, T.; Logan, J.; et al. Has the survival of patients with glioblastoma changed over the years? Br. J. Cancer 2016, 114, e20. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Taillibert, S.; Kanner, A.; Read, W.; Steinberg, D.; Lhermitte, B.; Toms, S.; Idbaih, A.; Ahluwalia, M.S.; Fink, K.; et al. Effect of tumor-treating fields plus maintenance temozolomide vs maintenance temozolomide alone on survival in patients with glioblastoma: A randomized clinical trial. JAMA 2017, 318, 2306–2316. [Google Scholar] [CrossRef] [PubMed]
- Friedman, H.S.; Kerby, T.; Calvert, H. Temozolomide and treatment of malignant glioma. Clin. Cancer Res. 2000, 6, 2585–2597. [Google Scholar] [PubMed]
- Lexi-Comp Online. Temozolomide: Drug information. In Lexi-Comp Online; 2018; Available online: https://online.lexi.com/lco/action/home (accessed on 15 May 2018).
- Lee, S.Y. Temozolomide resistance in glioblastoma multiforme. Genes Dis. 2016, 3, 198–210. [Google Scholar] [CrossRef]
- Fox, S.; Laws, E.R., Jr.; Anderson, F., Jr.; Farace, E. Complementary therapy use and quality of life in persons with high-grade gliomas. J. Neurosci. Nurs. 2006, 38, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Lawenda, B.D.; Kelly, K.M.; Ladas, E.J.; Sagar, S.M.; Vickers, A.; Blumberg, J.B. Should supplemental antioxidant administration be avoided during chemotherapy and radiation therapy? J. Natl. Cancer Inst. 2008, 100, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Ramiro, A.; Ramirez-Ortega, D.; Perez de la Cruz, V.; Hernandez-Pedro, N.Y.; Gonzalez-Esquivel, D.F.; Sotelo, J.; Pineda, B. Role of redox status in development of glioblastoma. Front. Immunol. 2016, 7, 156. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, M.; Caffo, M.; Minutoli, L.; Marini, H.; Abbritti, R.V.; Squadrito, F.; Trichilo, V.; Valenti, A.; Barresi, V.; Altavilla, D.; et al. Ros and brain gliomas: An overview of potential and innovative therapeutic strategies. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Sainz, R.M.; Antolin, I.; Mayo, J.C.; Herrera, F.; Rodriguez, C. Several antioxidant pathways are involved in astrocyte protection by melatonin. J. Pineal Res. 2002, 33, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Ladas, E.J.; Jacobson, J.S.; Kennedy, D.D.; Teel, K.; Fleischauer, A.; Kelly, K.M. Antioxidants and cancer therapy: A systematic review. J. Clin. Oncol. 2004, 22, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Mahoui, I. Targeting brain tumor stem cells through natural antioxidants. In American Junior Academy of Sciences Poster Session; American Association for Cancer Research: San Jose, CA, USA, 2015. [Google Scholar]
- Pinskey, J.M. Effects of 1α, 25-Dihydroxyvitamin D3 (Calcitriol) on Cultured Glioblastoma Multiforme Cells; Northern Michigan University: Marquette, MI, USA, 2012. [Google Scholar]
- Yuan, Y.; Xue, X.; Guo, R.B.; Sun, X.L.; Hu, G. Resveratrol enhances the antitumor effects of temozolomide in glioblastoma via ros-dependent ampk-tsc-mtor signaling pathway. CNS Neurosci. Ther. 2012, 18, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Bak, D.H.; Kang, S.H.; Choi, D.R.; Gil, M.N.; Yu, K.S.; Jeong, J.H.; Lee, N.S.; Lee, J.H.; Jeong, Y.G.; Kim, D.K.; et al. Autophagy enhancement contributes to the synergistic effect of vitamin d in temozolomide-based glioblastoma chemotherapy. Exp. Ther. Med. 2016, 11, 2153–2162. [Google Scholar] [CrossRef] [PubMed]
- Diesel, B.; Radermacher, J.; Bureik, M.; Bernhardt, R.; Seifert, M.; Reichrath, J.; Fischer, U.; Meese, E. Vitamin D-3 metabolism in human glioblastoma multiforme: Functionality of cyp27b1 splice variants, metabolism of calcidiol and effect of calcitriol. Clin. Cancer Res. 2005, 11, 5370–5380. [Google Scholar] [CrossRef] [PubMed]
- Na, M.H.; Seo, E.Y.; Kim, W.K. Effects of alpha-lipoic acid on cell proliferation and apoptosis in mda-mb-231 human breast cells. Nutr. Res. Pract. 2009, 3, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Q.; Wang, F.; Ling, E.A.; Liu, S.; Wang, L.; Yang, Y.; Yao, L.; Chen, X.; Wang, F.; et al. Melatonin antagonizes hypoxia-mediated glioblastoma cell migration and invasion via inhibition of hif-1alpha. J. Pineal Res. 2013, 55, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Dovinova, I.; Novotny, L.; Rauko, P.; Kvasnicka, P. Combined effect of lipoic acid and doxorubicin in murine leukemia. Neoplasma 1999, 46, 237–241. [Google Scholar] [PubMed]
- Huckabee, S. The Effect of R-alpha Lipoic Acid on Edelfosine’s Ability to Toxically Kill s. Cerevisae; 543rd Annual Ozarks Science and Engineering Fair: Springfield, MI, USA, 2013. [Google Scholar]
- Carmichael, J.; DeGraff, W.G.; Gazdar, A.F.; Minna, J.D.; Mitchell, J.B. Evaluation of a tetrazolium-based semiautomated colorimetric assay: Assessment of chemosensitivity testing. Cancer Res. 1987, 47, 936–942. [Google Scholar] [PubMed]
- Reichrath, S.; Müller, C.S.; Gleissner, B.; Pfreundschuh, M.; Vogt, T.; Reichrath, J. Notch- and vitamin D signaling in 1,25(OH)2D3-resistant glioblastoma multiforme (GBM) cell lines. J. Steroid Biochem. Mol. Biol. 2010, 121, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Van Ginkel, P.R.; Yang, W.; Marcet, M.M.; Chow, C.C.; Kulkarni, A.D.; Darjatmoko, S.; Lindstrom, M.J.; Lokken, J.; Bhattacharya, S.; Albert, D.M. 1 alpha-hydroxyvitamin D2 inhibits growth of human neuroblastoma. J. Neurooncol. 2007, 85, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Magrassi, L.; Butti, G.; Pezzotta, S.; Infuso, L.; Milanesi, G. Effects of vitamin d and retinoic acid on human glioblastoma cell lines. Acta Neurochir. 1995, 133, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, P.J. Vitamin D in cancer patients: Above all, do no harm. J. Clin. Oncol. 2009, 27, 2117–2119. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Sainz, R.M.; Mayo, J.C.; Lopez-Burillo, S. Melatonin: Reducing the toxicity and increasing the efficacy of drugs. J. Pharm. Pharmacol. 2002, 54, 1299–1321. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Hidalgo, M.; Guerrero, J.M.; Villegas, I.; Packham, G.; de la Lastra, C.A. Melatonin, a natural programmed cell death inducer in cancer. Curr. Med. Chem. 2012, 19, 3805–3821. [Google Scholar] [CrossRef] [PubMed]
- Galley, H.F.; Lowes, D.A.; Allen, L.; Cameron, G.; Aucott, L.S.; Webster, N.R. Melatonin as a potential therapy for sepsis: A phase I dose escalation study and an ex vivo whole blood model under conditions of sepsis. J. Pineal Res. 2014, 56, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Hao, A.; Li, X.; Du, Z.; Li, H.; Wang, H.; Yang, H.; Fang, Z. Melatonin inhibits tumorigenicity of glioblastoma stem-like cells via the AKT-EZH2-STAT3 signaling axis. J. Pineal Res. 2016, 61, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Jardim-Perassi, B. What Is the Equivalent Concentration Melatonin for Use in Humans? Mezencev, R., Ed.; ResearchGate: Berlin, Germany, 2015. [Google Scholar]
- Martin, V.; Herrera, F.; Carrera-Gonzalez, P.; Garcia-Santos, G.; Antolin, I.; Rodriguez-Blanco, J.; Rodriguez, C. Intracellular signaling pathways involved in the cell growth inhibition of glioma cells by melatonin. Cancer Res. 2006, 66, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.M.; Blask, D.E. Effects of the pineal hormone melatonin on the proliferation and morphological characteristics of human breast cancer cells (mcf-7) in culture. Cancer Res. 1988, 48, 6121–6126. [Google Scholar] [PubMed]
- Cos, S.; Fernandez, R.; Guezmes, A.; Sanchez-Barcelo, E.J. Influence of melatonin on invasive and metastatic properties of mcf-7 human breast cancer cells. Cancer Res. 1998, 58, 4383–4390. [Google Scholar] [PubMed]
- Marelli, M.M.; Limonta, P.; Maggi, R.; Motta, M.; Moretti, R.M. Growth-inhibitory activity of melatonin on human androgen-independent du 145 prostate cancer cells. Prostate 2000, 45, 238–244. [Google Scholar] [CrossRef]
- Goder, A.; Nagel, G.; Kraus, A.; Dorsam, B.; Seiwert, N.; Kaina, B.; Fahrer, J. Lipoic acid inhibits the DNA repair protein o 6-methylguanine-DNA methyltransferase (mgmt) and triggers its depletion in colorectal cancer cells with concomitant autophagy induction. Carcinogenesis 2015, 36, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.B.; Wang, Z.; Shu, F.; Jin, Y.H.; Liu, H.Y.; Wang, Q.J.; Yang, Y. Activation of amp-activated protein kinase by temozolomide contributes to apoptosis in glioblastoma cells via p53 activation and mtorc1 inhibition. J. Biol. Chem. 2010, 285, 40461–40471. [Google Scholar] [CrossRef] [PubMed]
- Sak, K. Chemotherapy and Dietary Phytochemical Agents. Chemother. Res. Pract. 2012, 2012, 282570. [Google Scholar] [CrossRef] [PubMed]
- Glasauer, A.; Chandel, N.S. Targeting antioxidants for cancer therapy. Biochem. Pharmacol. 2014, 92, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Kuban-Jankowska, A.; Gorska-Ponikowska, M.; Wozniak, M. Lipoic acid decreases the viability of breast cancer cells and activity of PTP1B AND SHP2. Anticancer Res. 2017, 37, 2893–2898. [Google Scholar] [CrossRef] [PubMed]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yi, J. Cancer cell killing via ros: To increase or decrease, that is the question. Cancer Biol. Ther. 2008, 7, 1875–1884. [Google Scholar] [CrossRef] [PubMed]
- Hawk, M.A.; McCallister, C.; Schafer, Z.T. Antioxidant activity during tumor progression: A necessity for the survival of cancer cells? Cancers 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Dokic, I.; Hartmann, C.; Herold-Mende, C.; Regnier-Vigouroux, A. Glutathione peroxidase 1 activity dictates the sensitivity of glioblastoma cells to oxidative stress. Glia 2012, 60, 1785–1800. [Google Scholar] [CrossRef] [PubMed]
- Rocha, C.R.; Kajitani, G.S.; Quinet, A.; Fortunato, R.S.; Menck, C.F. NRF2 and glutathione are key resistance mediators to temozolomide in glioma and melanoma cells. Oncotarget 2016, 7, 48081–48092. [Google Scholar] [CrossRef] [PubMed]
- Koren, R.; Hadari-Naor, I.; Zuck, E.; Rotem, C.; Liberman, U.A.; Ravid, A. Vitamin D is a prooxidant in breast cancer cells. Cancer Res. 2001, 61, 1439–1444. [Google Scholar] [PubMed]
- Valdecantos, M.P.; Perez-Matute, P.; Quintero, P.; Martinez, J.A. Vitamin C, resveratrol and lipoic acid actions on isolated rat liver mitochondria: All antioxidants but different. Redox Rep. 2010, 15, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Damnjanovic, I.; Kocic, G.; Najman, S.; Stojanovic, S.; Stojanovic, D.; Veljkovic, A.; Conic, I.; Langerholc, T.; Pesic, S. Chemopreventive potential of alpha lipoic acid in the treatment of colon and cervix cancer cell lines. Bratisl. Lek. Listy 2014, 115, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Manik, C. Mgmt modulates glioblastoma angiogenesis and response to the tyrosine kinase inhibitor sunitinib. Neuro Oncol. 2010, 12, 822–833. [Google Scholar]
- Lissa, N.M. Mechanisms of chemoresistance in cancer stem cells. Clin. Transl. Med. 2013, 2, 3. [Google Scholar]
- So, J.Y.; Suh, N. Targeting cancer stem cells in solid tumors by vitamin D. J. Steroid Biochem. Mol. Biol. 2015, 148, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Akbardazeh, M. The potential therapeutic effect of melatonin on human ovarian cancer by inhibition of invasion and migration of cancer stem cells. Sci. Rep. 2017, 7, 17062. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MTT U87-MG | MTT MU1454 | MTT scU87-MG | MTT scMU1454 | ROS Activity U87-MG | ROS Activity MU1454 | GP×1 Activity U87-MG | GP×1 Activity MU1454 | |
|---|---|---|---|---|---|---|---|---|
| Vitamin D3 100 nM | ↓*** | ↓* | - | - | ↓** | - | - | - |
| Vitamin D3 10 nM | ↓*** | ↔ | - | - | - | - | - | - |
| Vitamin D3 1 nM | ↓*** | ↔ | - | - | - | - | - | - |
| Vitamin D3 100 nM + TMZ serial | ↓*** | ↓* | - | - | - | - | - | - |
| Vitamin D3 10 nM + TMZ serial | ↓*** | ↔ | - | - | - | - | - | - |
| Vitamin D3 1 nM +TMZ serial | ↔ | ↔ | - | - | - | - | - | - |
| Vitamin D3 100 nM + TMZ co-treatment | - | - | - | - | ↔ | ↑* | ↓* | ↔ |
| Melatonin 1 mM | ↓**** | ↓**** | - | - | ↔ | ↔ | ↔ | ↔ |
| Melatonin 1 µM | ↔ | ↓*** | - | - | - | - | - | - |
| Melatonin 50 nM | ↔ | ↓*** | - | - | ↔ | ↓** | ↑* | ↔ |
| Melatonin 1 mM + TMZ serial | ↓**** | ↓**** | ↓*** | - | - | - | - | - |
| Melatonin 1 µM + TMZ serial | ↔ | ↓**** | - | - | - | - | - | - |
| Melatonin 50 nM + TMZ serial | ↓**** | ↓**** | ↔ | ↓**** | - | - | - | - |
| Melatonin 1 mM + TMZ co-treatment | ↓*** | ↓*** | - | - | ↔ | ↔ | ↔ | ↔ |
| Melatonin 50 nM + TMZ co-treatment | - | - | - | - | ↔ | ↔ | ↑** | ↔ |
| Lipoic Acid 1mM | ↓**** | ↓*** | - | - | - | - | - | - |
| Lipoic Acid 250 µM | ↓**** | ↓**** | - | - | ↓** | ↓*** | ↓** | ↔ |
| Lipoic Acid 100 nM | ↓*** | ↓** | - | - | - | - | - | - |
| Lipoic Acid 1mM + TMZ serial | ↓**** | ↓**** | - | - | - | - | - | - |
| Lipoic Acid 250 µM + TMZ serial | ↓**** | ↓*** | - | - | - | - | - | - |
| Lipoic Acid 100 nM + TMZ serial | ↓*** | ↓*** | - | - | - | - | - | - |
| Lipoic acid 250 µM + TMZ co-treatment | ↓*** | ↓*** | - | - | ↓**** | ↓**** | ↑* | ↔ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McConnell, D.D.; McGreevy, J.W.; Williams, M.N.; Litofsky, N.S. Do Anti-Oxidants Vitamin D3, Melatonin, and Alpha-Lipoic Acid Have Synergistic Effects with Temozolomide on Cultured Glioblastoma Cells? Medicines 2018, 5, 58. https://doi.org/10.3390/medicines5020058
McConnell DD, McGreevy JW, Williams MN, Litofsky NS. Do Anti-Oxidants Vitamin D3, Melatonin, and Alpha-Lipoic Acid Have Synergistic Effects with Temozolomide on Cultured Glioblastoma Cells? Medicines. 2018; 5(2):58. https://doi.org/10.3390/medicines5020058
Chicago/Turabian StyleMcConnell, Diane D., Joe W. McGreevy, Macy N. Williams, and N. Scott Litofsky. 2018. "Do Anti-Oxidants Vitamin D3, Melatonin, and Alpha-Lipoic Acid Have Synergistic Effects with Temozolomide on Cultured Glioblastoma Cells?" Medicines 5, no. 2: 58. https://doi.org/10.3390/medicines5020058
APA StyleMcConnell, D. D., McGreevy, J. W., Williams, M. N., & Litofsky, N. S. (2018). Do Anti-Oxidants Vitamin D3, Melatonin, and Alpha-Lipoic Acid Have Synergistic Effects with Temozolomide on Cultured Glioblastoma Cells? Medicines, 5(2), 58. https://doi.org/10.3390/medicines5020058