
Essential Oils and Their Components as Modulators of Antibiotic Activity against Gram-Negative Bacteria

 ,
,  ,
,
Abstract
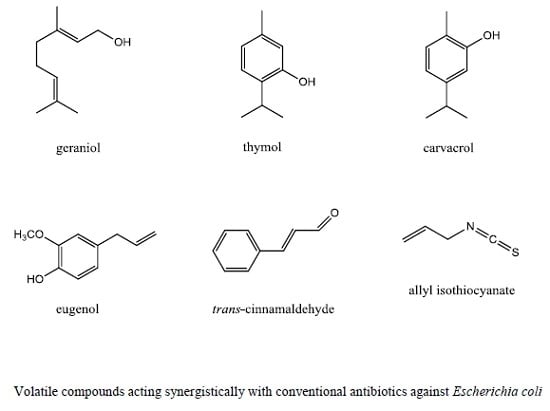
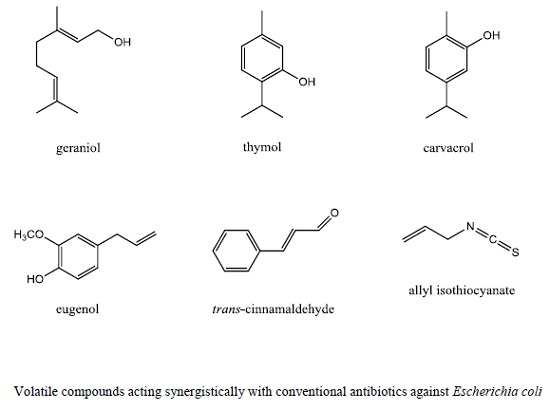
:
1. Introduction
2. Essential Oils and Volatile Compounds Modulating Antibiotic Activity against Acinetobacter baumannii
3. Essential Oils and Volatile Compounds Modulating Antibiotic Activity against Escherichia coli
4. Essential Oils and Volatile Compounds Modulating Antibiotic Activity against Klebsiella pneumoniae
5. Essential Oils and Volatile Compounds Modulating Antibiotic Activity against Pseudomonas aeruginosa
6. Essential Oils and Volatile Compounds Modulating Antibiotic Activity against Other Gram-Negative Bacteria
7. Conclusions
Author Contributions
Conflicts of Interest
References
- Abat, C.; Chaudet, H.; Rolain, J.M.; Colson, P.; Raoult, D. Traditional and syndromic surveillance of infectious diseases and pathogens. Int. J. Infect. Dis. 2016, 48, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Van Vuuren, S.F.; Suliman, S.; Viljoen, A.M. The antimicrobial activity of four commercial essential oils in combination with conventional antimicrobials. Lett. Appl. Microbiol. 2009, 48, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Summary of the Latest Data on Antibiotic Resistance in the European Union. Available online: http://ecdc.europa.eu/en/eaad/Documents/antibiotics-resistance-EU-data-2013.pdf (accessed on 10 June 2016).
- Wolska, K.I.; Grześ, K.; Kurek, A. Synergy between novel antimicrobials and conventional antibiotics or bacteriocins. Pol. J. Microbiol. 2012, 61, 95–104. [Google Scholar] [PubMed]
- Elshafie, H.S.; Mancini, E.; Sakr, S.; de Martino, L.; Mattia, C.A.; de Feo, V.; Camele, I. Antifungal activity of some constituents of Origanum vulgare L. essential oil against postharvest disease of peach fruit. J. Med. Food 2015, 18, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Mancini, E.; Camele, I.; Elshafie, H.S.; de Martino, L.; Pellegrino, C.; Grulova, D.; de Feo, V. Chemical composition and biological activity of the essential oil of Origanum vulgare ssp. hirtum from different areas in the Southern Apennines (Italy). Chem. Biodivers. 2014, 11, 639–651. [Google Scholar] [CrossRef] [PubMed]
- Van Vuuren, S.; Viljoen, A. Plant-based antimicrobial studies—Methods and approaches to study the interaction between natural products. Planta Med. 2011, 77, 1168–1182. [Google Scholar] [CrossRef] [PubMed]
- Bassolé, I.H.; Juliani, H.R. Essential oils in combination and their antimicrobial properties. Molecules 2012, 17, 3989–4006. [Google Scholar] [CrossRef] [PubMed]
- Basri, D.F.; Xian, L.W.; Abdul Shukor, N.I.; Latip, J. Bacteriostatic antimicrobial combination: Antagonistic interaction between epsilon-viniferin and vancomycin against methicillin-resistant Staphylococcus aureus. Biomed. Res. Int. 2014. [Google Scholar] [CrossRef] [PubMed]
- Langeveld, W.T.; Veldhuizen, E.J.; Burt, S.A. Synergy between essential oil components and antibiotics: A review. Crit. Rev. Microbiol. 2014, 40, 76–94. [Google Scholar] [CrossRef] [PubMed]
- Kyaw, B.M.; Arora, S.; Lim, C.S. Bactericidal antibiotic-phytochemical combinations against methicillin resistant Staphylococcus aureus. Braz. J. Microbiol. 2012, 43, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.E.; Kim, H.Y.; Cha, J.D. Synergistic effect between clove oil and its major compounds and antibiotics against oral bacteria. Arch. Oral. Biol. 2011, 56, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003. [Google Scholar] [CrossRef] [PubMed]
- Hendry, E.R.; Worthington, T.; Conway, B.R.; Lambert, P.A. Antimicrobial efficacy of eucalyptus oil and 1,8-cineole alone and in combination with chlorhexidine digluconate against microorganisms grown in planktonic and biofilm cultures. J. Antimicrob. Chemother. 2009, 64, 1219–1225. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, S. Phytochemicals for bacterial resistance—Strengths, weaknesses and opportunities. Planta Med. 2008, 74, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Kourtesi, C.; Ball, A.R.; Huang, Y.Y.; Jachak, S.M.; Vera, D.M.; Khondkar, P.; Gibbons, S.; Hamblin, M.R.; Tegos, G.P. Microbial efflux systems and inhibitors: Approaches to drug discovery and the challenge of clinical implementation. Open Microbiol. J. 2013, 7, 34–52. [Google Scholar] [CrossRef] [PubMed]
- Garvey, M.I.; Rahman, M.M.; Gibbons, S.; Piddock, L.J. Medicinal plant extracts with efflux inhibitory activity against Gram-negative bacteria. Int. J. Antimicrob. Agents 2011, 37, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Hemaiswarya, S.; Kruthiventi, A.K.; Doble, M. Synergism between natural products and antibiotics against infectious diseases. Phytomedicine 2008, 15, 639–652. [Google Scholar] [CrossRef] [PubMed]
- Cole, E.R.; dos Santos, R.B.; Lacerda Júnior, V.; Martins, J.; Greco, S.; Cunha Neto, A. Chemical composition of essential oil from ripe fruit of Schinus terebinthifolius raddi and evaluation of its activity against wild strains of hospital origin. Braz. J. Microbiol. 2014, 45, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, H.S.; Ghanney, N.; Mang, S.M.; Ferchichi, A.; Camele, I. An in vitro attempt for controlling severe phytopathogens and human pathogens using essential oils from mediterranean plants of genus schinus. J. Med. Food 2016, 19, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Howard, A.; O’Donoghue, M.; Feeney, A.; Sleator, R.D. Acinetobacter baumannii: An emerging opportunistic pathogen. Virulence 2012, 3, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.C.; Gajbhiye, S.R.; Agrawal, G.N. Clinical and antimicrobial profile of Acinetobacter spp.: An emerging nosocomial superbug. Adv. Biomed. Res. 2014. [Google Scholar] [CrossRef] [PubMed]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [PubMed]
- Aleksic, V.; Dukic, N.M.; Simin, N.; Nedeljkovic, N.S.; Knezevic, P. Synergistic effect of Myrtus communis L. essential oils and conventional antibiotics against multi-drug resistant Acinetobacter baumannii wound isolates. Phytomedicine 2014, 21, 1666–1674. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, P.; Aleksic, V.; Simin, N.; Svircev, E.; Petrovic, A.; Mimica-Dukic, N. Antimicrobial activity of Eucalyptus camaldulensis essential oils and their interactions with conventional antimicrobial agents against multi-drug resistant Acinetobacter baumannii. J. Ethnopharmacol. 2016, 178, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.C.; Hood, I.; Boyd, K.L.; Olson, P.D.; Morrison, J.M.; Carson, S.; Sayood, K.; Iwen, P.C.; Skaar, E.P.; Dunman, P.M. Inactivation of phospholipase D diminishes Acinetobacter baumannii pathogenesis. Infect. Immun. 2010, 78, 1952–1962. [Google Scholar] [CrossRef] [PubMed]
- Camarena, L.; Bruno, V.; Euskirchen, G.; Poggio, S.; Snyder, M. Molecular mechanisms of ethanol-induced patho-genesis revealed by RNA-sequencing. PLoS Pathog. 2010. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Price, S.; Weinstein, R.A. Acinetobacter Infection. N. Engl. J. Med. 2008, 358, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Doi, Y.; Husain, S.; Potoski, B.A.; McCurry, K.R.; Paterson, D.L. Extensively drug-resistant Acinetobacter baumannii. Emerg. Infect. Dis. 2009, 15, 980–982. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.; Ferreira, S.; Silva, F.; Domingues, F.C. Synergistic activity of coriander oil and conventional antibiotics against Acinetobacter baumannii. Phytomedicine 2012, 19, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Ferreira, S.; Queiroz, J.A.; Domingues, F.C. Coriander (Coriandrum sativum L.) essential oil: Its antibacterial activity and mode of action evaluated by flow cytometry. J. Med. Microbiol. 2011, 60, 1479–1486. [Google Scholar] [CrossRef] [PubMed]
- Prabuseenivasan, S.; Jayakumar, M.; Ignacimuthu, S. In vitro antibacterial activity of some plant essential oils. BMC Complement. Altern. Med. 2006. [Google Scholar] [CrossRef] [PubMed]
- Zaika, L.L. Spices and herbs: Their antibacterial activity and its determination. J. Food Saf. 1988, 23, 97–118. [Google Scholar]
- Rosato, A.; Piarulli, M.; Corbo, F.; Muraglia, M.; Carone, A.; Vitali, M.E.; Vitali, C. In vitro synergistic antibacterial action of certain combinations of gentamicin and essential oils. Curr. Med. Chem. 2010, 17, 3289–3295. [Google Scholar] [CrossRef] [PubMed]
- Tam, V.H.; Schilling, A.N.; Vo, G.; Kabbara, S.; Kwa, A.L.; Wiederhold, N.P.; Lewis, R.E. Pharmacodynamics of polymyxin B against Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2005, 49, 3624–3630. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Romero, J.C.; González-Ríos, H.; Borges, A.; Simões, M. Antibacterial effects and mode of action of selected essential oils components against Escherichia coli and Staphylococcus aureus. Evid. Based Complement. Altern. Med. 2015. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, V.; Muselli, A.; Bernardini, A.F.; Berti, L.; Pagès, J.M.; Amaral, L.; Bolla, J.M. Geraniol restores antibiotic activities against multidrug-resistant isolates from Gram-negative species. Antimicrob. Agents Chemother. 2009, 53, 2209–2211. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, M.M.; Ramalho, P.; Silva, A.P.; Teixeira-Santos, R.; Pina-Vaz, C.; Rodrigues, A.G. Polyethyleneimine and polyethyleneimine-based nanoparticles: Novel bacterial and yeast biofilm inhibitors. J. Med. Microbiol. 2014, 63, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, Z.W.; Yin, Z.Q.; Wei, Q.; Jia, R.Y.; Zhou, L.J.; Xu, J.; Song, X.; Zhou, Y.; Du, Y.H.; et al. Antibacterial activity of leaf essential oil and its constituents from Cinnamomum longepaniculatum. Int. J. Clin. Exp. Med. 2014, 7, 1721–1727. [Google Scholar] [PubMed]
- Guerra, F.Q.; Mendes, J.M.; Sousa, J.P.; Morais-Braga, M.F.; Santos, B.H.; Coutinho, M.H.D.; Lima, E.O. Increasing antibiotic activity against a multidrug-resistant Acinetobacter spp. by essential oils of Citrus limon and Cinnamomum zeylanicum. Nat. Prod. Res. 2012, 26, 2235–2238. [Google Scholar] [CrossRef] [PubMed]
- Darfeuille-Michaud, A.; Boudeau, J.; Bulois, P.; Neut, C.; Glasser, A.L.; Barnich, N.; Bringer, M.A.; Swidsinski, A.; Beaugerie, L.; Colombel, J.F. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn’s disease. Gastroenterology 2004, 127, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Allocati, N.; Masulli, M.; Alexeyev, M.F.; di Ilio, C. Escherichia coli in Europe: An overview. Int. J. Environ. Res. Public Health 2013, 10, 6235–6254. [Google Scholar] [CrossRef] [PubMed]
- Croxen, M.A.; Finlay, B.B. Molecular mechanisms of Escherichia coli pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar] [PubMed]
- Johnson, J.R.; Stell, A.L. Extended virulence genotypes of Escherichia coli strains from patients with urosepsis in relation to phylogeny and host compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Qadri, F.; Svennerholm, A.M.; Faruque, A.S.; Sack, R.B. Enterotoxigenic Escherichia coli in developing countries: Epidemiology, microbiology, clinical features, treatment, and prevention. Clin. Microbiol. Rev. 2005, 18, 465–483. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Bonnin, R.A.; Nordmann, P. Genetic support and diversity of acquired extended-spectrum β-lactamases in Gram-negative rods. Infect. Genet. Evol. 2012, 12, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile beta-lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [PubMed]
- Robicsek, A.; Jacoby, G.A.; Hooper, D.C. The worldwide emergence of plasmid-mediated quinolone resistance. Lancet Infect. Dis. 2006, 6, 629–640. [Google Scholar] [CrossRef]
- Si, H.; Hu, J.; Liu, Z.; Zeng, Z.L. Antibacterial effect of oregano essential oil alone and in combination with antibiotics against extended-spectrum beta-lactamase-producing Escherichia coli. FEMS Immunol. Med. Microbiol. 2008, 53, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Okusu, H.; Ma, D.; Nikaido, H. AcrAB efflux pump plays a major role in the antibiotic resistance phenotype of Escherichia coli multiple-antibiotic-resistance (Mar) mutants. J. Bacteriol. 1996, 178, 306–308. [Google Scholar] [PubMed]
- Fadli, M.; Chevalier, J.; Saad, A.; Mezrioui, N.E.; Hassani, L.; Pagès, J.M. Essential oils from Moroccan plants as potential chemosensitisers restoring antibiotic activity in resistant Gram-negative bacteria. Int. J. Antimicrob. Agents 2011, 38, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhou, F.; Ji, B.P.; Pei, R.S.; Xu, N. The antibacterial mechanism of carvacrol and thymol against Escherichia coli. Lett. Appl. Microbiol. 2008, 47, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Helander, I.M.; Alakomi, H.L.; Latva-Kala, K.; Mattila-Sandholm, T.; Pol, I.; Smid, E.J.; Gorris, L.G.M.; von Wright, A. Characterization of the action of selected essential oil components on Gram-negative bacteria. J. Agric. Food Chem. 1998, 46, 3590–3595. [Google Scholar] [CrossRef]
- Ultee, A.; Bennik, M.H.; Moezelaar, R. The phenolic hydroxyl group of carvacrol is essential for action against the food-borne pathogen Bacillus cereus. Appl. Environ. Microbiol. 2002, 68, 1561–1568. [Google Scholar] [CrossRef] [PubMed]
- Fadli, M.; Saada, A.; Sayadib, S.; Chevalierc, J.; Mezriouia, N.E.; Pagès, J.M.; Hassani, L. Antibacterial activity of Thymus maroccanus and Thymus broussonetii essential oils against nosocomial infection-bacteria and their synergistic potential with antibiotics. Phytomedicine 2012, 19, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Fadli, M.; Bollab, J.M.; Mezriouia, N.E.; Pagès, J.M.; Hassania, L. First evidence of antibacterial and synergistic effects of Thymus riatarum essential oil with conventional antibiotics. Ind. Crop. Prod. 2014, 61, 370–376. [Google Scholar] [CrossRef]
- Guinoiseau, E.; Lorenzi, V.; Luciani, A.; Muselli, A.; Costa, J.; Casanova, J.; Berti, L. Biological properties and resistance reversal effect of Helichrysum italicum (Roth) G. Don. Formatex 2013, 2, 1073–1080. [Google Scholar]
- Miladinović, D.L.; Ilić, B.S.; Kocić, B.D.; Miladinović, M.D. An in vitro antibacterial study of savory essential oil and geraniol in combination with standard antimicrobials. Nat. Prod. Commun. 2014, 9, 1629–1632. [Google Scholar] [PubMed]
- Barreto, H.M.; Silva Filho, E.C.; Lima, E.D.O.; Coutinho, H.D.M.; Morais Braga, M.F.B.; Tavares, C.C.A.; Tintinoc, S.R.; Rego, J.V.; de Abreu, A.P.L.; Lustosa Mdo, C.; et al. Chemical composition and possible use as adjuvant of the antibiotic therapy of the essential oil of Rosmarinus officinalis L. Ind. Crop. Prod. 2014, 59, 290–294. [Google Scholar] [CrossRef]
- Costa-Lotufo, L.V.; Araújo, E.C.; Lima, M.A.; Moraes, M.E.; Pessoa, C.; Silviera, E.R.; Moraes, M.O. Antiproliferative effects of abietane diterpenoids isolated from Hyptis martiusii Benth (Labiatae). Pharmazie 2004, 59, 78–79. [Google Scholar] [PubMed]
- Caldas, G.F.; do Amaral Costa, I.M.; da Silva, J.B.; da Nóbrega, R.F.; Rodrigues, F.F.; da Costa, J.G.; Wanderley, A.G. Antiulcerogenic activity of the essential oil of Hyptis martiusii Benth. (Lamiaceae). J. Ethnopharmacol. 2011, 137, 886–892. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.D.; Galvao Rodrigue, F.F.; Coutinho, D.M.H.; da Costa, J.G.; de Menezes, I.R. Chemical composition, modulatory bacterial resistance and antimicrobial activity of essential oil the Hyptis martiusii benth. by direct and gaseous contact. Jundishapur J. Nat. Pharm. Prod. 2014, 9, e13521. [Google Scholar] [CrossRef] [PubMed]
- Gallucci, M.N.; Casero, C.; de las Mercedes, M.; Demo, M.S. Interaction between terpenes and penicillin on bacterial strains resistant to beta-lactam antibiotics. Mol. Med. Chem. 2006, 10, 30–32. [Google Scholar]
- Palaniappan, K.; Holley, R.A. Use of natural antimicrobials to increase antibiotic susceptibility of drug resistant bacteria. Int. J. Food. Microbiol. 2010, 140, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Ben Arfa, A.; Combes, S.; Preziosi-Belloy, L.; Gontard, N.; Chalier, P. Antimicrobial activity of carvacrol related to its chemical structure. Lett. Appl. Microbiol. 2006, 43, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Di Pasqua, R.; Hoskins, N.; Betts, G.; Mauriello, G. Changes in membrane fatty acids composition of microbial cells induced by addiction of thymol, carvacrol, limonene, cinnamaldehyde, and eugenol in the growing media. J. Agric. Food Chem. 2006, 54, 2745–2749. [Google Scholar] [CrossRef] [PubMed]
- Novak, J.; Zitterl-Eglseer, K.; Deans, S.G.; Franz, C.M. Essential oils of different cultivars of Cannabis sativa L. and their antimicrobial activity. Flavour Fragr. J. 2001, 16, 259–262. [Google Scholar] [CrossRef]
- Bernotienë, G.; Nivinskienë, O.; Butkienë, R.; Mockutë, D. Chemical composition of essential oil of hops (Humulus lupulus L.) growing in Aukstaitija. Chemija 2004, 2, 31–36. [Google Scholar]
- Alves, R.J.V.; Medeiros, D.; de Moura, R.L.; de Gusmão, L.C.T.; da Silva, N.G.; Antunes, K.; Matozinhos, C.; Blanco, M.A. First report of naturalization of Houttuynia cordata Thunb. 1783 (Piperales: Saururaceae) in South America. Check List 2013, 9, 642–644. [Google Scholar] [CrossRef]
- Dai, D.N.; Thang, T.D.; Ogunmoye, A.R.; Olanrewaju, I.A.; Ogunwande, I.A. Chemical constituents of essential oils from the leaves of Tithonia diversifolia, Houttuynia cordata and Asarum glabrum grown in Vietnam. Am. J. Essent. Oils Nat. Prod. 2015, 2, 17–21. [Google Scholar]
- Tajidin, N.E.; Ahmad, S.H.; Rosenani, A.B.; Azimah, H.; Munirah, M. Chemical composition and citral content in lemongrass (Cymbopogon citratus) essential oil at three maturity stages. Afr. J. Biotechnol. 2012, 7, 2685–2693. [Google Scholar] [CrossRef]
- Miladinović, D.L.; Ilic, B.S.; Miladinovic, L.C.; Kocic, B.D.; Ciric, V.M.; Stankov-Jovanovic, V.P.; Cvetkovic, O.G. Antibacterial activity of Thymus pulegioides essential oil and its synergistic potential with antibiotics: A chemometric approach. In Recent Progress in Medicinal Plants, Essential Oils III and Phytopharmacology; Govil, J.N., Bhattacharya, S., Eds.; Studium Press LLC: Houston, TX, USA, 2013; pp. 101–136. [Google Scholar]
- Djouahri, A.; Saka, B.; Boudarene, L.; Benseradj, L.; Aberrane, S.; Aitmoussa, S.; Chelghoum, C.; Lamari, L.; Sabaou, L.; Baaliouame, A. In vitro synergistic/antagonistic antibacterial and anti-inflammatory effect of various extracts/essential oil from cones of Tetraclinis articulata (Vahl) Masters with antibiotic and anti-inflammatory agents. Ind. Crop. Prod. 2014, 56, 60–66. [Google Scholar] [CrossRef]
- Kasrati, A.; Jamall, C.A.; Fadli, M.; Bekkouche, K.; Hassani, L.; Wohlmuth, H.; Leach, D.; Abbad, A. Antioxidative activity and synergistic effect of Thymus saturejoides Coss. essential oils with cefixime against selected food-borne bacteria. Ind. Crop. Prod. 2014, 61, 338–344. [Google Scholar] [CrossRef]
- Brisse, S.; Grimont, F.; Grimont, P.A.D. The Genus Klebsiella. Prokaryotes 2006, 6, 159–196. [Google Scholar]
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods and pathogenicity factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [PubMed]
- Fung, C.P.; Chang, F.Y.; Lee, S.C.; Hu, B.S.; Kuo, B.I.; Liu, C.Y.; Ho, M.; Siu, L.K. A global emerging disease of Klebsiella pneumoniae liver abscess: Is serotype K1 an important factor for complicated endophthalmitis? Gut 2002, 50, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.H.; Kao, C.Y.; Yang, D.C.; Tseng, C.C.; Wu, A.B.; Teng, C.H.; Wang, M.C.; Wu, J.J. Clinical and microbiological characteristics of Klebsiella pneumoniae from community-acquired recurrent urinary tract infections. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1533–1539. [Google Scholar] [CrossRef] [PubMed]
- Ko, W.C.; Paterson, D.L.; Sagnimeni, A.J.; Hansen, D.S.; von Gottberg, A.; Mohapatra, S.; Casellas, J.M.; Goossens, H.; Mulazimoglu, L.; Trenholme, G.; et al. Community acquired Klebsiella pneumoniae bacteremia: Global differences in clinical patterns. Emerg. Infect. Dis. 2002, 8, 160–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Zhao, Y.; Liu, C.; Chen, Z.; Zhou, D. Molecular pathogenesis of Klebsiella pneumoniae. Future Microbiol. 2014, 9, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Vuotto, C.; Longo, F.; Balice, M.P.; Donelli, G.; Varaldo, P.E. Antibiotic resistance related to biofilm formation in Klebsiella pneumoniae. Pathogens 2014, 3, 743–758. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Cuzon, G.; Naas, T. The real threat of Klebsiella pneumoniae carbapenemase-producing bacteria. Lancet Infect. Dis. 2009, 9, 228–236. [Google Scholar] [CrossRef]
- Petrosillo, N.; Giannella, M.; Lewis, R.; Viale, P. Treatment of carbapenem-resistant Klebsiella pneumoniae: The state of the art. Expert Rev. Anti Infect. Ther. 2013, 11, 159–177. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Price, L.S.; Poirel, L.; Bonomo, R.A.; Schwaber, M.J.; Daikos, G.L.; Cormican, M.; Cornaglia, G.; Garau, J.; Gniadkowski, M.; Hayden, M.K.; et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect. Dis. 2013, 13, 785–796. [Google Scholar] [CrossRef]
- Parisi, S.G.; Bartolini, A.; Santacatterina, E.; Castellani, E.; Ghirardo, R.; Berto, A.; Franchin, E.; Menegotto, N.; de Canale, E.; Tommasini, T.; et al. Prevalence of Klebsiella pneumoniae strains producing carbapenemases and increase of resistance to colistin in an Italian teaching hospital from January 2012 to December 2014. BMC Infect. Dis. 2015. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, Z.A.; Paterson, D.L.; Potoski, B.A.; Kilayko, M.C.; Sandovsky, G.; Sordillo, E.; Polsky, B.; Adams-Haduch, J.M.; Doi, Y. Treatment outcome of bacteremia due to KPC-producing Klebsiella pneumoniae: Superiority of combination antimicrobial regimens. Antimicrob. Agents Chemother. 2012, 56, 2108–2113. [Google Scholar] [CrossRef] [PubMed]
- Ultee, A.; Kets, E.P.W.; Smid, E.J. Mechanisms of action of carvacrol on the food-borne pathogen Bacillus cereus. Appl. Environ. Microbiol. 1999, 65, 4606–4610. [Google Scholar] [PubMed]
- Miladinović, D.L.; Ilić, B.S.; Mihajilov-Krstev, T.M.; Jović, J.L.; Marković, MS. In vitro antibacterial activity of Libanotis montana essential oil in combination with conventional antibiotics. Nat. Prod. Commun. 2014, 9, 281–286. [Google Scholar] [PubMed]
- Sikkema, J.; de Bont, J.A.; Poolman, B. Mechanisms of membrane toxicity of hydrocarbons. Microbiol. Rev. 1995, 59, 201–222. [Google Scholar] [PubMed]
- Yap, P.S.X.; Yiap, B.C.; Ping, H.C.; Lim, S.H.E. Essential oils, a new horizon in combating bacterial antibiotic resistance. Open Microbiol. J. 2014, 8, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Moore, N.M.; Flaws, M.L. Antimicrobial resistance mechanisms in Pseudomonas aeruginosa. Clin. Lab. Sci. 2011, 24, 47–51. [Google Scholar] [PubMed]
- Fujitani, S.; Sun, H.Y.; Yu, V.L.; Weingarten, J.A. Pneumonia due to Pseudomonas aeruginosa: Part I: epidemiology, clinical diagnosis and source. Chest 2011, 139, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Mesaros, N.; Nordmann, P.; Plésiat, P.; Roussel-Delvallez, M.; van Eldere, J.; Glupczynski, Y.; van Laethem, Y.; Jacobs, F.; Lebecque, P.; Malfroot, A.; et al. Pseudomonas aeruginosa: Resistance and therapeutic options at the turn of the new millennium. Clin. Microbiol. Infect. 2007, 13, 560–578. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.; Maciá, M.D.; Gutiérrez, O.; Vidal, C.; Pérez, J.L.; Oliver, A. Molecular mechanisms of β-lactam resistance mediated by AmpC hyperproduction in Pseudomonas aeruginosa clinical strains. Antimicrob. Agents Chemother. 2005, 49, 4733–4738. [Google Scholar] [CrossRef] [PubMed]
- Ben Haj Khalifa, A.; Moissenet, D.; vu Thien, H.; Khedher, M. Virulence factors in Pseudomonas aeruginosa: Mechanisms and modes of regulation. Ann. Biol. Clin. 2011, 69, 393–403. [Google Scholar]
- Lister, P.D.; Wolter, D.J.; Hanson, N.D. Antibacterial-resistant Pseudomonas aeruginosa: Clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin. Microbiol. Rev. 2009, 22, 582–610. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.V.; da Sousa, P.J.; Pessôa, L.F.H.; de Freitas, F.R.A.; Coutinho, D.M.H.; Alves, B.N.L.; Lima, O.E. Ocimum basilicum: Antibacterial activity and association study with antibiotics against bacteria of clinical importance. Pharm. Biol. 2015, 54, 863–867. [Google Scholar] [CrossRef] [PubMed]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How antibiotics kill bacteria: From targets to networks. Nat. Rev. Microbiol. 2010, 8, 423–435. [Google Scholar] [CrossRef] [PubMed]
- El-Hosseiny, L.; el-Shenawy, M.; Haroun, M.; Abdullah, F. Comparative evaluation of the inhibition effect of some essential oils with antibiotics against Pseudomonas aeruginosa. Int. J. Antibiot. 2014. [Google Scholar] [CrossRef]
- Veras, H.N.H.; Rodrigues, F.G.F.; Botelho, M.A.; Menezes, I.R.A.; Coutinho, H.D.M.; Costa, J.G.M. Enhancement of aminoglycosides and β-lactams antibiotic activity by essential oil of Lippia sidoides Cham. and the Thymol. Arab. J. Chem. 2013. [Google Scholar] [CrossRef]
- Faleiro, M.L. The mode of antibacterial action of essential oils. In Science against Microbial Pathogens: Communicating Current Research and Technological Advances; Méndez Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2011; pp. 1143–1156. [Google Scholar]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Griffin, S.G.; Wyllie, S.G.; Markham, J.L.; Leach, D.N. The role of structure and molecular properties of terpenoids in determining their antimicrobial activity. Flavour Fragr. J. 1999, 14, 322–332. [Google Scholar] [CrossRef]
- Rodrigues, F.F.; Costa, J.G.; Coutinho, H.D. Enhancement of the antibiotic activity of gentamicin by volatile compounds of Zanthoxylum articulatum. Indian J. Med. Res. 2010, 131, 833–835. [Google Scholar]
- Bitu, V.C.; Fecundo, H.D.; da Costa, J.G.; Coutinho, H.D.; Rodrigues, F.F.; de Santana, N.M.; Botelho, M.A.; Menezes, I.R. Chemical composition of the essential oil of Lippia gracilis schauer leaves and its potential as modulator of bacterial resistance. Nat. Prod. Res. 2014, 28, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Santos, N.K.A.; Rodrigues, F.F.G.; Coutinho, H.D.M.; Viana, G.S.B.; Costa, J.G.M. Isolation of alpha-bisabolol from the essential oil of Vanillosmopsis arborea baker and modulation of antibiotic activity using gaseous contact. J. Essent. Oil Bear. Plants 2013, 16, 826–831. [Google Scholar] [CrossRef]
- Rodrigues, F.F.; Costa, J.G.; Coutinho, H.D. Synergy effects of the antibiotics gentamicin and the essential oil of Croton zehntneri. Phytomedicine 2009, 16, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Hilmer, S.N.; Tran, K.; Rubie, P.; Wright, J.; Gnjidic, D.; Mitchell, S.J.; Matthews, S. Gentamicin pharmacokinetics in old age and frailty. Br. J. Clin. Pharmacol. 2011, 71, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Brehm-Stecher, B.F.; Johnson, E.A. Sensitization of Staphylococcus aureus and Escherichia coli to antibiotics by the sesquiterpenoids nerolidol, farnesol, bisabolol, and apritone. Antimicrob. Agents Chemother. 2003, 47, 3357–3360. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, H.D.M.; Rodrigues, F.F.G.; Nascimento, E.M.M.; Costa, J.M.C.; Falcão-Silva, V.S.; Siqueira-Júnior, J.P. Synergism of gentamicin and norfloxacin with the volatile compounds of Lippia microphylla cham. (Verbenaceae). J. Essent. Oil Res. 2011, 23, 24–28. [Google Scholar]
- De Sousa, E.O.; Rodrigues, F.F.; Campos, A.R.; Lima, S.G.; da Costa, J.G. Chemical composition and synergistic interaction between aminoglycosides antibiotics and essential oil of Lantana montevidensis Briq. Nat. Prod. Res. 2013, 27, 942–945. [Google Scholar] [CrossRef] [PubMed]
- Dahham, S.S.; Tabana, Y.M.; Iqbal, M.A.; Ahamed, M.B.; Ezzat, M.O.; Majid, A.S.; Majid, A.M. The anticancer, antioxidant and antimicrobial properties of the sesquiterpene β-caryophyllene from the essential oil of Aquilaria crassna. Molecules 2015, 20, 11808–11829. [Google Scholar] [CrossRef] [PubMed]
- Hemaiswarya, S.; Doble, M. Synergistic interaction of eugenol with antibiotics against Gram negative bacteria. Phytomedicine 2009, 16, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Thiolas, A.; Bollet, C.; la Scola, B.; Raoult, D.; Pagès, J.M. Successive emergence of Enterobacter aerogenes strains resistant to imipenem and colistin in a patient. Antimicrob. Agents Chemother. 2005, 49, 1354–1358. [Google Scholar] [CrossRef] [PubMed]
- Davin-Regli, A.; Pagès, J.M. Enterobacter aerogenes and Enterobacter cloacae; versatile bacterial pathogens confronting antibiotic treatment. Front. Microbiol. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavigne, J.P.; Sotto, A.; Nicolas-Chanoine, M.H.; Bouziges, N.; Bourg, G.; Davin-Regli, A.; Pagès, J.M. Membrane permeability, a pivotal function involved in antibiotic resistance and virulence in Enterobacter aerogenes clinical isolates. Clin. Microbiol. Infect. 2012, 18, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Fadli, M.; Chevalier, J.; Bolla, J.M.; Mezrioui, N.E.; Hassani, L.; Pagès, J.M. Thymus maroccanus essential oil, a membranotropic compound active on Gram-negative bacteria and resistant isolates. J. Appl. Microbiol. 2012, 113, 1120–1129. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.A.; Morey, P.; Bengoechea, J.A. Quinolones sensitize Gram-negative bacteria to antimicrobial peptides. Antimicrob. Agents Chemother. 2006, 50, 2361–2367. [Google Scholar] [CrossRef] [PubMed]
- Mobley, H.L.; Chippendale, G.R.; Swihart, K.G.; Welch, R.A. Cytotoxicity of the HpmA hemolysin and urease of Proteus mirabilis and Proteus vulgaris against cultured human renal proximal tubular epithelial cells. Infect. Immun. 1991, 59, 2036–2042. [Google Scholar] [PubMed]
- Datz, M.; Joris, B.; Azab, E.A.; Galleni, M.; van Beeumen, J.; Frère, J.M.; Martin, H.H. A common system controls the induction of very different genes. The class-A beta-lactamase of Proteus vulgaris and the Enterobacterial class-C beta-lactamase. Eur. J. Biochem. 1994, 226, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Thenmozhi, S.; Moorthy, K.; Sureshkumar, B.T.; Suresh, M. Antibiotic resistance mechanism of ESBL producing Enterobacteriaceae in clinical field: A review. Int. J. Pure App. Biosci. 2014, 2, 207–226. [Google Scholar]
- Choi, J.G.; Kang, O.H.; Lee, Y.S.; Oh, Y.C.; Chae, H.S.; Jang, H.J.; Shin, D.W.; Kwon, D.Y. Antibacterial activity of methyl gallate isolated from Galla Rhois or carvacrol combined with nalidixic acid against nalidixic acid resistant bacteria. Molecules 2009, 14, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Eng, S.K.; Pusparajaha, P.; Ab Mutalibc, N.S.; Hooi-Leng Sera, H.L.; Chand, K.G.; Leea, L.H. Salmonella: A review on pathogenesis, epidemiology and antibiotic resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef]
- Butler, T.; Sridhar, C.B.; Daga, M.K.; Pathak, K.; Pandit, R.B.; Khakhria, R.; Potkar, C.N.; Zelasky, M.T.; Johnson, R.B. Treatment of typhoid fever with azithromycin versus chloramphenicol in a randomized multicentre trial in India. J. Antimicrob. Chemother. 1999, 44, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, M.J.; Borges, A.; Dias, C.; Aires, A.; Bennett, R.N.; Rosa, E.S.; Simões, M. Antimicrobial activity of phenolics and glucosinolate hydrolysis products and their synergy with streptomycin against pathogenic bacteria. Med. Chem. 2010, 6, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.M.; Preston, J.F.; Wei, C.I. Antibacterial mechanism of allyl isothiocyanate. J. Food Prot. 2000, 63, 727–734. [Google Scholar] [PubMed]
- Bosnic, T.; Softic, D.; Grujic-Vasic, J. Antimicrobial activity of some essential oils and major constituents of essential oils. Acta Med. Acad. 2006, 35, 19–22. [Google Scholar]
- El Bouzidi, L.; Jamali, C.A.; Bekkouche, K.; Hassani, L.; Wohlmuth, H.; Leach, D.; Abbad, A. Chemical composition, antioxidant and antimicrobial activities of essential oils obtained from wild and cultivated Moroccan Thymus species. Ind. Crop. Prod. 2013, 43, 450–456. [Google Scholar] [CrossRef]
- Nostro, A.; Sudano Roccaro, A.; Bisignano, G.; Marino, A.; Cannatelli, M.A.; Pizzimenti, F.C.; Cioni, P.L.; Procopio, F.; Blanco, A.R. Effects of oregano, carvacrol and thymol on Staphylococcus aureus and Staphylococcus epidermidis biofilms. J. Med. Microbiol. 2007, 56, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Mechergui, K.; Jaouadi, W.; Coelho, J.P.; Khouja, M.L. Effect of harvest year on production, chemical composition and antioxidant activities of essential oil of oregano (Origanum vulgare subsp. glandulosum (Desf.) Ietswaart) growing in North Africa. Ind. Crop. Prod. 2016, 90, 32–37. [Google Scholar] [CrossRef]
- Asensio, C.M.; Grosso, N.R.; Juliani, H.R. Quality characters, chemical composition and biological activities of oregano (Origanum spp.) essential oils from central and Southern Argentina. Ind. Crop. Prod. 2015, 63, 203–213. [Google Scholar] [CrossRef]
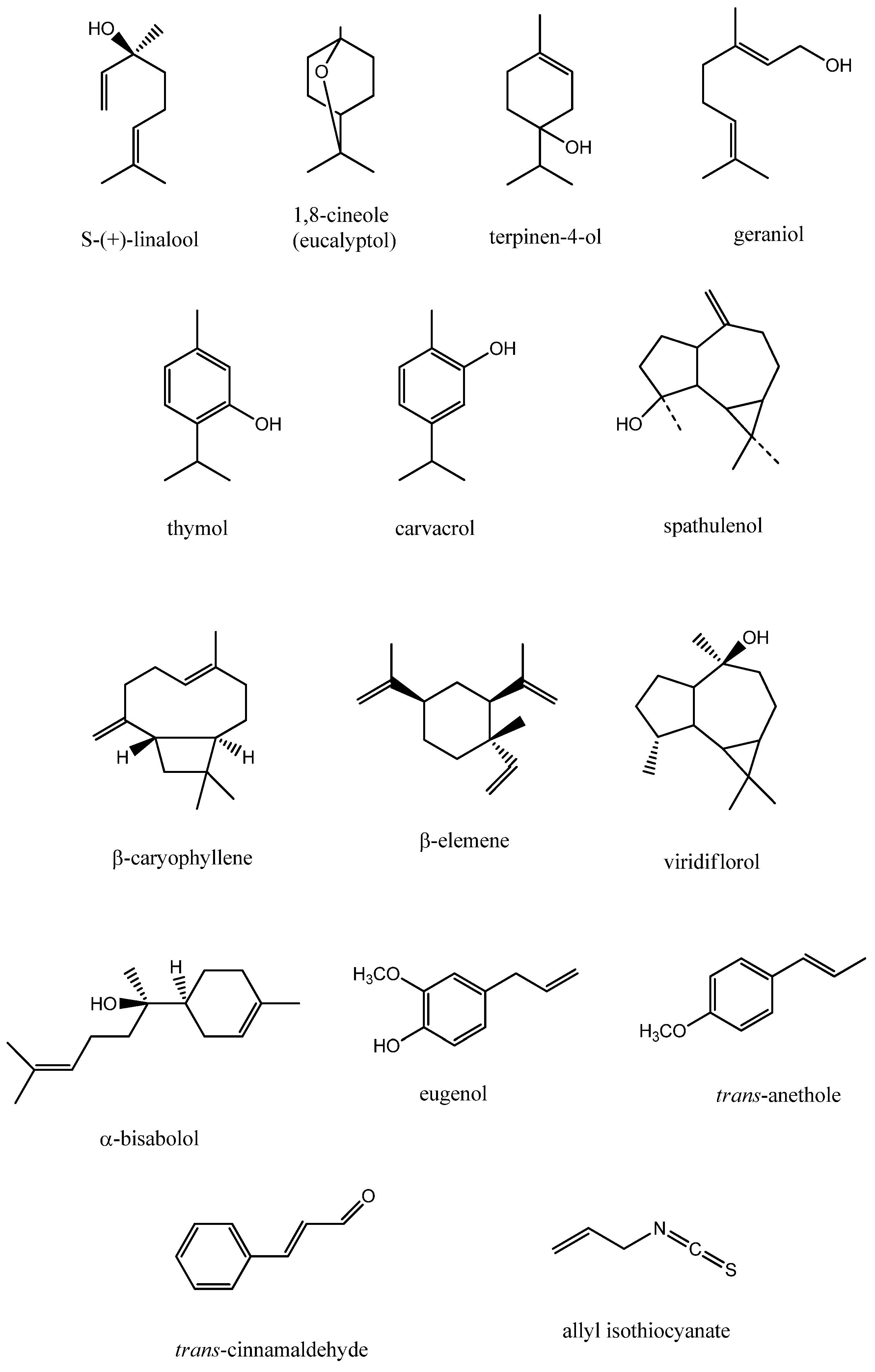

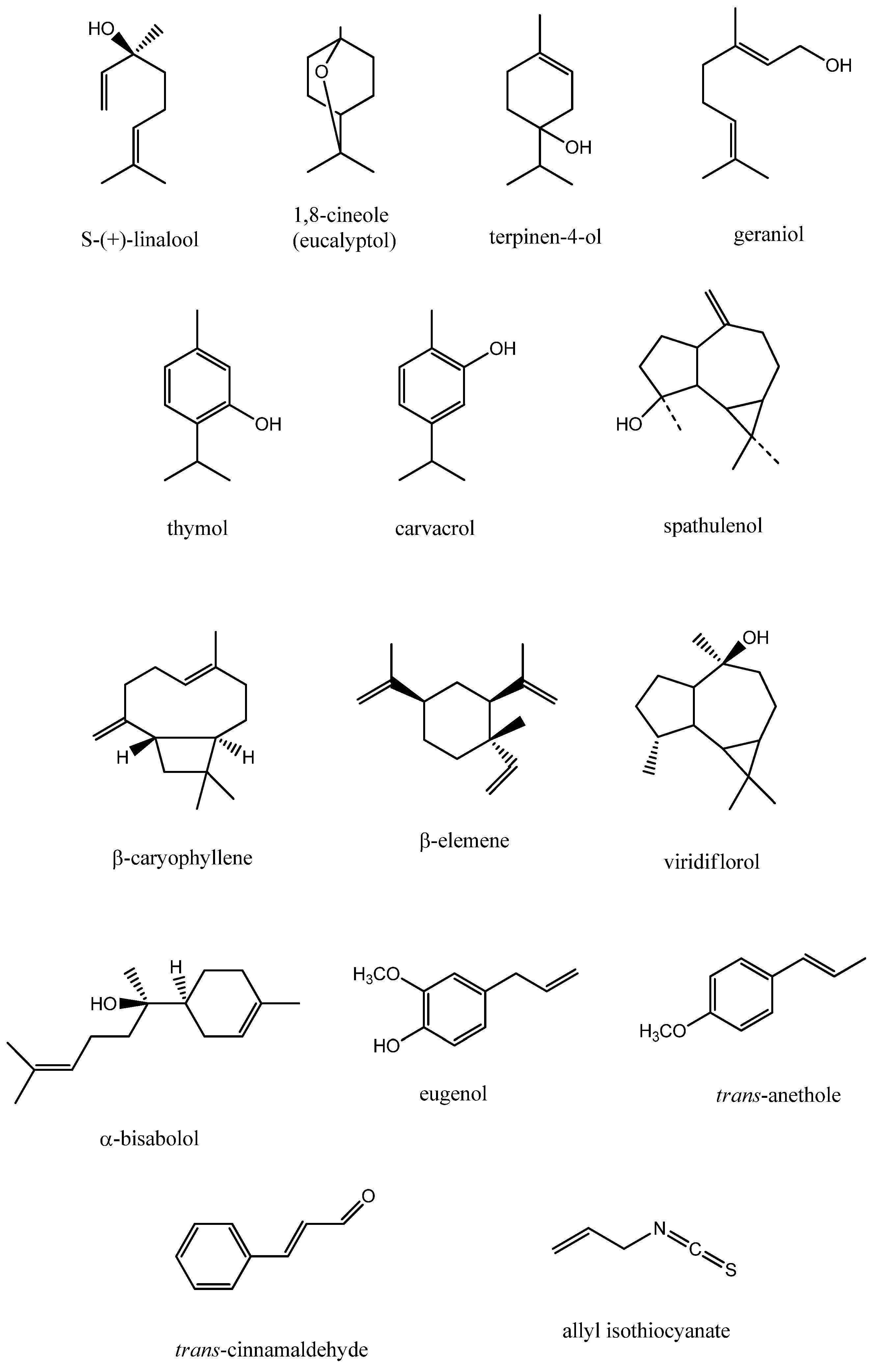
{kind=link}
{kind=link}
| Essential Oil/Volatile Compound | Vegetal Source | Major Constituent | Acinetobacter baumannii Strain | Antibiotic | Method | Effect | Ref. |
|---|---|---|---|---|---|---|---|
| Tea tree EO | Melaleucae folium (Melaleuca alternifolia (Maiden. and Beach.) Cheel., Myrtaceae, Tea tree) | Terpinen-4-ol (30.3%) | ATCC 19606 | Gentamicin | CB (FICI = 0.5) | S | [34] |
| Myrtle EO | Myrti folium (Myrtus communis L., Myrtaceae, Myrtle) | Linalool (22.276%) Myrtenyl acetate (16.561%) 1,8-Cineole (13.250%) | ATCC 19606 ATCC BAA747 and 22 clinical isolates of which MDR strains: Aba-6637, Aba-4914, Aba-5055 | Ciprofloxacin | CB (FICI < 0.5) TK | S | [24] |
| Linalool (18.320%) Myrtenyl acetate (18.009%) 1,8-Cineole (16.878%) | |||||||
| Polymyxin B | S | ||||||
| Linalool (26.591%) Myrtenyl acetate (18.489%) 1,8-Cineole (15.762%) | |||||||
| Longbeak eucalyptus EO | Eucalypti folium (Eucalyptus camaldulensis Dehnh., Myrtaceae, Longbeak eucalyptus) | Spathulenol (18.90%) | Aba-4914 Aba-5055 | Ciprofloxacin | CB (FICI < 0.5) | S | [25] |
| Aba-6673 | CB (FICI = 0.53) | Ad | |||||
| Aba-4914 Aba-6673 | Gentamicin | CB (FICI < 0.5) | S | ||||
| Spathulenol (21.39%) | |||||||
| CB (FICI = 0.5) | Ad | ||||||
| ATCC 19606 Aba-4914 Aba-5055 | Polymyxin B | CB (FICI < 0.5) TK | S | ||||
| Rosewood EO | Anibae lignum (Aniba rosaeodora Ducke, Lauraceae, Rosewood) | Linalool (60.1%) | ATCC 19606 | Gentamicin | CB (FICI = 0.11) | S | [34] |
| Cinnamon EO | Cinnamomi cortex (Cinnamomum zeylanicum Nees, Lauraceae, Cinnamon) | - | Clinical isolate A-06 | Amikacin | CB (FICI = 0.045) | S | [40] |
| Gentamicin | CB (FICI = 0.5) | Ad | |||||
| Imipenem | CB (FICI = 2) | I | |||||
| Meropenem | CB (FICI = 1.5) | I | |||||
| Coriander EO | Coriandri fructus (Coriandrum sativum L., Apiaceae, Coriander) | - | LMG 1025 | Cefoperazone | CB (FICI = 0.750) | Ad | [30] |
| Chloramphenicol | CB (FICI = 0.312) | S | |||||
| Ciprofloxacin | CB (FICI = 0.281) | S | |||||
| Gentamicin | CB (FICI = 0.250) | S | |||||
| Piperacillin | CB (FICI = 1) | Ad | |||||
| Tetracycline | CB (FICI = 0.312) | S | |||||
| LMG 1041 | Cefoperazone | CB (FICI = 1) | Ad | ||||
| Chloramphenicol | CB (FICI = 0.047) | S | |||||
| Ciprofloxacin | CB (FICI = 0.375) | S | |||||
| Gentamicin | CB (FICI = 0.375) | S | |||||
| Piperacillin | CB (FICI = 0.625) | Ad | |||||
| Tetracycline | CB (FICI = 0.185) | S | |||||
| Immortelle EO | Helichrysi flos (Helichrysum italicum (Roth) G. Don, Asteraceae, Immortelle) | - | ATCC 19606 Clinical isolate AB1 | Chloramphenicol | FR (8-fold reduction in MIC) | S | [37] |
| Oregano EO | Origani herba (Origanum vulgare Linn., Lamiaceae, Oregano) | Cymenol (58.6%) | ATCC 19606 | Gentamicin | CB (FICI = 0.65) | Ad | [34] |
| Geranium EO | Pelargonii herba (Pelargonium graveolens L., Geraniaceae, Geranium) | Citronellol (47.3%) | ATCC 19606 | Gentamicin | CB (FICI = 0.11) | S | |
| Lemon EO | Citri pericarpium (Citrus limon L. Burm. F., Rutaceae, Lemon) | - | Clinical isolate A-06 | Amikacin | CB (FICI = 0.037) | S | [40] |
| Gentamicin | CB (FICI = 0.5) | Ad | |||||
| Imipenem | CB (FICI = 2) | I | |||||
| Meropenem | CB (FICI = 2) | I |
| Essential Oil/Volatile Compound | Vegetal Source | Major Constituent | Escherichia coli Strain | Antibiotic | Method | Effect | Ref. |
|---|---|---|---|---|---|---|---|
| Oregano EO | Origani herba (Origanum vulgare Linn., Lamiaceae, Oregano) | Cymenol (58.6%) | ATCC 25922 | Gentamicin | CB (FICI = 0.65) | Ad | [34] |
| Thyme EO | Thymi herba (Thymus maroccanus L., Lamiaceae, Za’ater/Azukeni) | Carvacrol (76.35%) | AG100 | Chloramphenicol | FR (4-fold reduction in MIC) | S | [52] |
| AG102 | FR (32-fold reduction in MIC) | ||||||
| Thymi herba (Thymus broussonetii L., Lamiaceae, Za’atar Essaouiri) | Carvacrol (39.77%) Borneol (12.03%) | AG100 | FR (4-fold reduction in MIC) | ||||
| AG102 | FR (32-fold reduction in MIC) | ||||||
| Lemon thyme EO | Thymi herba (Thymus pulegioides L., Lamiaceae, Lemon thyme/broad-leaved thyme) | Geraniol (66.59%) | ATCC 25922 | Tetracycline | CB (FICI = 0.43–0.76) | S | [74] |
| CB (FICI = 0.98–1.09) | Ad | ||||||
| Streptomycin | CB (FICI = 1.20–2.80) | A | |||||
| Chloramphenicol | CB (FICI = 0.21–0.87) | S | |||||
| CB (FICI = 0.98–1.09) | Ad | ||||||
| Savory EO | Saturejae herba (Satureja kitaibelii Wierzb. ex Heuff., Lamiaceae, Savory) | Geraniol (50.4%) | ATCC 25922 | Chloramphenicol | CB (FICI = 0.21–0.87) | S | [60] |
| CB (FICI = 0.98–1.09) | Ad | ||||||
| Tetracycline | CB (FICI = 0.32–0.87) | S | |||||
| CB (FICI = 0.98–1.09) | Ad | ||||||
| Sandarac EO | Tetraclinis coni (Tetraclinis articulata (Vahl.) Masters., Cupressaceae, Sandarac) | α-Campholenal (16.34%) Trans-Pinocarveol (15.45%) Verbenone (13.36%) Cis-Verbenol (12.36%) | ATCC 10536 | Amoxicillin | CB (FICI = 1) | Ad | [75] |
| Rosemary EO | Rosmarini folium (Rosmarinus officinalis L., Lamiaceae, Rosemary) | 1,8-Cineole (30.87%) | Clinical isolate | Amikacin | FR (~250-fold reduction in MIC) | S | [61] |
| Neomycin | FR (~250-fold reduction in MIC) | ||||||
| Gentamicin | FR (~4200-fold reduction in MIC) | ||||||
| Cidreira-do-mato/cidreira-brava EO | Hyptis folium (Hyptis martiusii Benth., Lamiaceae, Cidreira-do-mato/cidreira-brava) | Bicyclogermacrene (10.6%) Trans-Caryophyllene (9.2%) Caryophyllene oxide (7.4%) 1,8-Cineole (7%) | ATCC 25922 | Amikacin | FR (–) | I | [64] |
| IZGC (+2.4%) | S | ||||||
| Gentamycin | FR (–) | I | |||||
| IZGC (+17.6%) | S | ||||||
| Neomycin | FR (4-fold reduction in MIC) | S | |||||
| Kanamycin | FR (–) | I | |||||
| Tobramycin | IZGC (+76.5%) | S | |||||
| Cidreira-do-mato/cidreira-brava EO | Hyptis folium (Hyptis martiusii Benth., Lamiaceae, Cidreira-do-mato/cidreira-brava) | Bicyclogermacrene (10.6%) Trans-Caryophyllene (9.2%) Caryophyllene oxide (7.4%) 1,8-Cineole (7%) | Clinical isolate | Amikacin | FR (–) | I | [64] |
| IZGC (+82.4%) | S | ||||||
| Gentamycin | FR (–) | I | |||||
| IZGC (+40.0%) | S | ||||||
| Neomycin | FR (–) | I | |||||
| Kanamycin | FR (–) | I | |||||
| Tobramycin | IZGC (+80.0%) | S | |||||
| Thyme EO | Thymi herba (Thymus riatarum Humbert and Maire., Lamiaceae, Moroccan thyme) | Borneol (41.67%) | AG100 | Chloramphenicol | FR (4-fold reduction in MIC) | S | [58] |
| AG100A | FR (2-fold reduction in MIC) | ||||||
| Thyme EO | Thymi herba (Thymus saturejoides Coss., Lamiaceae, Azoukni) | Carvacrol (25.3%) Borneol (19.7%) | ATCC 25922 | Cefixime | CB (FICI = 1.25) | I | [76] |
| Carvacrol (26.5%) Borneol (20.1%) | |||||||
| Carvacrol (45.3%) Borneol (7.5%) | |||||||
| Rosewood EO | Anibae lignum (Aniba rosaeodora Ducke, Lauraceae, Rosewood) | Linalool (60.1%) | ATCC 25922 | Gentamicin | CB (FICI = 0.35) | S | [34] |
| Geranium EO | Pelargonii herba (Pelargonium graveolens L’Hér., Geraniaceae, Geranium) | Citronellol (47.3%) | ATCC 25922 | Gentamicin | CB (FICI = 0.30) | S | [34] |
| Tea tree EO | Melaleucae folium (Melaleuca alternifolia (Maiden. and Beach.) Cheel., Myrtaceae, Tea tree) | Terpinen-4-ol (30.3%) | ATCC 25922 | Gentamicin | CB (FICI = 0.49) | S | |
| Citronellol | - | Clinical isolate | Penicillin | CB (FICI = 1.03) | I | [65] | |
| Geraniol | - | Clinical isolate | Penicillin | CB (FICI = 1.5) | I | [60] | |
| ATCC 25922 | Chloramphenicol | CB (FICI = 0.32–0.87) | S | ||||
| CB (FICI = 0.98–1.09) | Ad | ||||||
| Tetracycline | CB (FICI = 0.32–0.87) | S | |||||
| CB (FICI = 0.98–1.28) | Ad | ||||||
| Menthol | - | Clinical isolate | Penicillin | CB (FICI = 1.5) | I | [65] | |
| Myrcene | - | Clinical isolate | Penicillin | CB (FICI = 10) | A | ||
| Thymol | - | Clinical isolate | Penicillin | CB (FICI = 0.15) | S | [65] | |
| N00 666 | Ampicillin | CB (FICI = 0.12) | S | [66] | |||
| Penicillin | CB (FICI = 0.20) | ||||||
| Tetracycline | CB (FICI = 0.15) | ||||||
| Erythromycin | CB (FICI = 0.25) | ||||||
| Bacitracin | CB (FICI = 0.56) | I | |||||
| Novobiocin | CB (FICI = 0.37) | S | |||||
| Eugenol | - | Clinical isolate | Penicillin | CB (FICI = 0.16) | S | [65] | |
| N00 666 | Ampicillin | CB (FICI > 0.5) | I | [66] | |||
| Penicillin | |||||||
| Tetracycline | CB (FICI = 0.16) | S | |||||
| Erythromycin | CB (FICI = 1.1) | I | |||||
| Bacitracin | CB (FICI = 0.5) | S | |||||
| Novobiocin | CB (FICI = 1.1) | I | |||||
| Carvacrol | - | Clinical isolate | Penicillin | CB (FICI = 2) | I | [65] | |
| N00 666 | Ampicillin | CB (FICI = 0.25) | S | [66] | |||
| Penicillin | CB (FICI = 0.37) | ||||||
| Tetracycline | CB (FICI = 0.15) | ||||||
| Erythromycin | CB (FICI = 1.0) | I | |||||
| Bacitracin | CB (FICI = 0.25) | S | |||||
| Novobiocin | CB (FICI = 0.63) | I | |||||
| Cinnamaldehyde | - | N00 666 | Ampicillin | CB (FICI = 0.37) | S | [66] | |
| Penicillin | CB (FICI = 0.24) | ||||||
| Tetracycline | CB (FICI = 0.37) | ||||||
| Erythromycin | CB (FICI = 0.24) | ||||||
| Bacitracin | CB (FICI = 0.63) | I | |||||
| Novobiocin | CB (FICI = 0.24) | S | |||||
| Allyl isothiocyanate | - | N00 666 | Ampicillin | CB (FICI = 0.63) | I | ||
| Penicillin | CB (FICI = 1.0) | ||||||
| Tetracycline | CB (FICI = 0.75) | ||||||
| Erythromycin | CB (FICI = 0.73) | ||||||
| Bacitracin | CB (FICI = 0.5) | S | |||||
| Novobiocin | CB (FICI = 1.0) | I | |||||
| Essential Oil/Volatile Compound | Vegetal Source | Major Constituent | Klebsiella pneumoniae Strain | Antibiotic | Method | Effect | Ref. |
|---|---|---|---|---|---|---|---|
| Peppermint EO | Menthae folium (Mentha piperita L., Lamiaceae, Peppermint) | - | NCTC 9633 | Ciprofloxacin | CB (FICI = 0.68–0.90) | S | [2] |
| CB (FICI = 1.40–2.24) | A | ||||||
| Rosemary EO | Rosmarini folium (Rosmarinus officinalis L., Lamiaceae, Rosemary) | - | NCTC 9633 | Ciprofloxacin | CB (FICI = 0.28–0.97) | S | |
| CB (FICI = 1.03–1.07) | A | ||||||
| Thyme EO | Thymi herba (Thymus vulgaris L., Lamiaceae, Thyme) | - | NCTC 9633 | Ciprofloxacin | CB (FICI = 0.71–0.90) | S | |
| CB (FICI = 1.10–1.40) | A | ||||||
| Thyme EO | Thymi herba (Thymus maroccanus L., Lamiaceae, Za’ater/Azukeni) | Carvacrol (76.35%) | Clinical isolate | Ciprofloxacin | CB (FICI = 0.37) | S | [57] |
| Gentamicin | CB (FICI = 0.50) | S | |||||
| Pristinamycin | CB (FICI = 0.50) | S | |||||
| Cefixime | CB (FICI = 1) | I | |||||
| Thyme EO | Thymi herba (Thymus broussonetii L., Lamiaceae, Za’atar Essaouiri) | Carvacrol (39.77%) Borneol (12.03%) | Ciprofloxacin | CB (FICI = 0.62) | PS | ||
| Gentamicin | CB (FICI = 0.62) | PS | |||||
| Pristinamycin | CB (FICI = 0.50) | S | |||||
| Cefixime | CB (FICI = 1) | I | |||||
| Lemon thyme EO | Thymi herba (Thymus pulegioides L., Lamiaceae, Lemon thyme/broad-leaved thyme) | Geraniol (66.59%) | ATCC 700603 | Tetracycline | CB (FICI = 0.76–0.82) | S | [74] |
| CB (FICI = 0.92–1.10) | Ad | ||||||
| CB (FICI = 1.16–1.28) | A | ||||||
| Streptomycin | CB (FICI = 0.32–0.87) | S | |||||
| CB (FICI = 1.09) | Ad | ||||||
| CB (FICI = 1.16) | A | ||||||
| Chloramphenicol | CB (FICI = 0.32–0.87) | S | |||||
| CB (FICI = 0.98–1.09) | Ad | ||||||
| Savory EO | Saturejae herba (Satureja kitaibelii Wierzb. ex Heuff., Lamiaceae, Savory) | Geraniol (50.4%) | ATCC 700603 | Chloramphenicol | CB (FICI = 0.21–0.43) | S | [60] |
| CB (FICI = 0.54–0.98) | Ad | ||||||
| CB (FICI = 1.09) | I | ||||||
| Tetracycline | CB (FICI = 0.43) | S | |||||
| CB (FICI = 0.56–0.98) | Ad | ||||||
| CB (FICI = 1.09) | I | ||||||
| Thyme EO | Thymi herba (Thymus saturejoides Coss., Lamiaceae, Azoukni) | Carvacrol (25.3%) Borneol (19.7%) | Clinical isolate | Cefixime | CB (FICI = 0.75) | PS | [76] |
| Carvacrol (26.5%) Borneol (20.1%) | CB (FICI = 0.75) | PS | |||||
| Carvacrol (45.3%) Borneol (7.5%) | CB (FICI = 0.50) | S | |||||
| Sandarac EO | Tetraclinis coni (Tetraclinis articulata (Vahl.) Masters., Cupressaceae, Sandarac) | α-Campholenal (16.34%) Trans-Pinocarveol (15.45%) Verbenone (13.36%) Cis-Verbenol (12.36%) | CIP 8291 | Amoxicillin | CB (FICI = 0.8) | PS | [75] |
| Tea tree EO | Melaleucae folium (Melaleuca alternifolia, (Maiden. and Betch.) Chee., Myrtaceae, Tea tree) | - | NCTC 9633 | Ciprofloxacin | CB (FICI = 0.73–0.95) | S | [2] |
| CB (FICI = 1.03–1.85) | A | ||||||
| Geraniol | - | ATCC 700603 | Chloramphenicol | CB (FICI = 0.32–0.87) | S | [60] | |
| CB (FICI = 0.98–1.09) | Ad | ||||||
| Tetracycline | CB (FICI = 0.76–0.80) | S | |||||
| CB (FICI = 0.95–1.10) | Ad | ||||||
| CB (FICI = 1.25–1.66) | A | ||||||
| Essential Oil/Volatile Compound | Vegetal Source | Major Constituent | Pseudomonas aeruginosa Strain | Antibiotic | Method | Effect | Ref. |
|---|---|---|---|---|---|---|---|
| Thyme EO | Thymi herba (Thymus maroccanus L., Lamiaceae, Za’ater/Azukeni) | Carvacrol (76.35%) | Clinical isolate | Ciprofloxacin | CB (FICI = 0.15) | S | [57] |
| Gentamicin | CB (FICI = 0.18) | S | |||||
| Pristinamycin | CB (FICI = 0.75) | PS | |||||
| Cefixime | CB (FICI = 0.75) | PS | |||||
| Za’atar Essaouiri EO | Thymi herba (Thymus broussonetii L., Lamiaceae, Za’atar Essaouiri) | Carvacrol (39.77%) Borneol (12.03%) | Ciprofloxacin | CB (FICI = 0.14) | S | ||
| Gentamicin | CB (FICI = 0.28) | S | |||||
| Pristinamycin | CB (FICI = 0.75) | PS | |||||
| Cefixime | CB (FICI = 0.5) | S | |||||
| Lemon thyme EO | Thymi herba (Thymus pulegioides L., Lamiaceae, Lemon thyme/broad-leaved thyme) | Geraniol (66.59%) | ATCC 27853 | Tetracycline | CB (FICI = 0.54–0.82) | S | [74] |
| CB (FICI = 0.95–1.08) | Ad | ||||||
| CB (FICI = 1.16–1.28) | A | ||||||
| Streptomycin | CB (FICI = 1.20–2.00) | A | |||||
| Chloramphenicol | CB (FICI = 0.43–0.87) | S | |||||
| CB (FICI = 0.98–1.09) | Ad | ||||||
| Savory EO | Saturejae herba (Satureja kitaibelii Wierzb. ex Heuff., Lamiaceae, Savory) | Geraniol (50.4%) | ATCC 27853 | Tetracycline | CB (FICI = 0.54–0.82) | S | [60] |
| CB (FICI = 0.96–1.10) | Ad | ||||||
| CB (FICI = 1.21–1.47) | A | ||||||
| Chloramphenicol | CB (FICI = 0.43–0.87) | S | |||||
| CB (FICI = 0.98–1.09 ) | Ad | ||||||
| Thyme EO | Thymi herba (Thymus saturejoides Coss., Lamiaceae, Azoukni) | Carvacrol (25.3%) Borneol (19.7%) | ATCC 27853 | Cefixime | CB (FICI = 0.28) | S | [76] |
| Carvacrol (26.5%) Borneol (20.1%) | CB (FICI = 0.31) | ||||||
| Carvacrol (45.3%) Borneol (7.5%) | CB (FICI = 0.29) | ||||||
| Sandarac EO | Tetraclinis coni (Tetraclinis articulata (Vahl.) Masters., Cupressaceae, Sandarac) | α-Campholenal (16.34%) Trans-Pinocarveol (15.45%) Verbenone (13.36%) Cis-Verbenol (12.36%) | CIPA 22 | Amoxicillin | CB (FICI = 1.00) | Ad | [75] |
| Cidreira-do-mato/cidreira-brava EO | Hyptis folium (Hyptis martiusii Benth., Lamiaceae, Cidreira-do-mato/cidreira-brava) | Bicyclogermacrene (10.6%) Trans-Caryophyllene (9.2%) Caryophyllene oxide (7.4%) 1,8-Cineole (7%) | ATCC 15442 | Gentamycin | IZGC (+18.9%) | S | [64] |
| FR (–) | I | ||||||
| Amikacin | IZGC (+60%) | S | |||||
| FR4-fold increase in MIC | A | ||||||
| Tobramycin | IZGC (+12.5%) | S | |||||
| Neomycin | FR (–) | I | |||||
| Kanamycin | FR (–) | I | |||||
| Marjoram EO | Origani folium (Origanum majorana L., Lamiaceae, Marjoram) | 4-Terpineol (21.3%) | ATCC 9027 | Piperacillin | IZDC (+41.2%) | S | [101] |
| Cefepime | IZDC (+10%) | S | |||||
| Meropenem | IZDC (+20.5%) | S | |||||
| Gentamicin | IZDC (+31.03%) | S | |||||
| Norfloxacin | IZDC (−0.03%) | A | |||||
| Sage EO | Salviae folium (Salvia officinalis L., Lamiaceae, Sage) | 1,8-Cineole (29%) | Piperacillin | IZDC (+29.4%) | S | ||
| Cefepime | IZDC (−0.03%) | A | |||||
| Meropenem | IZDC (+23.5%) | S | |||||
| Gentamicin | IZDC (+13.7%) | S | |||||
| Norfloxacin | IZDC (−0.16%) | A | |||||
| Thyme EO | Thymi herba (Thymus vulgaris L., Lamiaceae, Thyme) | Thymol (33.6%) | Piperacillin | IZDC (+147%) | S | ||
| Cefepime | IZDC (+53.3%) | S | |||||
| Meropenem | IZDC (+52.9%) | S | |||||
| Gentamicin | IZDC (+37.9%) | S | |||||
| Basil EO | Basilici herba (Ocimum basilicum L., Lamiaceae, Basil) | Linalool (55.2%) | ATCC 25853 | Imipenem | CB (FICI = 0.75) | Ad | [99] |
| Ciprofloxacin | CB (FICI = 1.03) | I | |||||
| 1662339 | Imipenem | CB (FICI = 0.0625) | S | ||||
| Ciprofloxacin | CB (FICI = 0.09) | S | |||||
| Alecrim-de-tabuleiro EO | Lippiae folium (Lippia microphylla Cham., Verbenaceae, Alecrim-de-tabuleiro) | 1,8-Cineole (18.1%) Z-β-Ocimene (15.2%) Bicyclogermacrene (11.6%) | ATCC 15442 | Gentamicin | IZGC (+47%) | S | [112] |
| Tetracycline | IZGC (−14%) | A | |||||
| Creeping lantana EO | Lantanae folium (Lantana montevidensis Briq., Verbenaceae, Creeping lantana) | β-Caryophyllene (31.50%) Germacrene D (27.50%) | ATCC 15442 | Gentamicin | IZGC (+12%) | S | [113] |
| Amikacin | IZGC (+102%) | ||||||
| Alecrim pimento EO | Lippiae folium (Lippia sidoides Cham., Verbenaceae, Alecrim pimento) | Thymol (84.9%) | ATCC 15442 | Gentamycin | FR (4-fold reduction in MIC) | S | [102] |
| Neomycin | FR (2-fold reduction in MIC) | ||||||
| Alecrim-da-chapada EO | Lippiae folium (Lippia gracilis Schauer., Verbenaceae, Alecrim-da-chapada) | Thymol (44.4%) Carvacrol (22.2%) | ATCC 15442 | Amikacin | IZGC | S | [107] |
| Tobramycin | IZGC | ||||||
| Gentamycin | IZGC | ||||||
| Candeeiro EO | Vanillosmopsis cortex (Vanillosmopsis arborea Baker., Asteraceae Candeeiro) | α-Bisabolol (80.43%) | ATCC 15442 | Gentamycin | IZGC (+8.6%) | S | [108] |
| Tetracycline | IZGC (−8.0%) | A | |||||
| Tobramycin | IZGC (−18.0%) | A | |||||
| Immortelle EO | Helichrysi flos (Helichrysum italicum (Roth) G. Don, Asteraceae, Immortelle) | - | PAO1 PA124 | Chloramphenicol | FR (16-fold reduction in MIC) | S | [37] |
| FR (8-fold reduction in MIC) | S | ||||||
| Canela de Cunha EO | Crotonii folium (Croton zehntneri Pax et Hoffm., Euphorbiaceae Canela de Cunha) | Trans-Anethole | ATCC 15442 | Gentamycin | IZGC (+42.8%) | S | [109] |
| Tetracycline | IZGC (0%) | I | |||||
| Limão-bravo EO | Zanthoxylii folium (Zanthoxylum articulatum Engler Rutaceae, Limão-bravo) | Viridiflorol (35.4%) | ATCC 15442 | Gentamycin | IZGC (+43.8%) | S | [106] |
| Tetracycline | IZGC (+9.6%) | Ad | |||||
| Eugenol | - | NCIM 5029 | Ampicillin | CB (FICI < 0.50) | S | [115] | |
| Chloramphenicol | |||||||
| Erythromycin | |||||||
| Norfloxacin | |||||||
| Oxacillin | |||||||
| Penicillin | |||||||
| Polymyxin | |||||||
| Rifampicin | |||||||
| Erythromycin | |||||||
| Thymol | - | ATCC 15442 | Gentamycin | FR (4-fold reduction in MIC) | S | [102] | |
| Neomycin | FR (2-fold reduction in MIC) | ||||||
| Geraniol | - | ATCC 27853 | Chloramphenicol | CB (FICI = 0.54–0.87) | S | [60] | |
| CB (FICI = 0.92–1.09) | Ad | ||||||
| Tetracycline | CB (FICI = 0.76) | S | |||||
| CB (FICI = 0.92–1.10) | Ad | ||||||
| CB (FICI = 1.16–1.47) | A | ||||||
| Essential Oil/Volatile Compound | Vegetal Source | Major Constituent | Other Gram-negative Bacteria | Antibiotic | Method | Effect | Ref. |
|---|---|---|---|---|---|---|---|
| Enterobacter aerogenes Strain | |||||||
| Thyme EO | Thymi herba (Thymus maroccanus L., Lamiaceae, Za’ater/Azukeni) | Carvacrol (76.35%) | ATCC 13048 | Chloramphenicol | FR (4-fold reduction in MIC) | S | [52] |
| EA27clinical isolate | FR (8-fold reduction in MIC) | ||||||
| Thyme EO | Thymi herba (Thymus broussonetii L., Lamiaceae, Za’atar Essaouiri) | Carvacrol (39.77%) Borneol (12.03%) | ATCC 13048 | FR (4-fold reduction in MIC) | |||
| EA27clinical isolate | FR (8-fold reduction in MIC) | ||||||
| Immortelle EO | Helichrysi flos (Helichrysum italicum (Roth) G. Don, Asteraceae, Immortelle) | - | ATCC 13048 | Chloramphenicol | FR (2-fold reduction in MIC) | S | [37] |
| EAEP289 | Chloramphenicol | FR (8-fold reduction in MIC) | |||||
| Ampicillin | FR (–) | I | |||||
| Penicillin | FR (–) | ||||||
| Norfloxacin | FR (2-fold reduction in MIC) | S | |||||
| EAEP294 | Chloramphenicol | FR (128-fold reduction in MIC ) | |||||
| Ampicillin | FR (~7300-fold reduction in MIC) | ||||||
| Penicillin | FR (~14600-fold reduction in MIC) | ||||||
| Norfloxacin | FR (~914-fold reduction in MIC) | ||||||
| Eugenol | - | NCIM 5139 | Ampicillin | CB (FICI < 0.50) | S | [115] | |
| Penicillin | |||||||
| Oxacillin | |||||||
| Erythromycin | |||||||
| Norfloxacin | |||||||
| Chloramphenicol | |||||||
| Polymyxin B | |||||||
| Tetracycline | |||||||
| Vancomycin | |||||||
| Rifampin | |||||||
| Enterobacter cloacae Strain | |||||||
| Thyme EO | Thymi herba (Thymus maroccanus L. Lamiaceae, Za’ater/Azukeni) | Carvacrol (76.35%) | Clinical isolate | Ciprofloxacin | CB (FICI = 0.37) | S | [57] |
| Gentamicin | CB (FICI = 0.19) | ||||||
| Pristinamycin | CB (FICI = 0.50) | ||||||
| Cefixime | CB (FICI = 1.00) | I | |||||
| Thyme EO | Thymi herba (Thymus broussonetii L. Lamiaceae, Za’atar Essaouiri) | Carvacrol (39.77%) Borneol (12.03%) | Ciprofloxacin | CB (FICI = 0.50) | S | ||
| Gentamicin | CB (FICI = 0.50) | ||||||
| Pristinamycin | CB (FICI = 0.50) | ||||||
| Cefixime | CB (FICI = 1.00) | I | |||||
| Alecrim pimento EO | Lippiae folium (Lippia sidoides Cham. Verbenaceae, Alecrim pimento) | Thymol (84.9%) | ATCC 23355 | Gentamicin | FR (–) | I | [102] |
| Neomycin | |||||||
| Penicillin G | |||||||
| Ceftriaxone | |||||||
| Thymol | - | ATCC 23355 | Gentamicin | FR (–) | I | ||
| Neomycin | |||||||
| Penicillin G | |||||||
| Ceftriaxone | |||||||
| Proteus vulgaris Strain | |||||||
| α-Bisabolol | Candeeiro EO Candeeiro stem Vanillosmopsis arborea Baker. Asteraceae | - | ATCC 13315 | Gentamycin | IZGC (+4%) | I | [108] |
| Tetracycline | IZGC (−51.72%) | A | |||||
| Tobramycin | IZGC (−53%) | A | |||||
| Eugenol | - | NCIM 2813 | Ampicillin | CB (FICI < 0.50) | S | [115] | |
| Penicillin | |||||||
| Oxacillin | |||||||
| Erythromycin | |||||||
| Norfloxacin | |||||||
| Chloramphenicol | |||||||
| Polymyxin B | |||||||
| Tetracycline | |||||||
| Vancomycin | |||||||
| Rifampin | |||||||
| Salmonella typhimurium Strain | |||||||
| Thyme EO | Thymi herba (Thymus maroccanus L., Lamiaceae, Za’ater/Azukeni) | Carvacrol (76.35%) | Clinical isolate | Ciprofloxacin | CB (FICI = 0.37) | S | [57] |
| Gentamicin | CB (FICI = 0.75) | PS | |||||
| Pristinamycin | CB (FICI = 0.75) | ||||||
| Cefixime | CB (FICI = 0.18) | S | |||||
| Thyme EO | Thymi herba (Thymus broussonetii L., Lamiaceae, Za’atar Essaouiri) | Carvacrol (39.77%) Borneol (12.03%) | Ciprofloxacin | CB (FICI = 0.56) | PS | ||
| Gentamicin | CB (FICI = 0.62) | ||||||
| Pristinamycin | CB (FICI = 0.50) | S | |||||
| Cefixime | CB (FICI = 0.18) | ||||||
| Carvacrol | - | Clinical isolate | Nalidixic acid | CB (FICI = 0.31) | S | [124] | |
| SGI1 | Ampicillin | CB (FICI = 0.25) | [66] | ||||
| Penicillin | CB (FICI = 0.37) | ||||||
| Tetracycline | CB (FICI = 0.18) | ||||||
| Erythromycin | CB (FICI = 0.25) | ||||||
| Bacitracin | CB (FICI = 0.25) | ||||||
| Novobiocin | CB (FICI = 0.37) | ||||||
| Eugenol | - | SGI1 | Ampicillin | CB (FICI > 0.5) | I | ||
| Penicillin | CB (FICI > 0.5) | ||||||
| Tetracycline | CB (FICI = 0.22) | S | |||||
| Erythromycin | CB (FICI = 0.63) | I | |||||
| Bacitracin | CB (FICI > 0.5) | ||||||
| Novobiocin | CB (FICI = 0.40) | S | |||||
| Eugenol | - | NCIM 2501 | Ampicillin | CB (FICI < 0.50) | S | [115] | |
| Penicillin | |||||||
| Oxacillin | |||||||
| Erythromycin | |||||||
| Norfloxacin | |||||||
| Chloramphenicol | |||||||
| Polymyxin B | |||||||
| Tetracycline | |||||||
| Vancomycin | |||||||
| Rifampin | |||||||
| Thymol | - | SGI1 | Ampicillin | CB (FICI = 0.12) | S | [66] | |
| Penicillin | CB (FICI = 0.13) | ||||||
| Tetracycline | CB (FICI = 0.10) | ||||||
| Erythromycin | CB (FICI = 0.25) | ||||||
| Bacitracin | CB (FICI = 0.15) | ||||||
| Novobiocin | CB (FICI = 0.37) | ||||||
| Cinnamaldehyde | - | SGI1 | Ampicillin | CB (FICI = 0.25) | S | ||
| Penicillin | CB (FICI = 0.63) | I | |||||
| Tetracycline | CB (FICI = 0.37) | ||||||
| Erythromycin | CB (FICI = 0.24) | ||||||
| Bacitracin | CB (FICI = 0.24) | ||||||
| Novobiocin | CB (FICI = 0.24) | ||||||
| Allyl isothiocyanate | - | SGI1 | Ampicillin | CB (FICI = 0.35) | S | [66] | |
| Penicillin | CB (FICI = 0.63) | I | |||||
| Tetracycline | CB (FICI = 0.73) | ||||||
| Erythromycin | CB (FICI = 0.48) | S | |||||
| Bacitracin | CB (FICI = 0.50) | ||||||
| Novobiocin | CB (FICI = 1.00) | I | |||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aelenei, P.; Miron, A.; Trifan, A.; Bujor, A.; Gille, E.; Aprotosoaie, A.C. Essential Oils and Their Components as Modulators of Antibiotic Activity against Gram-Negative Bacteria. Medicines 2016, 3, 19. https://doi.org/10.3390/medicines3030019
Aelenei P, Miron A, Trifan A, Bujor A, Gille E, Aprotosoaie AC. Essential Oils and Their Components as Modulators of Antibiotic Activity against Gram-Negative Bacteria. Medicines. 2016; 3(3):19. https://doi.org/10.3390/medicines3030019
Chicago/Turabian StyleAelenei, Petruta, Anca Miron, Adriana Trifan, Alexandra Bujor, Elvira Gille, and Ana Clara Aprotosoaie. 2016. "Essential Oils and Their Components as Modulators of Antibiotic Activity against Gram-Negative Bacteria" Medicines 3, no. 3: 19. https://doi.org/10.3390/medicines3030019
APA StyleAelenei, P., Miron, A., Trifan, A., Bujor, A., Gille, E., & Aprotosoaie, A. C. (2016). Essential Oils and Their Components as Modulators of Antibiotic Activity against Gram-Negative Bacteria. Medicines, 3(3), 19. https://doi.org/10.3390/medicines3030019