
Lentinan Alleviated PM2.5 Exposure-Induced Epithelial–Mesenchymal Transition in Pulmonary Epithelial Cells by Inhibiting the GARP/TGF-β/Smad Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and PM2.5 Sampling and Preparation
2.2. Cell Proliferation Assay
2.3. Wound Healing Assay (WHA)
2.4. Transwell Assay (TWA)
2.5. Real-Time Polymerase Chain Reaction (RT-PCR)
2.6. Western Blot (WB) Assay
2.7. RNA Interference
2.8. Statistical Analysis
3. Results
3.1. Lentinan Reversed the PM2.5-Induced Decrease in Cell Activity
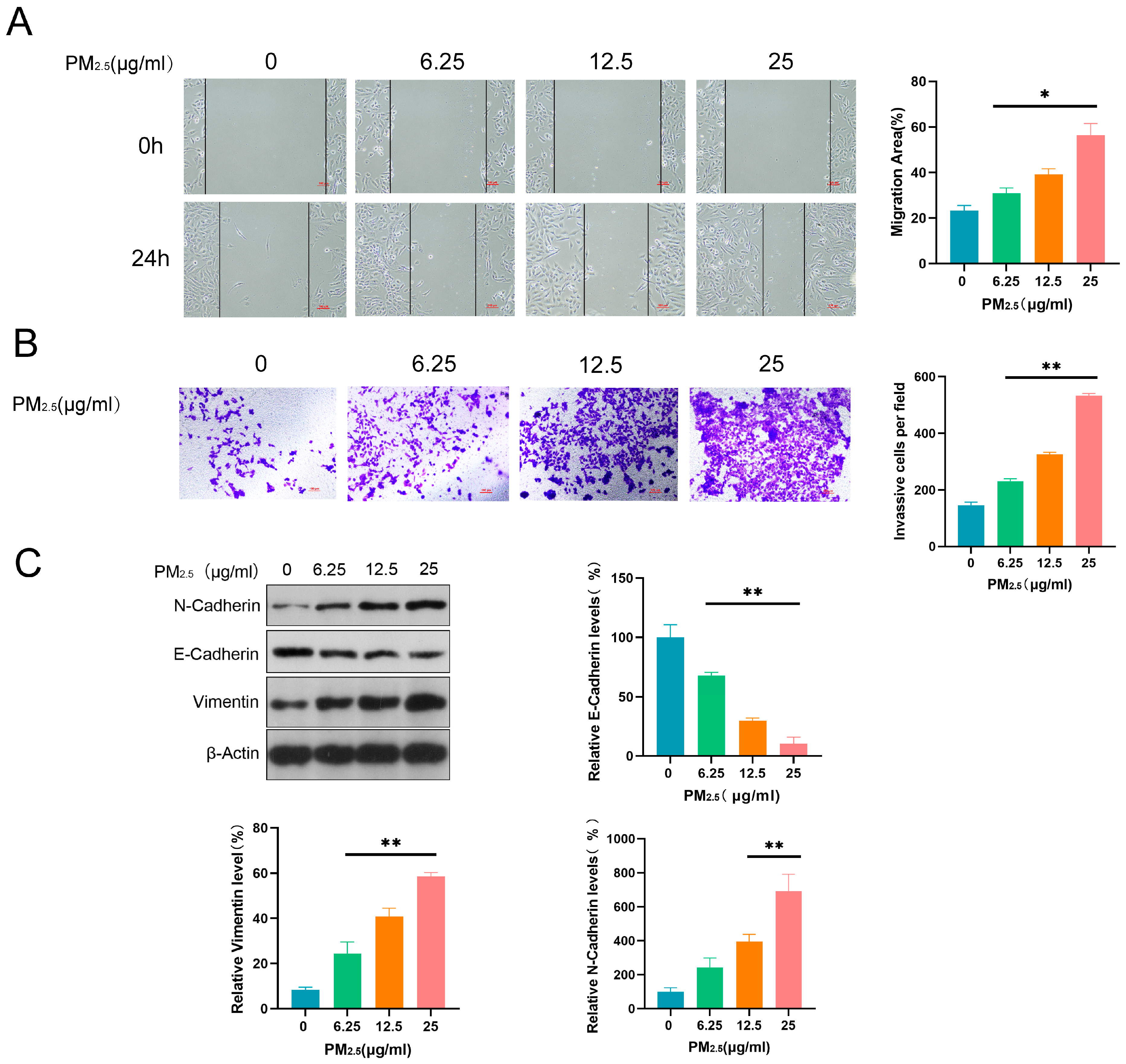
3.2. PM2.5 Promoted Invasive, Migratory, and EMT Induction Abilities in Beas-2B Cells
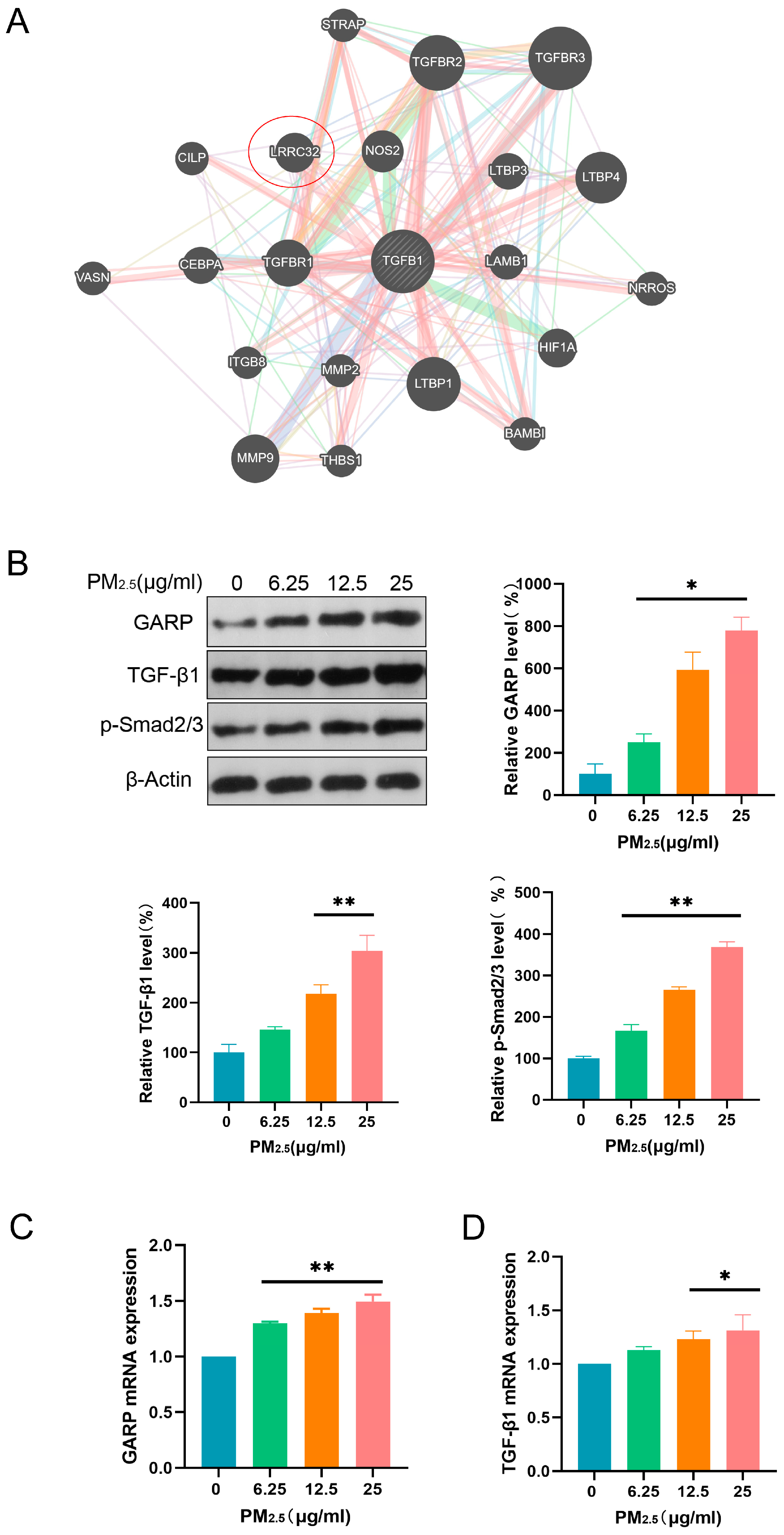
3.3. PM2.5 Activated the GARP/TGF-β/Smad Pathway
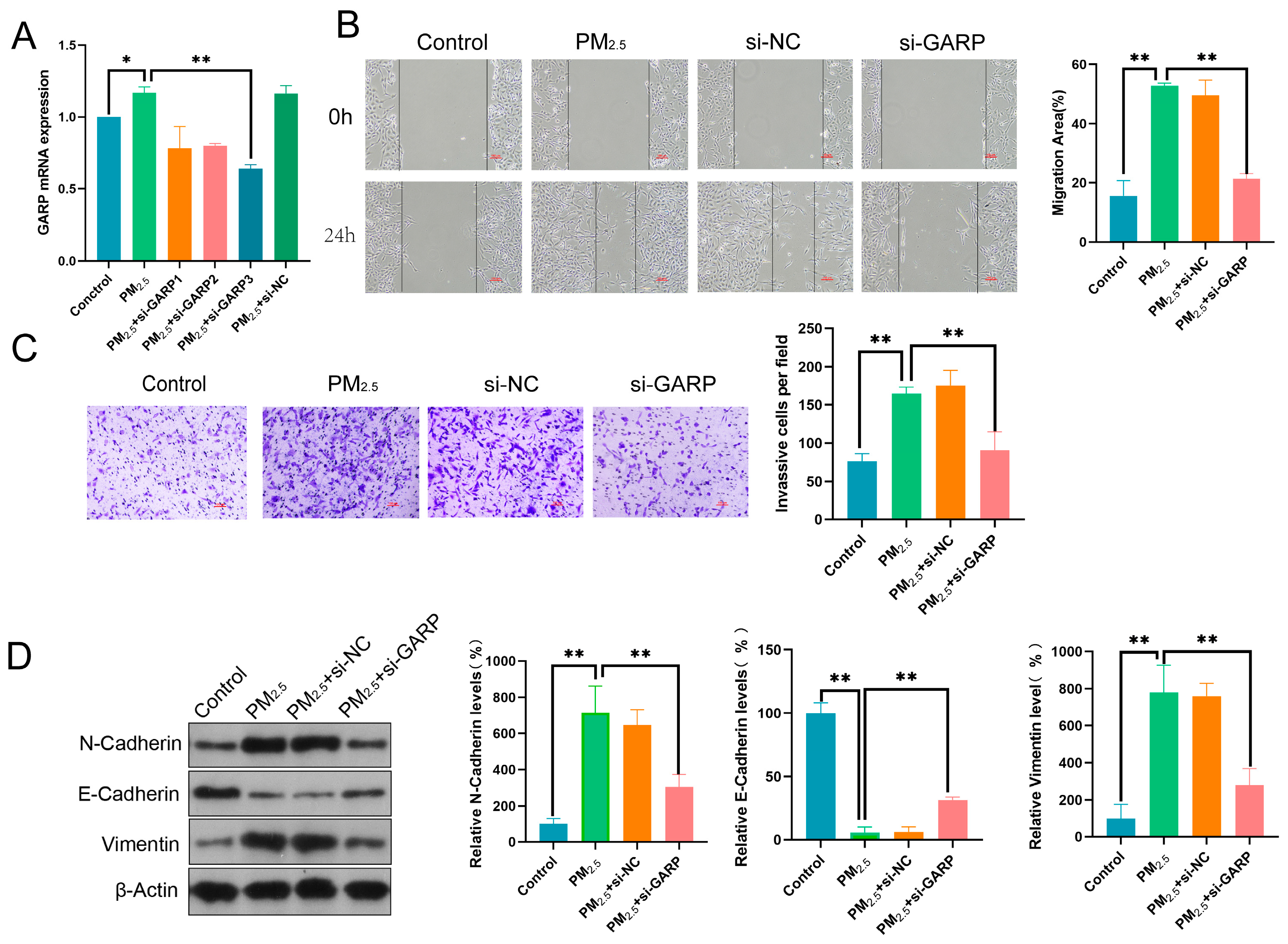
3.4. GARP Knockdown Reversed the Effects of PM2.5 on Beas-2B Cell Migratory, Invasive, and EMT Induction Abilities
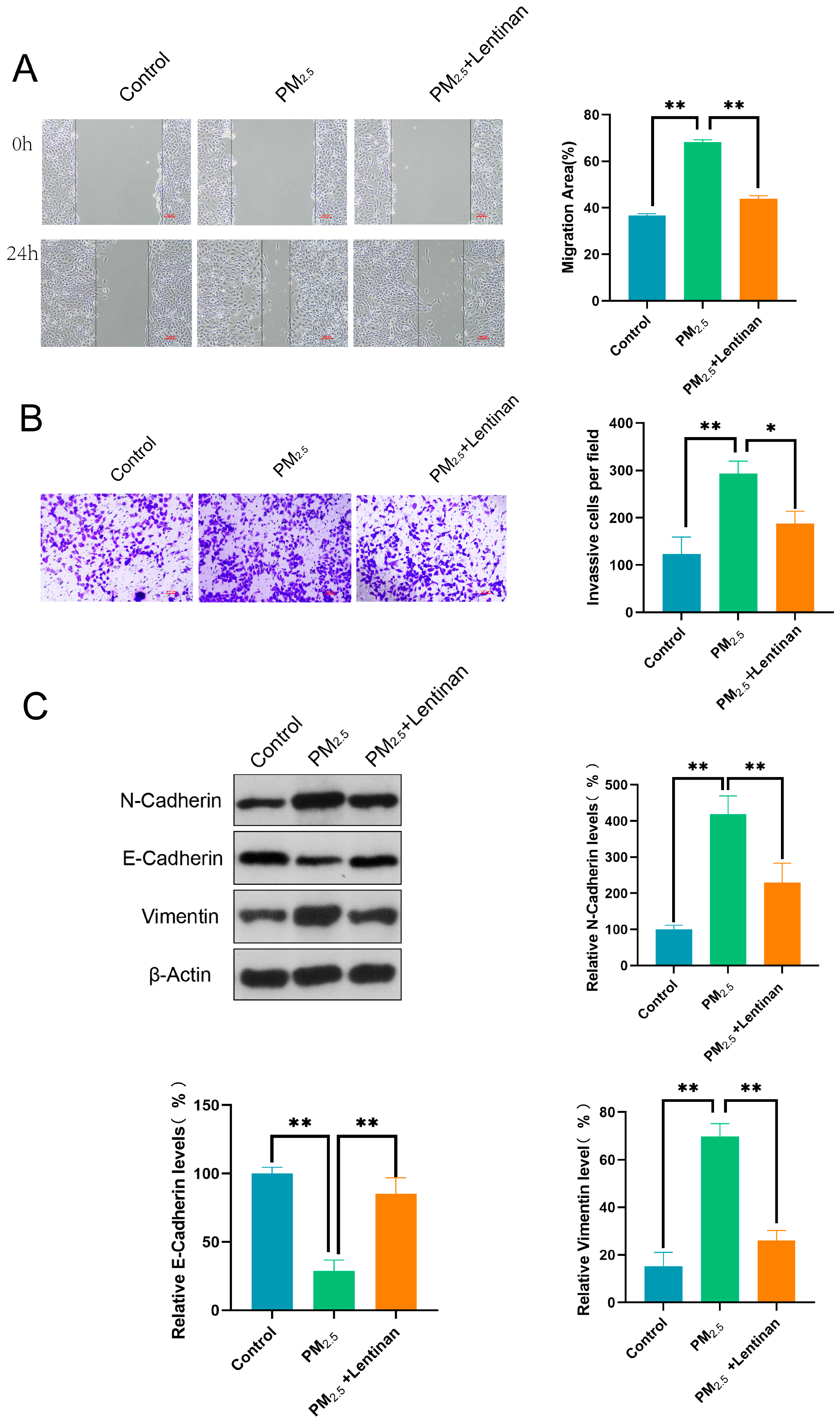
3.5. Lentinan Mitigated the Impact of PM2.5 on Beas-2B Cells
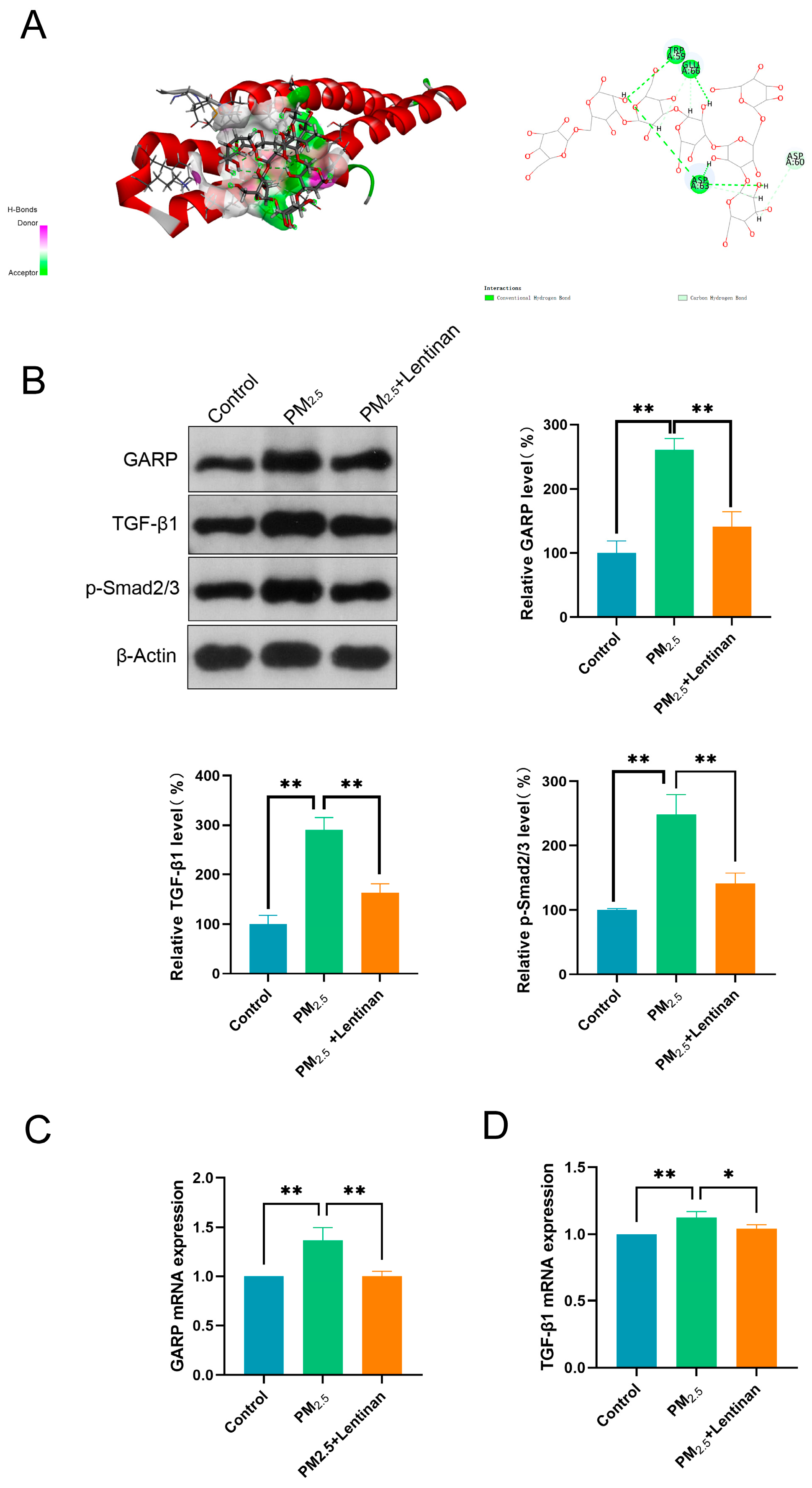
3.6. Lentinan Inhibited the PM2.5-Induced GARP/TGF-β/Smad Pathway Activation
4. Discussion
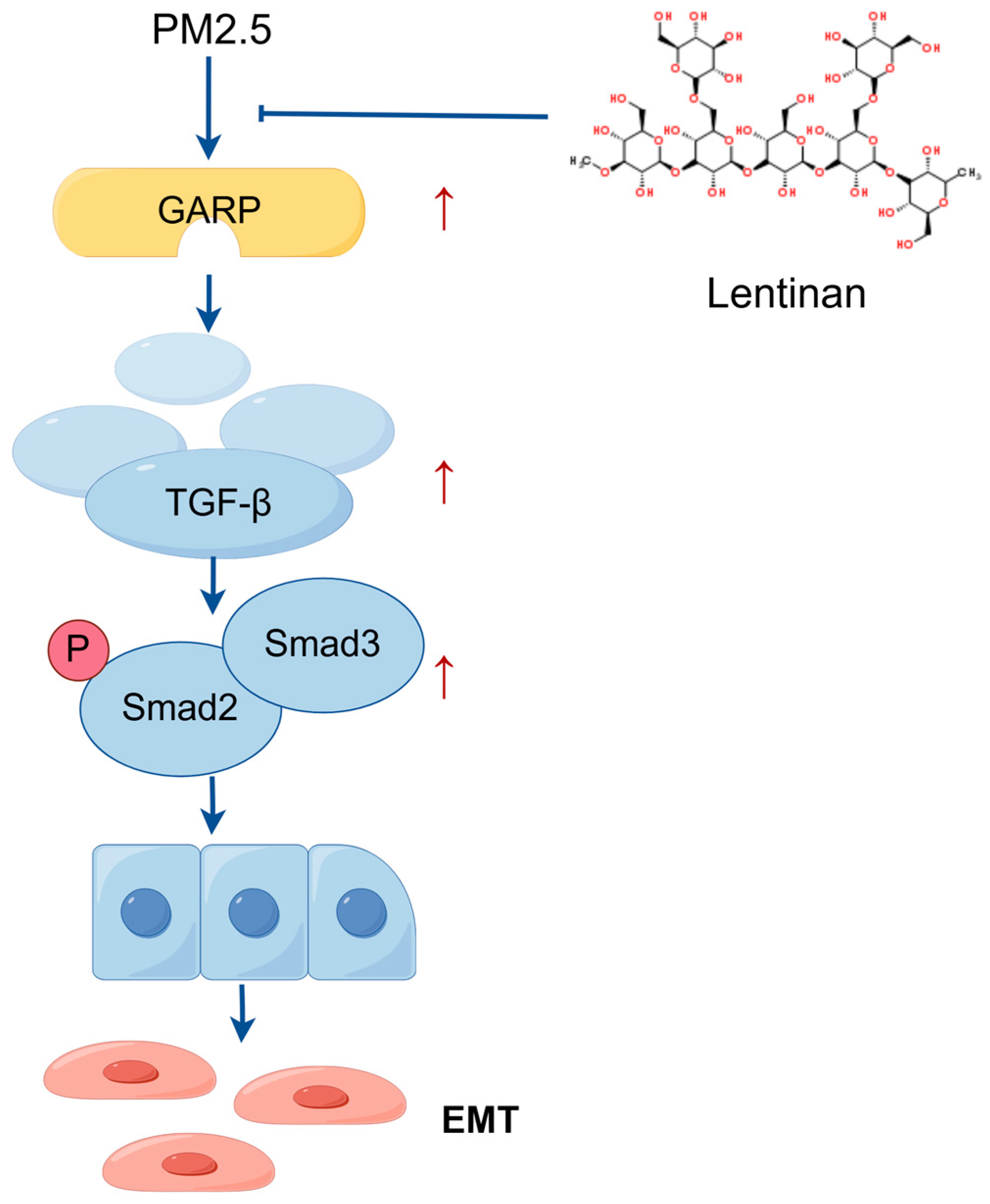
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, J.; Deng, Y.; Tin, M.S.; Lok, V.; Ngai, C.H.; Zhang, L.; Lucero-Prisno, D.E.; Xu, W.; Zheng, Z.-J.; Elcarte, E.; et al. Distribution, Risk Factors, and Temporal Trends for Lung Cancer Incidence and Mortality: A Global Analysis. Chest 2022, 161, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Zaręba, Ł.; Piszczatowska, K.; Dżaman, K.; Soroczynska, K.; Motamedi, P.; Szczepański, M.J.; Ludwig, N. The Relationship between Fine Particle Matter (PM2.5) Exposure and Upper Respiratory Tract Diseases. J. Pers. Med. 2024, 14, 98. [Google Scholar] [CrossRef] [PubMed]
- Behinaein, P.; Hutchings, H.; Knapp, T.; Okereke, I.C. The Growing Impact of Air Quality on Lung-Related Illness: A Narrative Review. J. Thorac. Dis. 2023, 15, 5055–5063. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Xu, A.; Guo, Y.; Bai, Q.; Wu, X.; Ji, S.-P.; Xia, R.-X. PM2.5 Exposure Induces Alveolar Epithelial Cell Apoptosis and Causes Emphysema through P53/Siva-1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3943–3950. [Google Scholar]
- Yue, D.; Zhang, Q.; Zhang, J.; Liu, W.; Chen, L.; Wang, M.; Li, R.; Qin, S.; Song, X.; Ji, Y. Diesel Exhaust PM2.5 Greatly Deteriorates Fibrosis Process in Pre-Existing Pulmonary Fibrosis via Ferroptosis. Environ. Int. 2023, 171, 107706. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, M.; Xiong, Y.; Zhang, L.; Xiong, A.; Wang, J.; He, X.; Li, G. Integrated Analysis of ATAC-Seq and RNA-Seq Unveils the Role of Ferroptosis in PM2.5-Induced Asthma Exacerbation. Int. Immunopharmacol. 2023, 125 Pt B, 111209. [Google Scholar] [CrossRef]
- Obeng, G.M.; Aram, S.A.; Agyei, D.; Saalidong, B.M. Exposure to Particulate Matter (PM2.5) and Volatile Organic Compounds (VOCs), and Self-Reported Health Symptoms among Fish Smokers: A Case Study in the Western Region of Ghana. PLoS ONE 2023, 18, e0283438. [Google Scholar] [CrossRef]
- Wu, C.; Zhang, Y.; Wei, J.; Zhao, Z.; Norbäck, D.; Zhang, X.; Lu, C.; Yu, W.; Wang, T.; Zheng, X.; et al. Associations of Early-Life Exposure to Submicron Particulate Matter With Childhood Asthma and Wheeze in China. JAMA Netw. Open 2022, 5, e2236003. [Google Scholar] [CrossRef]
- Zeng, X.; Xu, X.; Zheng, X.; Reponen, T.; Chen, A.; Huo, X. Heavy Metals in PM2.5 and in Blood, and Children’s Respiratory Symptoms and Asthma from an e-Waste Recycling Area. Environ. Pollut. 2016, 210, 346–353. [Google Scholar] [CrossRef]
- Amnuaylojaroen, T.; Parasin, N. Pathogenesis of PM2.5-Related Disorders in Different Age Groups: Children, Adults, and the Elderly. Epigenomes 2024, 8, 13. [Google Scholar] [CrossRef]
- Firoozi, Z.; Shahi, A.; Mohammadisoleimani, E.; Afzali, S.; Mansoori, B.; Bahmanyar, M.; Mohaghegh, P.; Dastsooz, H.; Pezeshki, B.; Nikfar, G.; et al. CircRNA-Associated ceRNA Networks (circCeNETs) in Chronic Obstructive Pulmonary Disease (COPD). Life Sci. 2024, 349, 122715. [Google Scholar] [CrossRef] [PubMed]
- Cochard, M.; Ledoux, F.; Landkocz, Y. Atmospheric Fine Particulate Matter and Epithelial Mesenchymal Transition in Pulmonary Cells: State of the Art and Critical Review of the in Vitro Studies. J. Toxicol. Environ. Health B Crit. Rev. 2020, 23, 293–318. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Hu, S.; Li, C.; Ma, H.; Wang, Q.; Meng, G.; Guo, T.; Zhang, J. Cigarette Smoke Induced Lung Barrier Dysfunction, EMT, and Tissue Remodeling: A Possible Link between COPD and Lung Cancer. BioMed Res. Int. 2019, 2019, 2025636. [Google Scholar] [CrossRef]
- Wang, Y.; Liao, S.; Pan, Z.; Jiang, S.; Fan, J.; Yu, S.; Xue, L.; Yang, J.; Ma, S.; Liu, T.; et al. Hydrogen Sulfide Alleviates Particulate Matter-Induced Emphysema and Airway Inflammation by Suppressing Ferroptosis. Free Radic. Biol. Med. 2022, 186, 1–16. [Google Scholar] [CrossRef]
- Xu, Z.; Ding, W.; Deng, X. PM2.5, Fine Particulate Matter: A Novel Player in the Epithelial-Mesenchymal Transition? Front. Physiol. 2019, 10, 1404. [Google Scholar] [CrossRef]
- Liu, X.; Xu, D.; Liu, Z.; Li, Y.; Zhang, C.; Gong, Y.; Jiang, Y.; Xing, B. THBS1 Facilitates Colorectal Liver Metastasis through Enhancing Epithelial-Mesenchymal Transition. Clin. Transl. Oncol. 2020, 22, 1730–1740. [Google Scholar] [CrossRef]
- Hu, C.; Xin, Z.; Sun, X.; Hu, Y.; Zhang, C.; Yan, R.; Wang, Y.; Lu, M.; Huang, J.; Du, X.; et al. Activation of ACLY by SEC63 Deploys Metabolic Reprogramming to Facilitate Hepatocellular Carcinoma Metastasis upon Endoplasmic Reticulum Stress. J. Exp. Clin. Cancer Res. 2023, 42, 108. [Google Scholar] [CrossRef]
- Song, Y.; Chen, Y.; Cai, H.; Zhu, G.; Zeng, Y.; Abuduxukuer, Z.; Chen, K.; Wang, J.; Ye, L.; Jin, M. Lentinan Attenuates Allergic Airway Inflammation and Epithelial Barrier Dysfunction in Asthma via Inhibition of the PI3K/AKT/NF-κB Pathway. Phytomedicine 2024, 134, 155965. [Google Scholar] [CrossRef]
- Zhang, Z.; Zha, Z.; Zhao, Z.; Liu, W.; Li, W. Lentinan Inhibits AGE-Induced Inflammation and the Expression of Matrix-Degrading Enzymes in Human Chondrocytes. Drug Des. Dev. Ther. 2020, 14, 2819–2829. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, J.; Zhao, Y.; Zong, S.; Tian, Y.; Chen, S.; Li, M.; Liu, H.; Zhang, Q.; Jing, X.; et al. Therapeutic Effects of Lentinan on Inflammatory Bowel Disease and Colitis-Associated Cancer. J. Cell. Mol. Med. 2019, 23, 750–760. [Google Scholar] [CrossRef]
- Qi, H.; Liu, Y.; Wang, N.; Xiao, C. Lentinan Attenuated the PM2.5 Exposure-Induced Inflammatory Response, Epithelial-Mesenchymal Transition and Migration by Inhibiting the PVT1/miR-199a-5p/Caveolin1 Pathway in Lung Cancer. DNA Cell Biol. 2021, 40, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Metelli, A.; Salem, M.; Wallace, C.H.; Wu, B.X.; Li, A.; Li, X.; Li, Z. Immunoregulatory Functions and the Therapeutic Implications of GARP-TGF-β in Inflammation and Cancer. J. Hematol. Oncol. 2018, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Miyazono, K.; Katsuno, Y.; Koinuma, D.; Ehata, S.; Morikawa, M. Intracellular and Extracellular TGF-β Signaling in Cancer: Some Recent Topics. Front. Med. 2018, 12, 387–411. [Google Scholar] [CrossRef] [PubMed]
- Metelli, A.; Wu, B.X.; Riesenberg, B.; Guglietta, S.; Huck, J.D.; Mills, C.; Li, A.; Rachidi, S.; Krieg, C.; Rubinstein, M.P.; et al. Thrombin Contributes to Cancer Immune Evasion via Proteolysis of Platelet-Bound GARP to Activate LTGF-β. Sci. Transl. Med. 2020, 12, eaay4860. [Google Scholar] [CrossRef]
- Li, A.; Chang, Y.; Song, N.-J.; Wu, X.; Chung, D.; Riesenberg, B.P.; Velegraki, M.; Giuliani, G.D.; Das, K.; Okimoto, T.; et al. Selective Targeting of GARP-LTGFβ Axis in the Tumor Microenvironment Augments PD-1 Blockade via Enhancing CD8+ T Cell Antitumor Immunity. J. Immunother. Cancer 2022, 10, e005433. [Google Scholar] [CrossRef]
- Hou, J.; Wang, X.; Su, C.; Ma, W.; Zheng, X.; Ge, X.; Duan, X. Reduced Frequencies of Foxp3+ GARP+ Regulatory T Cells in COPD Patients Are Associated with Multi-Organ Loss of Tissue Phenotype. Respir. Res. 2022, 23, 176. [Google Scholar] [CrossRef]
- Nookala, S.; Mukundan, S.; Fife, A.; Alagarsamy, J.; Kotb, M. Heterogeneity in FoxP3- and GARP/LAP-Expressing T Regulatory Cells in an HLA Class II Transgenic Murine Model of Necrotizing Soft Tissue Infections by Group A Streptococcus. Infect. Immun. 2018, 86, e00432-18. [Google Scholar] [CrossRef]
- Yang, T.; Gao, Y.; Shi, X.; Zhang, Q.; Ren, Q.; Fu, Y.; Li, H.; Wang, Y.; He, T.; He, M.; et al. Analysis of Atmospheric PM2.5 Composition and Source in an Industrial City in Northern China. Chin. Rare Earths 2023, 44, 113–122. (In Chinese) [Google Scholar]
- Wang, Y.; Zhong, Y.; Hou, T.; Liao, J.; Zhang, C.; Sun, C.; Wang, G. PM2.5 Induces EMT and Promotes CSC Properties by Activating Notch Pathway in Vivo and Vitro. Ecotoxicol. Environ. Saf. 2019, 178, 159–167. [Google Scholar] [CrossRef]
- Wang, Y.; Zhong, Y.; Zhang, C.; Liao, J.; Wang, G. PM2.5 Downregulates MicroRNA-139-5p and Induces EMT in Bronchiolar Epithelium Cells by Targeting Notch1. J. Cancer 2020, 11, 5758–5767. [Google Scholar] [CrossRef]
- Liu, K.; Hua, S.; Song, L. PM2.5 Exposure and Asthma Development: The Key Role of Oxidative Stress. Oxid. Med. Cell. Longev. 2022, 2022, 3618806. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.; Liu, S.; Ye, D.; Bai, G.; Guo, M.; Sun, R.; Ran, P. PM2.5 Induces Lung Inflammation and Fibrosis via Airway Smooth Muscle Cell Expression of the Wnt5a/JNK Pathway. J. Thorac. Dis. 2023, 15, 6094–6105. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Lin, Y.; Zhong, Y.; Guo, X.; Lin, Y.; Yang, S.; Liu, J.; Xie, X.; Sun, Y.; Wang, D.; et al. Impaired AT2 to AT1 Cell Transition in PM2.5-Induced Mouse Model of Chronic Obstructive Pulmonary Disease. Respir. Res. 2022, 23, 70. [Google Scholar] [CrossRef] [PubMed]
- Leilei, L.; Xue, S.; Yan, L.; Yuyuan, L.; Ying, W.; Wenke, Q.; Xuesong, Y.; Ming, L. PM2.5-Exposed Hepatocytes Induce Hepatic Stellate Cells Activation by Releasing TGF-Β1. Biochem. Biophys. Res. Commun. 2021, 569, 125–131. [Google Scholar] [CrossRef]
- Lin, C.-H.; Liu, W.-S.; Wan, C.; Wang, H.-H. Pentraxin 3 Mediates Early Inflammatory Response and EMT Process in Human Tubule Epithelial Cells Induced by PM2.5. Int. Immunopharmacol. 2022, 112, 109258. [Google Scholar] [CrossRef]
- Zhao, C.; Pu, W.; Wazir, J.; Jin, X.; Wei, L.; Song, S.; Su, Z.; Li, J.; Deng, Y.; Wang, H. Long-Term Exposure to PM2.5 Aggravates Pulmonary Fibrosis and Acute Lung Injury by Disrupting Nrf2-Mediated Antioxidant Function. Environ. Pollut. 2022, 313, 120017. [Google Scholar] [CrossRef]
- Wang, H.; Wang, G.; Meng, Y.; Liu, Y.; Yao, X.; Feng, C. Modified Guo-Min Decoction Ameliorates PM2.5-Induced Lung Injury by Inhibition of PI3K-AKT and MAPK Signaling Pathways. Phytomedicine 2024, 123, 155211. [Google Scholar] [CrossRef]
- Chao, X.; Yi, L.; Lan, L.L.; Wei, H.Y.; Wei, D. Long-Term PM2.5 Exposure Increases the Risk of Non-Small Cell Lung Cancer (NSCLC) Progression by Enhancing Interleukin-17a (IL-17a)-Regulated Proliferation and Metastasis. Aging 2020, 12, 11579–11602. [Google Scholar] [CrossRef]
- Hao, Y.; Long, Z.; Gu, X. Farrerol Suppresses Epithelial-Mesenchymal Transition in Hepatocellular Carcinoma via Suppression of TGF-Β1/Smad2/3 Signaling. Pathol. Res. Pract. 2024, 264, 155719. [Google Scholar] [CrossRef]
- Moreau, J.M.; Velegraki, M.; Bolyard, C.; Rosenblum, M.D.; Li, Z. Transforming Growth Factor-Β1 in Regulatory T Cell Biology. Sci. Immunol. 2022, 7, eabi4613. [Google Scholar] [CrossRef]
- Campbell, M.G.; Cormier, A.; Ito, S.; Seed, R.I.; Bondesson, A.J.; Lou, J.; Marks, J.D.; Baron, J.L.; Cheng, Y.; Nishimura, S.L. Cryo-EM Reveals Integrin-Mediated TGF-β Activation without Release from Latent TGF-β. Cell 2020, 180, 490–501.e16. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Y.; Li, Y.; Kou, X.; Xue, Z. Mechanisms of Biochanin A Alleviating PM2.5 Organic Extracts-Induced EMT of A549 Cells through the PI3K/Akt Pathway. J. Nat. Prod. 2022, 85, 2290–2301. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Huo, R.; Wang, Y.; Ma, N.; Shi, X.; Shen, X.; Chang, G. Lentinan Inhibits Oxidative Stress and Alleviates LPS-Induced Inflammation and Apoptosis of BMECs by Activating the Nrf2 Signaling Pathway. Int. J. Biol. Macromol. 2022, 222 Pt B, 2375–2391. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, B.; Zhang, S.; Fan, M.; Ji, X.; Zhang, S.; Wang, Z.; Qiao, K. Lentinan Protects Caenorhabditis Elegans against Fluopyram-Induced Toxicity through DAF-16 and SKN-1 Pathways. Ecotoxicol. Environ. Saf. 2023, 265, 115510. [Google Scholar] [CrossRef]
- Yap, P.G.; Gan, C.Y. Optimized Extraction and Characterization of Ramie Leaf Polysaccharides Using Deep Eutectic Solvent and Microwave: Antioxidant, Metal Chelation, and UV Protection Properties. Int. J. Biol. Macromol. 2024, 282 Pt 3, 136927. [Google Scholar] [CrossRef]
- Yu, L.; Gao, Y.; Ye, Z.; Duan, H.; Zhao, J.; Zhang, H.; Narbad, A.; Tian, F.; Zhai, Q.; Chen, W. Interaction of Beta-Glucans with Gut Microbiota: Dietary Origins, Structures, Degradation, Metabolism, and Beneficial Function. Crit. Rev. Food Sci. Nutr. 2024, 64, 9884–9909. [Google Scholar] [CrossRef]
- Jiang, Y.; Chang, Z.; Xu, Y.; Zhan, X.; Wang, Y.; Gao, M. Advances in Molecular Enzymology of β-1,3-Glucanases: A Comprehensive Review. Int. J. Biol. Macromol. 2024, 279 Pt 3, 135349. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Xu, S.; Bian, B.; Hu, Z.; Wu, F.; Zhao, S.; Wang, X.; Wang, L.; Ma, T. Lentinan Alleviated PM2.5 Exposure-Induced Epithelial–Mesenchymal Transition in Pulmonary Epithelial Cells by Inhibiting the GARP/TGF-β/Smad Pathway. Toxics 2025, 13, 166. https://doi.org/10.3390/toxics13030166
Wang Z, Xu S, Bian B, Hu Z, Wu F, Zhao S, Wang X, Wang L, Ma T. Lentinan Alleviated PM2.5 Exposure-Induced Epithelial–Mesenchymal Transition in Pulmonary Epithelial Cells by Inhibiting the GARP/TGF-β/Smad Pathway. Toxics. 2025; 13(3):166. https://doi.org/10.3390/toxics13030166
Chicago/Turabian StyleWang, Zhi, Shiqing Xu, Bohao Bian, Zhida Hu, Feiyang Wu, Siqi Zhao, Xiaohui Wang, Li Wang, and Teng Ma. 2025. "Lentinan Alleviated PM2.5 Exposure-Induced Epithelial–Mesenchymal Transition in Pulmonary Epithelial Cells by Inhibiting the GARP/TGF-β/Smad Pathway" Toxics 13, no. 3: 166. https://doi.org/10.3390/toxics13030166
APA StyleWang, Z., Xu, S., Bian, B., Hu, Z., Wu, F., Zhao, S., Wang, X., Wang, L., & Ma, T. (2025). Lentinan Alleviated PM2.5 Exposure-Induced Epithelial–Mesenchymal Transition in Pulmonary Epithelial Cells by Inhibiting the GARP/TGF-β/Smad Pathway. Toxics, 13(3), 166. https://doi.org/10.3390/toxics13030166