
Zebrafish Get Connected: Investigating Neurotransmission Targets and Alterations in Chemical Toxicity



Abstract
:1. Introduction
2. Zebrafish as a Biomedical Model
3. Review of Neurotransmitter Systems
3.1. Glutamate
3.2. GABA
3.3. Catecholamines
3.4. Serotonin
3.5. Acetylcholine
3.6. Histamine
3.7. Glycine
3.8. Other Neurotransmitters
4. Evaluation of Neurotoxicity
4.1. Evaluation of Neurotransmitters
4.2. Evaluation of Gene Expression Changes in Neurotransmitter Pathways
4.3. Evaluation of Neurobehavior
4.4. Visualization of Neurotransmitters and Neurotransmitter Systems
4.5. Pharmacology Screens
5. Specific Examples of Chemical Toxicity Targeting Neurotransmitter Systems
5.1. Drugs
5.2. Pesticides
5.3. Metals
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Megha, K.; Deshmukh, P.S.; Ravi, A.K.; Tripathi, A.K.; Abegaonkar, M.P.; Banerjee, B.D. Effect of low-intensity microwave radiation on monoamine neurotransmitters and their key regulating enzymes in rat brain. Cell Biochem. Biophys. 2015, 73, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, S.V.; Limbocker, R.A.; Gehringer, R.C.; Divis, J.L.; Osterhaus, G.L.; Newby, M.D.; Sofis, M.J.; Jarmolowicz, D.P.; Newman, B.D.; Mathews, T.A.; et al. Impaired brain dopamine and serotonin release and uptake in wistar rats following treatment with carboplatin. ACS Chem. Neurosci. 2016, 7, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Parng, C.; Roy, N.M.; Ton, C.; Lin, Y.; McGrath, P. Neurotoxicity assessment using zebrafish. J. Pharmacol. Toxicol. Methods 2007, 55, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Andersen, H.R.; Nielsen, J.B.; Grandjean, P. Toxicologic evidence of developmental neurotoxicity of environmental chemicals. Toxicology 2000, 144, 121–127. [Google Scholar] [CrossRef]
- Grandjean, P.; Landrigan, P.J. Developmental neurotoxicity of industrial chemicals. Lancet 2006, 368, 2167–2178. [Google Scholar] [CrossRef]
- Costa, L.G. Interactions of neurotoxicants with neurotransmitter systems. Toxicology 1988, 49, 359–366. [Google Scholar] [CrossRef]
- Beitz, J.M. Parkinson’s disease: A review. Front. Biosci. (Sch. Ed.) 2014, 6, 65–74. [Google Scholar] [CrossRef]
- Sarter, M.; Bruno, J.P.; Parikh, V. Abnormal neurotransmitter release underlying behavioral and cognitive disorders: Toward concepts of dynamic and function-specific dysregulation. Neuropsychopharmacology 2006, 32, 1452–1461. [Google Scholar] [CrossRef] [PubMed]
- Werner, F.M.; Covenas, R. Classical neurotransmitters and neuropeptides involved in major depression: A review. Int. J. Neurosci. 2010, 120, 455–470. [Google Scholar] [CrossRef] [PubMed]
- De Esch, C.; Slieker, R.; Wolterbeek, A.; Woutersen, R.; de Groot, D. Zebrafish as potential model for developmental neurotoxicity testing: A mini review. Neurotoxicol. Teratol. 2012, 34, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Rico, E.P.; Rosemberg, D.B.; Seibt, K.J.; Capiotti, K.M.; Da Silva, R.S.; Bonan, C.D. Zebrafish neurotransmitter systems as potential pharmacological and toxicological targets. Neurotoxicol. Teratol. 2011, 33, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.-Y.; Cowden, J.; Simmons, S.O.; Padilla, S.; Ramabhadran, R. Gene expression changes in developing zebrafish as potential markers for rapid developmental neurotoxicity screening. Neurotoxicol. Teratol. 2010, 32, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Murakami, S.; Ashikawa, Y.; Sasagawa, S.; Umemoto, N.; Shimada, Y.; Tanaka, T. Zebrafish as a systems toxicology model for developmental neurotoxicity testing. Congenit. Anom. 2015, 55, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.K.; Jesuthasan, S.J.; Penney, T.B. Zebrafish forebrain and temporal conditioning. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2014, 369, 20120462. [Google Scholar] [CrossRef] [PubMed]
- Mueller, T.; Wullimann, M.F. Atlas of Early Zebrafish Brain Development, 2nd ed.; Elsevier: San Diego, CA, USA, 2016. [Google Scholar]
- Panula, P.; Chen, Y.C.; Priyadarshini, M.; Kudo, H.; Semenova, S.; Sundvik, M.; Sallinen, V. The comparative neuroanatomy and neurochemistry of zebrafish CNS systems of relevance to human neuropsychiatric diseases. Neurobiol. Dis. 2010, 40, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Bowman, T.V.; Zon, L.I. Swimming into the future of drug discovery: In vivo chemical screens in zebrafish. ACS Chem. Biol. 2010, 5, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Lele, Z.; Krone, P.H. The zebrafish as a model system in developmental, toxicological and transgenic research. Biotechnol. Adv. 1996, 14, 57–72. [Google Scholar] [CrossRef]
- Yang, L.; Ho, N.Y.; Alshut, R.; Legradi, J.; Weiss, C.; Reischl, M.; Mikut, R.; Liebel, U.; Müller, F.; Strähle, U. Zebrafish embryos as models for embryotoxic and teratological effects of chemicals. Reprod. Toxicol. 2009, 28, 245–253. [Google Scholar] [CrossRef] [PubMed]
- McGrath, P.; Li, C.-Q. Zebrafish: A predictive model for assessing drug-induced toxicity. Drug Discov. Today 2008, 13, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Penberthy, W.T.; Shafizadeh, E.; Lin, S. The zebrafish as a model for human disease. Front. Biosci. 2002, 7, d1439–d1453. [Google Scholar] [CrossRef] [PubMed]
- Vascotto, S.G.; Beckham, Y.; Kelly, G.M. The zebrafish’s swim to fame as an experimental model in biology. Biochem. Cell Biol. 1997, 75, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Barbazuk, W.B.; Korf, I.; Kadavi, C.; Heyen, J.; Tate, S.; Wun, E.; Bedell, J.A.; McPherson, J.D.; Johnson, S.L. The syntenic relationship of the zebrafish and human genomes. Genome Res. 2000, 10, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Vandepoele, K.; De Vos, W.; Taylor, J.S.; Meyer, A.; Van de Peer, Y. Major events in the genome evolution of vertebrates: Paranome age and size differ considerably between ray-finned fishes and land vertebrates. Proc. Natl. Acad. Sci. USA 2004, 101, 1638–1643. [Google Scholar] [CrossRef] [PubMed]
- Christoffels, A.; Koh, E.G.; Chia, J.M.; Brenner, S.; Aparicio, S.; Venkatesh, B. Fugu genome analysis provides evidence for a whole-genome duplication early during the evolution of ray-finned fishes. Mol. Biol. Evol. 2004, 21, 1146–1151. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Hashiguchi, Y.; Nishida, M. Temporal pattern of loss/persistence of duplicate genes involved in signal transduction and metabolic pathways after teleost-specific genome duplication. BMC Evolut. Biol. 2009, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Glasauer, S.M.; Neuhauss, S.C. Whole-genome duplication in teleost fishes and its evolutionary consequences. Mol. Genet. Genom. 2014, 289, 1045–1060. [Google Scholar] [CrossRef] [PubMed]
- Postlethwait, J.; Amores, A.; Cresko, W.; Singer, A.; Yan, Y.L. Subfunction partitioning, the teleost radiation and the annotation of the human genome. Trends Genet. 2004, 20, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Braitenberg, V.; Schüz, A. Cortex: Statistics and Geometry of Neuronal Connectivity, 2nd ed.; Springer: Berlin, Germang, 1998. [Google Scholar]
- Meldrum, B.S. Glutamate as a neurotransmitter in the brain: Review of physiology and pathology. J. Nutr. 2000, 130, 1007s–1015s. [Google Scholar] [PubMed]
- Niciu, M.J.; Kelmendi, B.; Sanacora, G. Overview of glutamatergic neurotransmission in the nervous system. Pharmacol. Biochem. Behav. 2012, 100, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Sherman, S.M. The function of metabotropic glutamate receptors in thalamus and cortex. Neuroscientist 2014, 20, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Danbolt, N.C. Glutamate as a neurotransmitter in the healthy brain. J. Neural Transm. 2014, 121, 799–817. [Google Scholar] [CrossRef] [PubMed]
- Yelamanchi, S.D.; Jayaram, S.; Thomas, J.K.; Gundimeda, S.; Khan, A.A.; Singhal, A.; Keshava Prasad, T.S.; Pandey, A.; Somani, B.L.; Gowda, H. A pathway map of glutamate metabolism. J. Cell Commun. Signal. 2016, 10, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Fremeau, R.T., Jr.; Voglmaier, S.; Seal, R.P.; Edwards, R.H. Vgluts define subsets of excitatory neurons and suggest novel roles for glutamate. Trends Neurosci. 2004, 27, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Rubio, M.E.; Wenthold, R.J. Differential distribution of intracellular glutamate receptors in dendrites. J. Neurosci. 1999, 19, 5549–5562. [Google Scholar] [PubMed]
- Collingridge, G.L.; Olsen, R.W.; Peters, J.; Spedding, M. A nomenclature for ligand-gated ion channels. Neuropharmacology 2009, 56, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Keramidas, A.; Moorhouse, A.J.; Schofield, P.R.; Barry, P.H. Ligand-gated ion channels: Mechanisms underlying ion selectivity. Prog. Biophys. Mol. Biol. 2004, 86, 161–204. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.J.; Trudell, J.R.; Harris, R.A. Seeking structural specificity: Direct modulation of pentameric ligand-gated ion channels by alcohols and general anesthetics. Pharmacol. Rev. 2014, 66, 396–412. [Google Scholar] [CrossRef] [PubMed]
- Connolly, C.N.; Wafford, K.A. The cys-loop superfamily of ligand-gated ion channels: The impact of receptor structure on function. Biochem. Soc. Trans. 2004, 32, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Kozuska, J.L.; Paulsen, I.M. The cys-loop pentameric ligand-gated ion channel receptors: 50 years on. Can. J. Physiol. Pharmacol. 2012, 90, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, D.M.; Rasmussen, S.G.F.; Kobilka, B.K. The structure and function of G-protein-coupled receptors. Nature 2009, 459, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Trkulja, V.; Salkovic, M.; Lackovic, Z. Signalling promiscuity of the metabotropic neurotransmitter receptors: Are there any functional consequences? Period. Biol. 1998, 100, 157–164. [Google Scholar]
- Fryxell, K.J. The evolutionary divergence of neurotransmitter receptors and second-messenger pathways. J. Mol. Evol. 1995, 41, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Katritch, V.; Cherezov, V.; Stevens, R.C. Structure-function of the G-protein-coupled receptor superfamily. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 531–556. [Google Scholar] [CrossRef] [PubMed]
- Pierce, K.L.; Premont, R.T.; Lefkowitz, R.J. Seven-transmembrane receptors. Nat. Rev. Mol. Cell Biol. 2002, 3, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Strader, C.D.; Fong, T.M.; Tota, M.R.; Underwood, D.; Dixon, R.A.F. Structure and function of G-protein-coupled receptors. Annu. Rev. Biochem. 1994, 63, 101–132. [Google Scholar] [CrossRef] [PubMed]
- Willard, S.S.; Koochekpour, S. Glutamate, glutamate receptors, and downstream signaling pathways. Int. J. Biol. Sci. 2013, 9, 948–959. [Google Scholar] [CrossRef] [PubMed]
- Karakas, E.; Regan, M.C.; Furukawa, H. Emerging structural insights into the function of ionotropic glutamate receptors. Trends Biochem. Sci. 2015, 40, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Harvey, B.H.; Shahid, M. Metabotropic and ionotropic glutamate receptors as neurobiological targets in anxiety and stress-related disorders: Focus on pharmacology and preclinical translational models. Pharmacol. Biochem. Behav. 2012, 100, 775–800. [Google Scholar] [CrossRef] [PubMed]
- Hassel, B.; Dingledine, R. Chapter 17—Glutamate and glutamate receptors. In Basic Neurochemistry, 8th ed.; Brady, S.T., Siegel, G.J., Albers, R.W., Price, D.L., Eds.; Academic Press: New York, NY, USA, 2012; pp. 342–366. [Google Scholar]
- Niswender, C.M.; Conn, P.J. Metabotropic glutamate receptors: Physiology, pharmacology, and disease. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 295–322. [Google Scholar] [CrossRef] [PubMed]
- Page, G.; Khidir, F.A.; Pain, S.; Barrier, L.; Fauconneau, B.; Guillard, O.; Piriou, A.; Hugon, J. Group I metabotropic glutamate receptors activate the p70S6 kinase via both mammalian target of rapamycin (mTOR) and extracellular signal-regulated kinase (ERK 1/2) signaling pathways in rat striatal and hippocampal synaptoneurosomes. Neurochem. Int. 2006, 49, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Saugstad, J.A.; Ingram, S.L. Group I metabotropic glutamate receptors (mGlu1 and mGlu5). In The Glutamate Receptors; Gereau, R.W., Swanson, G.T., Eds.; Humana Press: Totowa, NJ, USA, 2008; pp. 387–463. [Google Scholar]
- Hermans, E.; Challiss, R.A. Structural, signalling and regulatory properties of the group I metabotropic glutamate receptors: Prototypic family C G-protein-coupled receptors. Biochem. J. 2001, 359, 465–484. [Google Scholar] [CrossRef] [PubMed]
- Willard, S.S.; Koochekpour, S. Glutamate signaling in benign and malignant disorders: Current status, future perspectives, and therapeutic implications. Int. J. Biol. Sci. 2013, 9, 728–742. [Google Scholar] [CrossRef] [PubMed]
- Iacovelli, L.; Bruno, V.; Salvatore, L.; Melchiorri, D.; Gradini, R.; Caricasole, A.; Barletta, E.; De Blasi, A.; Nicoletti, F. Native group-III metabotropic glutamate receptors are coupled to the mitogen-activated protein kinase/phosphatidylinositol-3-kinase pathways. J. Neurochem. 2002, 82, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Dhanasiri, A.K.S.; Fernandes, J.M.O.; Kiron, V. Glutamine synthetase activity and the expression of three glul paralogues in zebrafish during transport. Comp. Biochem. Physiol. Part B 2012, 163, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.-H.; Wu, C.-H.; Chen, Y.-C.; Chow, W.-Y. Embryonic expression of zebrafish ampa receptor genes: Zygotic gria2α expression initiates at the midblastula transition. Brain Res. 2006, 1110, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Hoppmann, V.; Wu, J.J.; Søviknes, A.M.; Helvik, J.V.; Becker, T.S. Expression of the eight AMPA receptor subunit genes in the developing central nervous system and sensory organs of zebrafish. Dev. Dyn. 2008, 237, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.A.; Kucenas, S.; Voigt, M.M. Molecular characterization and embryonic expression of the family of N-methyl-D-aspartate receptor subunit genes in the zebrafish. Dev. Dyn. 2005, 234, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Haug, M.F.; Gesemann, M.; Mueller, T.; Neuhauss, S.C.F. Phylogeny and expression divergence of metabotropic glutamate receptor genes in the brain of zebrafish (Danio rerio). J. Comp. Neurol. 2013, 521, 1533–1560. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-Y.; Haug, M.F.; Gesemann, M.; Neuhauss, S.C.F. Novel expression patterns of metabotropic glutamate receptor 6 in the zebrafish nervous system. PLoS ONE 2012, 7, e35256. [Google Scholar] [CrossRef] [PubMed]
- Rico, E.P.; de Oliveira, D.L.; Rosemberg, D.B.; Mussulini, B.H.; Bonan, C.D.; Dias, R.D.; Wofchuk, S.; Souza, D.O.; Bogo, M.R. Expression and functional analysis of Na+-dependent glutamate transporters from zebrafish brain. Brain Res. Bull. 2010, 81, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Gesemann, M.; Lesslauer, A.; Maurer, C.M.; Schönthaler, H.B.; Neuhauss, S.C. Phylogenetic analysis of the vertebrate excitatory/neutral amino acid transporter (SLC1/EAAT) family reveals lineage specific subfamilies. BMC Evolut. Biol. 2010, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Mueller, T.; Wullimann, M.F. Anatomy of neurogenesis in the early zebrafish brain. Dev. Brain Res. 2003, 140, 137–155. [Google Scholar] [CrossRef]
- Schuurmans, C.; Guillemot, F. Molecular mechanisms underlying cell fate specification in the developing telencephalon. Curr. Opin. Neurobiol. 2002, 12, 26–34. [Google Scholar] [CrossRef]
- Liguz-Lecznar, M.; Skangiel-Kramska, J. Vesicular glutamate transporters (VGLUTs): The three musketeers of glutamatergic system. Acta Neurobiol. Exp. 2007, 67, 207–218. [Google Scholar]
- Bezzi, P.; Gundersen, V.; Galbete, J.L.; Seifert, G.; Steinhauser, C.; Pilati, E.; Volterra, A. Astrocytes contain a vesicular compartment that is competent for regulated exocytosis of glutamate. Nat. Neurosci. 2004, 7, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Gallo, V.; Ghiani, C.A. Glutamate receptors in glia: New cells, new inputs and new functions. Trends Pharmacol. Sci. 2000, 21, 252–258. [Google Scholar] [CrossRef]
- Olsen, R.W.; Li, G.-D. Chapter 18—GABA. In Basic Neurochemistry, 8th ed.; Brady, S.T., Siegel, G.J., Albers, R.W., Price, D.L., Eds.; Academic Press: New York, NY, USA, 2012; pp. 367–376. [Google Scholar]
- Pehrson, A.L.; Sanchez, C. Altered γ-aminobutyric acid neurotransmission in major depressive disorder: A critical review of the supporting evidence and the influence of serotonergic antidepressants. Drug Des. Devel. Ther. 2015, 9, 603–624. [Google Scholar] [CrossRef] [PubMed]
- Kantrowitz, J.; Citrome, L.; Javitt, D. GABAB receptors, schizophrenia and sleep dysfunction. CNS Drugs 2009, 23, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, L.J. Molecular mechanisms of antiseizure drug activity at GABAA receptors. Seizure 2013, 22, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ari, Y. Excitatory actions of GABA during development: The nature of the nurture. Nat. Rev. Neurosci. 2002, 3, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.; Brustein, E.; Liao, M.; Mercado, A.; Babilonia, E.; Mount, D.B.; Drapeau, P. Neurogenic role of the depolarizing chloride gradient revealed by global overexpression of KCC2 from the onset of development. J. Neurosci. 2008, 28, 1588–1597. [Google Scholar] [CrossRef] [PubMed]
- Rivera, C.; Voipio, J.; Payne, J.A.; Ruusuvuori, E.; Lahtinen, H.; Lamsa, K.; Pirvola, U.; Saarma, M.; Kaila, K. The K+/Cl− co-transporter KCC2 renders GABA hyperpolarizing during neuronal maturation. Nature 1999, 397, 251–255. [Google Scholar] [PubMed]
- Yamada, J.; Okabe, A.; Toyoda, H.; Kilb, W.; Luhmann, H.J.; Fukuda, A. Cl− uptake promoting depolarizing GABA actions in immature rat neocortical neurones is mediated by NKCC1. J. Physiol. 2004, 557, 829–841. [Google Scholar] [CrossRef] [PubMed]
- Bosma, P.T.; Blázquez, M.; Collins, M.A.; Bishop, J.D.; Drouin, G.; Priede, I.G.; Docherty, K.; Trudeau, V.L. Multiplicity of glutamic acid decarboxylases (GAD) in vertebrates: Molecular phylogeny and evidence for a new GAD paralog. Mol. Biol. Evol. 1999, 16, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Scimemi, A. Structure, function, and plasticity of GABA transporters. Front. Cell. Neurosci. 2014, 8, 161. [Google Scholar] [CrossRef] [PubMed]
- Schousboe, A.; Bak, L.K.; Waagepetersen, H.S. Astrocytic control of biosynthesis and turnover of the neurotransmitters glutamate and GABA. Front. Endocrinol. 2013, 4, 102. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.B.; Waagepetersen, H.S.; Bak, L.K.; Schousboe, A.; Sonnewald, U. The glutamine–glutamate/GABA cycle: Function, regional differences in glutamate and GABA production and effects of interference with GABA metabolism. Neurochem. Res. 2015, 40, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Sigel, E.; Steinmann, M.E. Structure, function, and modulation of GABAA receptors. J. Biol. Chem. 2012, 287, 40224–40231. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.; Wakimoto, H.; Fujita, N.; Lalande, M.; Barnard, E.A. Analysis of the set of GABAA receptor genes in the human genome. J. Biol. Chem. 2004, 279, 41422–41435. [Google Scholar] [CrossRef] [PubMed]
- Bowery, N.G.; Bettler, B.; Froestl, W.; Gallagher, J.P.; Marshall, F.; Raiteri, M.; Bonner, T.I.; Enna, S.J. International union of pharmacology. XXXIII. Mammalian γ-aminobutyric acidB receptors: Structure and function. Pharmacol. Rev. 2002, 54, 247–264. [Google Scholar] [CrossRef] [PubMed]
- Pinard, A.; Seddik, R.; Bettler, B. GABAB receptors: Physiological functions and mechanisms of diversity. Adv. Pharmacol. (San Diego Calif.) 2010, 58, 231–255. [Google Scholar]
- Padgett, C.L.; Slesinger, P.A. GABAB receptor coupling to G-proteins and ion channels. In Advances in Pharmacology; Thomas, P.B., Ed.; Academic Press: San Diego, CA, USA, 2010; Volume 58, pp. 123–147. [Google Scholar]
- Kullmann, D.M.; Ruiz, A.; Rusakov, D.M.; Scott, R.; Semyanov, A.; Walker, M.C. Presynaptic, extrasynaptic and axonal GABAA receptors in the CNS: Where and why? Prog. Biophys. Mol. Biol. 2005, 87, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Bettler, B.; Tiao, J.Y.-H. Molecular diversity, trafficking and subcellular localization of GABAB receptors. Pharmacol. Ther. 2006, 110, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.C.; Heinrich, G.; Sandell, J.H. Sequence and expression of glutamic acid decarboxylase isoforms in the developing zebrafish. J. Comp. Neurol. 1998, 396, 253–266. [Google Scholar] [CrossRef]
- Mueller, T.; Vernier, P.; Wullimann, M.F. A phylotypic stage in vertebrate brain development: GABA cell patterns in zebrafish compared with mouse. J. Comp. Neurol. 2006, 494, 620–634. [Google Scholar] [CrossRef] [PubMed]
- Mueller, T.; Wullimann, M.F. Chapter 3—Interpretation of data—How to use the atlas: Analysis. In Atlas of Early Zebrafish Brain Development, 2nd ed.; Elsevier: San Diego, CA, USA, 2016; pp. 159–204. [Google Scholar]
- Higashijima, S.-I.; Schaefer, M.; Fetcho, J.R. Neurotransmitter properties of spinal interneurons in embryonic and larval zebrafish. J. Comp. Neurol. 2004, 480, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Nam, R.H.; Yoo, Y.M.; Lee, C.J. Identification and functional evidence of GABAergic neurons in parts of the brain of adult zebrafish (Danio rerio). Neurosci. Lett. 2004, 355, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Mueller, T.; Guo, S. The distribution of GAD67-mRNA in the adult zebrafish (teleost) forebrain reveals a prosomeric pattern and suggests previously unidentified homologies to tetrapods. J. Comp. Neurol. 2009, 516, 553–568. [Google Scholar] [CrossRef] [PubMed]
- Delgado, L.; Schmachtenberg, O. Immunohistochemical localization of GABA, GAD65, and the receptor subunits GABAAα1 and GABAB1 in the zebrafish cerebellum. Cerebellum 2008, 7, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Goldman-Rakic, P.S. The cortical dopamine system: Role in memory and cognition. Adv. Pharmacol. (San Diego Calif.) 1998, 42, 707–711. [Google Scholar]
- Schultz, W. Updating dopamine reward signals. Curr. Opin. Neurobiol. 2013, 23, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Howes, O.D.; Kambeitz, J.; Kim, E.; Stahl, D.; Slifstein, M.; Abi-Dargham, A.; Kapur, S. The nature of dopamine dysfunction in schizophrenia and what this means for treatment: Meta-analysis of imaging studies. Arch. Gen. Psychiatry 2012, 69, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Daubner, S.C.; Le, T.; Wang, S. Tyrosine hydroxylase and regulation of dopamine synthesis. Arch. Biochem. Biophys. 2011, 508, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Neff, N.H.; Hadjiconstantinou, M. Regulation of tyrosine hydroxylase and aromatic l-amino acid decarboxylase by dopaminergic drugs. Eur. J. Pharmacol. 1997, 323, 149–157. [Google Scholar] [CrossRef]
- Gnegy, M.E. Chapter 14—Catecholamines. In Basic Neurochemistry, 8th ed.; Brady, S.T., Siegel, G.J., Albers, R.W., Price, D.L., Eds.; Academic Press: New York, NY, USA, 2012; pp. 283–299. [Google Scholar]
- Benarroch, E.E. Monoamine transporters: Structure, regulation, and clinical implications. Neurology 2013, 81, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Lawal, H.O.; Krantz, D.E. Slc18: Vesicular neurotransmitter transporters for monoamines and acetylcholine. Mol. Asp. Med. 2013, 34, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Rudnick, G.; Kramer, R.; Blakely, R.D.; Murphy, D.L.; Verrey, F. The SLC6 transporters: Perspectives on structure, functions, regulation, and models for transporter dysfunction. Pflüg. Arch.—Eur. J. Physiol. 2014, 466, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Shih, J.C.; Chen, K.; Ridd, M.J. Role of MAO A and B in neurotransmitter metabolism and behavior. Pol. J. Pharmacol. 1999, 51, 25–29. [Google Scholar] [PubMed]
- Callier, S.; Snapyan, M.; Le Crom, S.; Prou, D.; Vincent, J.D.; Vernier, P. Evolution and cell biology of dopamine receptors in vertebrates. Biol. Cell/Under Auspices Eur. Cell Biol. Organ. 2003, 95, 489–502. [Google Scholar] [CrossRef]
- Missale, C.; Nash, S.R.; Robinson, S.W.; Jaber, M.; Caron, M.G. Dopamine receptors: From structure to function. Physiol. Rev. 1998, 78, 189–225. [Google Scholar] [PubMed]
- Beaulieu, J.-M.; Espinoza, S.; Gainetdinov, R.R. Dopamine receptors—IUPHAR review 13. Br. J. Pharmacol. 2015, 172, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Candy, J.; Collet, C. Two tyrosine hydroxylase genes in teleosts. Biochim. Biophys. Acta 2005, 1727, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Filippi, A.; Mahler, J.; Schweitzer, J.; Driever, W. Expression of the paralogous tyrosine hydroxylase encoding genes th1 and th2 reveals the full complement of dopaminergic and noradrenergic neurons in zebrafish larval and juvenile brain. J. Comp. Neurol. 2010, 518, 423–438. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Ruuskanen, J.O.; Wullimann, M.F.; Vernier, P. Two tyrosine hydroxylase genes in vertebrates new dopaminergic territories revealed in the zebrafish brain. Mol. Cell. Neurosci. 2010, 43, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Ruuskanen, J.O.; Wullimann, M.F.; Vernier, P. Differential expression of dopaminergic cell markers in the adult zebrafish forebrain. J. Comp. Neurol. 2011, 519, 576–598. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Li, S.; Zhong, H.; Lin, S. Zebrafish tyrosine hydroxylase 2 gene encodes tryptophan hydroxylase. J. Biol. Chem. 2013, 288, 22451–22459. [Google Scholar] [CrossRef] [PubMed]
- Setini, A.; Pierucci, F.; Senatori, O.; Nicotra, A. Molecular characterization of monoamine oxidase in zebrafish (Danio rerio). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2005, 140, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Anichtchik, O.; Sallinen, V.; Peitsaro, N.; Panula, P. Distinct structure and activity of monoamine oxidase in the brain of zebrafish (Danio rerio). J. Comp. Neurol. 2006, 498, 593–610. [Google Scholar] [CrossRef] [PubMed]
- Sallinen, V.; Sundvik, M.; Reenilä, I.; Peitsaro, N.; Khrustalyov, D.; Anichtchik, O.; Toleikyte, G.; Kaslin, J.; Panula, P. Hyperserotonergic phenotype after monoamine oxidase inhibition in larval zebrafish. J. Neurochem. 2009, 109, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Alazizi, A.; Liu, M.-Y.; Williams, F.E.; Kurogi, K.; Sakakibara, Y.; Suiko, M.; Liu, M.-C. Identification, characterization, and ontogenic study of a catechol o-methyltransferase from zebrafish. Aquat. Toxicol. 2011, 102, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Sprague, J.; Bayraktaroglu, L.; Clements, D.; Conlin, T.; Fashena, D.; Frazer, K.; Haendel, M.; Howe, D.G.; Mani, P.; Ramachandran, S.; et al. The zebrafish information network: The zebrafish model organism database. Nucleic Acids Res. 2006, 34, D581–D585. [Google Scholar] [CrossRef] [PubMed]
- Ruuskanen, J.O.; Laurila, J.; Xhaard, H.; Rantanen, V.V.; Vuoriluoto, K.; Wurster, S.; Marjamaki, A.; Vainio, M.; Johnson, M.S.; Scheinin, M. Conserved structural, pharmacological and functional properties among the three human and five zebrafish alpha 2-adrenoceptors. Br. J. Pharmacol. 2005, 144, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, J.; Löhr, H.; Filippi, A.; Driever, W. Dopaminergic and noradrenergic circuit development in zebrafish. Dev. Neurobiol. 2012, 72, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Rink, E.; Wullimann, M.F. Development of the catecholaminergic system in the early zebrafish brain: An immunohistochemical study. Dev. Brain Res. 2002, 137, 89–100. [Google Scholar] [CrossRef]
- Rink, E.; Wullimann, M.F. The teleostean (zebrafish) dopaminergic system ascending to the subpallium (striatum) is located in the basal diencephalon (posterior tuberculum). Brain Res. 2001, 889, 316–330. [Google Scholar] [CrossRef]
- Becker, T.; Wullimann, M.F.; Becker, C.G.; Bernhardt, R.R.; Schachner, M. Axonal regrowth after spinal cord transection in adult zebrafish. J. Comp. Neurol. 1997, 377, 577–595. [Google Scholar] [CrossRef]
- Parker, M.O.; Brock, A.J.; Walton, R.T.; Brennan, C.H. The role of zebrafish (Danio rerio) in dissecting the genetics and neural circuits of executive function. Front. Neural Circuits 2013, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Panula, P.; Sundvik, M.; Karlstedt, K. Developmental roles of brain histamine. Trends Neurosci. 2014, 37, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Barreiro-Iglesias, A.; Mysiak, K.S.; Adrio, F.; Rodicio, M.C.; Becker, C.G.; Becker, T.; Anadon, R. Distribution of glycinergic neurons in the brain of glycine transporter-2 transgenic Tg(glyt2:Gfp) adult zebrafish: Relationship to brain-spinal descending systems. J. Comp. Neurol. 2013, 521, 389–425. [Google Scholar] [CrossRef] [PubMed]
- Haas, H.L.; Sergeeva, O.A.; Selbach, O. Histamine in the nervous system. Physiol. Rev. 2008, 88, 1183–1241. [Google Scholar] [CrossRef] [PubMed]
- Tomkins, D.M.; Sellers, E.M. Addiction and the brain: The role of neurotransmitters in the cause and treatment of drug dependence. Can. Med. Assoc. J. 2001, 164, 817–821. [Google Scholar]
- Dalley, J.W.; Roiser, J.P. Dopamine, serotonin and impulsivity. Neuroscience 2012, 215, 42–58. [Google Scholar] [CrossRef] [PubMed]
- Perry, E.; Walker, M.; Grace, J.; Perry, R. Acetylcholine in mind: A neurotransmitter correlate of consciousness? Trends Neurosci. 1999, 22, 273–280. [Google Scholar] [CrossRef]
- Filippi, A.; Mueller, T.; Driever, W. Vglut2 and gad expression reveal distinct patterns of dual GABAergic versus glutamatergic cotransmitter phenotypes of dopaminergic and noradrenergic neurons in the zebrafish brain. J. Comp. Neurol. 2014, 522, 2019–2037. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.M. Catecholaminergic systems in the zebrafish. III. Organization and projection pattern of medullary dopaminergic and noradrenergic neurons. J. Comp. Neurol. 1997, 381, 411–427. [Google Scholar] [CrossRef]
- McLean, D.L.; Fetcho, J.R. Ontogeny and innervation patterns of dopaminergic, noradrenergic, and serotonergic neurons in larval zebrafish. J. Comp. Neurol. 2004, 480, 38–56. [Google Scholar] [CrossRef] [PubMed]
- Kaslin, J.; Panula, P. Comparative anatomy of the histaminergic and other aminergic systems in zebrafish (Danio rerio). J. Comp. Neurol. 2001, 440, 342–377. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.M. Catecholaminergic systems in the zebrafish. II. Projection pathways and pattern of termination of the locus coeruleus. J. Comp. Neurol. 1994, 344, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Daubert, E.A.; Condron, B.G. Serotonin: A regulator of neuronal morphology and circuitry. Trends Neurosci. 2010, 33, 424–434. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, P.; Cases, O.; Maroteaux, L. The developmental role of serotonin: News from mouse molecular genetics. Nat. Rev. Neurosci. 2003, 4, 1002–1012. [Google Scholar] [CrossRef] [PubMed]
- Lucki, I. The spectrum of behaviors influenced by serotonin. Biol. Psychiatry 1998, 44, 151–162. [Google Scholar] [CrossRef]
- Parsey, R.V. Serotonin receptor imaging: Clinically useful? J. Nucl. Med. 2010, 51, 1495–1498. [Google Scholar] [CrossRef] [PubMed]
- Walther, D.J.; Bader, M. A unique central tryptophan hydroxylase isoform. Biochem. Pharmacol. 2003, 66, 1673–1680. [Google Scholar] [CrossRef]
- Mohammad-Zadeh, L.F.; Moses, L.; Gwaltney-Brant, S.M. Serotonin: A review. J. Vet. Pharmacol. Ther. 2008, 31, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Hensler, J.G. Chapter 15—Serotonin. In Basic Neurochemistry, 8th ed.; Brady, S.T., Siegel, G.J., Albers, R.W., Price, D.L., Eds.; Academic Press: New York, NY, USA, 2012; pp. 300–322. [Google Scholar]
- Nichols, D.E.; Nichols, C.D. Serotonin receptors. Chem. Rev. 2008, 108, 1614–1641. [Google Scholar] [CrossRef] [PubMed]
- Bellipanni, G.; Rink, E.; Bally-Cuif, L. Cloning of two tryptophan hydroxylase genes expressed in the diencephalon of the developing zebrafish brain. Mech. Dev. 2002, 119 (Suppl.), S215–S220. [Google Scholar] [CrossRef]
- Teraoka, H.; Russell, C.; Regan, J.; Chandrasekhar, A.; Concha, M.L.; Yokoyama, R.; Higashi, K.; Take-uchi, M.; Dong, W.; Hiraga, T.; et al. Hedgehog and fgf signaling pathways regulate the development of tphR-expressing serotonergic raphe neurons in zebrafish embryos. J. Neurobiol. 2004, 60, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Lillesaar, C. The serotonergic system in fish. J. Chem. Neuroanat. 2011, 41, 294–308. [Google Scholar] [CrossRef] [PubMed]
- Herculano, A.M.; Maximino, C. Serotonergic modulation of zebrafish behavior: Towards a paradox. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 55, 50–66. [Google Scholar] [CrossRef] [PubMed]
- Lillesaar, C.; Tannhauser, B.; Stigloher, C.; Kremmer, E.; Bally-Cuif, L. The serotonergic phenotype is acquired by converging genetic mechanisms within the zebrafish central nervous system. Dev. Dyn. 2007, 236, 1072–1084. [Google Scholar] [CrossRef] [PubMed]
- Norton, W.H.J.; Folchert, A.; Bally-Cuif, L. Comparative analysis of serotonin receptor (HTR1A/HTR1B families) and transporter (slc6a4a/b) gene expression in the zebrafish brain. J. Comp. Neurol. 2008, 511, 521–542. [Google Scholar] [CrossRef] [PubMed]
- Picciotto, M.R.; Higley, M.J.; Mineur, Y.S. Acetylcholine as a neuromodulator: Cholinergic signaling shapes nervous system function and behavior. Neuron 2012, 76, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Alkondon, M.; Rocha, E.S.; Maelicke, A.; Albuquerque, E.X. Diversity of nicotinic acetylcholine receptors in rat brain. V. Alpha-bungarotoxin-sensitive nicotinic receptors in olfactory bulb neurons and presynaptic modulation of glutamate release. J. Pharmacol. Exp. Ther. 1996, 278, 1460–1471. [Google Scholar] [PubMed]
- Kenney, J.W.; Florian, C.; Portugal, G.S.; Abel, T.; Gould, T.J. Involvement of hippocampal jun-n terminal kinase pathway in the enhancement of learning and memory by nicotine. Neuropsychopharmacology 2010, 35, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Fisher, S.K.; Wonnacott, S. Chapter 13—Acetylcholine. In Basic Neurochemistry, 8th ed.; Brady, S.T., Siegel, G.J., Albers, R.W., Price, D.L., Eds.; Academic Press: New York, NY, USA, 2012; pp. 258–282. [Google Scholar]
- Yakel, J.L. Cholinergic receptors: Functional role of nicotinic ach receptors in brain circuits and disease. Pflüg. Arch. Eur. J. Physiol. 2013, 465, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.A. Muscarinic acetylcholine receptors (mAChRs) in the nervous system: Some functions and mechanisms. J. Mol. Neurosci. 2010, 41, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Zirger, J.M.; Beattie, C.E.; McKay, D.B.; Thomas Boyd, R. Cloning and expression of zebrafish neuronal nicotinic acetylcholine receptors. Gene Expr. Patterns 2003, 3, 747–754. [Google Scholar] [CrossRef]
- Papke, R.L.; Ono, F.; Stokes, C.; Urban, J.M.; Boyd, R.T. The nicotinic acetylcholine receptors of zebrafish and an evaluation of pharmacological tools used for their study. Biochem. Pharmacol. 2012, 84, 352–365. [Google Scholar] [CrossRef] [PubMed]
- Williams, F.E.; Messer, W.S., Jr. Muscarinic acetylcholine receptors in the brain of the zebrafish (Danio rerio) measured by radioligand binding techniques. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2004, 137, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Kaslin, J.; Nystedt, J.M.; Ostergard, M.; Peitsaro, N.; Panula, P. The orexin/hypocretin system in zebrafish is connected to the aminergic and cholinergic systems. J. Neurosci. 2004, 24, 2678–2689. [Google Scholar] [CrossRef] [PubMed]
- Clemente, D.; Porteros, A.; Weruaga, E.; Alonso, J.R.; Arenzana, F.J.; Aijon, J.; Arevalo, R. Cholinergic elements in the zebrafish central nervous system: Histochemical and immunohistochemical analysis. J. Comp. Neurol. 2004, 474, 75–107. [Google Scholar] [CrossRef] [PubMed]
- Mueller, T.; Vernier, P.; Wullimann, M.F. The adult central nervous cholinergic system of a neurogenetic model animal, the zebrafish Danio rerio. Brain Res. 2004, 1011, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Arenzana, F.J.; Clemente, D.; Sánchez-González, R.; Porteros, Á.; Aijón, J.; Arévalo, R. Development of the cholinergic system in the brain and retina of the zebrafish. Brain Res. Bull. 2005, 66, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Haas, H.; Panula, P. The role of histamine and the tuberomamillary nucleus in the nervous system. Nat. Rev. Neurosci. 2003, 4, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Sundvik, M.; Kudo, H.; Toivonen, P.; Rozov, S.; Chen, Y.C.; Panula, P. The histaminergic system regulates wakefulness and orexin/hypocretin neuron development via histamine receptor H1 in zebrafish. FASEB J. 2011, 25, 4338–4347. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.C.; Arrang, J.M.; Garbarg, M.; Pollard, H.; Ruat, M. Histaminergic transmission in the mammalian brain. Physiol. Rev. 1991, 71, 1–51. [Google Scholar] [PubMed]
- Leurs, R.; Hough, L.B.; Blandina, P.; Haas, H.L. Chapter 16—Histamine. In Basic Neurochemistry, 8th ed.; Brady, S.T., Siegel, G.J., Albers, R.W., Price, D.L., Eds.; Academic Press: New York, NY, USA, 2012; pp. 323–341. [Google Scholar]
- Eriksson, K.S.; Peitsaro, N.; Karlstedt, K.; Kaslin, J.; Panula, P. Development of the histaminergic neurons and expression of histidine decarboxylase mRNA in the zebrafish brain in the absence of all peripheral histaminergic systems. Eur. J. Neurosci. 1998, 10, 3799–3812. [Google Scholar] [CrossRef] [PubMed]
- Onodera, K.; Yamatodani, A.; Watanabe, T.; Wadas, H. Neuropharmacology of the histaminergic neuron system in the brain and its relationship with behavioral disorders. Prog. Neurobiol. 1994, 42, 685–702. [Google Scholar] [CrossRef]
- Kukko-Lukjanov, T.K.; Panula, P. Subcellular distribution of histamine, GABA and galanin in tuberomamillary neurons in vitro. J. Chem. Neuroanat. 2003, 25, 279–292. [Google Scholar] [CrossRef]
- Connelly, W.M.; Shenton, F.C.; Lethbridge, N.; Leurs, R.; Waldvogel, H.J.; Faull, R.L.; Lees, G.; Chazot, P.L. The histamine h4 receptor is functionally expressed on neurons in the mammalian CNS. Br. J. Pharmacol. 2009, 157, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Drutel, G.; Peitsaro, N.; Karlstedt, K.; Wieland, K.; Smit, M.J.; Timmerman, H.; Panula, P.; Leurs, R. Identification of rat H3 receptor isoforms with different brain expression and signaling properties. Mol. Pharmacol. 2001, 59, 1–8. [Google Scholar] [PubMed]
- Giovannini, M.G.; Efoudebe, M.; Passani, M.B.; Baldi, E.; Bucherelli, C.; Giachi, F.; Corradetti, R.; Blandina, P. Improvement in fear memory by histamine-elicited ERK2 activation in hippocampal CA3 cells. J. Neurosci. 2003, 23, 9016–9023. [Google Scholar] [PubMed]
- Mariottini, C.; Scartabelli, T.; Bongers, G.; Arrigucci, S.; Nosi, D.; Leurs, R.; Chiarugi, A.; Blandina, P.; Pellegrini-Giampietro, D.E.; Passani, M.B. Activation of the histaminergic H3 receptor induces phosphorylation of the Akt/GSK-3β pathway in cultured cortical neurons and protects against neurotoxic insults. J. Neurochem. 2009, 110, 1469–1478. [Google Scholar] [CrossRef] [PubMed]
- Peitsaro, N.; Sundvik, M.; Anichtchik, O.V.; Kaslin, J.; Panula, P. Identification of zebrafish histamine H1, H2 and H3 receptors and effects of histaminergic ligands on behavior. Biochem. Pharmacol. 2007, 73, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Sundvik, M.; Panula, P. Organization of the histaminergic system in adult zebrafish (Danio rerio) brain: Neuron number, location, and cotransmitters. J. Comp. Neurol. 2012, 520, 3827–3845. [Google Scholar] [CrossRef] [PubMed]
- McDearmid, J.R.; Liao, M.; Drapeau, P. Glycine receptors regulate interneuron differentiation during spinal network development. Proc. Natl. Acad. Sci. USA 2006, 103, 9679–9684. [Google Scholar] [CrossRef] [PubMed]
- Hernandes, M.S.; Troncone, L.R. Glycine as a neurotransmitter in the forebrain: A short review. J. Neural Trans. (Vienna Austria: 1996) 2009, 116, 1551–1560. [Google Scholar] [CrossRef] [PubMed]
- Raiteri, L.; Stigliani, S.; Usai, C.; Diaspro, A.; Paluzzi, S.; Milanese, M.; Raiteri, M.; Bonanno, G. Functional expression of release-regulating glycine transporters GLYT1 on GABAergic neurons and GLY2 on astrocytes in mouse spinal cord. Neurochem. Int. 2008, 52, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Bowery, N.G.; Smart, T.G. GABA and glycine as neurotransmitters: A brief history. Br. J. Pharmacol. 2006, 147, S109–S119. [Google Scholar] [CrossRef] [PubMed]
- Cummings, K.A.; Popescu, G.K. Glycine-dependent activation of nmda receptors. J. Gen. Physiol. 2015, 145, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Yevenes, G.E.; Peoples, R.W.; Tapia, J.C.; Parodi, J.; Soto, X.; Olate, J.; Aguayo, L.G. Modulation of glycine-activated ion channel function by G-protein βγ subunits. Nat. Neurosci. 2003, 6, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Fucile, S.; de Saint Jan, D.; David-Watine, B.; Korn, H.; Bregestovski, P. Comparison of glycine and GABA actions on the zebrafish homomeric glycine receptor. J. Physiol. 1999, 517 Pt 2, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Imboden, M.; Devignot, V.; Goblet, C. Phylogenetic relationships and chromosomal location of five distinct glycine receptor subunit genes in the teleost Danio rerio. Dev. Genes Evol. 2001, 211, 415–422. [Google Scholar] [PubMed]
- Higashijima, S.; Mandel, G.; Fetcho, J.R. Distribution of prospective glutamatergic, glycinergic, and GABAergic neurons in embryonic and larval zebrafish. J. Comp. Neurol. 2004, 480, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Moly, P.K.; Ikenaga, T.; Kamihagi, C.; Islam, A.F.M.T.; Hatta, K. Identification of initially appearing glycine-immunoreactive neurons in the embryonic zebrafish brain. Dev. Neurobiol. 2014, 74, 616–632. [Google Scholar] [CrossRef] [PubMed]
- Joca, S.R.; Moreira, F.A.; Wegener, G. Atypical neurotransmitters and the neurobiology of depression. CNS Neurol. Disord.-Drug Targets 2015, 14, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Holmqvist, B.; Ebbesson, L.; Alm, P. Nitric oxide and the zebrafish (Danio rerio): Developmental neurobiology and brain neurogenesis. In Advances in Experimental Biology; Bruno, T., Barry, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 1, pp. 229–465. [Google Scholar]
- Chatterjee, D.; Gerlai, R. High precision liquid chromatography analysis of dopaminergic and serotoninergic responses to acute alcohol exposure in zebrafish. Behav. Brain Res. 2009, 200, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Milanese, C.; Sager, J.J.; Bai, Q.; Farrell, T.C.; Cannon, J.R.; Greenamyre, J.T.; Burton, E.A. Hypokinesia and reduced dopamine levels in zebrafish lacking β- and γ1-synucleins. J. Biol. Chem. 2012, 287, 2971–2983. [Google Scholar] [CrossRef] [PubMed]
- Wirbisky, S.E.; Weber, G.J.; Lee, J.W.; Cannon, J.R.; Freeman, J.L. Novel dose-dependent alterations in excitatory GABA during embryonic development associated with lead (Pb) neurotoxicity. Toxicol. Lett. 2014, 229, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Chatterjee, D.; Gerlai, R. Strain dependent gene expression and neurochemical levels in the brain of zebrafish: Focus on a few alcohol related targets. Physiol. Behav. 2012, 107, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Tufi, S.; Leonards, P.; Lamoree, M.; de Boer, J.; Legler, J.; Legradi, J. Changes in neurotransmitter profiles during early zebrafish (Danio rerio) development and after pesticide exposure. Environ. Sci. Technol. 2016, 50, 3222–3230. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.J.; McCutcheon, J.E.; Young, A.M.J.; Norton, W.H.J. Neurochemical measurements in the zebrafish brain. Front. Behav. Neurosci. 2015, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Lopez Patino, M.A.; Yu, L.; Yamamoto, B.K.; Zhdanova, I.V. Gender differences in zebrafish responses to cocaine withdrawal. Physiol. Behav. 2008, 95, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Cambier, S.; Gonzalez, P.; Mesmer-Dudons, N.; Brèthes, D.; Fujimura, M.; Bourdineaud, J.-P. Effects of dietary methylmercury on the zebrafish brain: Histological, mitochondrial, and gene transcription analyses. BioMetals 2012, 25, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Wirbisky, S.E.; Weber, G.J.; Sepulveda, M.S.; Xiao, C.; Cannon, J.R.; Freeman, J.L. Developmental origins of neurotransmitter and transcriptome alterations in adult female zebrafish exposed to atrazine during embryogenesis. Toxicology 2015, 333, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.L.; Weber, G.J.; Peterson, S.M.; Nie, L.H. Embryonic ionizing radiation exposure results in expression alterations of genes associated with cardiovascular and neurological development, function, and disease and modified cardiovascular function in zebrafish. Front. Genet. 2014, 5, 268. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.M.; Zhang, J.; Weber, G.; Freeman, J.L. Global gene expression analysis reveals dynamic and developmental stage-dependent enrichment of lead-induced neurological gene alterations. Environ. Health Perspect. 2011, 119, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.J.; Sepulveda, M.S.; Peterson, S.M.; Lewis, S.S.; Freeman, J.L. Transcriptome alterations following developmental atrazine exposure in zebrafish are associated with disruption of neuroendocrine and reproductive system function, cell cycle, and carcinogenesis. Toxicol. Sci. 2013, 132, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Scholz, S. Zebrafish embryos as an alternative model for screening of drug-induced organ toxicity. Arch. Toxicol. 2013, 87, 767–769. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Aanes, H.; Winata, C.L.; Lin, C.H.; Chen, J.P.; Srinivasan, K.G.; Lee, S.G.P.; Lim, A.Y.M.; Hajan, H.S.; Collas, P.; Bourque, G.; et al. Zebrafish mRNA sequencing deciphers novelties in transcriptome dynamics during maternal to zygotic transition. Genome Res. 2011, 21, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.; Richardson, M.K.; Noldus, L.P.J.J.; Tegelenbosch, R.A.J. Zebrafish embryos and larvae in behavioural assays. Behaviour 2012, 149, 1241–1281. [Google Scholar] [CrossRef]
- Bailey, J.; Oliveri, A.; Levin, E.D. Zebrafish model systems for developmental neurobehavioral toxicology. Birth Defects Res. Part C Embryo Today 2013, 99, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Blaser, R.E.; Chadwick, L.; McGinnis, G.C. Behavioral measures of anxiety in zebrafish (Danio rerio). Behav. Brain Res. 2010, 208, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Budick, S.A.; O’Malley, D.M. Locomotor repertoire of the larval zebrafish: Swimming, turning and prey capture. J. Exp. Biol. 2000, 203, 2565–2579. [Google Scholar] [PubMed]
- Buske, C.; Gerlai, R. Diving deeper into zebrafish development of social behavior: Analyzing high resolution data. J. Neurosci. Methods 2014, 234, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Champagne, D.L.; Hoefnagels, C.C.M.; de Kloet, R.E.; Richardson, M.K. Translating rodent behavioral repertoire to zebrafish (Danio rerio): Relevance for stress research. Behav. Brain Res. 2010, 214, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Egan, R.J.; Bergner, C.L.; Hart, P.C.; Cachat, J.M.; Canavello, P.R.; Elegante, M.F.; Elkhayat, S.I.; Bartels, B.K.; Tien, A.K.; Tien, D.H.; et al. Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. Behav. Brain Res. 2009, 205, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Gerlai, R. Fish in behavior research: Unique tools with a great promise! J. Neurosci. Methods 2014, 234, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S.; et al. Towards a comprehensive catalog of zebrafish behavior 1.0 and beyond. Zebrafish 2013, 10, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Stewart, A.M.; Gerlai, R. Zebrafish as an emerging model for studying complex brain disorders. Trends Pharmacol. Sci. 2014, 35, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Maximino, C.; de Brito, T.M.; da Silva Batista, A.W.; Herculano, A.M.; Morato, S.; Gouveia, A., Jr. Measuring anxiety in zebrafish: A critical review. Behav. Brain Res. 2010, 214, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.; Stewart, A.M.; Kalueff, A.V. Aquatic blues: Modeling depression and antidepressant action in zebrafish. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 55, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Steenbergen, P.J.; Richardson, M.K.; Champagne, D.L. The use of the zebrafish model in stress research. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 1432–1451. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Echevarria, D.J.; Homechaudhuri, S.; Stewart, A.M.; Collier, A.D.; Kaluyeva, A.A.; Li, S.; Liu, Y.; Chen, P.; Wang, J.; et al. Zebrafish neurobehavioral phenomics for aquatic neuropharmacology and toxicology research. Aquat. Toxicol. 2016, 170, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.O. Adult vertebrate behavioural aquatic toxicology: Reliability and validity. Aquat. Toxicol. 2016, 170, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Connaughton, V.P.; Behar, T.N.; Liu, W.L.S.; Massey, S.C. Immunocytochemical localization of excitatory and inhibitory neurotransmitters in the zebrafish retina. Vis. Neurosci. 1999, 16, 483–490. [Google Scholar] [CrossRef] [PubMed]
- DeCarvalho, T.N.; Subedi, A.; Rock, J.; Harfe, B.D.; Thisse, C.; Thisse, B.; Halpern, M.E.; Hong, E. Neurotransmitter map of the asymmetric dorsal habenular nuclei of zebrafish. Genesis 2014, 52, 636–655. [Google Scholar] [CrossRef] [PubMed]
- Fetcho, J.R.; O’Malley, D.M. Imaging neuronal networks in behaving animals. Curr. Opin. Neurobiol. 1997, 7, 832–838. [Google Scholar] [CrossRef]
- Marvin, J.S.; Borghuis, B.G.; Tian, L.; Cichon, J.; Harnett, M.T.; Akerboom, J.; Gordus, A.; Renninger, S.L.; Chen, T.W.; Bargmann, C.I.; et al. An optimized fluorescent probe for visualizing glutamate neurotransmission. Nat. Methods 2013, 10, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, M.B.; Orger, M.B.; Robson, D.N.; Li, J.M.; Keller, P.J. Whole-brain functional imaging at cellular resolution using light-sheet microscopy. Nat. Methods 2013, 10, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Garaschuk, O.; Milos, R.-I.; Konnerth, A. Targeted bulk-loading of fluorescent indicators for two-photon brain imaging in vivo. Nat. Protoc. 2006, 1, 380–386. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, D.M.; Zhou, Q.; Gahtan, E. Probing neural circuits in the zebrafish: A suite of optical techniques. Methods 2003, 30, 49–63. [Google Scholar] [CrossRef]
- Arrenberg, A.B.; Driever, W. Integrating anatomy and function for zebrafish circuit analysis. Front. Neural Circuits 2013, 7. [Google Scholar] [CrossRef] [PubMed]
- Bruni, G.; Lakhani, P.; Kokel, D. Discovering novel neuroactive drugs through high-throughput behavior-based chemical screening in the zebrafish. Front. Pharmacol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Magno, L.D.P.; Fontes, A.; Goncalves, B.M.N.; Gouveia, A. Pharmacological study of the light/dark preference test in zebrafish (Danio rerio): Waterborne administration. Pharmacol. Biochem. Behav. 2015, 135, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Maximino, C.; da Silva, A.W.B.; Gouveia, A., Jr.; Herculano, A.M. Pharmacological analysis of zebrafish (Danio rerio) scototaxis. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 624–631. [Google Scholar] [CrossRef] [PubMed]
- McCarroll, M.N.; Gendelev, L.; Keiser, M.J.; Kokel, D. Leveraging large-scale behavioral profiling in zebrafish to explore neuroactive polypharmacology. ACS Chem. Biol. 2016, 11, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Rihel, J.; Schier, A.F. Behavioral screening for neuroactive drugs in zebrafish. Dev. Neurobiol. 2012, 72, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.M.; Grieco, F.; Tegelenbosch, R.A.J.; Kyzar, E.J.; Nguyen, M.; Kaluyeva, A.; Song, C.; Noldus, L.; Kalueff, A.V. A novel 3D method of locomotor analysis in adult zebrafish: Implications for automated detection of CNS drug-evoked phenotypes. J. Neurosci. Methods 2015, 255, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Maximino, C.; Herculano, A.M. A review of monoaminergic neuropsychopharmacology in zebrafish. Zebrafish 2010, 7, 359–378. [Google Scholar] [CrossRef] [PubMed]
- Rico, E.P.; Rosemberg, D.B.; Dias, R.D.; Bogo, M.R.; Bonan, C.D. Ethanol alters acetylcholinesterase activity and gene expression in zebrafish brain. Toxicol. Lett. 2007, 174, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, D.; Shams, S.; Gerlai, R. Chronic and acute alcohol administration induced neurochemical changes in the brain: Comparison of distinct zebrafish populations. Amino Acids 2014, 46, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Puttonen, H.; Sundvik, M.; Rozov, S.; Chen, Y.-C.; Panula, P. Acute ethanol treatment upregulates th1, th2 and hdc in larval zebrafish in stable networks. Front. Neural Circuits 2013, 7. [Google Scholar] [CrossRef] [PubMed]
- Gerlai, R.; Ahmad, F.; Prajapati, S. Differences in acute alcohol-induced behavioral responses among zebrafish populations. Alcohol. Clin. Exp. Res. 2008, 32, 1763–1773. [Google Scholar] [CrossRef] [PubMed]
- Gerlai, R.; Lee, V.; Blaser, R. Effects of acute and chronic ethanol exposure on the behavior of adult zebrafish (Danio rerio). Pharmacol. Biochem. Behav. 2006, 85, 752–761. [Google Scholar] [CrossRef] [PubMed]
- Tran, S.; Gerlai, R. Time-course of behavioural changes induced by ethanol in zebrafish (Danio rerio). Behav. Brain Res. 2013, 252, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Kurta, A.; Palestis, B.G. Effects of ethanol on the shoaling behavior of zebrafish (Danio rerio). Dose-Response 2010, 8, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Pannia, E.; Tran, S.; Rampersad, M.; Gerlai, R. Acute ethanol exposure induces behavioural differences in two zebrafish (Danio rerio) strains: A time course analysis. Behav. Brain Res. 2014, 259, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.M.; Oliveri, A.N.; Zhang, C.; Frazier, J.M.; Mackinnon, S.; Cole, G.J.; Levin, E.D. Long-term behavioral impairment following acute embryonic ethanol exposure in zebrafish. Neurotoxicol. Teratol. 2015, 48, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Luchiari, A.C.; Salajan, D.C.; Gerlai, R. Acute and chronic alcohol administration: Effects on performance of zebrafish in a latent learning task. Behav. Brain Res. 2015, 282, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Echevarria, D.J.; Toms, C.N.; Jouandot, D.J. Alcohol-induced behavior change in zebrafish models. Rev. Neurosci. 2011, 22, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Zenki, K.C.; Mussulini, B.H.M.; Rico, E.P.; Oliveira, D.L.d.; Rosemberg, D.B. Effects of ethanol and acetaldehyde in zebrafish brain structures: An in vitro approach on glutamate uptake and on toxicity-related parameters. Toxicol. In Vitro 2014, 28, 822–828. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.D.; Bencan, Z.; Cerutti, D.T. Anxiolytic effects of nicotine in zebrafish. Physiol. Behav. 2007, 90, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Bencan, Z.; Levin, E.D. The role of α7 and α4β2 nicotinic receptors in the nicotine-induced anxiolytic effect in zebrafish. Physiol. Behav. 2008, 95, 408–412. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.D.; Chen, E. Nicotinic involvement in memory function in zebrafish. Neurotoxicol. Teratol. 2004, 26, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Eddins, D.; Cerutti, D.; Williams, P.; Linney, E.; Levin, E.D. Zebrafish provide a sensitive model of persisting neurobehavioral effects of developmental chlorpyrifos exposure: Comparison with nicotine and pilocarpine effects and relationship to dopamine deficits. Neurotoxicol. Teratol. 2010, 32, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Zhang, X.; Guo, L.; Tian, H.; Wang, W.; Ru, S. Anti-estrogenic effect of semicarbazide in female zebrafish (Danio rerio) and its potential mechanisms. Aquat. Toxicol. 2016, 170, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Roy, N.M.; Arpie, B.; Lugo, J.; Linney, E.; Levin, E.D.; Cerutti, D. Brief embryonic strychnine exposure in zebrafish causes long-term adult behavioral impairment with indications of embryonic synaptic changes. Neurotoxicol. Teratol. 2012, 34, 587–591. [Google Scholar] [CrossRef] [PubMed]
- Wigle, D.T.; Arbuckle, T.E.; Walker, M.; Wade, M.G.; Liu, S.L.; Krewski, D. Environmental hazards: Evidence for effects on child health. J. Toxicol. Environ. Health Part B 2007, 10, 3–39. [Google Scholar] [CrossRef] [PubMed]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. In Molecular, Clinical and Environmental Toxicology: Volume 3: Environmental Toxicology; Luch, A., Ed.; Springer Basel: Basel, Switzerland, 2012; pp. 133–164. [Google Scholar]
- Depledge, M.H.; Weeks, J.M.; Bjerregaard, P. Heavy metals. In Handbook of Ecotoxicology; Blackwell Publishing Ltd.: Oxford, UK, 2009; pp. 543–569. [Google Scholar]
- Kuhlmann, A.C.; McGlothan, J.L.; Guilarte, T.R. Developmental lead exposure causes spatial learning deficits in adult rats. Neurosci. Lett. 1997, 233, 101–104. [Google Scholar] [CrossRef]
- Schwartz, J. Low-level lead exposure and children′s IQ: A metaanalysis and search for a threshold. Environ. Res. 1994, 65, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Baghurst, P.A.; McMichael, A.J.; Wigg, N.R.; Vimpani, G.V.; Robertson, E.F.; Roberts, R.J.; Tong, S.-L. Environmental exposure to lead and children’s intelligence at the age of seven years. N. Engl. J. Med. 1992, 327, 1279–1284. [Google Scholar] [CrossRef] [PubMed]
- Joe, M.B.; Kahn, R.S.; Tanya, F.; Auinger, P.; Lanphear, B.P. Exposures to environmental toxicants and attention deficit hyperactivity disorder in U.S. Children. Environ. Health Perspect. 2006, 114, 1904–1909. [Google Scholar]
- Lee, J.; Freeman, J.L. Embryonic exposure to 10 µg·L−1 lead results in female-specific expression changes in genes associated with nervous system development and function and alzheimer’s disease in aged adult zebrafish brain. Metallomics 2016, 8, 589–596. [Google Scholar] [CrossRef]
- Richetti, S.K.; Rosemberg, D.B.; Ventura-Lima, J.; Monserrat, J.M.; Bogo, M.R.; Bonan, C.D. Acetylcholinesterase activity and antioxidant capacity of zebrafish brain is altered by heavy metal exposure. Neurotoxicology 2011, 32, 116–122. [Google Scholar] [CrossRef] [PubMed]
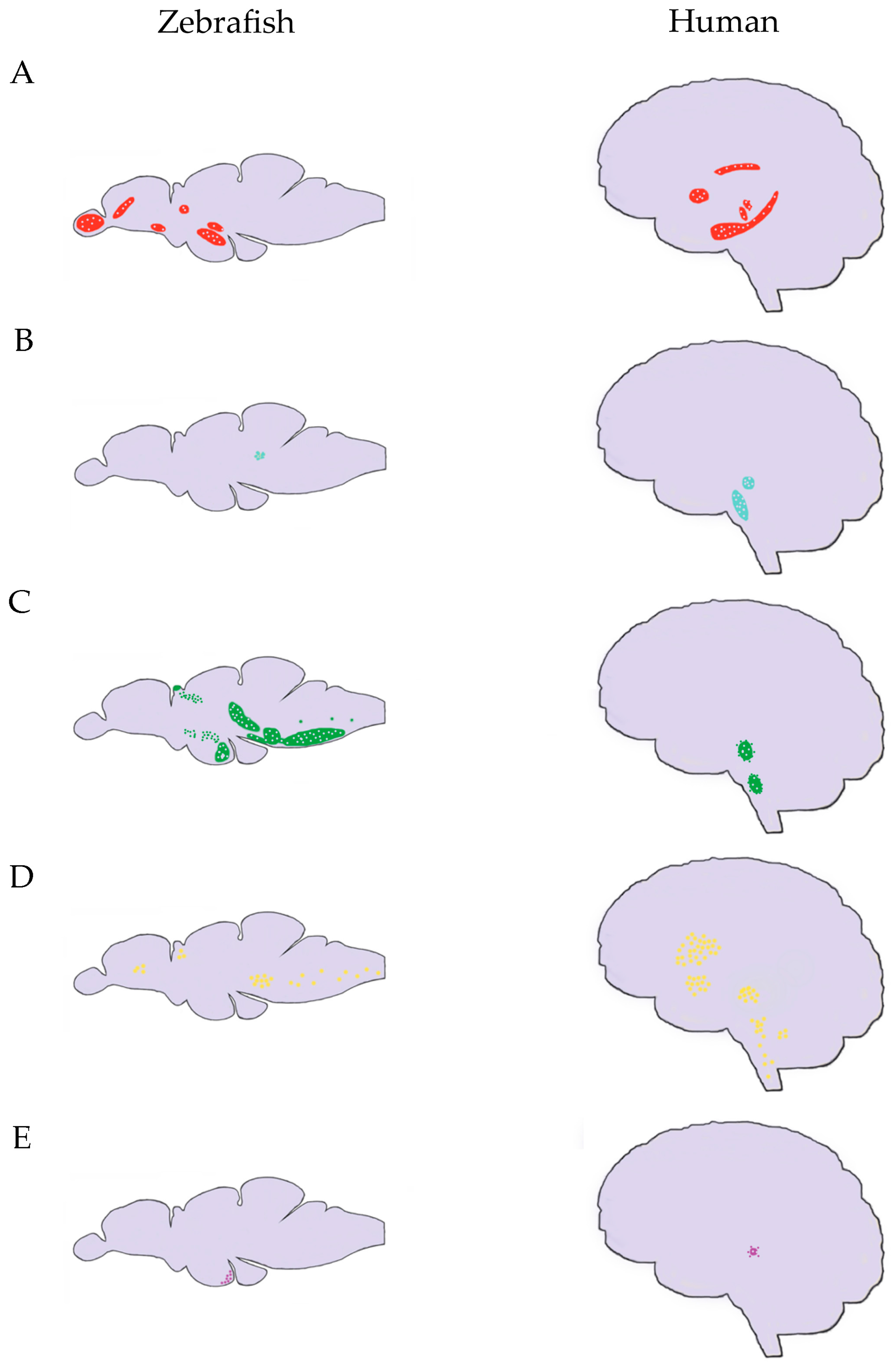

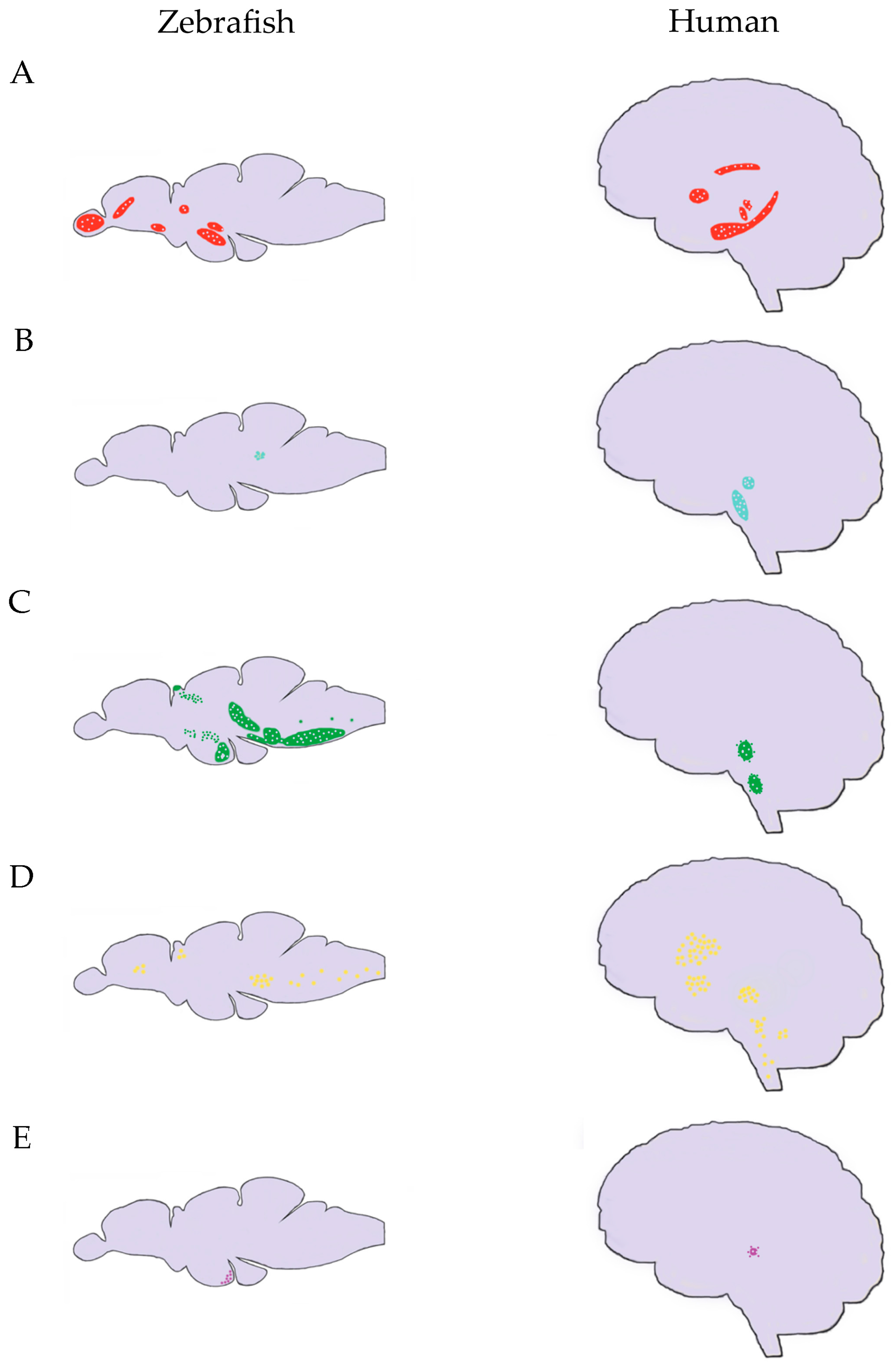{kind=link}
| Common Name | Human | Zebrafish | Gene ID | RefSeq | ZFIN ID |
|---|---|---|---|---|---|
| Glutamic-oxaloacetic transaminase 1 | GOT1 | got1 | 406330 | NM_213057 | ZDB-GENE-040426-2003 |
| Vesicular glutamate transporter 1 (VGLUT1) | SLC17A7 | slc17a7a | 795293 | NM_001098755 | ZDB-GENE-050105-5 |
| slc17a7b | 100331980 | XM_009297642 | ZDB-GENE-131125-32 | ||
| Vesicular glutamate transporter 2 (VGLUT2) | SLC17A6 | slc17a6a | 494492 | NM_001009982 | ZDB-GENE-050105-4 |
| slc17a6b | 100149756 | NM_001128821 | ZDB-GENE-030616-554 | ||
| Vesicular glutamate transporter 3 (VGLUT3) | SLC17A8 | slc17a8 | 563467 | NM_001082835 | ZDB-GENE-060503-416 |
| Glutamate-ammonia ligase (Glutamine synthetase) | GLUL | glula | 100000775 | NM_181559 | ZDB-GENE-030131-688 |
| glulb | 336473 | NM_182866 | ZDB-GENE-030131-8417 | ||
| glulc | 566165 | NM_001075114 | ZDB-GENE-060929-540 | ||
| Glutaminase | GLS | glsa | 564147 | NM_001045044 | ZDB-GENE-050204-3 |
| glsb | 564746 | XM_688079 | ZDB-GENE-030616-550 | ||
| Ionotropic Reptors | |||||
| AMPA Receptors | GRIA1 | gria1a | 798689 | NM_205598 | ZDB-GENE-020125-1 |
| gria1b | 403044 | NM_205730 | ZDB-GENE-020125-2 | ||
| GRIA2 | gria2a | 170450 | NM_131894 | ZDB-GENE-020125-3 | |
| gria2b | 170451 | NM_131895 | ZDB-GENE-020125-4 | ||
| GRIA3 | gria3a | 170452 | NM_198339 | ZDB-GENE-020125-5 | |
| gria3b | 368416 | NM_198360 | ZDB-GENE-030616-53 | ||
| GRIA4 | gria4a | 407735 | NM_214806 | ZDB-GENE-020125-7 | |
| gria4b | 336069 | NM_212752 | ZDB-GENE-030131-8013 | ||
| Kainate Receptors | GRIK1 | grik1a | 798001 | XM_009305317 | ZDB-GENE-030131-6502 |
| grik1b | 561540 | XM_684948 | ZDB-GENE-070821-1 | ||
| GRIK2 | grik2 | 556013 | XM_009300832 | ZDB-GENE-080414-1 | |
| GRIK3 | grik3 | 100334689 | XM_009300849.1 | - | |
| GRIK4 | grik4 | 556582 | XM_009291736 | ZDB-GENE-070821-5 | |
| GRIK5 | grik5 | 798791 | NM_001328156 | ZDB-GENE-070821-6 | |
| NMDA Receptors | GRIN1 | grin1a | 767745 | NM_001076714 | ZDB-GENE-051202-1 |
| grin1b | 100005675 | NM_001144131 | ZDB-GENE-051202-2 | ||
| GRIN2A | grin2aa | 563297 | XM_686662 | ZDB-GENE-070424-129 | |
| grin2ab | 570493 | XM_009306215 | ZDB-GENE-070424-223 | ||
| GRIN2B | grin2ba | - | - | ZDB-GENE-090821-2 | |
| grin2bb | 559976 | NM_001128337 | ZDB-GENE-061207-27 | ||
| GRIN2C | grin2ca | 100003342 | XM_002661129 | ZDB-GENE-070822-3 | |
| grin2cb | 100333648 | XM_009306796 | ZDB-GENE-100308-2 | ||
| GRIN2D | grin2da | 449864 | XM_009294079 | ZDB-GENE-041008-124 | |
| grin2db | - | - | ZDB-GENE-100920-7 | ||
| GRIN3A | grin3a | 564832 | XM_009305086 | ZDB-GENE-130530-780 | |
| grin3ba | 566411 | XM_009298558 | ZDB-GENE-070912-354 | ||
| GRIN3B | grin3bb | 100333101 | XM_009305920 | ZDB-GENE-131122-77 | |
| Metabotropic Receptors | |||||
| Group I | GRM1 | grm1a | 555576 | NM_001044788 | ZDB-GENE-030131-7893 |
| grm1b | 100150246 | NM_001302252 | ZDB-GENE-090821-3 | ||
| GRM5 | grm5a | 568406 | NM_001328710 | ZDB-GENE-090821-9 | |
| grm5b | 100332913 | NM_001302238 | ZDB-GENE-090821-6 | ||
| Group II | GRM2 | grm2a | 336153 | NM_001302225 | ZDB-GENE-030131-8097 |
| grm2b | 564461 | NM_001287547 | ZDB-GENE-060201-5 | ||
| GRM3 | grm3 | 565256 | NM_001128343 | ZDB-GENE-061009-13 | |
| Group III | GRM4 | grm4 | 567181 | NM_001302241 | ZDB-GENE-030131-5781 |
| GRM6 | grm6a | 568484 | NM_001123292 | ZDB-GENE-060208-1 | |
| grm6b | 565450 | NM_001080020 | ZDB-GENE-021120-2 | ||
| GRM7 | * | - | - | - | |
| GRM8 | grm8a | 792371 | NM_001302228 | ZDB-GENE-110421-2 | |
| grm8b | 569768 | NM_001287539 | ZDB-GENE-110421-3 | ||
| Glutamate Transporters | SLC1A1 | slc1a1 | 436939 | NM_001002666 | ZDB-GENE-040718-414 |
| SLC1A2 | slc1a2a | 560802 | NM_001190305 | ZDB-GENE-100422-11 | |
| slc1a2b | 335836 | NM_199979 | ZDB-GENE-030131-7779 | ||
| SLC1A3 | slc1a3a | 323439 | NM_212640 | ZDB-GENE-030131-2159 | |
| slc1a3b | 556181 | NM_001190303 | ZDB-GENE-090708-3 | ||
| SLC1A4 | slc1a4 | 368885 | NM_001002513 | ZDB-GENE-030616-566 | |
| SLC1A5 | slc1a5 | 100002129 | NM_001190755 | ZDB-GENE-070501-4 | |
| SLC1A6 | slc1a6 | 559270 | NM_001109703 | ZDB-GENE-071004-45 | |
| SCL1A7 | slc1a7a | 100170783 | NM_001291344 | ZDB-GENE-061009-24 | |
| slc1a7b | 100463517 | NM_001190760 | ZDB-GENE-101111-7 | ||
| SLC1A8 | slc1a8a | 570702 | XM_694211 | ZDB-GENE-101111-8 | |
| slc1a8b | 564474 | NM_001190816 | ZDB-GENE-070912-552 | ||
| SLC1A9 | slc1a9 | 100463516 | NM_001190759 | ZDB-GENE-101111-9 | |
| Common Name | Human | Zebrafish | Gene ID | RefSeq | ZFIN ID |
|---|---|---|---|---|---|
| Na-K-Cl cotransporter 1 (NKCC1) | SLC12A2 | slc12a2 | 415170 | NM_001002080 | ZDB-GENE-040625-53 |
| K-Cl cotransporter 2 (KCC1) | SLC12A5 | slc12a5 | 797331 | NM_001302243 | ZDB-GENE-120927-3 |
| 4-aminobutyrate transaminase | ABAT | abat | 378968 | NM_201498 | ZDB-GENE-031006-4 |
| Glutamate decarboxylase | GAD1 | gad1a | 100329827 | XM_005167412 | ZDB-GENE-070912-472 |
| gad1b | 378441 | NM_194419 | ZDB-GENE-030909-3 | ||
| GAD2 | gad2 | 550403 | NM_001017708 | ZDB-GENE-030909-9 | |
| Vesicular GABA transporter (VGAT) | SLC32A | slc32a1 | 798575 | NM_001080701 | ZDB-GENE-061201-1 |
| GABA Transporter 1 (GAT1) | SLC6A1 | slc6a1a | 692318 | NM_001045287 | ZDB-GENE-060519-23 |
| slc6a1b | 492490 | NM_001007362 | ZDB-GENE-041114-57 | ||
| slc6a1l | 568985 | XM_692346 | ZDB-GENE-041210-296 | ||
| GABA Transporter 3 (GAT3) | SLC6A11 | slc6a11a | 558960 | NM_001098387 | ZDB-GENE-030131-3729 |
| slc6a11b | 100150472 | XM_001919885 | ZDB-GENE-121116-2 | ||
| Succinic semialdehyde dehydrogenase | ALDH5A | aldh5a1 | 565235 | NM_001110468 | ZDB-GENE-070228-2 |
| GABA Receptors | |||||
| GABAA Receptor Subunit α | GABRA1 | gabra1 | 768183 | NM_001077326 | ZDB-GENE-061013-194 |
| GABRA2 | gabra2 | 100150704 | XM_009307207 | ZDB-GENE-141216-16 | |
| GABRA3 | gabra3 | 100538116 | XM_009295708 | ZDB-GENE-091204-365 | |
| GABRA4 | - | - | - | - | |
| GABRA5 | gabra5 | 799124 | XM_001339475 | ZDB-GENE-081104-30 | |
| GABRA6 | gabra6a | 393704 | NM_200731 | ZDB-GENE-040426-1692 | |
| gabra6b | 559693 | XM_002667357 | ZDB-GENE-080815-1 | ||
| GABAA Receptor Subunit β | GABRB1 | gabrb1 | 100331377 | XM_002664133 | ZDB-GENE-090313-230 |
| GABRB2 | gabrb2 | 336252 | NM_001024387 | ZDB-GENE-030131-8196 | |
| gabrb2l | 100332196 | XM_005174450 | ZDB-GENE-111215-5 | ||
| GABRB3 | gabrb3 | 566922 | XM_005166079 | ZDB-GENE-101102-2 | |
| - | gabrb4 | 566514 | XM_005173874XM_017353011 | ZDB-GENE-070424-211 | |
| GABAA Receptor Subunit γ | GABRG1 | - | - | - | - |
| GABRG2 | gabrg2 | 553402 | NM_001256250 | ZDB-GENE-091118-65 | |
| GABRG3 | gabrg3 | 567057 | XM_009302568 | ZDB-GENE-070718-5 | |
| GABAA Receptor Subunit δ | GARBD | gabrd | 571422 | XM_695007 | ZDB-GENE-081105-170 |
| GABAA Receptor Subunit π | GABRP | gabrp | 566633 | XM_005173293 | ZDB-GENE-081028-62 |
| GABAA Receptor Subunit ρ | GABRR1 | gabrr1 | 568984 | NM_001025553 | ZDB-GENE-040724-212 |
| GABR2 | gabrr2a | 751659 | NM_001045376 | ZDB-GENE-060825-164 | |
| gabrr2b | 569032 | XM_692394 | ZDB-GENE-041014-174 | ||
| GABR3 | gabrr3a | 570876 | NM_001128760 | ZDB-GENE-080722-20 | |
| gabrr3b | - | - | ZDB-GENE-131120-131 | ||
| GABAA Receptor Subunit ζ | GABRZ | gabrz | 561738 | NM_001114742 | ZDB-GENE-080303-26 |
| GABAB Receptor 1 | GABBR1 | gabbr1a | 373873 | XM_689405 | ZDB-GENE-030904-5 |
| gabbr1b | 558708 | XM_005170102 | ZDB-GENE-060503-5 | ||
| GABAB Receptor 2 | GABBR2 | gabbr2 | 560267 | NM_001144043 | ZDB-GENE-060503-620 |
| Common Name | Human | Zebrafish | Gene ID | RefSeq | ZFIN ID |
|---|---|---|---|---|---|
| Tyrosine hydroxylase | TH | th | 30384 | NM_131149 | ZDB-GENE-990621-5 |
| Aromatic amino acid decarboxylase | AAAD | aaad | 406651 | NM_213342 | ZDB-GENE-040426-2656 |
| Dopamine-β-hydroxylase | DBH | dbh | 30505 | NM_001109694 | ZDB-GENE-990621-3 |
| Phenylethanolamine-N-methyltransferase | PNMT | pnmt | 100332609 | XM_002666341 | - |
| Vesicular monoamine transporter 2 (VMAT2) | SLC18A2 | slc18a2 | 553304 | NM_001256225 | ZDB-GENE-080514-1 |
| Dopamine transporter (DAT) | SLC6A3 | slc6a3 | 80787 | NM_131755 | ZDB-GENE-010316-1 |
| Norepinephrine transporter (NET) | SLC6A2 | slc6a2 | 565776 | XM_689046 | ZDB-GENE-110408-4 |
| Catechol-O-methyltransferase | COMT | comta | 561372 | NM_001030157 | ZDB-GENE-050913-117 |
| comtb | 565370 | NM_001083843 | ZDB-GENE-040724-164 | ||
| Dopamine Receptors | DRD1 | drd1a | 792634 | XM_017359120 | ZDB-GENE-130522-1 |
| drd1b | 568126 | NM_001135976 | ZDB-GENE-070524-2 | ||
| DRD2 | drd2a | 282557 | NM_183068 | ZDB-GENE-021119-2 | |
| drd2b | 378719 | NM_197936 | ZDB-GENE-030910-2 | ||
| drd2l | 378718 | NM_197935 | ZDB-GENE-030910-1 | ||
| DRD3 | drd3 | 282554 | NM_183067 | ZDB-GENE-021119-1 | |
| DRD4 | drd4a | 503564 | NM_001012616 | ZDB-GENE-070112-996 | |
| drd4b | 503565 | NM_001012618 | ZDB-GENE-070508-3 | ||
| DRD5 | drd5a | 100536970 | XM_003199767 | ZDB-GENE-130522-2 | |
| drd5b | - | - | ZDB-GENE-130522-3 | ||
| Adrenergic Receptors | ADRA1A | adra1aa | 798498 | NM_001324454 | ZDB-GENE-030131-2831 |
| adra1ab | 557259 | XM_680297 | ZDB-GENE-060503-384 | ||
| ADRA1B | adra1ba | 100149100 | XM_001921978 | ZDB-GENE-120510-1 | |
| adra1bb | 492486 | NM_001007358 | ZDB-GENE-041114-51 | ||
| ADRA1D | adra1d | 568614 | XM_691951 | ZDB-GENE-090312-203 | |
| ADRA2 | adra2a | 266750 | NM_207637 | ZDB-GENE-021010-1 | |
| ADRA2B | adra2b | 266751 | NM_207638 | ZDB-GENE-021010-2 | |
| ADRA2C | adra2c | 266752 | NM_207639 | ZDB-GENE-021010-3 | |
| adra2da | 266754 | NM_194364 | ZDB-GENE-021010-4 | ||
| adra2db | 266755 | NM_194365 | ZDB-GENE-021010-5 | ||
| ADRB1 | adrb1 | 557194 | NM_001128689 | ZDB-GENE-081022-145 | |
| ADRB2 | adrb2a | 565838 | NM_001102652 | ZDB-GENE-100414-3 | |
| adrb2b | 100037315 | NM_001089471 | ZDB-GENE-070410-32 | ||
| ADRB3 | adrb3a | 558248 | NM_001128335 | ZDB-GENE-080917-21 | |
| adrb3b | 792519 | NM_001135134 | ZDB-GENE-081022-154 |
| Common Name | Human | Zebrafish | Gene ID | RefSeq | ZFIN ID |
|---|---|---|---|---|---|
| Tryptophan hydroxylase | TPH1 | tph1a | 352943 | NM_178306 | ZDB-GENE-030317-1 |
| tph1b | 415103 | NM_001001843 | ZDB-GENE-030805-6 | ||
| TPH2 | tph2 | 407712 | NM_001310068 | ZDB-GENE-040624-4 | |
| tph3/th2 | 414844 | NM_001001829 | ZDB-GENE-050201-1 | ||
| Aromatic amino acid decarboxylase | AAAD | aaad | 406651 | NM_212827 | ZDB-GENE-040426-2656 |
| Monoamine oxidase | MAO-A, MAO-B | mao | 404730 | NM_001039972 | ZDB-GENE-040329-3 |
| Serotonin Transporter (SERT) | SLC6A4 | slc6a4a | 664719 | NM_001177459 | ZDB-GENE-060314-1 |
| slc6a4b | 664770 | NM_001123321 | ZDB-GENE-060314-2 | ||
| Serotonin Receptors | HTR1A | htr1aa | 100001828 | NM_001145766 | ZDB-GENE-071203-1 |
| htr1ab | 797538 | NM_001128709 | ZDB-GENE-090409-2 | ||
| HTR1B | htr1b | 561647 | NM_001145686 | ZDB-GENE-081022-141 | |
| htr1d | 556429 | NM_001145686 | ZDB-GENE-090409-3 | ||
| htr1fa | 100005344 | XM_017357893 | ZDB-GENE-081105-125 | ||
| HTR2 | htr2a | 560808 | NM_001044743 | ZDB-GENE-070912-500 | |
| htr2b | 751784 | NM_001044743 | ZDB-GENE-081022-57 | ||
| HTR2CL1 | htr2cl1 | 100000981 | XM_001339004 | ZDB-GENE-081104-48 | |
| HTR2CL2 | htr2cl2 | 798599 | XM_001339004 | ZDB-GENE-120215-109 | |
| HTR3 | htr3a | 571641 | XM_009295409 | ZDB-GENE-071012-5 | |
| htr3b | 571632 | NM_001126410 | ZDB-GENE-071012-4 | ||
| HTR5 | htr5a | 100038775 | NM_001007121 | ZDB-GENE-060531-129 | |
| HTR5-like | htr5al | 368475 | XM_009297078 | ZDB-GENE-030616-574 | |
| HTR6 | htr6 | 568269 | XM_685507 | ZDB-GENE-030131-7839 | |
| HTR7 | htr7 | 562111 | NM_178306 | ZDB-GENE-130530-666 |
| Common Name | Human | Zebrafish | Gene ID | RefSeq | ZFIN ID |
|---|---|---|---|---|---|
| High-affinity choline transporter | SLC5A7 | slc5a7a | 100005589 | XM_005159931 | ZDB-GENE-090313-273 |
| slc5a7b | - | - | ZDB-GENE-140429-1 | ||
| Choline acetyltransferase | CHAT | chata | 100170938 | NM_001130719 | ZDB-GENE-080102-2 |
| chatb | 103171573 | NM_001291882 | ZDB-GENE-140429-2 | ||
| Vesicular ACh transporter (VAChT) | SLC18A3 | slc18a3a | 559347 | NM_001077550 | ZDB-GENE-060929-990 |
| slc18a3b | 394082 | NM_201107 | ZDB-GENE-040426-1410 | ||
| Acetylcholinesterase | ACHE | ache | 114549 | NM_131846 | ZDB-GENE-010906-1 |
| Nicotinic Cholinergic Receptors | CHRNA1 | chrna1 | 30725 | NM_131445 | ZDB-GENE-980526-137 |
| CHRNA2 | chrna2a | 678575 | NM_001040327 | ZDB-GENE-040108-2 | |
| chrna2b | 568849 | XM_692206 | ZDB-GENE-041001-99 | ||
| CHRNA3 | chrna3 | 568467 | XM_001921279 | ZDB-GENE-070822-1 | |
| CHRNA4 | chrna4a | - | - | ZDB-GENE-130530-903 | |
| chrna4b | 556619 | NM_001048063 | ZDB-GENE-090505-3 | ||
| CHRNA5 | chrna5 | 550584 | NM_001017885 | ZDB-GENE-050417-440 | |
| CHRNA6 | chrna6 | 555747 | NM_001042684 | ZDB-GENE-090312-91 | |
| CHRNA7 | chrna7 | 394199 | NM_201219 | ZDB-GENE-040108-3 | |
| CHRNA9 | chrna9 | 568807 | XM_001920859 | ZDB-GENE-090312-63 | |
| CHRNA10 | chrna10a | 556507 | NM_001044804 | ZDB-GENE-060503-725 | |
| chrna10b | - | - | ZDB-GENE-130530-624 | ||
| Muscarinic Cholinergic Receptors | CHRM1 | chrm1a | 792708 | XM_001332257 | ZDB-GENE-090410-9 |
| chrm1b | 794658 | NM_178301 | ZDB-GENE-070705-188 | ||
| CHRM2 | chrm2a | 352938 | NM_178301 | ZDB-GENE-030314-1 | |
| chrm2b | 555516 | XM_678041 | ZDB-GENE-090410-3 | ||
| CHRM3 | chrm3a | 571679 | XM_695289 | ZDB-GENE-090410-4 | |
| chrm3b | 100149598 | XM_001919125 | ZDB-GENE-090410-5 | ||
| CHRM4 | chrm4a | 100150701 | XM_001922407 | ZDB-GENE-090410-6 | |
| chrm4b | - | - | ZDB-GENE-090410-7 | ||
| CHRM5 | chrm5a | 553978 | NM_001020803 | ZDB-GENE-080723-32 | |
| chrm5b | 561491 | NM_001030160 | ZDB-GENE-041001-169 |
| Common Name | Human | Zebrafish | Gene ID | RefSeq | ZFIN ID |
|---|---|---|---|---|---|
| l-Histidine decarboxylase | HDC | hdc | 793609 | NM_001102593 | ZDB-GENE-080102-5 |
| Amine oxidase | AOC1 | aoc1 | 555401 | NM_001077598 | ZDB-GENE-061103-112 |
| Histamine N-methyltransferase | HNMT | hnmt | 445242 | NM_001003636 | ZDB-GENE-040801-157 |
| Histamine Receptors | HRH1 | hrh1 | 735302 | NM_001042731 | ZDB-GENE-070531-3 |
| HRH2 | hrh2a | 735303 | NM_001045338 | ZDB-GENE-070531-4 | |
| hrh2b | 100005590 | NM_001109738 | ZDB-GENE-070928-20 | ||
| HRH3 | hrh3 | 561773 | NM_001025518 | ZDB-GENE-040724-204 |
| Common Name | Human | Zebrafish | Gene ID | RefSeq | ZFIN ID |
|---|---|---|---|---|---|
| Glycine dehydrogenase | GLDC | gldc | 321621 | NM_199554 | ZDB-GENE-030131-340 |
| Serine hydroxymethyltransferase 1 | SHMT1 | shmt1 | 394021 | NM_201046 | ZDB-GENE-040426-1558 |
| Serine hydroxymethyltransferase 2 | SMHT2 | shmt2 | 100144628 | NM_001123374 | ZDB-GENE-071213-1 |
| Inhibitory amino acid transporter | SLC32A1 | slc32a1 | 798575 | NM_001080701 | ZDB-GENE-061201-1 |
| Glycine transporter 1 | SLC6A9 | slc6a9 | 494490 | NM_001030073 | ZDB-GENE-050105-3 |
| Glycine transporter 2 | SLC6A5 | slc6a5 | 494450 | NM_001009557 | ZDB-GENE-050105-2 |
| Glycine receptor α | GLRA1 | glra1 | 30676 | NM_131402 | ZDB-GENE-991117-1 |
| GLRA2 | glra2 | 793646 | NM_001167899 | ZDB-GENE-090407-1 | |
| GLRA3 | glra3 | 192124 | NM_152965 | ZDB-GENE-020402-1 | |
| GLRA4 | glra4a | 83413 | NM_131782 | ZDB-GENE-010410-3 | |
| glra4b | 192125 | NM_001202511 | ZDB-GENE-020402-2 | ||
| Glycine receptor β | GLRB | glrba | 83412 | NM_131781 | ZDB-GENE-010410-2 |
| glrbb | 445193 | NM_001003587 | ZDB-GENE-040801-106 |
| Reference (Age, Sex, and Strain if Known) | Glutamate | GABA | DA | NE | 5-HT | ACh |
|---|---|---|---|---|---|---|
| Adult Zebrafish | ||||||
| Panula et al. [16] (Sex, age, and strain unknown) | - | - | 2.09 ± 0.42 nmol/g | 4.53 ± 0.97 nmol/g | - | - |
| López Patiño et al. [197] (Male and female 9 ± 1 mpf AB wildtype) | - | - | 1.5–2 pg/ug protein | - | - | - |
| Chatterjee and Gerlai [191] (Male and female 90 dpf AB wildtype) | - | - | 4.18 ± 0.28 ng/mg protein | - | - | - |
| Embryonic/Larval Zebrafish | ||||||
| Wirbisky et al. [193] (Embryos raised at 28.5 °C) | - | - | 78.31 ± 2.26 ng/fish (48 hpf) 99.17 ± 6.54 ng/fish (72 hpf) | - | - | - |
| Tufi et al. [195] (Embryos raised at 26 °C) | 9.1 ± 0.5 ng/embryo (48 hpf) 12 ± 0.3 ng/embryo (72 hpf) | 1.8 ± 0.03 ng/embryo (48 hpf) 2.2 ± 0.1 ng/embryo (72 hpf) | - | - | 7.2 ± 0.01 pg/embryo (48 hpf) 7.1 ± 0.1 pg/embryo (72 hpf) | 3.0 ± 0.1 pg/embyro (48 hpf) 4.0 ± 0.1 pg/embyro (72 hpf) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horzmann, K.A.; Freeman, J.L. Zebrafish Get Connected: Investigating Neurotransmission Targets and Alterations in Chemical Toxicity. Toxics 2016, 4, 19. https://doi.org/10.3390/toxics4030019
Horzmann KA, Freeman JL. Zebrafish Get Connected: Investigating Neurotransmission Targets and Alterations in Chemical Toxicity. Toxics. 2016; 4(3):19. https://doi.org/10.3390/toxics4030019
Chicago/Turabian StyleHorzmann, Katharine A., and Jennifer L. Freeman. 2016. "Zebrafish Get Connected: Investigating Neurotransmission Targets and Alterations in Chemical Toxicity" Toxics 4, no. 3: 19. https://doi.org/10.3390/toxics4030019
APA StyleHorzmann, K. A., & Freeman, J. L. (2016). Zebrafish Get Connected: Investigating Neurotransmission Targets and Alterations in Chemical Toxicity. Toxics, 4(3), 19. https://doi.org/10.3390/toxics4030019