
Embryonic Ethanol Exposure Affects Early- and Late-Added Cardiac Precursors and Produces Long-Lasting Heart Chamber Defects in Zebrafish


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry and Ethanol Treatment
2.2. Histology
2.3. Immunofluorescence
2.4. Fluorescence in Situ Hybridization (FISH)
2.5. Microscopy and Cell Counting
2.6. Statistical Analysis
3. Results
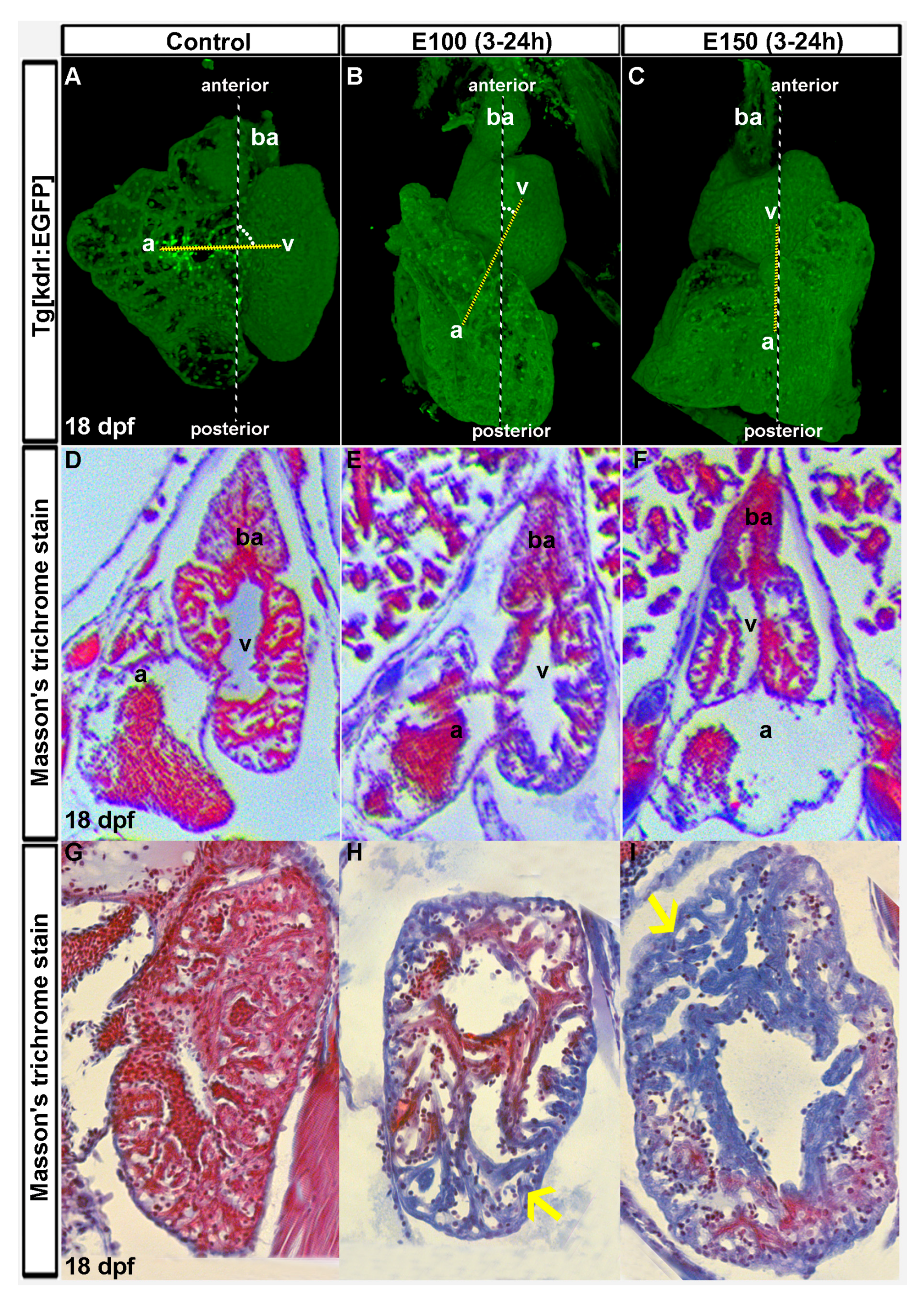
3.1. Ethanol-induced Cardiac Chamber Defects Occurred during Zebrafish Embryonic Development and Persisted in Larvae
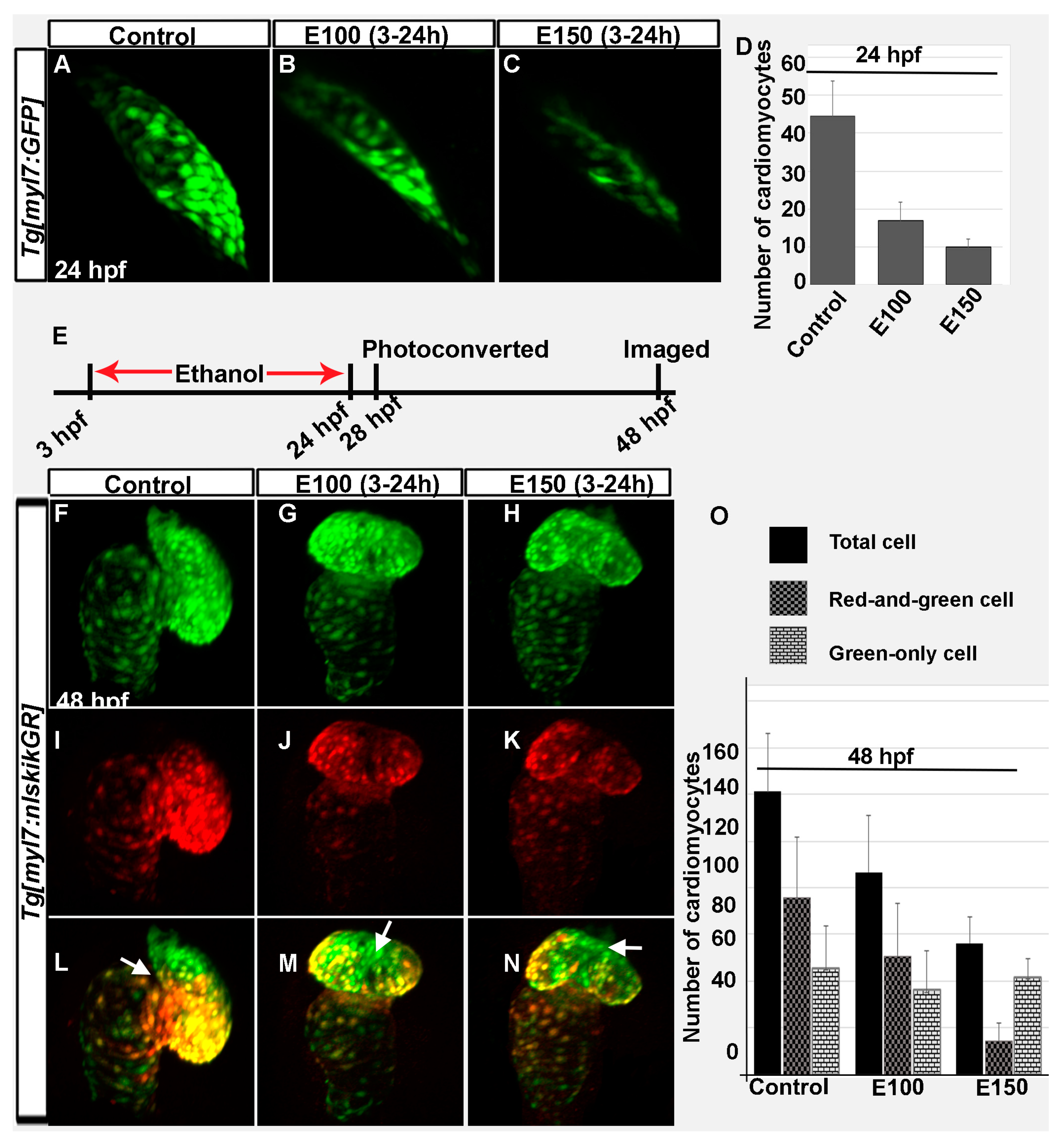
3.2. Embryonic Ethanol Exposure Reduced Early and Late Added Cardiomyocytes in the Heart
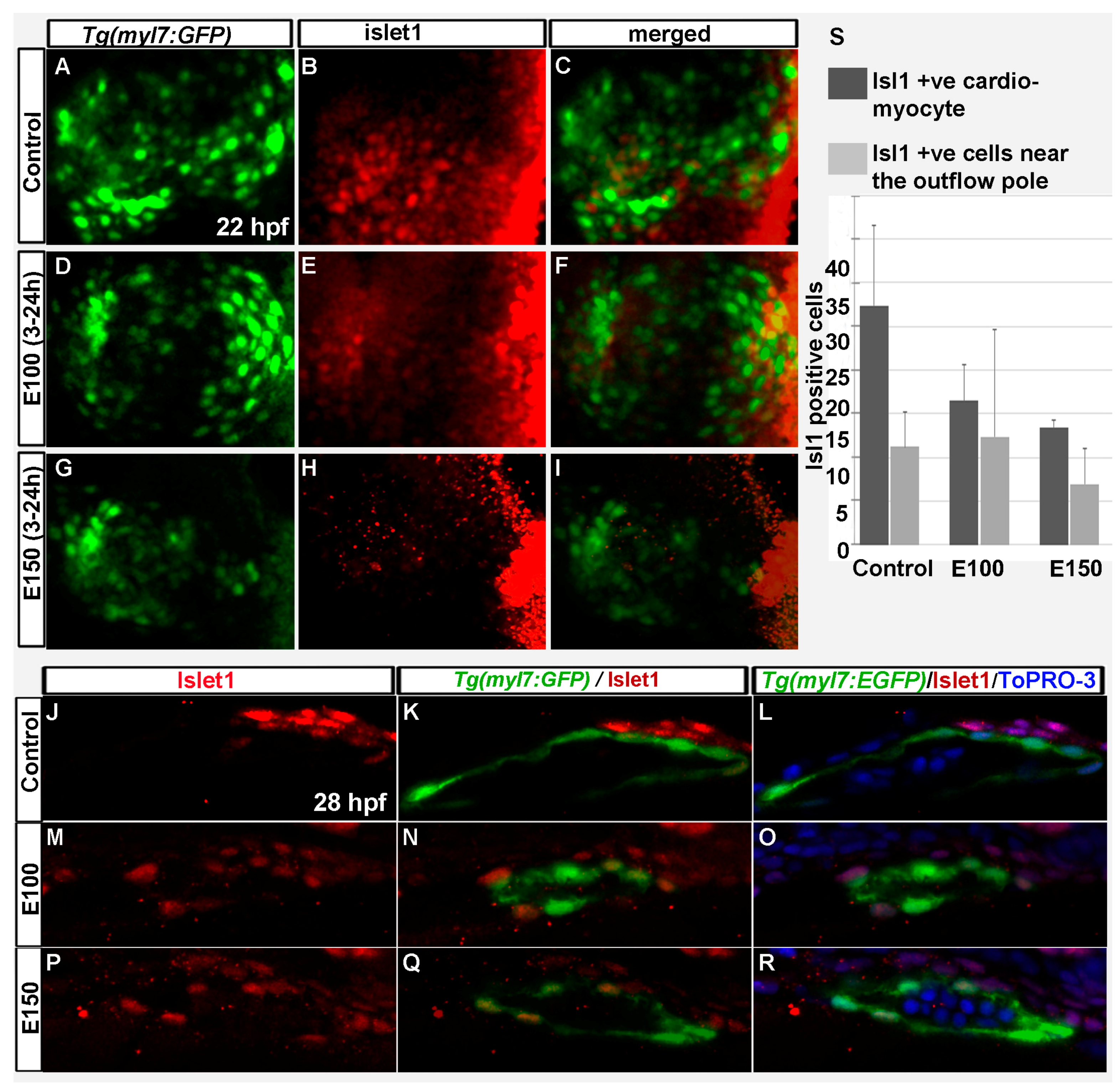
3.3. Early Ethanol Exposure Delayed Cardiac Differentiation
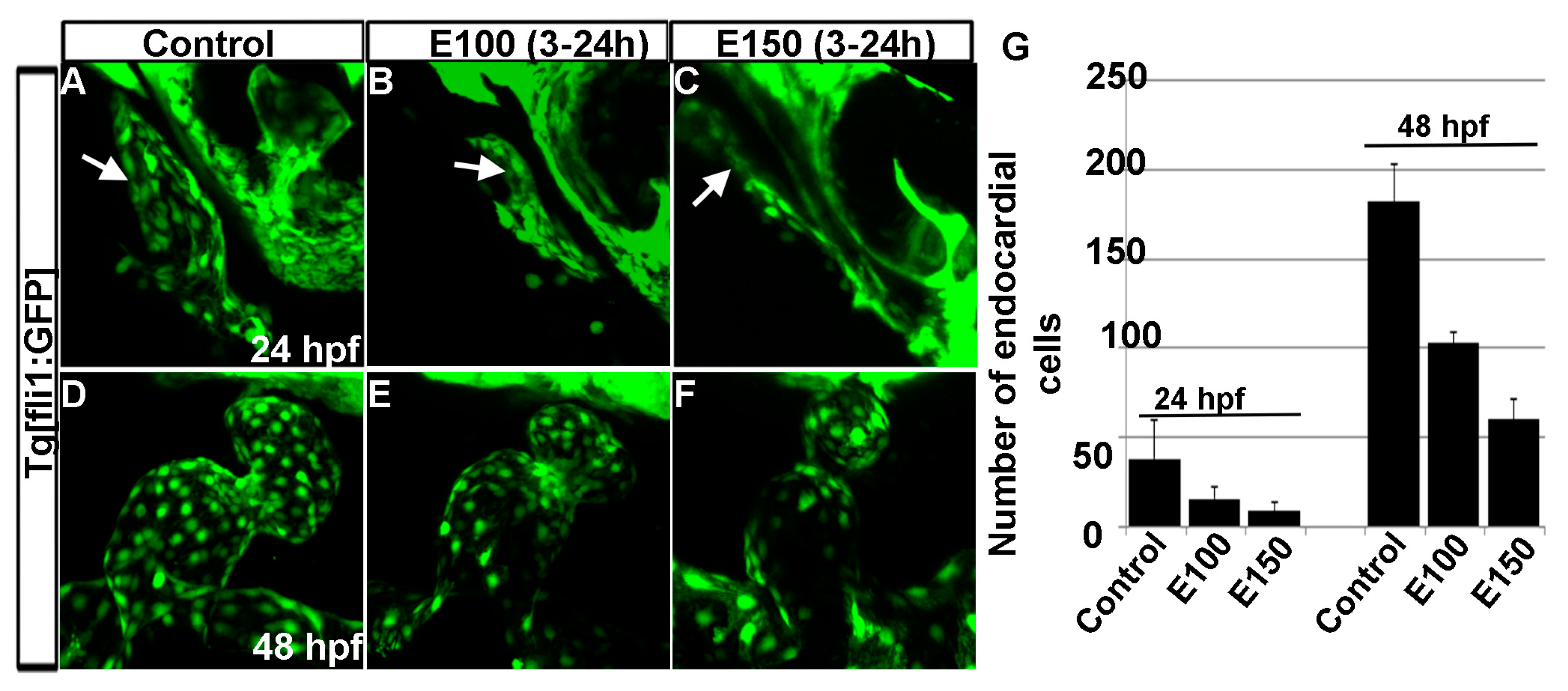
3.4. Embryonic Ethanol Exposure Reduced Endocardial Cell Number
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- May, P.A.; Gossage, J.P. Estimating the prevalence of fetal alcohol syndrome. A summary. Alcohol Res. Health 2001, 25, 159–167. [Google Scholar] [PubMed]
- Nash, A.; Davies, L. Fetal Alcohol Spectrum Disorders: What Pediatric Providers Need to Know. J. Pediatr. Health Care 2017, 31, 594–606. [Google Scholar] [CrossRef] [PubMed]
- Abel, E.L. An update on incidence of FAS: FAS is not an equal opportunity birth defect. Neurotoxicol. Teratol. 1995, 17, 437–443. [Google Scholar] [CrossRef]
- Abel, E.L.; Hannigan, J.H. Maternal risk factors in fetal alcohol syndrome: provocative and permissive influences. Neurotoxicol. Teratol. 1995, 17, 445–462. [Google Scholar] [CrossRef]
- Burd, L.; Deal, E.; Rios, R.; Adickes, E.; Wynne, J.; Klug, M.G. Congenital heart defects and fetal alcohol spectrum disorders. Congenit. Heart Dis. 2007, 2, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.J.; Lin, C.Y.; Chen, C.H.; Zhou, B.; Chang, C.P. Partitioning the heart: mechanisms of cardiac septation and valve development. Development 2012, 139, 3277–3299. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Cashman, T.J.; Nevis, K.R.; Obregon, P.; Carney, S.A.; Liu, Y.; Gu, A.; Mosimann, C.; Sondalle, S.; Peterson, R.E.; et al. Latent TGF-beta binding protein 3 identifies a second heart field in zebrafish. Nature 2011, 474, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Vincent, S.D.; Buckingham, M.E. How to make a heart: the origin and regulation of cardiac progenitor cells. Curr. Top. Dev. Biol. 2010, 90, 1–41. [Google Scholar] [PubMed]
- Staudt, D.; Stainier, D. Uncovering the molecular and cellular mechanisms of heart development using the zebrafish. Annu. Rev. Genet. 2012, 46, 397–418. [Google Scholar] [CrossRef] [PubMed]
- Hami, D.; Grimes, A.C.; Tsai, H.J.; Kirby, M.L. Zebrafish cardiac development requires a conserved secondary heart field. Development 2011, 138, 2389–2398. [Google Scholar] [CrossRef] [PubMed]
- Lazic, S.; Scott, I.C. Mef2cb regulates late myocardial cell addition from a second heart field-like population of progenitors in zebrafish. Dev. Biol. 2011, 354, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Stainier, D.Y. Zebrafish genetics and vertebrate heart formation. Nat. Rev. Genet. 2001, 2, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Lints, T.J.; Parsons, L.M.; Hartley, L.; Lyons, I.; Harvey, R.P. Nkx-2.5: A novel murine homeobox gene expressed in early heart progenitor cells and their myogenic descendants. Development 1993, 119, 969. [Google Scholar] [PubMed]
- Chen, J.N.; Fishman, M.C. Zebrafish tinman homolog demarcates the heart field and initiates myocardial differentiation. Development 1996, 122, 3809–3816. [Google Scholar] [PubMed]
- Stainier, D.Y.; Lee, R.K.; Fishman, M.C. Cardiovascular development in the zebrafish. I. Myocardial fate map and heart tube formation. Development 1993, 119, 31–40. [Google Scholar] [PubMed]
- De Pater, E.; Clijsters, L.; Marques, S.R.; Lin, Y.F.; Garavito-Aguilar, Z.V.; Yelon, D.; Bakkers, J. Distinct phases of cardiomyocyte differentiation regulate growth of the zebrafish heart. Development 2009, 136, 1633–1641. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.X.; Zdanowicz, M.; Young, L.; Kumiski, D.; Leatherbury, L.; Kirby, M.L. Cardiac neural crest in zebrafish embryos contributes to myocardial cell lineage and early heart function. Dev. Dyn. 2003, 226, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Yost, H.J. Cardiac neural crest contributes to cardiomyogenesis in zebrafish. Dev. Biol. 2003, 257, 127–139. [Google Scholar] [CrossRef]
- Cavanaugh, A.M.; Huang, J.; Chen, J.N. Two developmentally distinct populations of neural crest cells contribute to the zebrafish heart. Dev. Biol. 2015, 404, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Cudd, T.A. Animal model systems for the study of alcohol teratology. Exp. Biol. Med. (Maywood) 2005, 230, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Haycock, P.C. Fetal alcohol spectrum disorders: The epigenetic perspective. Biol. Reprod. 2009, 81, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Sulik, K.K. Genesis of alcohol-induced craniofacial dysmorphism. Exp. Biol. Med. (Maywood) 2005, 230, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, Y.; Buckley, D.M.; Eberhart, J.K. Diving into the world of alcohol teratogenesis: A review of zebrafish models of fetal alcohol spectrum disorders. Biochem. Cell Biol. Biochim. Biol. Cell. 2017. [Google Scholar] [CrossRef] [PubMed]
- Lovely, C.; Rampersad, M.; Fernandes, Y.; Eberhart, J. Gene-environment interactions in development and disease. Wiley Interdiscip. Rev. Dev. Biol. 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Sarmah, S.; Muralidharan, P.; Curtis, C.L.; McClintick, J.N.; Buente, B.B.; Holdgrafer, D.J.; Ogbeifun, O.; Olorungbounmi, O.C.; Patino, L.; Lucas, R.; et al. Ethanol exposure disrupts extraembryonic microtubule cytoskeleton and embryonic blastomere cell adhesion, producing epiboly and gastrulation defects. Biol. Open 2013, 2, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Muralidharan, P.; Sarmah, S.; Zhou, F.C.; Marrs, J.A. Fetal Alcohol Spectrum Disorder (FASD) Associated Neural Defects: Complex Mechanisms and Potential Therapeutic Targets. Brain Sci. 2013, 3, 964–991. [Google Scholar] [CrossRef] [PubMed]
- Muralidharan, P.; Sarmah, S.; Marrs, J.A. Zebrafish retinal defects induced by ethanol exposure are rescued by retinoic acid and folic acid supplement. Alcohol 2015, 49, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Sarmah, S.; Barrallo-Gimeno, A.; Melville, D.B.; Topczewski, J.; Solnica-Krezel, L.; Knapik, E.W. Sec24D-dependent transport of extracellular matrix proteins is required for zebrafish skeletal morphogenesis. PLoS ONE 2010, 5, e10367. [Google Scholar] [CrossRef] [PubMed]
- Sarmah, S.; Muralidharan, P.; Marrs, J.A. Embryonic Ethanol Exposure Dysregulates BMP and Notch Signaling, Leading to Persistent Atrio-Ventricular Valve Defects in Zebrafish. PLoS ONE 2016, 11, e0161205. [Google Scholar] [CrossRef] [PubMed]
- Gut, P.; Reischauer, S.; Stainier, D.Y.R.; Arnaout, R. Little Fish, Big Data: Zebrafish as a Model for Cardiovascular and Metabolic Disease. Physiol. Rev. 2017, 97, 889–938. [Google Scholar] [CrossRef] [PubMed]
- Grant, M.G.; Patterson, V.L.; Grimes, D.T.; Burdine, R.D. Modeling Syndromic Congenital Heart Defects in Zebrafish. Curr. Top. Dev. Biol. 2017, 124, 1–40. [Google Scholar] [PubMed]
- Sarmah, S.; Marrs, J.A. Complex cardiac defects after ethanol exposure during discrete cardiogenic events in zebrafish: prevention with folic acid. Dev. Dyn. 2013, 242, 1184–1201. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book; The University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- Brend, T.; Holley, S.A. Zebrafish whole mount high-resolution double fluorescent in situ hybridization. J. Vis. Exp. JoVE 2009, 25, 1229. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rosa, J.M.; Martin, V.; Peralta, M.; Torres, M.; Mercader, N. Extensive scar formation and regression during heart regeneration after cryoinjury in zebrafish. Development 2011, 138, 1663–1674. [Google Scholar] [CrossRef] [PubMed]
- Black, B.L. Transcriptional pathways in second heart field development. Semin. Cell Dev. Biol. 2007, 18, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Cardiovascular Diseases (CVDs). Available online: http://www.who.int/mediacentre/factsheets/fs317/en/ (accessed on 13 December 2016).
- Sarmah, S.; Muralidharan, P.; Marrs, J.A. Common congenital anomalies: Environmental causes and prevention with folic acid containing multivitamins. Birth Defects Res. Part C Embryo Today Rev. 2016, 108, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Sarmah, S.; Marrs, J.A. Zebrafish as a Vertebrate Model System to Evaluate Effects of Environmental Toxicants on Cardiac Development and Function. Int. J. Mol. Sci. 2016, 17, 2123. [Google Scholar] [CrossRef] [PubMed]
- Bruyere, H.J., Jr.; Stith, C.E. Strain-dependent effect of ethanol on ventricular septal defect frequency in White Leghorn chick embryos. Teratology 1993, 48, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Cavieres, M.F.; Smith, S.M. Genetic and developmental modulation of cardiac deficits in prenatal alcohol exposure. Alcohol. Clin. Exp. Res. 2000, 24, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Daft, P.A.; Johnston, M.C.; Sulik, K.K. Abnormal heart and great vessel development following acute ethanol exposure in mice. Teratology 1986, 33, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rosa, J.M.; Burns, C.E.; Burns, C.G. Zebrafish heart regeneration: 15 years of discoveries. Regeneration 2017, 4, 105–123. [Google Scholar] [CrossRef] [PubMed]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarmah, S.; Marrs, J.A. Embryonic Ethanol Exposure Affects Early- and Late-Added Cardiac Precursors and Produces Long-Lasting Heart Chamber Defects in Zebrafish. Toxics 2017, 5, 35. https://doi.org/10.3390/toxics5040035
Sarmah S, Marrs JA. Embryonic Ethanol Exposure Affects Early- and Late-Added Cardiac Precursors and Produces Long-Lasting Heart Chamber Defects in Zebrafish. Toxics. 2017; 5(4):35. https://doi.org/10.3390/toxics5040035
Chicago/Turabian StyleSarmah, Swapnalee, and James A. Marrs. 2017. "Embryonic Ethanol Exposure Affects Early- and Late-Added Cardiac Precursors and Produces Long-Lasting Heart Chamber Defects in Zebrafish" Toxics 5, no. 4: 35. https://doi.org/10.3390/toxics5040035
APA StyleSarmah, S., & Marrs, J. A. (2017). Embryonic Ethanol Exposure Affects Early- and Late-Added Cardiac Precursors and Produces Long-Lasting Heart Chamber Defects in Zebrafish. Toxics, 5(4), 35. https://doi.org/10.3390/toxics5040035