
Prepubertal Exposure to Tris(2-chloroethyl) Phosphate Disrupts Blood-Testis Barrier Integrity via Ferritinophagy-Mediated Ferroptosis

Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Testis Coefficient
2.3. Hematoxylin-Eosin (H&E) Staining
2.4. TM4 Cell Culture
2.5. CCK-8 Analysis
2.6. Transmission Electron Microscopy (TEM)
2.7. Analysis of Ferrous Iron
2.8. Detection of Glutathione (GSH)
2.9. Determination of Lipid Peroxidation
2.10. Western-Blot Analysis
2.11. Statistical Analysis
3. Results
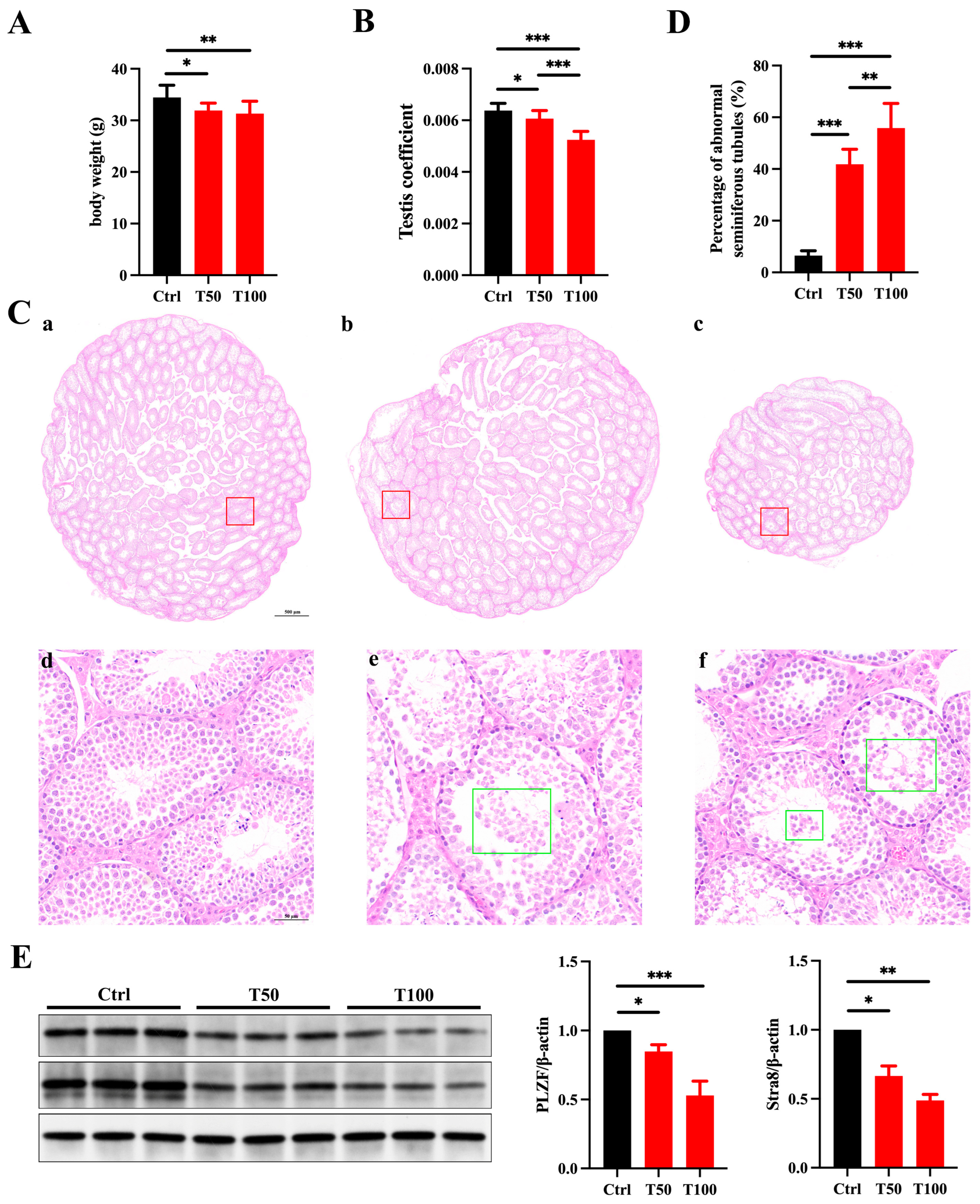
3.1. Prepubertal Exposure to TCEP Leads to Testicular Developmental Toxicity
3.2. TCEP Exposure Disrupts BTB Integrity
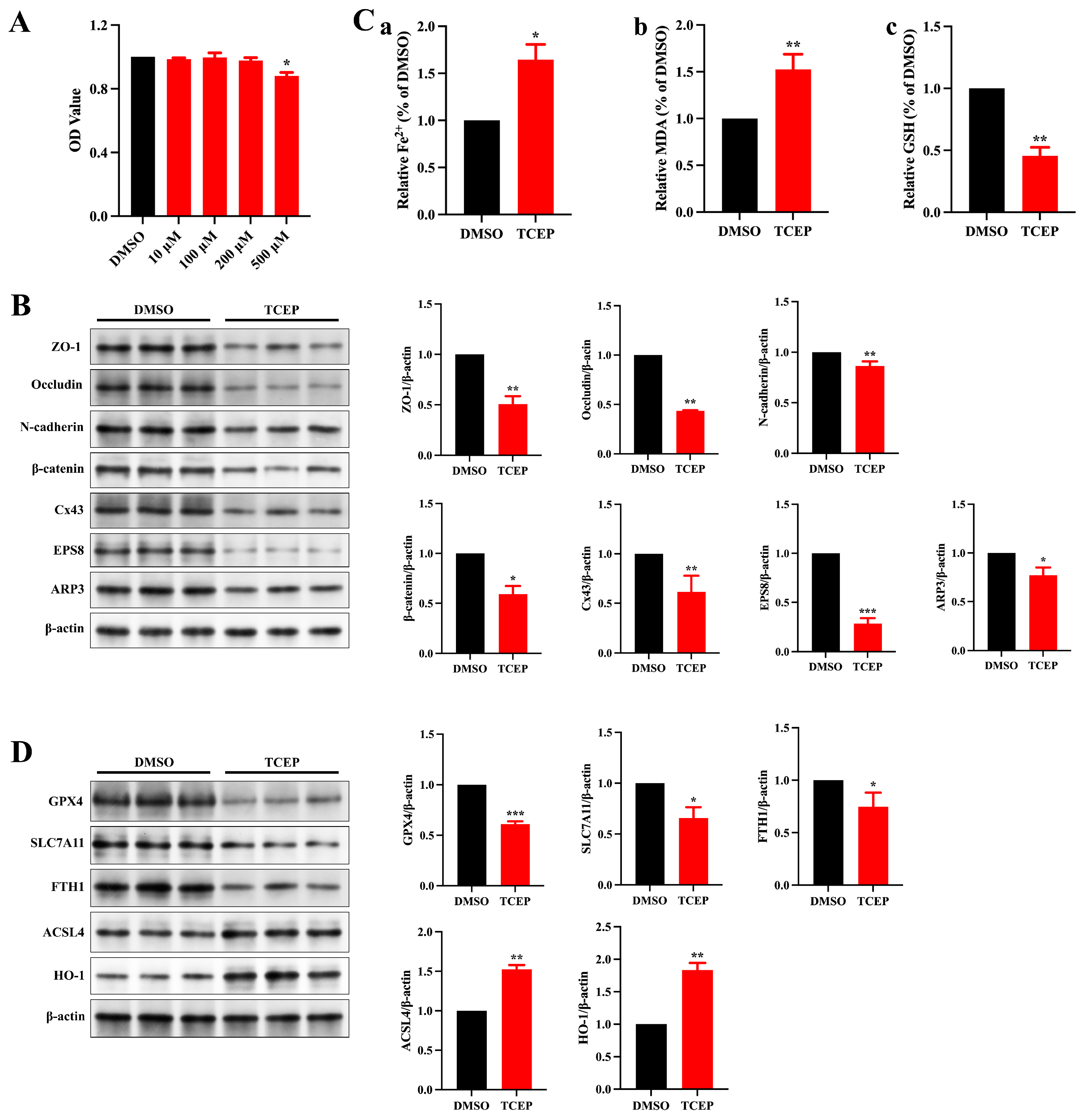
3.3. TCEP Exposure Induces Sertoli Cell Ferroptosis
3.4. TCEP Exposure Impairs Sertoli Cell Junctional Functions via Ferroptosis
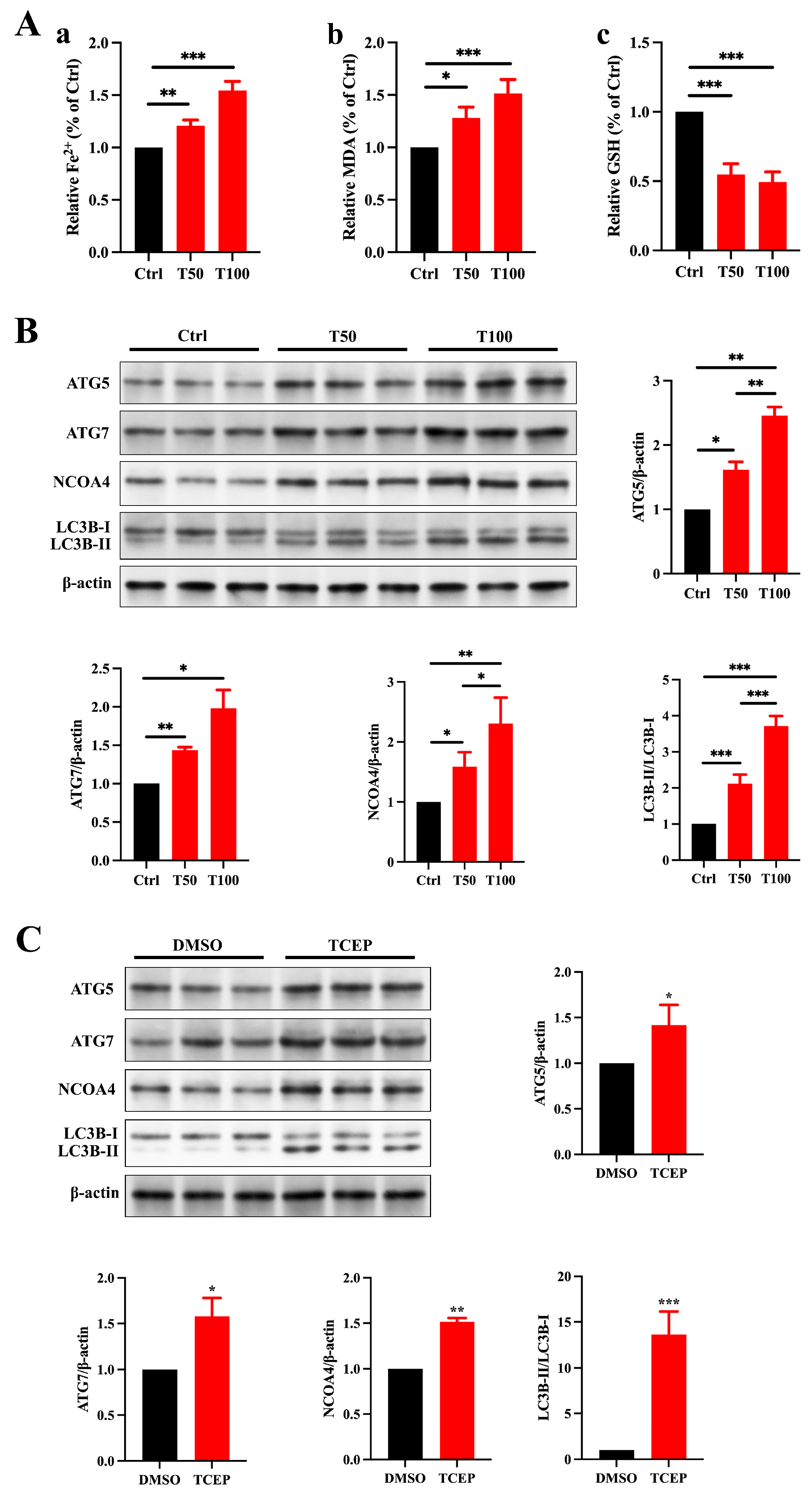
3.5. TCEP Induces Ferroptosis in Sertoli Cells and the Testis via Ferritinophagy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blum, A.; Behl, M.; Birnbaum, L.S.; Diamond, M.L.; Phillips, A.; Singla, V.; Sipes, N.S.; Stapleton, H.M.; Venier, M. Organophosphate Ester Flame Retardants: Are They a Regrettable Substitution for Polybrominated Diphenyl Ethers? Environ. Sci. Technol. Lett. 2019, 6, 638–649. [Google Scholar] [PubMed]
- Lee, S.; Jeong, W.; Kannan, K.; Moon, H.-B. Occurrence and exposure assessment of organophosphate flame retardants (OPFRs) through the consumption of drinking water in Korea. Water Res. 2016, 103, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Fang, B.; Wang, C.; Du, X.; Sun, G.; Jia, B.; Liu, X.; Qu, Y.; Zhang, Q.; Yang, Y.; Li, Y.Q.; et al. Structure-dependent destructive adsorption of organophosphate flame retardants on lipid membranes. J. Hazard. Mater. 2024, 478, 135494. [Google Scholar]
- Yang, J.; Li, X.; Yang, H.; Zhao, W.; Li, Y. OPFRs in e-waste sites: Integrating in silico approaches, selective bioremediation, and health risk management of residents surrounding. J. Hazard. Mater. 2022, 429, 128304. [Google Scholar]
- Ma, H.; He, J.; Fan, H.; Zhang, N.; Wu, Q.; Zhang, S.; Zhang, C.; Huang, T.; Gao, H.; Ma, J.; et al. The influence of emerging atmospheric organophosphorus flame retardants from land source emissions on the East China Sea. J. Hazard. Mater. 2024, 465, 133404. [Google Scholar] [PubMed]
- Castro-Jiménez, J.; González-Gaya, B.; Pizarro, M.; Casal, P.; Pizarro-Álvarez, C.; Dachs, J. Organophosphate Ester Flame Retardants and Plasticizers in the Global Oceanic Atmosphere. Environ. Sci. Technol. 2016, 50, 12831–12839. [Google Scholar]
- Mendy, A.; Percy, Z.; Braun, J.M.; Lanphear, B.; La Guardia, M.J.; Hale, R.C.; Yolton, K.; Chen, A. Prenatal and postnatal exposure to organophosphate esters and replacement flame retardant mixtures and childhood respiratory outcomes. Environ. Res. 2024, 266, 120514. [Google Scholar]
- Win-Shwe, T.T.; Yanagisawa, R.; Lwin, T.T.; Kawakami, F.; Koike, E.; Takano, H. Dietary Exposure to Flame Retardant Tris (2-Butoxyethyl) Phosphate Altered Neurobehavior and Neuroinflammatory Responses in a Mouse Model of Allergic Asthma. Int. J. Mol. Sci. 2022, 23, 655. [Google Scholar] [CrossRef]
- Zhang, B.; Lu, S.; Huang, M.; Zhou, M.; Zhou, Z.; Zheng, H.; Jiang, Y.; Bai, X.; Zhang, T. Urinary metabolites of organophosphate flame retardants in 0-5-year-old children: Potential exposure risk for inpatients and home-stay infants. Environ. Pollut. 2018, 243, 318–325. [Google Scholar] [CrossRef]
- Li, K.; Qi, Z.; Xie, Z.; Li, W.; Yang, X.; Zhai, Y.; Zhou, X.; Xie, X.; Song, W. TDMPP activation of estrogen receptor 2a regulates smc2 and p53 signaling to interfere with liver development in zebrafish (Danio rerio). J. Hazard. Mater. 2024, 477, 135379. [Google Scholar]
- Tsai, K.-F.; Cheng, F.-J.; Huang, W.-T.; Kung, C.-T.; Lee, C.-T.; Cheng, B.-C.; Chen, J.-B.; Li, S.-H.; Wang, C.-C.; Wang, L.-J.; et al. The associations between renal disease severity and exposure to organophosphate flame retardants in patients with chronic kidney disease. Environ. Int. 2022, 170, 107573. [Google Scholar] [PubMed]
- Cui, H.; Chang, Y.; Jiang, X.; Li, M. Triphenyl phosphate exposure induces kidney structural damage and gut microbiota disorders in mice under different diets. Environ. Int. 2020, 144, 106054. [Google Scholar] [PubMed]
- Chiang, C.; Mahalingam, S.; Flaws, J.A. Environmental Contaminants Affecting Fertility and Somatic Health. Semin. Reprod. Med. 2017, 35, 241–249. [Google Scholar] [PubMed]
- Wang, H.; Ding, J.; Luo, S.; Yan, M.; Hu, F. Unveiling the mechanisms of reproductive toxicity induced by full life-cycle exposure to environmentally relevant concentrations of tris(2-chloroethyl) phosphate in male zebrafish. Aquat. Toxicol. 2024, 276, 107079. [Google Scholar]
- Chen, G.; Jin, Y.; Wu, Y.; Liu, L.; Fu, Z. Exposure of male mice to two kinds of organophosphate flame retardants (OPFRs) induced oxidative stress and endocrine disruption. Environ. Toxicol. Pharmacol. 2015, 40, 310–318. [Google Scholar]
- Chen, G.; Zhang, S.; Jin, Y.; Wu, Y.; Liu, L.; Qian, H.; Fu, Z. TPP and TCEP induce oxidative stress and alter steroidogenesis in TM3 Leydig cells. Reprod. Toxicol. 2015, 57, 100–110. [Google Scholar]
- Washburn, R.L.; Hibler, T.; Kaur, G.; Dufour, J.M. Sertoli Cell Immune Regulation: A Double-Edged Sword. Front. Immunol. 2022, 13, 913502. [Google Scholar] [CrossRef]
- Li, N.; Mruk, D.D.; Lee, W.M.; Wong, C.K.; Cheng, C.Y. Is toxicant-induced Sertoli cell injury in vitro a useful model to study molecular mechanisms in spermatogenesis? Semin. Cell Dev. Biol. 2016, 59, 141–156. [Google Scholar]
- Gao, Y.; Mruk, D.D.; Cheng, C.Y. Sertoli cells are the target of environmental toxicants in the testis—A mechanistic and therapeutic insight. Expert Opin. Ther. Targets 2015, 19, 1073–1090. [Google Scholar]
- Huang, X.; Fu, Y.; Wang, S.; Guo, Q.; Wu, Y.; Zheng, X.; Wang, J.; Wu, S.; Shen, L.; Wei, G. 2,2′,4,4′-Tetrabromodiphenyl ether exposure disrupts blood-testis barrier integrity through CMA-mediated ferroptosis. Sci. Total Environ. 2024, 948, 174738. [Google Scholar]
- Jiang, X.; Stockwell, B.R.; Conrad, M. Ferroptosis: Mechanisms, biology and role in disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 266–282. [Google Scholar] [CrossRef] [PubMed]
- Santana-Codina, N.; Gikandi, A.; Mancias, J.D. The Role of NCOA4-Mediated Ferritinophagy in Ferroptosis. Adv. Exp. Med. Biol. 2021, 1301, 41–57. [Google Scholar] [PubMed]
- Allen, C.M.; Lopes, F.; Mitchell, R.T.; Spears, N. How does chemotherapy treatment damage the prepubertal testis? Reproduction 2018, 156, R209–R233. [Google Scholar]
- Kanatsu-Shinohara, M.; Shinohara, T. Spermatogonial stem cell self-renewal and development. Annu. Rev. Cell Dev. Biol. 2013, 29, 163–187. [Google Scholar]
- Li, N.; Tang, E.I.; Cheng, C.Y. Regulation of blood-testis barrier by actin binding proteins and protein kinases. Reproduction 2016, 151, R29–R41. [Google Scholar]
- Lie, P.P.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Epidermal growth factor receptor pathway substrate 8 (Eps8) is a novel regulator of cell adhesion and the blood-testis barrier integrity in the seminiferous epithelium. FASEB J. 2009, 23, 2555–2567. [Google Scholar]
- Lie, P.P.; Chan, A.Y.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Restricted Arp3 expression in the testis prevents blood-testis barrier disruption during junction restructuring at spermatogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 11411–11416. [Google Scholar] [PubMed]
- Qadeer, A.; Mubeen, S.; Liu, M.; Bekele, T.G.; Ohoro, C.R.; Adeniji, A.O.; Alraih, A.M.; Ajmal, Z.; Alshammari, A.S.; Al-Hadeethi, Y.; et al. Global environmental and toxicological impacts of polybrominated diphenyl ethers versus organophosphate esters: A comparative analysis and regrettable substitution dilemma. J. Hazard. Mater. 2024, 466, 133543. [Google Scholar] [CrossRef]
- Hoehn, R.M.; Jahl, L.G.; Herkert, N.J.; Hoffman, K.; Soehl, A.; Diamond, M.L.; Blum, A.; Stapleton, H.M. Flame Retardant Exposure in Vehicles Is Influenced by Use in Seat Foam and Temperature. Environ. Sci. Technol. 2024, 58, 8825–8834. [Google Scholar]
- Fu, J.; Fu, K.; Hu, B.; Zhou, W.; Fu, Y.; Gu, L.; Zhang, Q.; Zhang, A.; Fu, J.; Jiang, G. Source Identification of Organophosphate Esters through the Profiles in Proglacial and Ocean Sediments from Ny-Ålesund, the Arctic. Environ. Sci. Technol. 2023, 57, 1919–1929. [Google Scholar]
- Yan, Z.; Feng, C.; Leung, K.M.; Luo, Y.; Wang, J.; Jin, X.; Wu, F. Insights into the geographical distribution, bioaccumulation characteristics, and ecological risks of organophosphate esters. J. Hazard. Mater. 2023, 445, 130517. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhao, Y.; Li, M.; Du, M.; Li, X.; Li, Y. A Review of a Class of Emerging Contaminants: The Classification, Distribution, Intensity of Consumption, Synthesis Routes, Environmental Effects and Expectation of Pollution Abatement to Organophosphate Flame Retardants (OPFRs). Int. J. Mol. Sci. 2019, 20, 2874. [Google Scholar] [CrossRef]
- Yang, T.; Zhou, X.; Wu, Y.; Liang, Y.; Zeng, X.; Yu, Z. Metagenomic analyses of aerobic bacterial enrichment cultures that degraded Tris(2-chloroethyl) phosphate (TCEP) and its transformation products. Environ. Pollut. 2024, 361, 124825. [Google Scholar]
- Reproductive toxicology. Tris(2-chloroethyl)phosphate. Environ. Health Perspect. 1997, 105 (Suppl. S1), 365–366.
- Cheng, C.Y.; Mruk, D.D. The blood-testis barrier and its implications for male contraception. Pharmacol. Rev. 2012, 64, 16–64. [Google Scholar] [PubMed]
- Li, N.; Mruk, D.D.; Cheng, C.Y. Actin binding proteins in blood-testis barrier function. Curr. Opin. Endocrinol. Diabetes Obes. 2015, 22, 238–247. [Google Scholar] [PubMed]
- Chen, R.; Wang, F.; Chen, Y.; Han, D. Immune homeostasis and disorder in the testis—Roles of Sertoli cells. J. Reprod. Immunol. 2022, 152, 103625. [Google Scholar]
- Wang, J.; Wei, Y.; Wu, Y.; Zhao, T.; Kang, L.; Han, L.; Chen, J.; Long, C.; Wei, G.; Shen, L.; et al. Di-(2-ethylhexyl) phthalate induces prepubertal testicular injury through MAM-related mitochondrial calcium overload in Leydig and Sertoli cell apoptosis. Toxicology 2024, 509, 153956. [Google Scholar] [CrossRef]
- Wu, H.; Wei, Y.; Zhou, Y.; Long, C.; Hong, Y.; Fu, Y.; Zhao, T.; Wang, J.; Wu, Y.; Wu, S.; et al. Bisphenol S perturbs Sertoli cell junctions in male rats via alterations in cytoskeletal organization mediated by an imbalance between mTORC1 and mTORC2. Sci. Total Environ. 2021, 762, 144059. [Google Scholar] [CrossRef]
- Wei, Y.; Zhou, Y.; Long, C.; Wu, H.; Hong, Y.; Fu, Y.; Wang, J.; Wu, Y.; Shen, L.; Wei, G. Polystyrene microplastics disrupt the blood-testis barrier integrity through ROS-Mediated imbalance of mTORC1 and mTORC2. Environ. Pollut. 2021, 289, 117904. [Google Scholar] [CrossRef]
- Perobelli, J.E. The male peripubertal phase as a developmental window for reproductive toxicology studies. Curr. Pharm. Des. 2014, 20, 5398–5415. [Google Scholar]
- van den Driesche, S.; Kilcoyne, K.R.; Wagner, I.; Rebourcet, D.; Boyle, A.; Mitchell, R.; McKinnell, C.; Macpherson, S.; Donat, R.; Shukla, C.J.; et al. Experimentally induced testicular dysgenesis syndrome originates in the masculinization programming window. JCI Insight 2017, 2, e91204. [Google Scholar] [PubMed]
- Kilcoyne, K.R.; Mitchell, R.T. Effect of environmental and pharmaceutical exposures on fetal testis development and function: A systematic review of human experimental data. Hum. Reprod. Update 2019, 25, 397–421. [Google Scholar] [PubMed]
- Shi, J.; Gao, S.; Chen, Z.; Chen, Z.; Yun, D.; Wu, X.; Sun, F. Absence of MerTK disrupts spermatogenesis in an age-dependent manner. Mol. Cell. Endocrinol. 2023, 560, 111815. [Google Scholar]
- Wang, H.; Wang, H.; Xiong, W.; Chen, Y.; Ma, Q.; Ma, J.; Ge, Y.; Han, D. Evaluation on the phagocytosis of apoptotic spermatogenic cells by Sertoli cells in vitro through detecting lipid droplet formation by Oil Red O staining. Reproduction 2006, 132, 485–492. [Google Scholar] [PubMed]
- Hu, F.; Li, W.; Wang, H.; Peng, H.; He, J.; Ding, J.; Zhang, W. Environmentally relevant concentrations of tris (2-chloroethyl) phosphate (TCEP) induce hepatotoxicity in zebrafish (Danio rerio): A whole life-cycle assessment. Fish Physiol. Biochem. 2023, 49, 1421–1433. [Google Scholar]
- Meng, Y.; Xu, X.; Niu, D.; Xu, Y.; Qiu, Y.; Zhu, Z.; Zhang, H.; Yin, D. Organophosphate flame retardants induce oxidative stress and Chop/Caspase 3-related apoptosis via Sod1/p53/Map3k6/Fkbp5 in NCI-1975 cells. Sci. Total Environ. 2022, 819, 153160. [Google Scholar] [CrossRef]
- Wang, C.; Chen, Z.; Lu, Y.; Wang, L.; Zhang, Y.; Zhu, X.; Song, J. Neurotoxicity and related mechanisms of flame retardant TCEP exposure in mice. Toxicol. Mech. Methods 2020, 30, 490–496. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, J.; Zhao, T.; Chen, J.; Kang, L.; Wei, Y.; Han, L.; Shen, L.; Long, C.; Wu, S.; et al. Di-(2-ethylhexyl) phthalate exposure leads to ferroptosis via the HIF-1α/HO-1 signaling pathway in mouse testes. J. Hazard. Mater. 2022, 426, 127807. [Google Scholar] [CrossRef]
- Peng, H.; Wang, H.; Li, W.; Jing, C.; Zhang, W.; Zhao, H.; Hu, F. Life-cycle exposure to tris (2-chloroethyl) phosphate (TCEP) causes alterations in antioxidative status, ion regulation and histology of zebrafish gills. Comp. Biochem. Physiol. Toxicol. Pharmacol. CBP 2023, 274, 109746. [Google Scholar] [CrossRef]
- Muhoberac, B.B.; Vidal, R. Iron, Ferritin, Hereditary Ferritinopathy, and Neurodegeneration. Front. Neurosci. 2019, 13, 1195. [Google Scholar] [CrossRef] [PubMed]
- Sui, H.; Sun, Z.; Liu, C.; Xi, H. Ferritinophagy promotes microglia ferroptosis to aggravate neuroinflammation induced by cerebral ischemia-reperfusion injury via activation of the cGAS-STING signaling pathway. Neurochem. Int. 2024, 183, 105920. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.; Lin, J.; Ji, L.; Ye, L.; Ling, M.; Liao, X.; Lin, F.; Wang, Y.; Luo, B.; Hu, Z.; et al. Ferritinophagy mediated by the AMPK/ULK1 pathway is involved in ferroptosis subsequent to ventilator-induced lung injury. Respir. Res. 2024, 25, 440. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Liu, Z.; Liu, J.; Li, Z.; Bao, Y.; Sun, X.; Zhao, W.; Zhou, A.; Wu, H. NCOA4 linked to endothelial cell ferritinophagy and ferroptosis:a key regulator aggravate aortic endothelial inflammation and atherosclerosis. Redox Biol. 2024, 79, 103465. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Peng, M.; Liu, H.; Zhang, X.; Fu, D. Prepubertal Exposure to Tris(2-chloroethyl) Phosphate Disrupts Blood-Testis Barrier Integrity via Ferritinophagy-Mediated Ferroptosis. Toxics 2025, 13, 285. https://doi.org/10.3390/toxics13040285
Zhao Y, Peng M, Liu H, Zhang X, Fu D. Prepubertal Exposure to Tris(2-chloroethyl) Phosphate Disrupts Blood-Testis Barrier Integrity via Ferritinophagy-Mediated Ferroptosis. Toxics. 2025; 13(4):285. https://doi.org/10.3390/toxics13040285
Chicago/Turabian StyleZhao, Yonggang, Mo Peng, Honglei Liu, Xiaoyu Zhang, and Dan Fu. 2025. "Prepubertal Exposure to Tris(2-chloroethyl) Phosphate Disrupts Blood-Testis Barrier Integrity via Ferritinophagy-Mediated Ferroptosis" Toxics 13, no. 4: 285. https://doi.org/10.3390/toxics13040285
APA StyleZhao, Y., Peng, M., Liu, H., Zhang, X., & Fu, D. (2025). Prepubertal Exposure to Tris(2-chloroethyl) Phosphate Disrupts Blood-Testis Barrier Integrity via Ferritinophagy-Mediated Ferroptosis. Toxics, 13(4), 285. https://doi.org/10.3390/toxics13040285