Bisphenol S Modulates Type 1 Diabetes Development in Non-Obese Diabetic (NOD) Mice with Diet- and Sex-Related Effects

Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Husbandry
2.2. Bisphenol S (BPS) Exposure
2.3. Body Weight, Blood Glucose Measurement and Diabetic Incidence
2.4. Glucose and Insulin Tolerance Tests
2.5. Organ Collection and Flow Cytometry
2.6. Statistical Analysis
3. Results
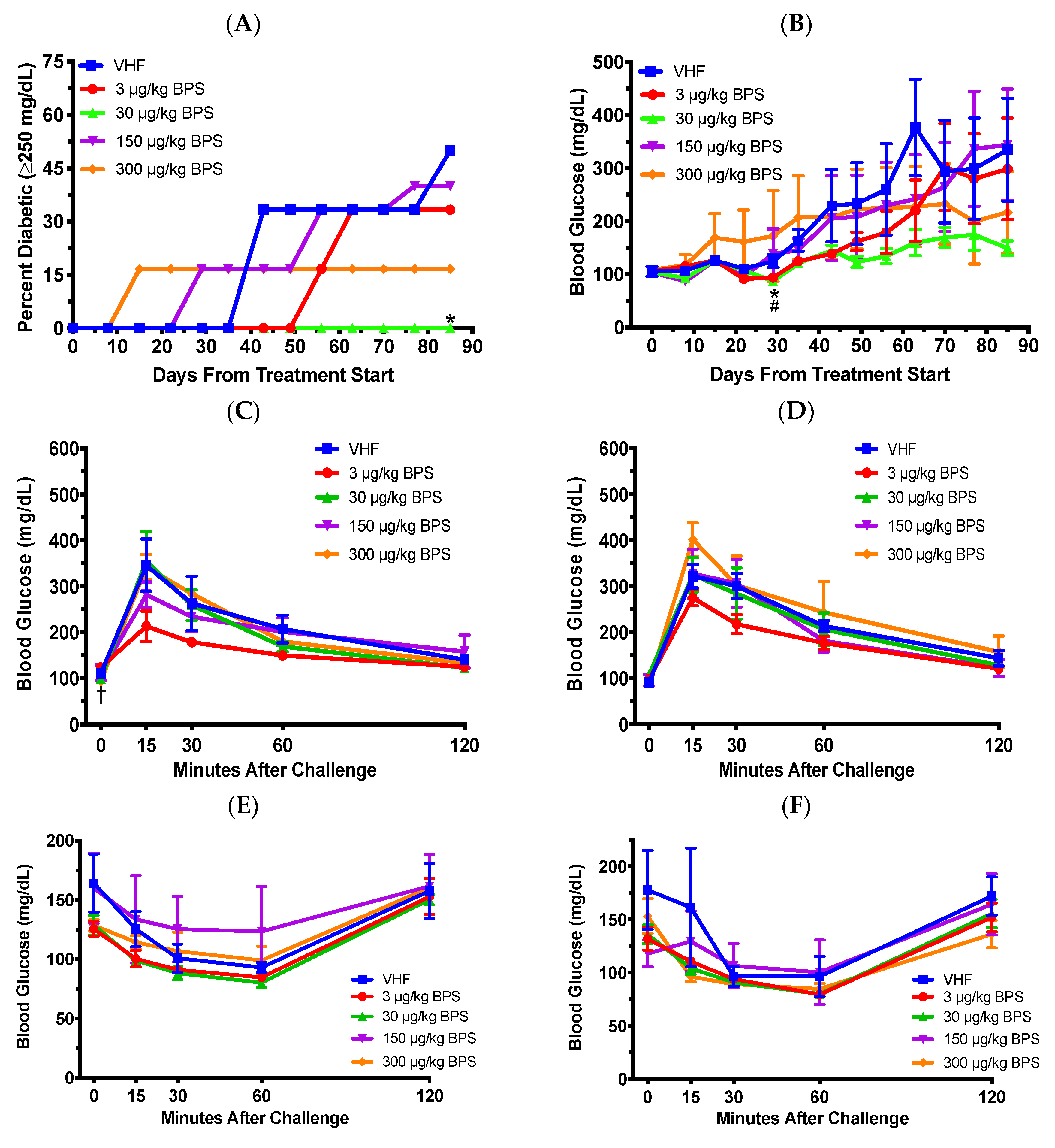
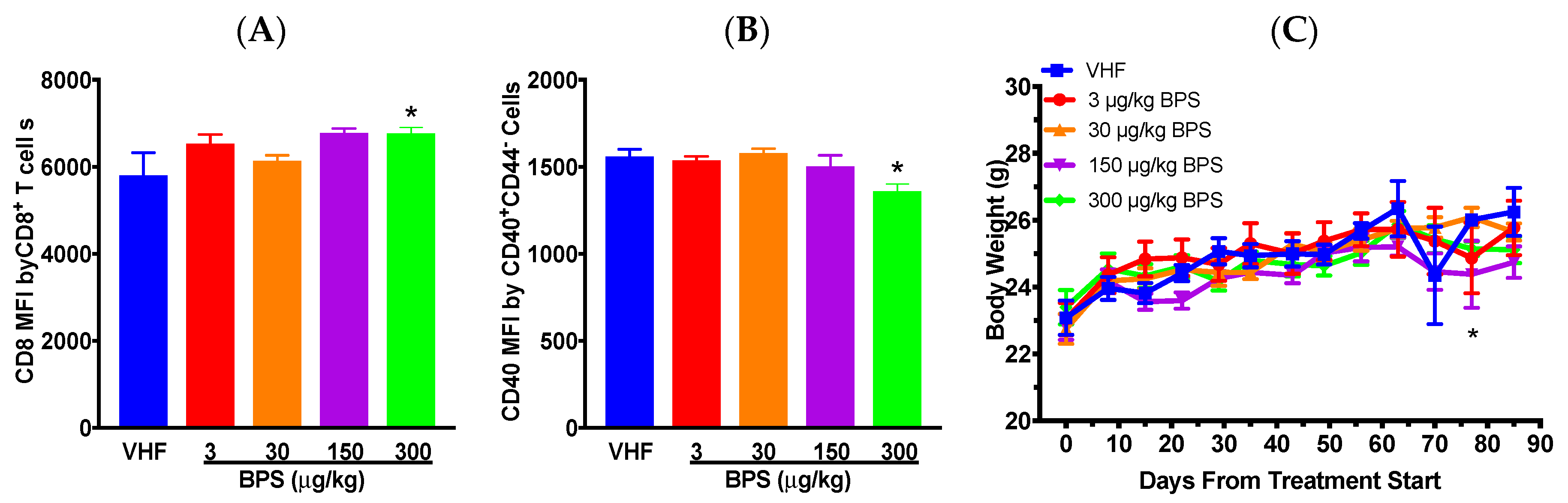
3.1. BPS Exposure Had Protective Effects on Type 1 Diabetes (T1D) In Females on A Soy-Based Diet
3.2. BPS Exposure Varied Blood Glucose Levels (BGLs) in Males on a Soy-Based Diet
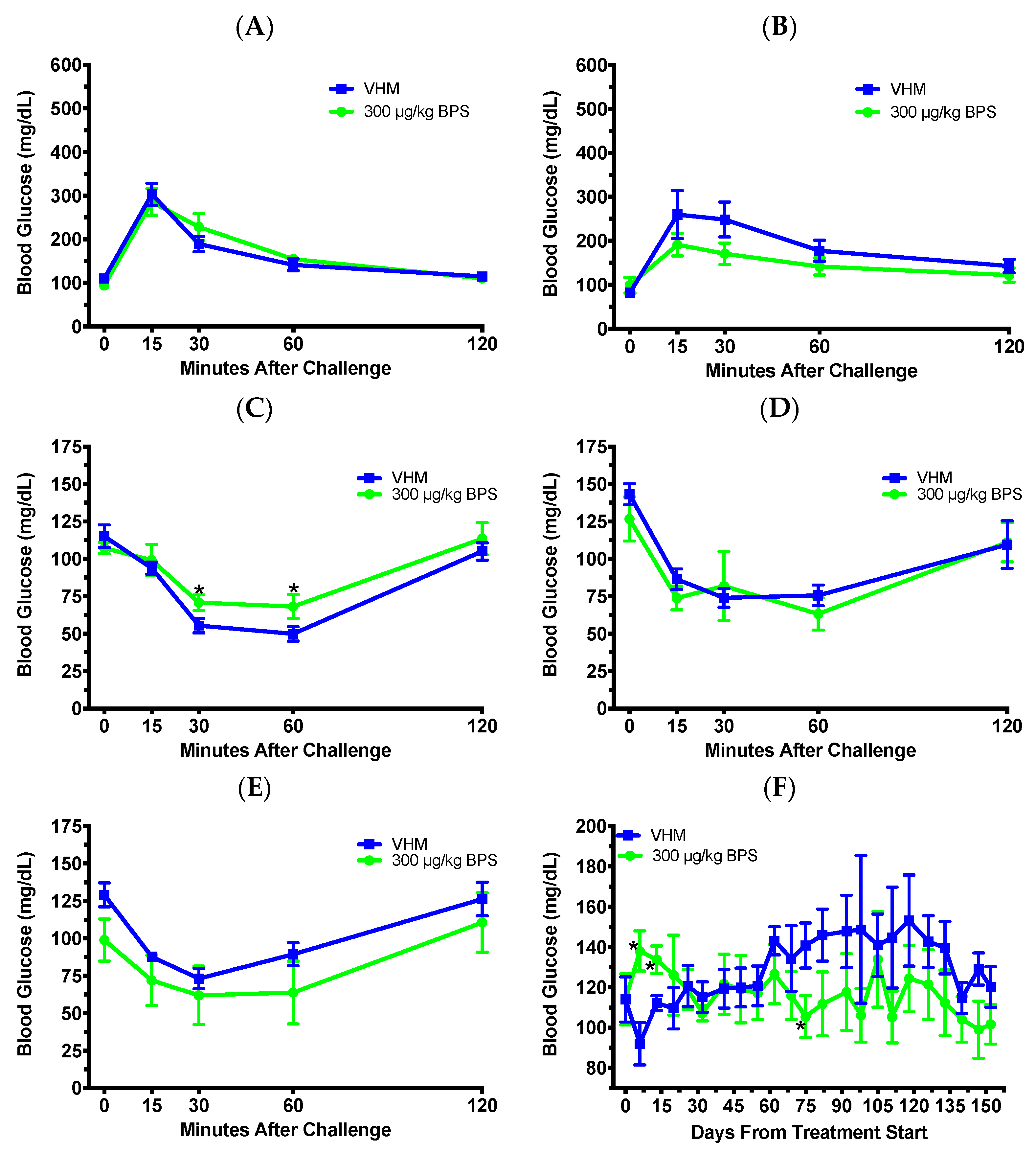
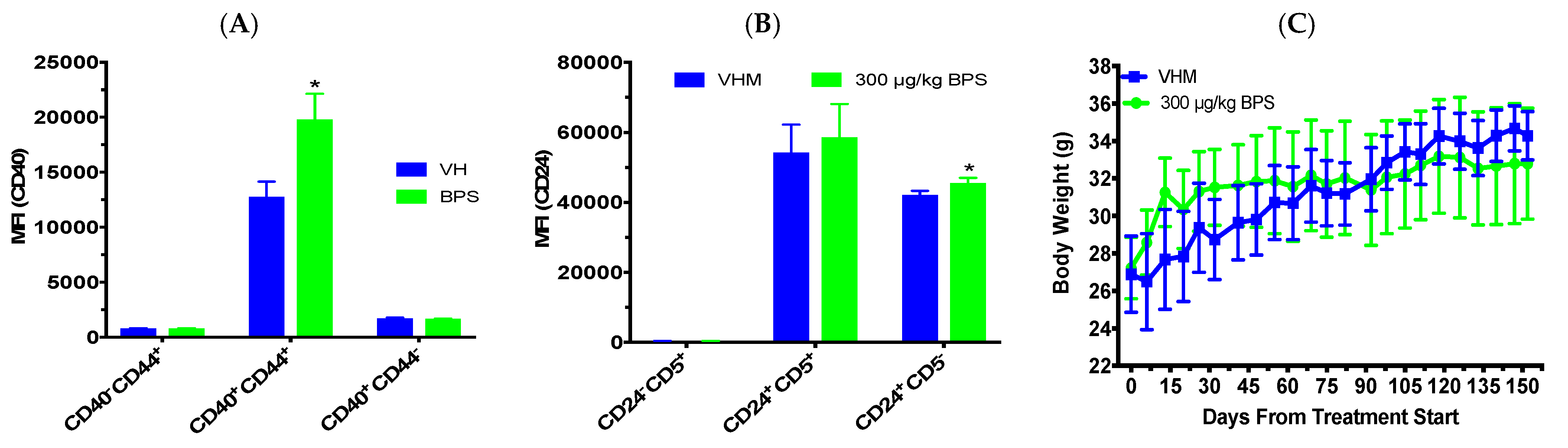
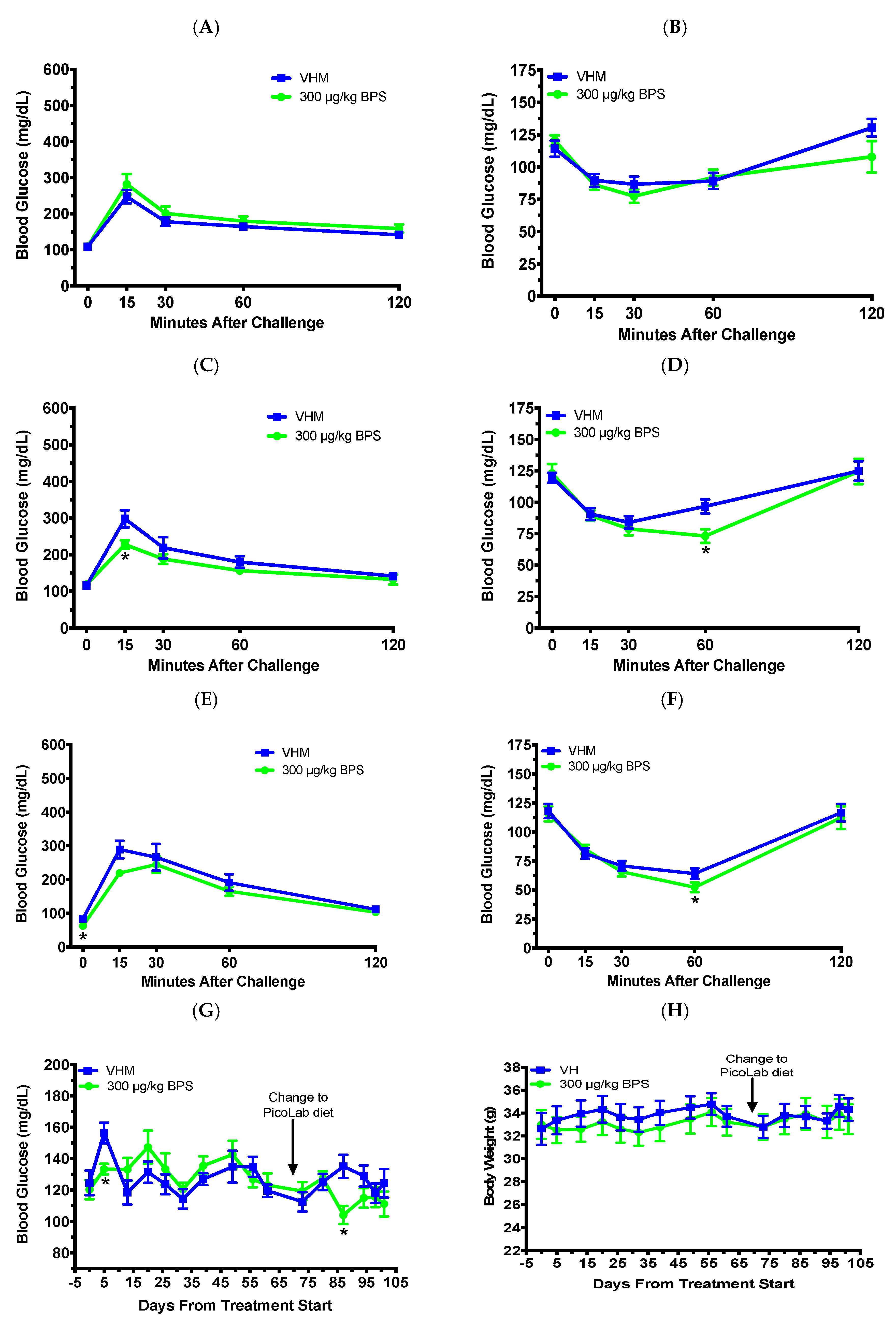
3.3. BPS Exposure Improved Glucose Tolerance and Insulin Sensitivity in Males on a Phytoestrogen-Free Diet
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wu, L.-H.; Zhang, X.-M.; Wang, F.; Gao, C.-J.; Chen, D.; Palumbo, J.R.; Guo, Y.; Zeng, E.Y. Occurrence of bisphenol S in the environment and implications for human exposure: A short review. Sci. Total Environ. 2018, 615, 87–98. [Google Scholar] [CrossRef]
- Usman, A.; Ahmad, M. From BPA to its analogues: Is it a safe journey? Chemosphere 2016, 158, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Del Moral, L.I.; Le Corre, L.; Poirier, H.; Niot, I.; Truntzer, T.; Merlin, J.-F.; Rouimi, P.; Besnard, P.; Rahmani, R.; Chagnon, M.-C. Obesogen effects after perinatal exposure of 4, 4′-sulfonyldiphenol (Bisphenol S) in C57BL/6 mice. Toxicology 2016, 357, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Tang, Z.; Yan, J.; Fang, J.; Wang, H.; Cai, Z. Bisphenol S exposure modulate macrophage phenotype as defined by cytokines profiling, global metabolomics and lipidomics analysis. Sci. Total Environ. 2017, 592, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Jiang, G.; Wei, P.; Wang, H.; Ru, S. Bisphenol S exposure impairs glucose homeostasis in male zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2018, 147, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.L.; Kirkman, M.S.; Laffel, L.M.; Peters, A.L. Type 1 diabetes through the life span: A position statement of the American Diabetes Association. Diabetes Care 2014, 37, 2034–2054. [Google Scholar] [CrossRef] [PubMed]
- Beydoun, H.A.; Khanal, S.; Zonderman, A.B.; Beydoun, M.A. Sex differences in the association of urinary bisphenol-A concentration with selected indices of glucose homeostasis among US adults. Ann. Epidemiol. 2014, 24, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Bodin, J.; Bolling, A.K.; Samuelsen, M.; Becher, R.; Lovik, M.; Nygaard, U.C. Long-term bisphenol A exposure accelerates insulitis development in diabetes-prone NOD mice. Immunopharmacol. Immunotoxicol. 2013, 35, 349–358. [Google Scholar] [CrossRef]
- Bodin, J.; Bølling, A.K.; Wendt, A.; Eliasson, L.; Becher, R.; Kuper, F.; Løvik, M.; Nygaard, U.C. Exposure to bisphenol A, but not phthalates, increases spontaneous diabetes type 1 development in NOD mice. Toxicol. Rep. 2015, 2, 99–110. [Google Scholar] [CrossRef]
- Xu, J.; Huang, G.; Nagy, T.; Teng, Q.; Guo, T.L. Sex-dependent effects of bisphenol A on type 1 diabetes development in non-obese diabetic (NOD) mice. Arch. Toxicol. 2019, 93, 997–1008. [Google Scholar] [CrossRef]
- Xu, J.; Huang, G.; Nagy, T.; Guo, T.L. Bisphenol A alteration of type 1 diabetes in non-obese diabetic (NOD) female mice is dependent on window of exposure. Arch. Toxicol. 2019, 93, 1083–1093. [Google Scholar] [CrossRef] [PubMed]
- Cetkovic-Cvrlje, M.; Thinamany, S.; Bruner, K.A. Bisphenol A (BPA) aggravates multiple low-dose streptozotocin-induced Type 1 diabetes in C57BL/6 mice. J. Immunotoxicol. 2017, 14, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.L.; Wang, Y.; Xiong, T.; Ling, X.; Zheng, J. Genistein modulation of streptozotocin diabetes in male B6C3F1 mice can be induced by diet. Toxicol. Appl. Pharmacol. 2014, 280, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.A.; Wong, F.S.; Wen, L. The importance of the Non Obese Diabetic (NOD) mouse model in autoimmune diabetes. J. Autoimmun. 2016, 66, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Delclos, K.B.; Camacho, L.; Lewis, S.M.; Vanlandingham, M.M.; Latendresse, J.R.; Olson, G.R.; Davis, K.J.; Patton, R.E.; da Costa, G.G.; Woodling, K.A. Toxicity evaluation of bisphenol A administered by gavage to Sprague-Dawley rats from gestation day 6 through postnatal day 90. Toxicol. Sci. 2014, 139, 174–197. [Google Scholar] [CrossRef]
- Johnson, S.A.; Javurek, A.B.; Painter, M.S.; Ellersieck, M.R.; Welsh, T.H.; Camacho, L.; Lewis, S.M.; Vanlandingham, M.M.; Ferguson, S.A.; Rosenfeld, C.S. Effects of developmental exposure to bisphenol A on spatial navigational learning and memory in rats: A CLARITY-BPA study. Horm. Behav. 2016, 80, 139–148. [Google Scholar] [CrossRef]
- King, A.J. The use of animal models in diabetes research. Br. J. Pharmacol. 2012, 166, 877–894. [Google Scholar] [CrossRef]
- Huang, G.; Xu, J.; Lefever, D.E.; Glenn, T.C.; Nagy, T.; Guo, T.L. Genistein prevention of hyperglycemia and improvement of glucose tolerance in adult non-obese diabetic mice are associated with alterations of gut microbiome and immune homeostasis. Toxicol. Appl. Pharmacol. 2017, 332, 138–148. [Google Scholar] [CrossRef]
- Katchy, A.; Pinto, C.; Jonsson, P.; Nguyen-Vu, T.; Pandelova, M.; Riu, A.; Schramm, K.-W.; Samarov, D.; Gustafsson, J.-Å.; Bondesson, M. Coexposure to phytoestrogens and bisphenol a mimics estrogenic effects in an additive manner. Toxicol. Sci. 2014, 138, 21–35. [Google Scholar] [CrossRef]
- Schmidt, S.; Degen, G.H.; Seibel, J.; Hertrampf, T.; Vollmer, G.; Diel, P. Hormonal activity of combinations of genistein, bisphenol A and 17β-estradiol in the female Wistar rat. Arch. Toxicol. 2006, 80, 839–845. [Google Scholar] [CrossRef]
- Taylor, J.A.; vom Saal, F.S.; Welshons, W.V.; Drury, B.; Rottinghaus, G.; Hunt, P.A.; Toutain, P.-L.; Laffont, C.M.; VandeVoort, C.A. Similarity of bisphenol A pharmacokinetics in rhesus monkeys and mice: Relevance for human exposure. Environ. Health Perspect. 2011, 119, 422. [Google Scholar] [CrossRef] [PubMed]
- Susiarjo, M.; Xin, F.; Bansal, A.; Stefaniak, M.; Li, C.; Simmons, R.A.; Bartolomei, M.S. Bisphenol a exposure disrupts metabolic health across multiple generations in the mouse. Endocrinology 2015, 156, 2049–2058. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.-B.; Luan, J.-N.; Ye, J.; Chen, S.-Y. RGC32 deficiency protects against high-fat diet-induced obesity and insulin resistance in mice. J. Endocrinol. 2015, 224, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, B.D.; Brandt, J.Z.; Grassi, T.F.; Silveira, L.T.R.; Scarano, W.R.; Barbisan, L.F. Genistein reduces the noxious effects of in utero bisphenol A exposure on the rat prostate gland at weaning and in adulthood. Food Chem. Toxicol. 2015, 84, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jenkins, S.; Lamartiniere, C.A. Cell proliferation and apoptosis in rat mammary glands following combinational exposure to bisphenol A and genistein. BMC Cancer 2014, 14, 379. [Google Scholar] [CrossRef] [PubMed]
- Gioiosa, L.; Palanza, P.; Parmigiani, S.; vom Saal, F.S. Risk evaluation of endocrine-disrupting chemicals: Effects of developmental exposure to low doses of bisphenol A on behavior and physiology in mice (Mus musculus). Dose-Response 2015, 13, 1559325815610760. [Google Scholar] [CrossRef] [PubMed]
- Kinch, C.D.; Ibhazehiebo, K.; Jeong, J.-H.; Habibi, H.R.; Kurrasch, D.M. Low-dose exposure to bisphenol A and replacement bisphenol S induces precocious hypothalamic neurogenesis in embryonic zebrafish. Proc. Natl. Acad. Sci. USA 2015, 112, 1475–1480. [Google Scholar] [CrossRef] [PubMed]
- D’Alise, A.M.; Auyeung, V.; Feuerer, M.; Nishio, J.; Fontenot, J.; Benoist, C.; Mathis, D. The defect in T-cell regulation in NOD mice is an effect on the T-cell effectors. Proc. Natl. Acad. Sci. USA 2008, 105, 19857–19862. [Google Scholar] [CrossRef] [PubMed]
- Apte, S.H.; Baz, A.; Groves, P.; Kelso, A.; Kienzle, N. Interferon-γ and interleukin-4 reciprocally regulate CD8 expression in CD8+ T cells. Proc. Natl. Acad. Sci. USA 2008, 105, 17475–17480. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Prins, G.S. Clarity in the face of confusion: New studies tip the scales on bisphenol A (BPA). Andrology 2016, 4, 561. [Google Scholar] [CrossRef]
- Bodin, J.; Bolling, A.K.; Becher, R.; Kuper, F.; Lovik, M.; Nygaard, U.C. Transmaternal bisphenol A exposure accelerates diabetes type 1 development in NOD mice. Toxicol. Sci. 2014, 137, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Dendrou, C.A.; Wicker, L.S. The IL-2/CD25 pathway determines susceptibility to T1D in humans and NOD mice. J. Clin. Immunol. 2008, 28, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.S.; Jung, U.J.; Yeo, J.; Kim, M.J.; Lee, M.K. Genistein and daidzein prevent diabetes onset by elevating insulin level and altering hepatic gluconeogenic and lipogenic enzyme activities in non-obese diabetic (NOD) mice. Diabetes Metab. Res. Rev. 2008, 24, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.L.; Germolec, D.R.; Zheng, J.F.; Kooistra, L.; Auttachoat, W.; Smith, M.J.; White, K.L.; Elmore, S.A. Genistein protects female nonobese diabetic mice from developing type 1 diabetes when fed a soy-and alfalfa-free diet. Toxicol. Pathol. 2015, 43, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, E.; Ladeira, C.; Viegas, S. EDCs mixtures: A stealthy hazard for human health? Toxics 2017, 5, 5. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody Combinations | Group | +/− (%) | +/+ (%) | −/+ (%) |
|---|---|---|---|---|
| CD3/CD45R | VHF | 9.87 ± 0.89 | 0.81 ± 0.16 | 36.93 ± 5.66 |
| 3 | 10.72 ± 1.91 | 0.93 ± 0.12 | 35.78 ± 2.17 | |
| 30 | 9.18 ± 0.42 | 0.86 ± 0.06 | 34.27 ± 1.29 | |
| 150 | 12.04 ± 1.58 | 1.17 ± 0.17 | 37.40 ± 1.93 | |
| 300 | 9.96 ± 0.59 | 1.18 ± 0.11 | 36.24 ± 1.36 | |
| CD4/CD8 | VHF | 7.44 ± 1.17 | 1.82 ± 0.10 | 9.26 ± 1.29 |
| 3 | 7.70 ± 1.31 | 1.62 ± 0.27 | 8.39 ± 0.83 | |
| 30 | 6.38 ± 0.47 | 1.94 ± 0.13 | 10.54 ± 0.87 | |
| 150 | 8.53 ± 1.17 | 2.39 ± 0.11 | 11.47 ± 0.37 | |
| 300 | 6.82 ± 0.34 | 2.42 ± 0.20 | 12.64 ± 1.46 | |
| CD4/CD25 | VHF | 7.68 ± 0.98 | 0.97 ± 0.12 | 0.36 ± 0.07 |
| 3 | 7.73 ± 1.33 | 1.11 ± 0.12 | 0.48 ± 0.05 | |
| 30 | 6.83 ± 0.43 | 0.91 ± 0.09 | 0.40 ± 0.03 | |
| 150 | 8.87 ± 1.22 | 1.36 ± 0.02 | 0.61 ± 0.09 * | |
| 300 | 7.63 ± 0.46 | 1.00 ± 0.93 | 0.50 ± 0.04 | |
| CD8/CD25 | VHF | 5.71 ± 0.61 | 0.37 ± 0.10 | 0.85 ± 0.11 |
| 3 | 5.66 ± 0.80 | 0.39 ± 0.05 | 1.06 ± 0.11 | |
| 30 | 6.85 ± 0.62 | 0.32 ± 0.03 | 0.87 ± 0.08 | |
| 150 | 8.14 ± 0.44 | 0.49 ± 0.05 | 1.36 ± 0.09 * | |
| 300 | 8.78 ± 1.03 | 0.49 ± 0.05 | 0.86 ± 0.06 | |
| CD24/CD5 | VHF | 67.40 ± 1.23 | 4.50 ± 0.03 | 16.47 ± 0.73 |
| 3 | 63.17 ± 2.60 | 4.18 ± 0.31 | 19.77 ± 3.02 | |
| 30 | 66.95 ± 1.05 | 4.35 ± 0.36 | 16.08 ± 0.61 | |
| 150 | 63.37 ± 2.05 | 4.36 ± 0.35 | 19.50 ± 2.85 | |
| 300 | 66.42 ± 1.50 | 4.47 ± 0.24 | 16.32 ± 1.09 | |
| CD40/CD44 | VHF | 32.77 ± 2.35 | 3.83 ± 0.64 | 2.61 ± 0.59 |
| 3 | 33.50 ± 2.68 | 3.89 ± 0.45 | 2.61 ± 0.28 | |
| 30 | 30.32 ± 1.58 | 3.67 ± 0.27 | 3.00 ± 0.13 | |
| 150 | 34.23 ± 2.95 | 3.59 ± 0.87 | 2.51 ± 0.55 | |
| 300 | 32.30 ± 1.60 | 2.61 ± 0.24 | 2.34 ± 0.20 | |
| B220/CD40L | VHF | 28.53 ± 5.97 | 0.59 ± 0.11 | 1.66 ± 0.16 |
| 3 | 33.10 ± 2.83 | 0.59 ± 0.08 | 1.62 ± 0.20 | |
| 30 | 29.97 ± 2.67 | 0.53 ± 0.09 | 1.83 ± 0.35 | |
| 150 | 34.10 ± 3.91 | 0.58 ± 0.11 | 1.48 ± 0.21 | |
| 300 | 29.52 ± 1.91 | 0.42 ± 0.07 | 1.21 ± 0.15 | |
| Mac3/CD45R | VHF | 5.57 ± 0.50 | 7.15 ± 1.29 | 29.33 ± 4.63 |
| 3 | 5.57 ± 0.27 | 8.32 ± 0.68 | 27.27 ± 1.76 | |
| 30 | 5.70 ± 0.32 | 7.94 ± 0.88 | 25.87 ± 0.70 | |
| 150 | 6.44 ± 0.67 | 11.10 ± 0.71 | 26.13 ± 1.47 | |
| 300 | 5.65 ± 0.67 | 10.32 ± 1.13 | 25.58 ± 0.76 |
| Antibody Combinations | Group | +/− (%) | +/+ (%) | -/+ (%) |
|---|---|---|---|---|
| CD4/CD8 | VHM | 10.03 ± 1.27 | 2.03 ± 0.51 | 13.02 ± 0.67 |
| 300 | 9.60 ± 0.44 | 1.56 ± 0.31 | 12.94 ± 1.45 | |
| CD4/CD25 | VHM | 10.56 ± 0.81 | 1.19 ± 0.09 | 1.24 ± 0.29 |
| 300 | 9.85 ± 0.65 | 1.03 ± 0.10 | 1.12 ± 0.30 | |
| CD8/CD25 | VHM | 8.44 ± 0.48 | 0.60 ± 0.11 | 1.67 ± 0.19 |
| 300 | 8.89 ± 1.73 | 0.53 ± 0.08 | 1.47 ± 0.32 | |
| CD24/CD5 | VHM | 66.40 ± 1.93 | 2.17 ± 0.10 | 23.00 ± 1.56 |
| 300 | 65.94 ± 2.33 | 2.25 ± 0.39 | 24.44 ± 2.37 | |
| CD40/CD44 | VHM | 18.74 ± 0.89 | 2.20 ± 0.27 | 3.12 ± 0.21 |
| 300 | 16.83 ± 1.43 | 1.77 ± 0.15 | 2.95 ± 0.26 | |
| B220/CD40L | VHM | 15.64 ±1.25 | 0.86 ±0.08 | 3.14±0.15 |
| 300 | 12.09 ±1.58 | 0.71 ±0.11 | 3.13±0.26 |
| Antibody Combinations | Group | +/− (%) | +/+ (%) | -/+ (%) |
|---|---|---|---|---|
| CD4/CD8 | VHM | 10.17 ± 0.68 | 1.23 ± 0.12 | 10.48 ± 0.55 |
| 300 | 7.83 ± 0.91 | 2.08 ± 0.66 | 10.02 ± 1.29 | |
| CD4/CD25 | VHM | 9.46 ± 0.78 | 1.62 ± 0.12 | 3.30 ± 0.48 |
| 300 | 8.61 ± 0.60 | 0.94 ± 0.19 * | 1.49 ± 0.31 * | |
| CD8/CD25 | VHM | 5.92 ± 0.40 | 1.14 ± 0.10 | 3.68 ± 0.48 |
| 300 | 6.28 ± 0.90 | 0.80 ± 0.08 * | 1.57 ± 0.44 * | |
| CD24/CD5 | VHM | 71.91 ± 1.63 | 1.88 ± 0.06 | 19.44 ± 1.30 |
| 300 | 73.37 ± 1.26 | 1.57 ± 0.13 | 17.73 ± 1.15 | |
| CD40/CD44 | VHM | 13.80 ± 0.69 | 2.10 ± 0.15 | 3.65 ± 0.29 |
| 300 | 12.07 ± 0.50 | 1.70 ± 0.16 | 3.91 ± 0.64 | |
| B220/CD40L | VHM | 12.21 ± 0.84 | 0.71 ± 0.05 | 2.60 ± 0.20 |
| 300 | 10.87 ± 0.72 | 0.71 ± 0.05 | 2.28 ± 0.13 | |
| MAC-3/CD18 | VHM | 1.92 ± 0.23 | 2.49 ± 0.27 | 1.88 ± 0.19 |
| 300 | 1.67 ± 0.09 | 2.15 ± 0.16 | 1.71 ± 0.32 |
| Absolute Organ Weights (mg) | |||||
| Treatment | Spleen | Pancreas | Kidneys | Liver | Thymus |
| Vehicle | 84.0 ± 3.5 | 236.5 ± 12.3 | 487.8 ± 12.8 | 1750.7 ± 59.8 | 28.5 ± 1.7 |
| 300 μg/kg BPS | 80.0 ± 6.0 | 219.2 ± 12.8 | 494.8 ± 20.5 | 1696.8 ± 70.3 | 19.1 ± 1.4 * |
| Organ to Body Weight (%) | |||||
| Treatment | %Spleen | %Pancreas | %Kidneys | %Liver | %Thymus |
| Vehicle | 0.25 ± 0.01 | 0.69 ± 0.03 | 1.42 ± 0.02 | 5.10 ± 0.07 | 0.08 ± 0.01 |
| 300 μg/kg BPS | 0.24 ± 0.01 | 0.66 ± 0.03 | 1.50 ± 0.02 * | 5.07 ± 0.08 | 0.06 ± 0.00 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Huang, G.; Guo, T.L. Bisphenol S Modulates Type 1 Diabetes Development in Non-Obese Diabetic (NOD) Mice with Diet- and Sex-Related Effects. Toxics 2019, 7, 35. https://doi.org/10.3390/toxics7020035
Xu J, Huang G, Guo TL. Bisphenol S Modulates Type 1 Diabetes Development in Non-Obese Diabetic (NOD) Mice with Diet- and Sex-Related Effects. Toxics. 2019; 7(2):35. https://doi.org/10.3390/toxics7020035
Chicago/Turabian StyleXu, Joella, Guannan Huang, and Tai L. Guo. 2019. "Bisphenol S Modulates Type 1 Diabetes Development in Non-Obese Diabetic (NOD) Mice with Diet- and Sex-Related Effects" Toxics 7, no. 2: 35. https://doi.org/10.3390/toxics7020035
APA StyleXu, J., Huang, G., & Guo, T. L. (2019). Bisphenol S Modulates Type 1 Diabetes Development in Non-Obese Diabetic (NOD) Mice with Diet- and Sex-Related Effects. Toxics, 7(2), 35. https://doi.org/10.3390/toxics7020035