

The Neurobehavioral Impact of Zinc Chloride Exposure in Zebrafish: Evaluating Cognitive Deficits and Probiotic Modulation

 ,
, 
 , , ,
, , ,  , and
, and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Ethical Note
2.2. Animal Maintenance
2.3. Chemical Compounds Administration
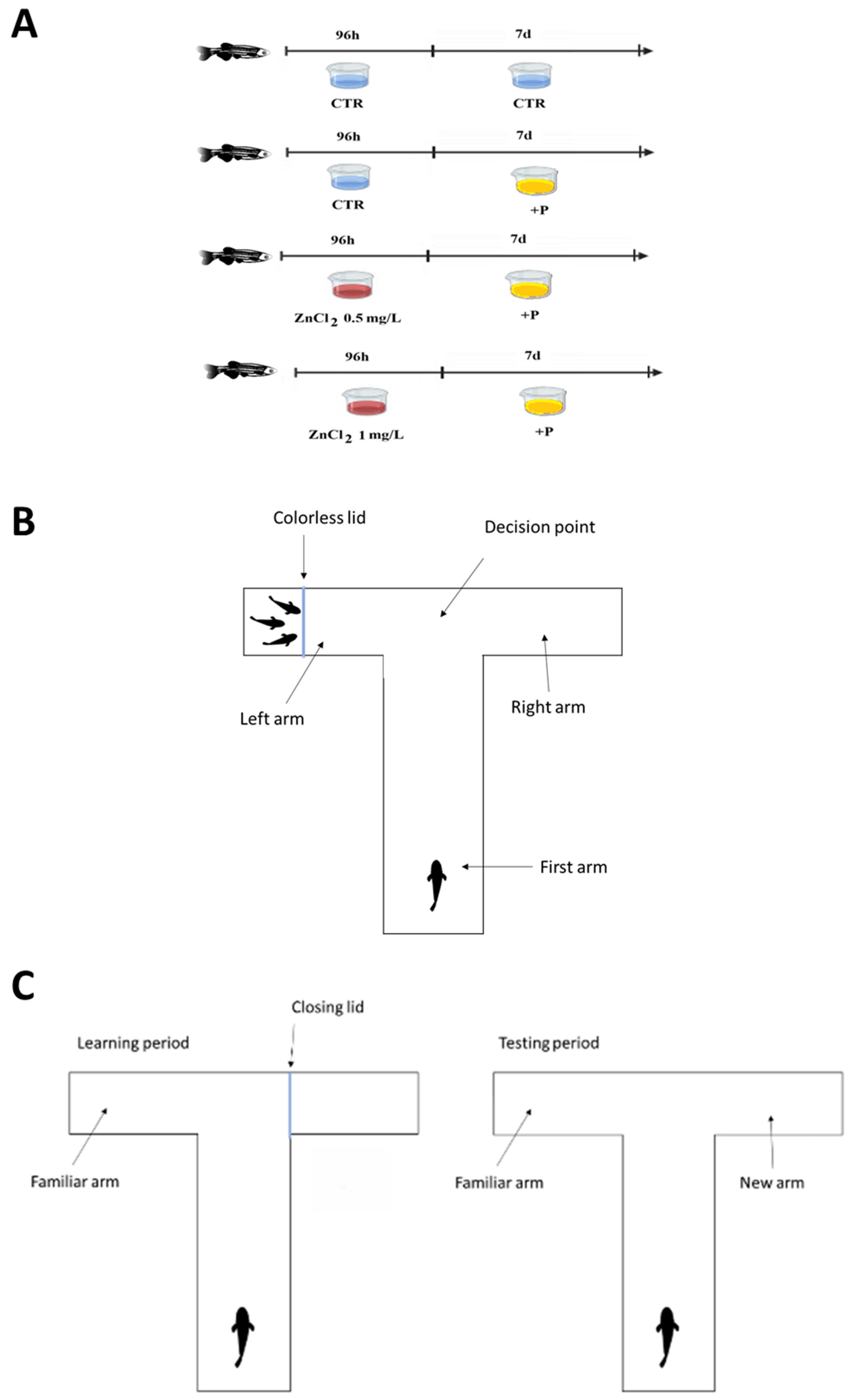
2.4. Experimental Design
2.4.1. Social Preference Test
2.4.2. Short-Term Memory Test
2.4.3. Long-Term Memory Test
2.4.4. Statistical Analysis
3. Results
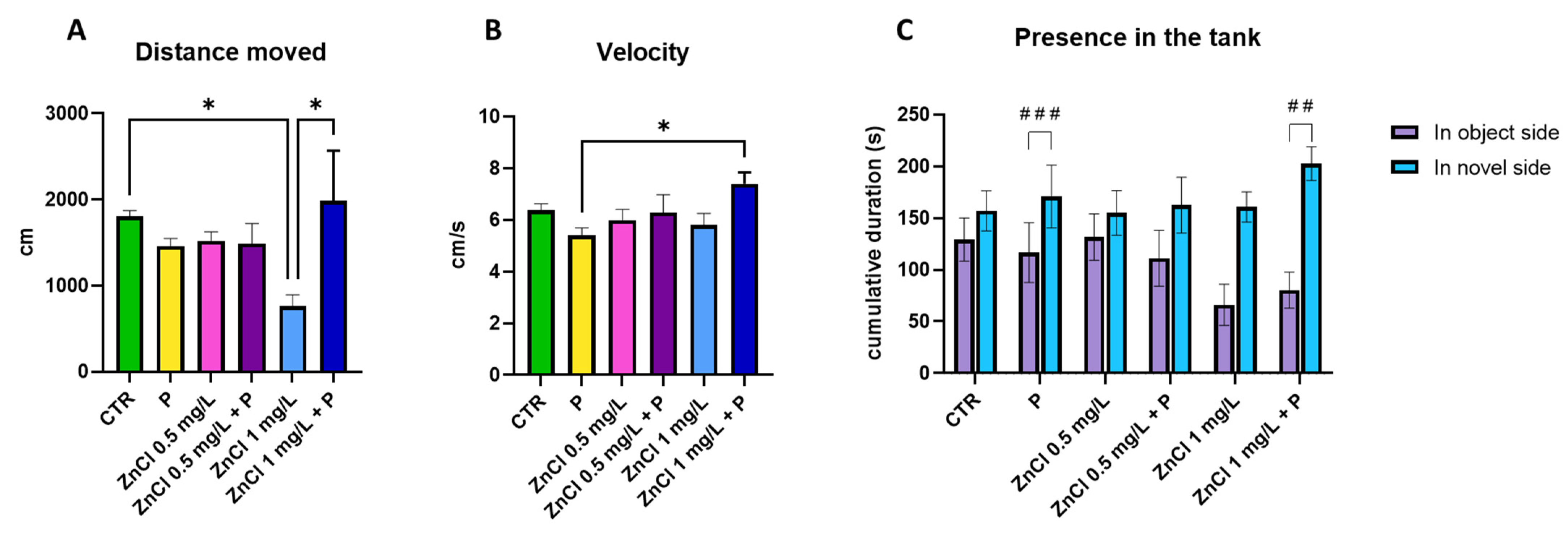
3.1. Social Preference Test
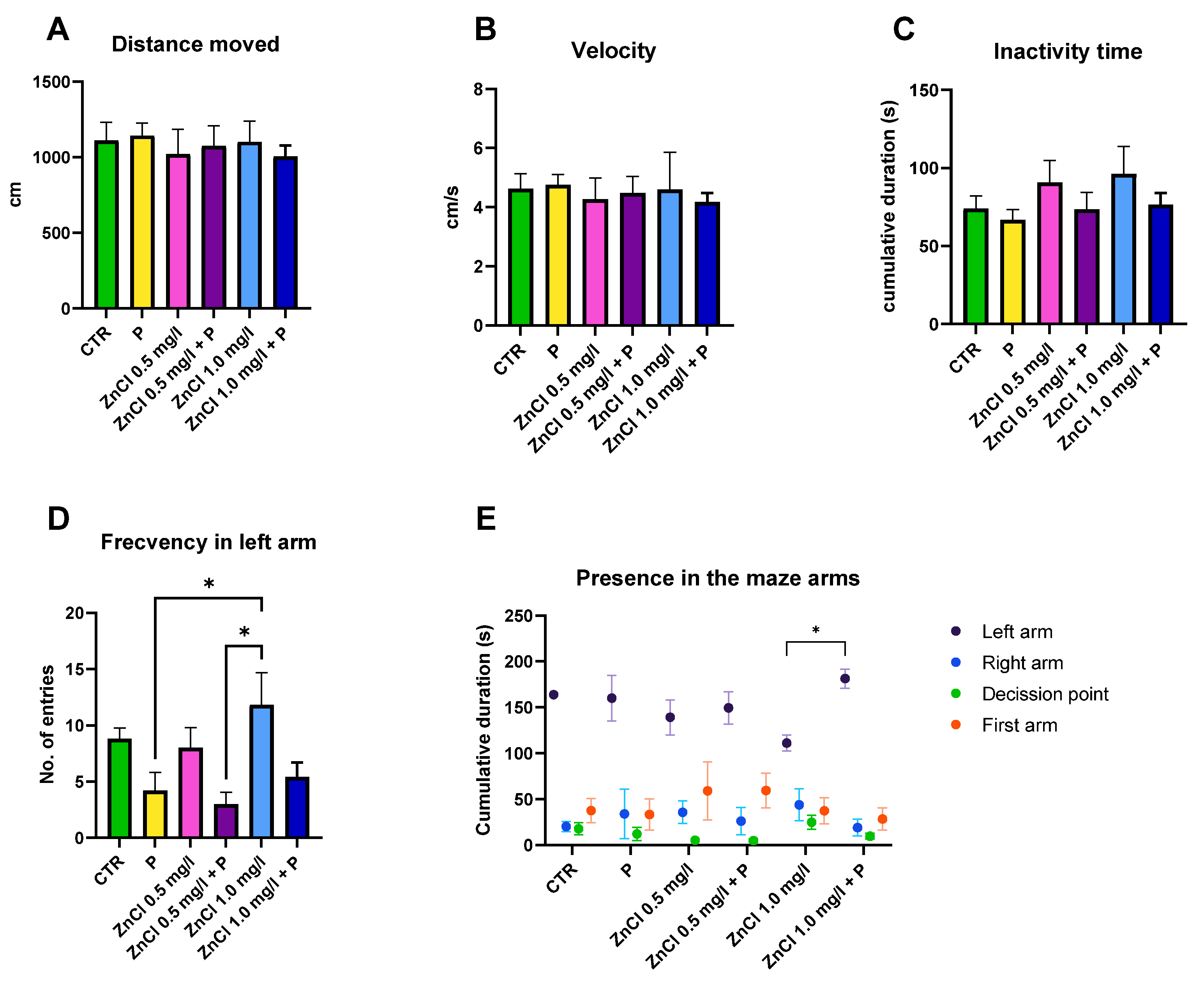
3.2. Short-Term Memory Test
3.3. Long-Term Memory Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.Q. Heavy Metals and Pesticides Toxicity in Agricultural Soil and Plants: Ecological Risks and Human Health Implications. Toxics 2021, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Zeb, M.; Khan, K.; Younas, M.; Farooqi, A.; Cao, X.; Kavil, Y.N.; Alelyani, S.S.; Alkasbi, M.M.; Al-Sehemi, A.G. A review of heavy metals pollution in riverine sediment from various Asian and European countries: Distribution, sources, and environmental risk. Mar. Pollut. Bull. 2024, 206, 116775. [Google Scholar] [CrossRef] [PubMed]
- Iordache, A.M.; Nechita, C.; Voica, C.; Pluhacek, T.; Schug, K.A. Climate change extreme and seasonal toxic metal occurrence in Romanian freshwaters in the last two decades—Case study and critical review. NPJ Clean Water 2022, 5, 2. [Google Scholar] [CrossRef]
- Saravanan, P.; Saravanan, V.; Rajeshkannan, R.; Arnica, G.; Rajasimman, M.; Baskar, G.; Pugazhendhi, A. Comprehensive review on toxic heavy metals in the aquatic system: Sources, identification, treatment strategies, and health risk assessment. Environ. Res. 2024, 258, 119440. [Google Scholar] [CrossRef]
- DHAC: Australian Government Department of Health and Aged Care. Environmental Fate and Effects of Zinc Ions; Draft Evaluation Statement: EVA00143 [Internet]; Australian Government Department of Health and Aged Care: Canberra, Australia, 2024. Available online: https://www.industrialchemicals.gov.au/sites/default/files/2024-04/EVA00143%20-%20Draft%20Evaluation%20Statement%20-%2015%20April%202024.pdf (accessed on 7 January 2025).
- Pakulska, D.; Czerczak, S. Health hazards resulting from exposure to zinc and its inorganic compounds in industry. Med. Pr. 2017, 68, 779–794. [Google Scholar] [CrossRef]
- Seydi, E.; Soltani, M.; Ramazani, M.; Zarei, M.H.; Pourahmad, J. Occupational exposure in lead and zinc mines induces oxidative stress in miners’ lymphocytes: Role of mitochondrial/lysosomal damage. Main Group Met. Chem. 2020, 43, 154–163. [Google Scholar] [CrossRef]
- DCCEEW: Australian Government, Department of Climate Change, Energy, the Environment and Water. Zinc and Compounds. National Pollutant Inventory Fact Sheet. 2022. Available online: https://www.dcceew.gov.au/environment/protection/npi/substances/fact-sheets/zinc-and-compounds (accessed on 7 January 2025).
- Schoofs, H.; Schmit, J.; Rink, L. Zinc Toxicity: Understanding the Limits. Molecules 2024, 29, 3130. [Google Scholar] [CrossRef]
- El Idrissi, A.; van Berkel, L.; Bonekamp, N.E.; Dalemans, D.J.; van der Heyden, M.A. The toxicology of zinc chloride smoke producing bombs and screens. Clin. Toxicol. 2017, 55, 167–174. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, Y.; Zhou, R.; Yang, Y.; Bu, Y. The effect of combined exposure of zinc and nickel on the development of zebrafish. J. Appl. Toxicol. 2021, 41, 1765–1778. [Google Scholar] [CrossRef]
- Sarasamma, S.; Audira, G.; Juniardi, S.; Sampurna, B.P.; Liang, S.T.; Hao, E.; Lai, Y.H.; Hsiao, C.D. Zinc Chloride Exposure Inhibits Brain Acetylcholine Levels, Produces Neurotoxic Signatures, and Diminishes Memory and Motor Activities in Adult Zebrafish. Int. J. Mol. Sci. 2018, 19, 3195. [Google Scholar] [CrossRef]
- Horie, Y.; Yonekura, K.; Suzuki, A.; Takahashi, C. Zinc chloride influences embryonic development, growth, and Gh/Igf-1 gene expression during the early life stage in zebrafish (Danio rerio). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 230, 108684. [Google Scholar] [CrossRef] [PubMed]
- Zixi, Y.; Ruixuan, L.; Shuangshuang, L.; Denggao, Q.; Guanyi, L.; Chun, W.; Jiajia, N.; Yingxue, S.; Hongying, H. Oxidative stress, neurotoxicity, and intestinal microbial regulation after a chronic zinc exposure: An experimental study on adult zebrafish (Danio rerio). Water Reuse 2023, 13, 82–96. [Google Scholar] [CrossRef]
- Toscano, M.; De Grandi, R.; Stronati, L.; De Vecchi, E.; Drago, L. Effect of Lactobacillus rhamnosus HN001 and Bifidobacterium longum BB536 on the healthy gut microbiota composition at phyla and species level: A preliminary study. World J. Gastroenterol. 2017, 23, 2696–2704. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Riasatian, M.; Mazloomi, S.M.; Ahmadi, A.; Derakhshan, Z.; Rajabi, S. Benefits of fermented synbiotic soymilk containing Lactobacillus acidophilus, Bifidobacterium lactis, and inulin towards lead toxicity alleviation. Heliyon 2023, 9, e17518. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gavzy, S.J.; Kensiski, A.; Lee, Z.L.; Mongodin, E.F.; Ma, B.; Bromberg, J.S. Bifidobacterium mechanisms of immune modulation and tolerance. Gut Microbes 2023, 15, 2291164. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Daisley, B.A.; Monachese, M.; Trinder, M.; Bisanz, J.E.; Chmiel, J.A.; Burton, J.P.; Reid, G. Immobilization of cadmium and lead by Lactobacillus rhamnosus GR-1 mitigates apical-to-basolateral heavy metal translocation in a Caco-2 model of the intestinal epithelium. Gut Microbes 2019, 10, 321–333. [Google Scholar] [CrossRef] [PubMed Central]
- Cheng, J.; Laitila, A.; Ouwehand, A.C. Bifidobacterium animalis subsp. lactis HN019 Effects on Gut Health: A Review. Front. Nutr. 2021, 8, 790561. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yang, X.; Li, W.; Ding, M.; Liu, K.J.; Qi, Z.; Zhao, Y. Contribution of zinc accumulation to ischemic brain injury and its mechanisms about oxidative stress, inflammation, and autophagy: An update. Metallomics 2024, 16, mfae012. [Google Scholar] [CrossRef]
- Wang, B.; Fang, T.; Chen, H. Zinc and Central Nervous System Disorders. Nutrients 2023, 15, 2140. [Google Scholar] [CrossRef]
- Porru, S.; Esplugues, A.; Llop, S.; Delgado-Saborit, J.M. The effects of heavy metal exposure on brain and gut microbiota: A systematic review of animal studies. Environ. Pollut. 2024, 348, 123732. [Google Scholar] [CrossRef]
- Loh, J.S.; Mak, W.Q.; Tan, L.K.S.; Ng, C.X.; Chan, H.H.; Yeow, S.H.; Foo, J.B.; Ong, Y.S.; How, C.W.; Khaw, K.Y. Microbiota-gut-brain axis and its therapeutic applications in neurodegenerative diseases. Signal Transduct. Target. Ther. 2024, 9, 37. [Google Scholar] [CrossRef]
- Lin, K.; Peng, F.; He, K.; Qian, Z.; Mei, X.; Su, Z.; Wujimaiti, Y.; Xia, X.; Zhang, T. Research progress on intestinal microbiota regulating cognitive function through the gut-brain axis. Neurol. Sci. 2024, 45, 3711–3721. [Google Scholar] [CrossRef]
- Socala, K.; Doboszewska, U.; Szopa, A.; Serefko, A.; Wlodarczyk, M.; Zielinska, A.; Poleszak, E.; Fichna, J.; Wlaz, P. The role of microbiota-gut-brain axis in neuropsychiatric and neurological disorders. Pharmacol. Res. 2021, 172, 105840. [Google Scholar] [CrossRef] [PubMed]
- Ashaolu, T.J.; Greff, B.; Varga, L. Action and immunomodulatory mechanisms, formulations, and safety concerns of probiotics. Biosci. Microbiota Food Health 2025, 44, 4–15. [Google Scholar] [CrossRef]
- Trisal, A.; Singh, I.; Garg, G.; Jorwal, K.; Singh, A.K. Gut-brain axis and brain health: Modulating neuroinflammation, cognitive decline, and neurodegeneration. 3 Biotech 2025, 15, 25. [Google Scholar] [CrossRef] [PubMed]
- Skalny, A.V.; Aschner, M.; Gritsenko, V.A.; Martins, A.C.; Tizabi, Y.; Korobeinikova, T.V.; Paoliello, M.M.B.; Tinkov, A.A. Modulation of gut microbiota with probiotics as a strategy to counteract endogenous and exogenous neurotoxicity. Adv. Neurotoxicol. 2024, 11, 133–176. [Google Scholar] [CrossRef]
- Kiran, N.S.; Yashaswini, C.; Chatterjee, A. Zebrafish: A trending model for gut-brain axis investigation. Aquat. Toxicol. 2024, 270, 106902. [Google Scholar] [CrossRef] [PubMed]
- Moradian, H.; Gabriel, T.; Barrau, M.; Roblin, X.; Paul, S. New methods to unveil host-microbe interaction mechanisms along the microbiota-gut-brain-axis. Gut Microbes 2024, 16, 2351520. [Google Scholar] [CrossRef]
- Zhong, X.; Li, J.; Lu, F.; Zhang, J.; Guo, L. Application of zebrafish in the study of the gut microbiome. Anim. Model. Exp. Med. 2022, 5, 323–336. [Google Scholar] [CrossRef]
- Firdous, S.M.; Pal, S.; Khanam, S.; Zakir, F. Behavioral neuroscience in zebrafish: Unravelling the complexity of brain-behavior relationships. Naunyn Schmiedebergs Arch. Pharmacol. 2024, 397, 9295–9313. [Google Scholar] [CrossRef]
- Duruibe, J.O.; Ogwuegbu, M.O.C.; Egwurugwu, J.N. Heavy metal pollution and human biotoxic effects. Int. J. Phys. Sci. 2007, 2, 112–118. [Google Scholar]
- Newman, M.; Ebrahimie, E.; Lardelli, M. Using the zebrafish model for Alzheimer’s disease research. Front. Genet. 2014, 5, 189. [Google Scholar] [CrossRef]
- Bautista-Gallego, J.; Rodríguez-Gómez, F.; Romero-Gil, V.; Benítez-Cabello, A.; Arroyo-López, F.N.; Garrido-Fernández, A. Reduction of the Bitter Taste in Packaged Natural Black Manzanilla Olives by Zinc Chloride. Front. Nutr. 2018, 5, 102. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Zhao, C.; Yang, M.; Yin, D. ZnCl2 treatment improves nutrient quality and Zn accumulation in peanut seeds and sprouts. Sci. Rep. 2020, 10, 2364. [Google Scholar] [CrossRef] [PubMed]
- CFR-Code of Federal Regulations-Title 21. Available online: https://www.ecfr.gov/current/title-21 (accessed on 18 February 2025).
- Bauer, K.C.; Huus, K.E.; Finlay, B.B. Microbes and the mind: Emerging hallmarks of the gut microbiota–brain axis. Cell Microbiol. 2016, 18, 632–644. [Google Scholar] [CrossRef]
- Zhu, X.; Li, B.; Lou, P.; Dai, T.; Chen, Y.; Zhuge, A.; Yuan, Y.; Li, L. The Relationship Between the Gut Microbiome and Neurodegenerative Diseases. Neurosci. Bull. 2021, 37, 1510–1522. [Google Scholar] [CrossRef]
- Everard, A.; Cani, P.D. Gut microbiota and GLP-1. Rev. Endocr. Metab. Disord. 2014, 15, 189–196. [Google Scholar] [CrossRef]
- Araujo, L.D.C.; Furlaneto, F.A.C.; da Silva, L.A.B.; Kapila, Y.L. Use of the Probiotic Bifidobacterium animalis subsp. lactis HN019 in Oral Diseases. Int. J. Mol. Sci. 2022, 23, 9334. [Google Scholar] [CrossRef]
- Pinto-Sanchez, M.I.; Hall, G.B.; Ghajar, K.; Nardelli, A.; Bolino, C.; Lau, J.T.; Martin, F.P.; Cominetti, O.; Welsh, C.; Rieder, A.; et al. Probiotic Bifidobacterium longum NCC3001 Reduces Depression Scores and Alters Brain Activity: A Pilot Study in Patients With Irritable Bowel Syndrome. Gastroenterology 2017, 153, 448–459.e8. [Google Scholar] [CrossRef]
- Capurso, L.M.D. Thirty Years of Lactobacillus rhamnosus GG: A Review. J. Clin. Gastroenterol. 2019, 53, S1–S41. [Google Scholar] [CrossRef]
- Shi, C.; Cheng, M.; Yang, X.; Lu, Y.; Yin, H.; Zeng, Y.; Wang, C. Probiotic Lactobacillus rhamnosus GG Promotes Mouse Gut Microbiota Diversity and T Cell Differentiation. Front. Microbiol. 2020, 11, 607735. [Google Scholar] [CrossRef] [PubMed]
- Catanzaro, R.; Anzalone, M.; Calabrese, F.; Milazzo, M.; Capuana, M.; Italia, A.; Occhipinti, S.; Marotta, F. The gut microbiota and its correlations with the central nervous system disorders. Panminerva Med. 2014, 57, 127–143. [Google Scholar]
- Luczynski, P.; McVey Neufeld, K.A.; Oriach, C.S.; Clarke, G.; Dinan, T.G.; Cryan, J.F. Growing up in a bubble: Using germ-free animals to assess the influence of the gut microbiota on brain and behavior. Int. J. Neuropsychopharmacol. 2016, 19, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Grochowska, M.; Laskus, T.; Radkowski, M. Gut Microbiota in Neurological Disorders. Arch. Immunol. Ther. Exp. 2019, 67, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.; Dwivedi, R.; Bansal, M.; Tripathi, M.; Dada, R. Role of Gut Microbiota in Neurological Disorders and Its Therapeutic Significance. J. Clin. Med. 2023, 12, 1650. [Google Scholar] [CrossRef]
- Yang, T.; Huang, C.; Guo, F.; Zhang, X.; Rostami, A.; Liu, C.; Yu, S.; Wang, L.; Zhang, H.; Hu, C.; et al. Probiotics and the Gut-Brain Axis: A Review. Nutrients 2022, 12, 1650. [Google Scholar] [CrossRef]
- Ilie, O.-D.; Paduraru, E.; Robea, M.-A.; Balmus, I.-M.; Jijie, R.; Nicoara, M.; Ciobica, A.S.; Nita, I.-B.; Dobrin, R.; Doroftei, B. The Possible Role of Bifidobacterium longum BB536 and Lactobacillus rhamnosus HN001 on Locomotor Activity and Oxidative Stress in a Rotenone-Induced Zebrafish Model of Parkinson’s Disease. Oxidative Med. Cell. Longev. 2021, 2021, 9629102. [Google Scholar] [CrossRef]
- Yu, F.; Hou, Z.-S.; Luo, H.-R.; Cui, X.-F.; Xiao, J.; Kim, Y.-B.; Li, J.-L.; Feng, W.-R.; Tang, Y.-K.; Li, H.-X.; et al. Zinc alters behavioral phenotypes, neurotransmitter signatures, and immune homeostasis in male zebrafish (Danio rerio). Sci. Total Environ. 2022, 828, 154099. [Google Scholar] [CrossRef]
- Tan, J.K.; Nazar, F.H.; Makpol, S.; Teoh, S.L. Zebrafish: A Pharmacological Model for Learning and Memory Research. Molecules 2022, 27, 7374. [Google Scholar] [CrossRef]
- May, Z.; Morrill, A.; Holcombe, A.; Johnston, T.; Gallup, J.; Fouad, K.; Hamilton, T.J. Object recognition memory in zebrafish. Behav. Brain Res. 2016, 296, 199–210. [Google Scholar] [CrossRef]
- Oliveira, J.; Silveira, M.; Chacon, D.; Luchiari, A. The Zebrafish World of Colors and Shapes: Preference and Discrimination. Zebrafish 2015, 12, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Zoghi, A.; Khosravi-Darani, K.; Sohrabvandi, S. Surface Binding of Toxins and Heavy Metals by Probiotics. Mini-Rev. Med. Chem. 2014, 14, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Lucon-Xiccato, T.; Dadda, M. Assessing memory in zebrafish using the one-trial test. Behav. Process. 2014, 106, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Baralić, K.; Živančević, K.; Bozic, D.; Đukić-Ćosić, D. Probiotic cultures as a potential protective strategy against the toxicity of environmentally relevant chemicals: State-of-the-art knowledge. Food Chem. Toxicol. 2023, 172, 113582. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, J.; Liang, H.; Ye, L.; Lan, L.; Lu, F.; Wang, Q.; Lei, T.; Yang, X.; Cui, P.; et al. Differences in Alpha Diversity of Gut Microbiota in Neurological Diseases. Front. Neurosci. 2022, 16, 879318. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ene, M.; Savuca, A.; Ciobica, A.-S.; Jijie, R.; Gurzu, I.L.; Hritcu, L.D.; Chelaru, I.-A.; Plavan, G.-I.; Nicoara, M.N.; Gurzu, B. The Neurobehavioral Impact of Zinc Chloride Exposure in Zebrafish: Evaluating Cognitive Deficits and Probiotic Modulation. Toxics 2025, 13, 193. https://doi.org/10.3390/toxics13030193
Ene M, Savuca A, Ciobica A-S, Jijie R, Gurzu IL, Hritcu LD, Chelaru I-A, Plavan G-I, Nicoara MN, Gurzu B. The Neurobehavioral Impact of Zinc Chloride Exposure in Zebrafish: Evaluating Cognitive Deficits and Probiotic Modulation. Toxics. 2025; 13(3):193. https://doi.org/10.3390/toxics13030193
Chicago/Turabian StyleEne, Madalina, Alexandra Savuca, Alin-Stelian Ciobica, Roxana Jijie, Irina Luciana Gurzu, Luminita Diana Hritcu, Ionut-Alexandru Chelaru, Gabriel-Ionut Plavan, Mircea Nicusor Nicoara, and Bogdan Gurzu. 2025. "The Neurobehavioral Impact of Zinc Chloride Exposure in Zebrafish: Evaluating Cognitive Deficits and Probiotic Modulation" Toxics 13, no. 3: 193. https://doi.org/10.3390/toxics13030193
APA StyleEne, M., Savuca, A., Ciobica, A.-S., Jijie, R., Gurzu, I. L., Hritcu, L. D., Chelaru, I.-A., Plavan, G.-I., Nicoara, M. N., & Gurzu, B. (2025). The Neurobehavioral Impact of Zinc Chloride Exposure in Zebrafish: Evaluating Cognitive Deficits and Probiotic Modulation. Toxics, 13(3), 193. https://doi.org/10.3390/toxics13030193