
Impact of Ultraviolet-Based Combined Disinfection Processes on the Formation and Toxicity of Ciprofloxacin Disinfection Byproducts in Water

Abstract
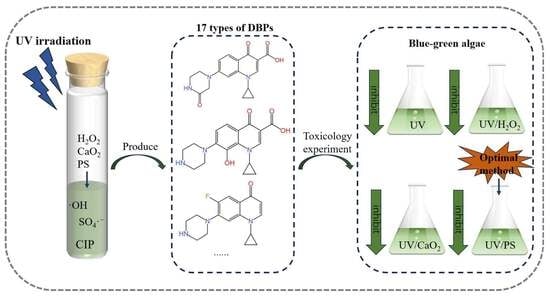
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Procedures
2.2.1. Degradation Kinetics of CIP and Analysis of DBP Formation Under Four Disinfection Systems
2.2.2. Toxicity of CIP and Its DBPs on M. aeruginosa
2.3. Analytical Methods
2.3.1. Determination of CIP Concentration
2.3.2. Determination of DBPs
2.3.3. Measurement of Algal Growth and Physiological/Biochemical Indicators
3. Results and Discussion
3.1. Effects of Different Disinfection Processes on CIP Degradation
3.1.1. Degradation of CIP Under Single UV Irradiation
3.1.2. Degradation of CIP Under the UV/H2O2 System
3.1.3. Degradation of CIP Under the UV/CaO2 System
3.1.4. Degradation of CIP Under the UV/PS System
3.1.5. Degradation of CIP at Different Initial Concentrations
3.2. Formation of DBPs During CIP Degradation
3.3. Effects of CIP and Its DBPs on M. aeruginosa
3.3.1. Effects on Algal Growth
3.3.2. Effects of CIP and DBPs on the Physiological and Biochemical Parameters of M. aeruginosa
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Z.; Chuang, Y.-H.; Szczuka, A.; Ishida, K.P.; Roback, S.; Plumlee, M.H.; Mitch, W.A. Pilot-Scale Evaluation of Oxidant Speciation, 1,4-Dioxane Degradation and Disinfection Byproduct Formation during UV/Hydrogen Peroxide, UV/Free Chlorine and UV/Chloramines Advanced Oxidation Process Treatment for Potable Reuse. Water Res. 2019, 164, 114939. [Google Scholar] [CrossRef]
- Gerrity, D.; Mayer, B.; Ryu, H.; Crittenden, J.; Abbaszadegan, M. A Comparison of Pilot-Scale Photocatalysis and Enhanced Coagulation for Disinfection Byproduct Mitigation. Water Res. 2009, 43, 1597–1610. [Google Scholar] [CrossRef]
- Imafidon, O.; Akpojivi, V.O.; Okosun, J.Z.; Ediagbonya, T.F. Assessment and Mitigation of Disinfection By-Products (DBPs) in Reclaimed Water: Balancing Treatment Efficacy and Environmental/Health Risks. Total Chem. 2025, 1, 100013. [Google Scholar] [CrossRef]
- Vu, T.N.; Kimura, S.Y.; Plewa, M.J.; Richardson, S.D.; Mariñas, B.J. Predominant N-Haloacetamide and Haloacetonitrile Formation in Drinking Water via the Aldehyde Reaction Pathway. Environ. Sci. Technol. 2019, 53, 850–859. [Google Scholar] [CrossRef]
- Liu, X.; Chen, L.; Yang, M.; Tan, C.; Chu, W. The Occurrence, Characteristics, Transformation and Control of Aromatic Disinfection by-Products: A Review. Water Res. 2020, 184, 116076. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-W.; Lin, A.Y.-C. Increased Acute Toxicity to Fish Caused by Pharmaceuticals in Hospital Effluents in a Pharmaceutical Mixture and after Solar Irradiation. Chemosphere 2015, 139, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, Z.; Geng, J.; Song, R.; Liu, X.; Fu, C.; Li, S. Current Advances in UV-Based Advanced Oxidation Processes for the Abatement of Fluoroquinolone Antibiotics in Wastewater. Chin. Chem. Lett. 2025, 36, 110138. [Google Scholar] [CrossRef]
- Nas, B.; Dolu, T.; Koyuncu, S. Behavior and Removal of Ciprofloxacin and Sulfamethoxazole Antibiotics in Three Different Types of Full-Scale Wastewater Treatment Plants: A Comparative Study. Water Air Soil Pollut. 2021, 232, 127. [Google Scholar] [CrossRef]
- Fang, Z.; Lin, Z.; Chen, P.; Feng, M.; Liu, H.; Xiao, Z.; Lin, Z.; Li, D.; Liu, D.; Zhang, Y.; et al. Enhanced Photochemical Degradation and Transformation of Ciprofloxacin in a UV/Calcium Peroxide System: pH Effects, Defluorination Kinetics, and Different Components Numerical Analysis. J. Clean. Prod. 2023, 414, 137706. [Google Scholar] [CrossRef]
- Liang, H.; Zhang, J.; Hu, J.; Li, X.; Li, B. Fluoroquinolone Residues in the Environment Rapidly Induce Heritable Fluoroquinolone Resistance in Escherichia coli. Environ. Sci. Technol. 2023, 57, 4784–4795. [Google Scholar] [CrossRef]
- Lin, Z.; Wu, Y.; Jin, X.; Liang, D.; Jin, Y.; Huang, S.; Wang, Z.; Liu, H.; Chen, P.; Lv, W.; et al. Facile Synthesis of Direct Z-Scheme UiO-66-NH2/PhC2Cu Heterojunction with Ultrahigh Redox Potential for Enhanced Photocatalytic Cr(VI) Reduction and NOR Degradation. J. Hazard. Mater. 2023, 443, 130195. [Google Scholar] [CrossRef]
- Zhao, R.; Wang, Y.; An, Y.; Yang, L.; Sun, Q.; Ma, J.; Zheng, H. Chitin-Biocalcium as a Novel Superior Composite for Ciprofloxacin Removal: Synergism of Adsorption and Flocculation. J. Hazard. Mater. 2022, 423, 126917. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.; Lyu, T.; Zhan, L.; Matamoros, V.; Angelidaki, I.; Cooper, M.; Pan, G. Mitigating Antibiotic Pollution Using Cyanobacteria: Removal Efficiency, Pathways and Metabolism. Water Res. 2021, 190, 116735. [Google Scholar] [CrossRef] [PubMed]
- Heberer, T. Occurrence, Fate, and Removal of Pharmaceutical Residues in the Aquatic Environment: A Review of Recent Research Data. Toxicol Lett. 2002, 131, 5–17. [Google Scholar] [CrossRef]
- Jia, A.; Wan, Y.; Xiao, Y.; Hu, J. Occurrence and Fate of Quinolone and Fluoroquinolone Antibiotics in a Municipal Sewage Treatment Plant. Water Res. 2012, 46, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Verlicchi, P.; Al Aukidy, M.; Zambello, E. Occurrence of Pharmaceutical Compounds in Urban Wastewater: Removal, Mass Load and Environmental Risk after a Secondary Treatment—A Review. Sci. Total. Environ. 2012, 429, 123–155. [Google Scholar] [CrossRef]
- Fang, H.; Oberoi, A.S.; He, Z.; Khanal, S.K.; Lu, H. Ciprofloxacin-Degrading Paraclostridium sp. Isolated from Sulfate-Reducing Bacteria-Enriched Sludge: Optimization and Mechanism. Water Res. 2021, 191, 116808. [Google Scholar] [CrossRef]
- Jia, Y.; Khanal, S.K.; Shu, H.; Zhang, H.; Chen, G.-H.; Lu, H. Ciprofloxacin Degradation in Anaerobic Sulfate-Reducing Bacteria (SRB) Sludge System: Mechanism and Pathways. Water Res. 2018, 136, 64–74. [Google Scholar] [CrossRef]
- Zhang, Q.; Zheng, D.; Bai, B.; Ma, Z.; Zong, S. Insight into Antibiotic Removal by Advanced Oxidation Processes (AOPs): Performance, Mechanism, Degradation Pathways, and Ecotoxicity Assessment. Chem. Eng. J. 2024, 500, 157134. [Google Scholar] [CrossRef]
- Sun, Y.; Jiao, Y.; Wang, M.; Zhao, Y.; Han, Y.; Wu, Q.; Toshiyuki, S.; Wang, C. Effect of Sludge Retention Time on the Removal of Fluoroquinolone Antibiotics in Wastewater Treatment of Activated Sludge. J. Water Process Eng. 2024, 59, 104941. [Google Scholar] [CrossRef]
- Tang, T.; Liu, M.; Du, Y.; Chen, Y. Deciphering the Internal Mechanisms of Ciprofloxacin Affected Anaerobic Digestion, Its Degradation and Detoxification Mechanism. Sci. Total Environ. 2022, 842, 156718. [Google Scholar] [CrossRef] [PubMed]
- Kovalakova, P.; Cizmas, L.; McDonald, T.J.; Marsalek, B.; Feng, M.; Sharma, V.K. Occurrence and Toxicity of Antibiotics in the Aquatic Environment: A Review. Chemosphere 2020, 251, 126351. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Daniels, K.D.; Park, M.; Nienhauser, A.B.; Clevenger, E.C.; Li, Y.; Snyder, S.A. Attenuation of Pharmaceutically Active Compounds in Aqueous Solution by UV/CaO2 Process: Influencing Factors, Degradation Mechanism and Pathways. Water Res. 2019, 164, 114922. [Google Scholar] [CrossRef] [PubMed]
- Straquadine, N.R.W.; Kudela, R.M.; Gobler, C.J. Hepatotoxic Shellfish Poisoning: Accumulation of Microcystins in Eastern Oysters (Crassostrea virginica) and Asian Clams (Corbicula fluminea) Exposed to Wild and Cultured Populations of the Harmful Cyanobacteria, Microcystis. Harmful Algae 2022, 115, 102236. [Google Scholar] [CrossRef]
- Wu, W.; Li, S.; Xie, P.; Li, X.; Chang, H.; Ho, S.-H. Algal-Fungal Interactions and Biomass Production in Wastewater Treatment: Current Status and Future Perspectives. Algal Res. 2023, 70, 103021. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, X.; Chen, L.; Han, Y.; Shen, Y.; Chen, B.; Wang, M. Deglycosylation Inactivation Initiated by a Novel Periplasmic Dehydrogenase Complex Provides a Novel Strategy for Eliminating the Recalcitrant Antibiotic Kanamycin. Environ. Sci. Technol. 2023, 57, 4298–4307. [Google Scholar] [CrossRef]
- Ren, H.; He, F.; Liu, S.; Li, T.; Zhou, R. Enhancing Fenton–like Process at Neutral pH by Fe(III)–GLDA Complexation for the Oxidation Removal of Organic Pollutants. J. Hazard. Mater. 2021, 416, 126077. [Google Scholar] [CrossRef]
- Wang, A.; Zhang, Y.; Zhong, H.; Chen, Y.; Tian, X.; Li, D.; Li, J. Efficient Mineralization of Antibiotic Ciprofloxacin in Acid Aqueous Medium by a Novel Photoelectro-Fenton Process Using a Microwave Discharge Electrodeless Lamp Irradiation. J. Hazard. Mater. 2018, 342, 364–374. [Google Scholar] [CrossRef]
- Karim, A.V.; Shriwastav, A. Degradation of Ciprofloxacin Using Photo, Sono, and Sonophotocatalytic Oxidation with Visible Light and Low-Frequency Ultrasound: Degradation Kinetics and Pathways. Chem. Eng. J. 2020, 392, 124853. [Google Scholar] [CrossRef]
- Niu, B.; Wang, L.; Li, M.; Yao, W.; Zang, K.; Zhou, L.; Hu, X.; Zheng, Y. Lattice B-Doping Evolved Ferromagnetic Perovskite-like Catalyst for Enhancing Persulfate-Based Degradation of Norfloxacin. J. Hazard. Mater. 2022, 425, 127949. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, C.; Jiang, M.; Xiang, Q.; Wang, S.; Su, Y.; Zhang, N.; Si, Z.; Mu, Y.; Yang, R. Dissolved Organic Matter Mitigates the Toxicity of Silver Nanoparticles to Freshwater Algae: Effects on Oxidative Stress and Membrane Function. Environ. Pollut. 2025, 372, 125973. [Google Scholar] [CrossRef] [PubMed]
- Gallo, M.; Morse, D.; Hollnagel, H.C.; Barros, M.P. Oxidative Stress and Toxicology of Cu2+ Based on Surface Areas in Mixed Cultures of Green Alga and Cyanobacteria: The Pivotal Role of H2O2. Aquat. Toxicol. 2020, 222, 105450. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Zhou, C.; Zhu, T.; Shao, K.; Wang, J. Impact of Ultraviolet-Based Combined Disinfection Processes on the Formation and Toxicity of Ciprofloxacin Disinfection Byproducts in Water. Toxics 2025, 13, 995. https://doi.org/10.3390/toxics13110995
Guo Y, Zhou C, Zhu T, Shao K, Wang J. Impact of Ultraviolet-Based Combined Disinfection Processes on the Formation and Toxicity of Ciprofloxacin Disinfection Byproducts in Water. Toxics. 2025; 13(11):995. https://doi.org/10.3390/toxics13110995
Chicago/Turabian StyleGuo, Yang, Chengyu Zhou, Tao Zhu, Kangle Shao, and Junhao Wang. 2025. "Impact of Ultraviolet-Based Combined Disinfection Processes on the Formation and Toxicity of Ciprofloxacin Disinfection Byproducts in Water" Toxics 13, no. 11: 995. https://doi.org/10.3390/toxics13110995
APA StyleGuo, Y., Zhou, C., Zhu, T., Shao, K., & Wang, J. (2025). Impact of Ultraviolet-Based Combined Disinfection Processes on the Formation and Toxicity of Ciprofloxacin Disinfection Byproducts in Water. Toxics, 13(11), 995. https://doi.org/10.3390/toxics13110995