


Methylomic, Proteomic, and Metabolomic Correlates of Traffic-Related Air Pollution in the Context of Cardiorespiratory Health: A Systematic Review, Pathway Analysis, and Network Analysis

 and
and 
Abstract
1. Introduction
2. Materials and Methods
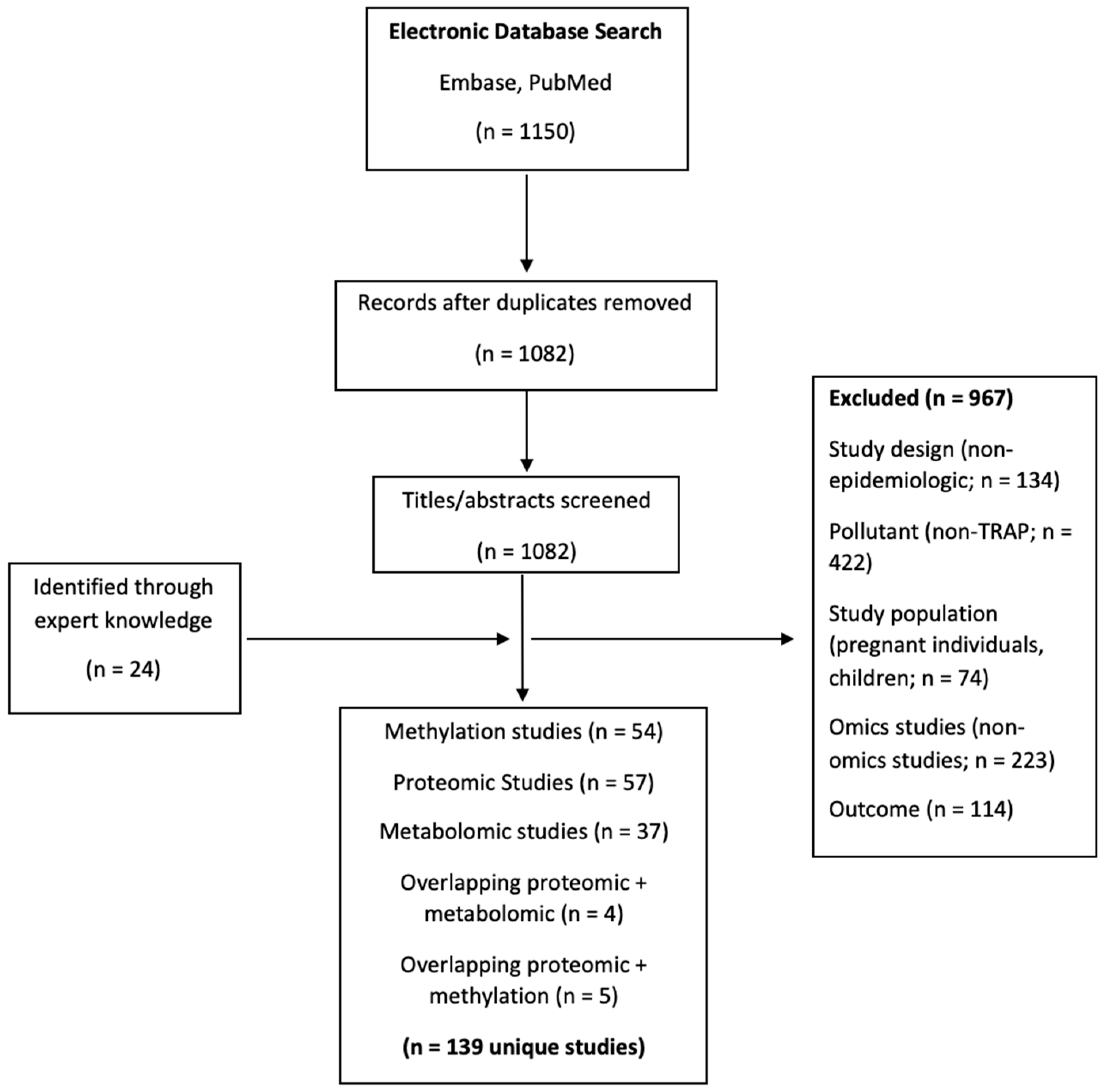
2.1. Search Strategy and Study Selection
2.2. Data Extraction and Organization
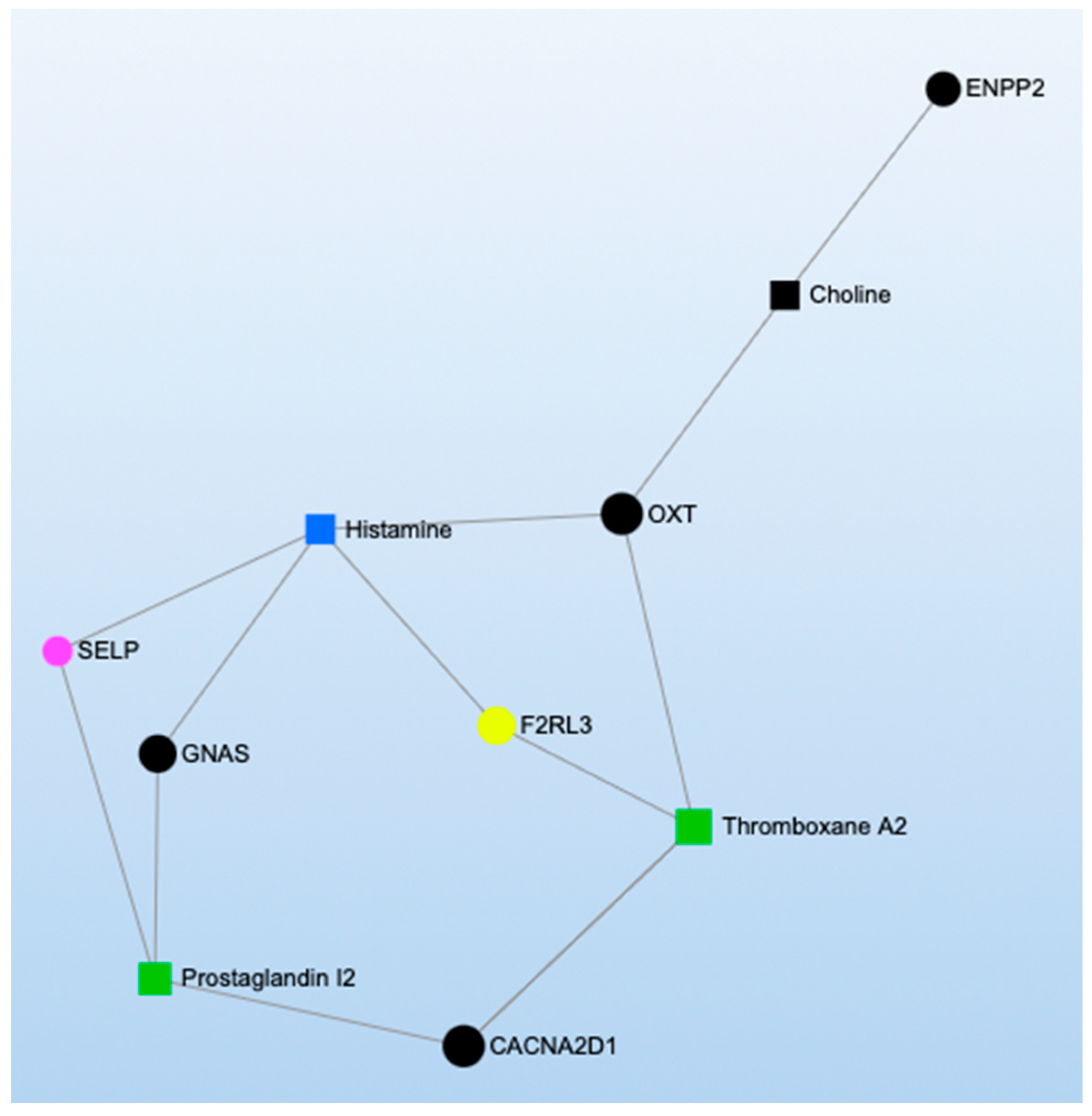
2.3. Pathway and Network Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Omics Type | Study Design | Exposure Assessment | Exposure Window | Study Populations a | Country | Sample Size | Sex Distribution | Omics Approach |
|---|---|---|---|---|---|---|---|---|
| Methylomics n = 54 studies | Cross-sectional: 29 Panel: 9 Cohort: 5 Cross-over: 9 Quasi-experimental: 2 | Fixed site measurement: 16 Spatiotemporal model: 21 Personal measurement: 12 Controlled exposure: 5 | Short-term: 29 Long-term: 25 | NAS: 10 [41,42,43,44,45,46,47,48,49,50] KORA: 3 [45,49,51] WHI: 3 [52,53,54] ARIC: 3 [52,53,54] EPIC-Italy: 2 [55,56] MESA: 2 [57,58] Sister Study: 2 [59,60] BAPE: 2 [61,62] Taiwan Biobank: 2 [63,64] REGICOR: 1 [55] EPIC-Netherlands: 1 [56] Lifelines: 1 [51] EXPOsOMICS: 1 [65] SAPALDIA: 1 [66] Lothian Birth Cohort: 1 [67] SPHERE: 1 [68] | USA: 17 China: 15 Italy: 8 Canada: 4 Netherlands: 3 Taiwan: 3 Germany: 2 Switzerland: 2 UK: 2 Belgium: 2 Spain: 1 South Korea: 1 Czech Republic: 1 | <50: 20 50–99: 3 100–1000: 20 >1000: 11 | 100% female: 4 100% male: 11 Other: 39 | Candidate gene: 26 Epigenome-wide association study: 24 Global methylation: 4 |
| Proteomics n = 57 studies | Cross-sectional: 28 Panel: 8 Cohort: 3 Cross-over: 10 Quasi-experimental: Case-control: 3 | Fixed site measurement: 24 Spatiotemporal mode: 19 Personal measurement: 9 Biomarker: 2 Controlled exposure: 4 | Short-term: 36 Long-term: 21 | NAS: 3 [69,70,71] SWAN: 3 [72,73,74] KORA: 3 [75,76,77] Heinz–Nixdorf Recall: 3 [75,78,79] Framingham Offspring: 2 [80,81] AIRCHD: 2 [82,83] EPIC-Italy: 1 [84] BPRHS: 1 [85] Malmo Diet and Cancer: 1 [86] AHAB-II: 1 [87] SAGE: 1 [88] Nurse’s Health Study: 1 [89] ELISABET: 1 [90] ESCAPE: 1 [91] SAPALDIA: 1 [75] FINRISK: 1 [75] TwinGene: 1 [75] MESA: 1 [92] CAFEH: 1 [93] CoLaus: 1 [94] | USA: 17 China: 17 Canada: 6 Germany: 4 India: 3 Taiwan: 3 Italy: 2 Sweden: 1 UK: 1 France: 1 Brazil: 1 Sweden: 1 Finland: 1 Switzerland: 1 | <50: 15 50–99: 10 100–1000: 13 >1000: 19 | 100% female: 3 100% male: 6 Other: 48 | Targeted: 54 Untargeted: 3 |
| Metabolomics n = 37 studies | Cross-sectional: 15 Panel: 7 Cohort: 2 Cross-over: 7 Natural Experiment: 1 | Fixed site measurement: 8 Spatiotemporal model: 10 Personal measurement: 14 Biomarker: 1 Controlled exposure: 4 | Short-term: 26 Long-term: 11 | DRIVE: 3 [95,96,97] NAS: 2 [98,99] Children’s Health Study: 2 [100,101] KORA: 2 [102,103] SAPALDIA: 1 [104] EPIC-Italy: 1 [104] ACE: 1 [105] ACE-2: 1 [106] Oxford St. 2: 1 [13] TAPAS II: 1 [13] CAFEH: 1 [107] EARTH: 1 [108] AIRCHD: 1 [83] SCOPE: 1 [109] TwinsUK: 1 [110] | USA: 17 China: 12 Germany: 2 UK: 2 Sweden: 1 Switzerland: 1 Italy: 1 India: 1 Spain: 1 Netherlands: 1 Brazil: 1 | <50: 15 50–99: 6 100–1000: 7 >1000: 4 | 100% female: 1 100% male: 5 Other: 31 | Targeted: 8 Untargeted: 29 |
3. Results and Discussion
3.1. Overview of the Literature
3.1.1. TRAP Exposure Assessment
3.1.2. Study Populations
3.1.3. Biological Matrices
3.1.4. Omics Assessment
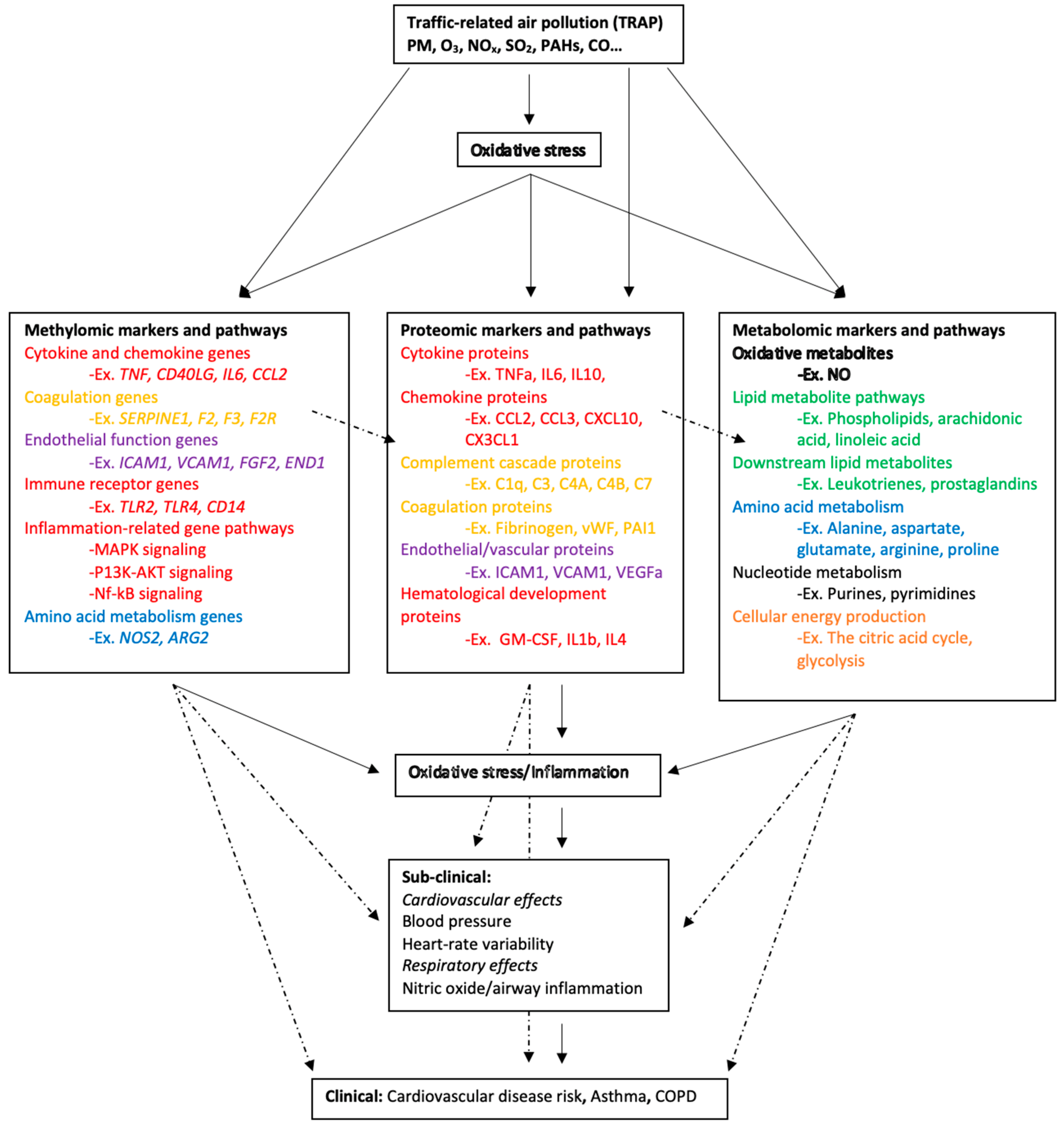
3.2. Omics Markers and Associated Biological Pathways
3.2.1. Lipid Metabolism
3.2.2. Cellular Energy Production
3.2.3. Amino Acid Metabolism
3.2.4. Inflammation and Immunity
3.2.5. Coagulation
3.2.6. Endothelial Function
3.2.7. Oxidative Stress
3.2.8. TRAP, Omics, and Respiratory Disease
3.2.9. TRAP, Omics, and CVD
4. Conclusions
4.1. Strengths and Limitations
4.2. Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brook, R.D.; Rajagopalan, S.; Pope, C.A.; Brook, J.R.; Bhatnagar, A.; Diez-Roux, A.V.; Holguin, F.; Hong, Y.; Luepker, R.V.; Mittleman, M.A.; et al. Particulate Matter Air Pollution and Cardiovascular Disease. Circulation 2010, 121, 2331–2378. [Google Scholar] [CrossRef]
- Hoek, G.; Krishnan, R.M.; Beelen, R.; Peters, A.; Ostro, B.; Brunekreef, B.; Kaufman, J.D. Long-Term Air Pollution Exposure and Cardio-Respiratory Mortality: A Review. Environ. Health 2013, 12, 43. [Google Scholar] [CrossRef] [PubMed]
- Künzli, N.; Kaiser, R.; Medina, S.; Studnicka, M.; Chanel, O.; Filliger, P.; Herry, M.; Horak, F.; Puybonnieux-Texier, V.; Quénel, P.; et al. Public-Health Impact of Outdoor and Traffic-Related Air Pollution: A European Assessment. Lancet 2000, 356, 795–801. [Google Scholar] [CrossRef]
- Anderson, J.O.; Thundiyil, J.G.; Stolbach, A. Clearing the Air: A Review of the Effects of Particulate Matter Air Pollution on Human Health. J. Med. Toxicol. 2012, 8, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Bourdrel, T.; Bind, M.-A.; Béjot, Y.; Morel, O.; Argacha, J.-F. Cardiovascular Effects of Air Pollution. Arch. Cardiovasc. Dis. 2017, 110, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Brucker, N.; do Nascimento, S.N.; Bernardini, L.; Charão, M.F.; Garcia, S.C. Biomarkers of Exposure, Effect, and Susceptibility in Occupational Exposure to Traffic-Related Air Pollution: A Review. J. Appl. Toxicol. 2020, 40, 722–736. [Google Scholar] [CrossRef]
- Laumbach, R.J.; Kipen, H.M. Acute Effects of Motor Vehicle Traffic-Related Air Pollution Exposures on Measures of Oxidative Stress in Human Airways. Ann. N. Y. Acad. Sci. 2010, 1203, 107–112. [Google Scholar] [CrossRef]
- Miller, M.R. Oxidative Stress and the Cardiovascular Effects of Air Pollution. Free Radic. Biol. Med. 2020, 151, 69–87. [Google Scholar] [CrossRef]
- Rajagopalan, S.; Al-Kindi, S.G.; Brook, R.D. Air Pollution and Cardiovascular Disease. J. Am. Coll. Cardiol. 2018, 72, 2054–2070. [Google Scholar] [CrossRef]
- Ferrari, L.; Carugno, M.; Bollati, V. Particulate Matter Exposure Shapes DNA Methylation through the Lifespan. Clin. Epigenet. 2019, 11, 129. [Google Scholar] [CrossRef]
- Rider, C.F.; Carlsten, C. Air Pollution and DNA Methylation: Effects of Exposure in Humans. Clin. Epigenet. 2019, 11, 131. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.; Bunkar, N.; Kumar, R.; Bhargava, A.; Tiwari, R.; Chaudhury, K.; Goryacheva, I.Y.; Mishra, P.K. Air Pollution Associated Epigenetic Modifications: Transgenerational Inheritance and Underlying Molecular Mechanisms. Sci. Total Environ. 2019, 656, 760–777. [Google Scholar] [CrossRef] [PubMed]
- van Veldhoven, K.; Kiss, A.; Keski-Rahkonen, P.; Robinot, N.; Scalbert, A.; Cullinan, P.; Chung, K.F.; Collins, P.; Sinharay, R.; Barratt, B.M.; et al. Impact of Short-Term Traffic-Related Air Pollution on the Metabolome—Results from Two Metabolome-Wide Experimental Studies. Environ. Int. 2019, 123, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Vlaanderen, J.J.; Janssen, N.A.; Hoek, G.; Keski-Rahkonen, P.; Barupal, D.K.; Cassee, F.R.; Gosens, I.; Strak, M.; Steenhof, M.; Lan, Q.; et al. The Impact of Ambient Air Pollution on the Human Blood Metabolome. Environ. Res. 2017, 156, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Hou, X.-Y.; Wei, Y.; Thai, P.; Chai, F. Biomarkers of the Health Outcomes Associated with Ambient Particulate Matter Exposure. Sci. Total Environ. 2017, 579, 1446–1459. [Google Scholar] [CrossRef] [PubMed]
- Leon-Mimila, P.; Wang, J.; Huertas-Vazquez, A. Relevance of Multi-Omics Studies in Cardiovascular Diseases. Front. Cardiovasc. Med. 2019, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Mars, R.A.T.; Yang, Y.; Ward, T.; Houtti, M.; Priya, S.; Lekatz, H.R.; Tang, X.; Sun, Z.; Kalari, K.R.; Korem, T.; et al. Longitudinal Multi-Omics Reveals Subset-Specific Mechanisms Underlying Irritable Bowel Syndrome. Cell 2020, 182, 1460–1473. [Google Scholar] [CrossRef]
- Schüssler-Fiorenza Rose, S.M.; Contrepois, K.; Moneghetti, K.J.; Zhou, W.; Mishra, T.; Mataraso, S.; Dagan-Rosenfeld, O.; Ganz, A.B.; Dunn, J.; Hornburg, D.; et al. A Longitudinal Big Data Approach for Precision Health. Nat. Med. 2019, 25, 792–804. [Google Scholar] [CrossRef]
- Beale, D.J.; Karpe, A.V.; Ahmed, W. Beyond Metabolomics: A Review of Multi-Omics-Based Approaches. In Microbial Metabolomics: Applications in Clinical, Environmental, and Industrial Microbiology; Beale, D.J., Kouremenos, K.A., Palombo, E.A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 289–312. ISBN 978-3-319-46326-1. [Google Scholar]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-Omics Approaches to Disease. Genome Biol. 2017, 18, 83. [Google Scholar] [CrossRef]
- Chu, S.H.; Huang, M.; Kelly, R.S.; Benedetti, E.; Siddiqui, J.K.; Zeleznik, O.A.; Pereira, A.; Herrington, D.; Wheelock, C.E.; Krumsiek, J.; et al. Integration of Metabolomic and Other Omics Data in Population-Based Study Designs: An Epidemiological Perspective. Metabolites 2019, 9, 117. [Google Scholar] [CrossRef]
- Everson, T.M.; Marsit, C.J. Integrating-Omics Approaches into Human Population-Based Studies of Prenatal and Early-Life Exposures. Curr. Environ. Health Rep. 2018, 5, 328–337. [Google Scholar] [CrossRef]
- Subramanian, I.; Verma, S.; Kumar, S.; Jere, A.; Anamika, K. Multi-Omics Data Integration, Interpretation, and Its Application. Bioinform. Biol. Insights 2020, 14, 1177932219899051. [Google Scholar] [CrossRef]
- Kellogg, R.A.; Dunn, J.; Snyder, M.P. Personal Omics for Precision Health. Circ. Res. 2018, 122, 1169–1171. [Google Scholar] [CrossRef]
- Olivier, M.; Asmis, R.; Hawkins, G.A.; Howard, T.D.; Cox, L.A. The Need for Multi-Omics Biomarker Signatures in Precision Medicine. Int. J. Mol. Sci. 2019, 20, 4781. [Google Scholar] [CrossRef] [PubMed]
- Riggs, D.W.; Yeager, R.A.; Bhatnagar, A. Defining the Human Envirome. Circ. Res. 2018, 122, 1259–1275. [Google Scholar] [CrossRef] [PubMed]
- Savaryn, J.P.; Catherman, A.D.; Thomas, P.M.; Abecassis, M.M.; Kelleher, N.L. The Emergence of Top-down Proteomics in Clinical Research. Genome Med. 2013, 5, 53. [Google Scholar] [CrossRef] [PubMed]
- Dedeurwaerder, S.; Defrance, M.; Calonne, E.; Denis, H.; Sotiriou, C.; Fuks, F. Evaluation of the Infinium Methylation 450K Technology. Epigenomics 2011, 3, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New Perspectives on Genomes, Pathways, Diseases and Drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A Web Server for Metabolomic Data Analysis and Interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef] [PubMed]
- Soma-Pillay, P.; Nelson-Piercy, C.; Tolppanen, H.; Mebazaa, A. Physiological Changes in Pregnancy. CVJA 2016, 27, 89–94. [Google Scholar] [CrossRef]
- Hill, D.P.; Smith, B.; McAndrews-Hill, M.S.; Blake, J.A. Gene Ontology Annotations: What They Mean and Where They Come From. BMC Bioinform. 2008, 9, S2. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a Reference Resource for Gene and Protein Annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed]
- Aoki-Kinoshita, K.F. Overview of KEGG Applications to Omics-Related Research. J. Pestic. Sci. 2006, 31, 296–299. [Google Scholar] [CrossRef][Green Version]
- Zahn-Zabal, M.; Michel, P.-A.; Gateau, A.; Nikitin, F.; Schaeffer, M.; Audot, E.; Gaudet, P.; Duek, P.D.; Teixeira, D.; Rech de Laval, V.; et al. The neXtProt Knowledgebase in 2020: Data, Tools and Usability Improvements. Nucleic Acids Res. 2020, 48, D328–D334. [Google Scholar] [CrossRef]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, M.; Jassal, B.; Stephan, R.; Milacic, M.; Rothfels, K.; Senff-Ribeiro, A.; Griss, J.; Sevilla, C.; Matthews, L.; Gong, C.; et al. The Reactome Pathway Knowledgebase 2022. Nucleic Acids Res. 2022, 50, D687–D692. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Pang, Z.; Xia, J. Comprehensive Investigation of Pathway Enrichment Methods for Functional Interpretation of LC–MS Global Metabolomics Data. Brief. Bioinform. 2023, 24, bbac553. [Google Scholar] [CrossRef]
- Jiang, D.; Armour, C.R.; Hu, C.; Mei, M.; Tian, C.; Sharpton, T.J.; Jiang, Y. Microbiome Multi-Omics Network Analysis: Statistical Considerations, Limitations, and Opportunities. Front. Genet. 2019, 10, 995. [Google Scholar] [CrossRef]
- Zhou, G.; Pang, Z.; Lu, Y.; Ewald, J.; Xia, J. OmicsNet 2.0: A Web-Based Platform for Multi-Omics Integration and Network Visual Analytics. Nucleic Acids Res. 2022, 50, W527–W533. [Google Scholar] [CrossRef]
- Bind, M.-A.; Lepeule, J.; Zanobetti, A.; Gasparrini, A.; Baccarelli, A.A.; Coull, B.A.; Tarantini, L.; Vokonas, P.S.; Koutrakis, P.; Schwartz, J. Air Pollution and Gene-Specific Methylation in the Normative Aging Study: Association, Effect Modification, and Mediation Analysis. Epigenetics 2014, 9, 448–458. [Google Scholar] [CrossRef]
- Dai, L.; Mehta, A.; Mordukhovich, I.; Just, A.C.; Shen, J.; Hou, L.; Koutrakis, P.; Sparrow, D.; Vokonas, P.S.; Baccarelli, A.A.; et al. Differential DNA Methylation and PM2.5 Species in a 450K Epigenome-Wide Association Study. Epigenetics 2017, 12, 139–148. [Google Scholar] [CrossRef]
- Wang, C.; Cardenas, A.; Hutchinson, J.N.; Just, A.; Heiss, J.; Hou, L.; Zheng, Y.; Coull, B.A.; Kosheleva, A.; Koutrakis, P.; et al. Short- and Intermediate-Term Exposure to Ambient Fine Particulate Elements and Leukocyte Epigenome-Wide DNA Methylation in Older Men: The Normative Aging Study. Environ. Int. 2022, 158, 106955. [Google Scholar] [CrossRef]
- Wang, C.; Koutrakis, P.; Gao, X.; Baccarelli, A.; Schwartz, J. Associations of Annual Ambient PM2.5 Components with DNAm PhenoAge Acceleration in Elderly Men: The Normative Aging Study. Environ. Pollut. 2020, 258, 113690. [Google Scholar] [CrossRef] [PubMed]
- Ward-Caviness, C.K.; Nwanaji-Enwerem, J.C.; Wolf, K.; Wahl, S.; Colicino, E.; Trevisi, L.; Kloog, I.; Just, A.C.; Vokonas, P.; Cyrys, J.; et al. Long-Term Exposure to Air Pollution Is Associated with Biological Aging. Oncotarget 2016, 7, 74510–74525. [Google Scholar] [CrossRef] [PubMed]
- Carmona, J.J.; Sofer, T.; Hutchinson, J.; Cantone, L.; Coull, B.; Maity, A.; Vokonas, P.; Lin, X.; Schwartz, J.; Baccarelli, A.A. Short-Term Airborne Particulate Matter Exposure Alters the Epigenetic Landscape of Human Genes Associated with the Mitogen-Activated Protein Kinase Network: A Cross-Sectional Study. Environ. Health 2014, 13, 94. [Google Scholar] [CrossRef] [PubMed]
- Nwanaji-Enwerem, J.C.; Bind, M.-A.; Dai, L.; Oulhote, Y.; Colicino, E.; Di, Q.; Just, A.C.; Hou, L.; Vokonas, P.; Coull, B.A.; et al. Editor’s Highlight: Modifying Role of Endothelial Function Gene Variants on the Association of Long-Term PM2.5 Exposure with Blood DNA Methylation Age: The VA Normative Aging Study. Toxicol. Sci. 2017, 158, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Nwanaji-Enwerem, J.C.; Dai, L.; Colicino, E.; Oulhote, Y.; Di, Q.; Kloog, I.; Just, A.C.; Hou, L.; Vokonas, P.; Baccarelli, A.A.; et al. Associations between Long-Term Exposure to PM2.5 Component Species and Blood DNA Methylation Age in the Elderly: The VA Normative Aging Study. Environ. Int. 2017, 102, 57–65. [Google Scholar] [CrossRef]
- Panni, T.; Mehta, A.J.; Schwartz, J.D.; Baccarelli, A.A.; Just, A.C.; Wolf, K.; Wahl, S.; Cyrys, J.; Kunze, S.; Strauch, K.; et al. Genome-Wide Analysis of DNA Methylation and Fine Particulate Matter Air Pollution in Three Study Populations: KORA F3, KORA F4, and the Normative Aging Study. Environ. Health Perspect. 2016, 124, 983–990. [Google Scholar] [CrossRef]
- Madrigano, J.; Baccarelli, A.; Mittleman, M.A.; Wright, R.O.; Sparrow, D.; Vokonas, P.S.; Tarantini, L.; Schwartz, J. Prolonged Exposure to Particulate Pollution, Genes Associated with Glutathione Pathways, and DNA Methylation in a Cohort of Older Men. Environ. Health Perspect. 2011, 119, 977–982. [Google Scholar] [CrossRef]
- De F.C. Lichtenfels, A.J.; Van Der Plaat, D.A.; De Jong, K.; Van Diemen, C.C.; Postma, D.S.; Nedeljkovic, I.; Van Duijn, C.M.; Amin, N.; La Bastide-van Gemert, S.; De Vries, M.; et al. Long-Term Air Pollution Exposure, Genome-Wide DNA Methylation and Lung Function in the LifeLines Cohort Study. Environ. Health Perspect. 2018, 126, 027004. [Google Scholar] [CrossRef]
- Gondalia, R.; Baldassari, A.; Holliday, K.M.; Justice, A.E.; Méndez-Giráldez, R.; Stewart, J.D.; Liao, D.; Yanosky, J.D.; Brennan, K.J.M.; Engel, S.M.; et al. Methylome-Wide Association Study Provides Evidence of Particulate Matter Air Pollution-Associated DNA Methylation. Environ. Int. 2019, 132, 104723. [Google Scholar] [CrossRef] [PubMed]
- Holliday, K.M.; Gondalia, R.; Baldassari, A.; Justice, A.E.; Stewart, J.D.; Liao, D.; Yanosky, J.D.; Jordahl, K.M.; Bhatti, P.; Assimes, T.L.; et al. Gaseous Air Pollutants and DNA Methylation in a Methylome-Wide Association Study of an Ethnically and Environmentally Diverse Population of U.S. Adults. Environ. Res. 2022, 212, 113360. [Google Scholar] [CrossRef] [PubMed]
- Gondalia, R.; Baldassari, A.; Holliday, K.M.; Justice, A.E.; Stewart, J.D.; Liao, D.; Yanosky, J.D.; Engel, S.M.; Sheps, D.; Jordahl, K.M.; et al. Epigenetically Mediated Electrocardiographic Manifestations of Sub-Chronic Exposures to Ambient Particulate Matter Air Pollution in the Women’s Health Initiative and Atherosclerosis Risk in Communities Study. Environ. Res. 2021, 198, 111211. [Google Scholar] [CrossRef]
- Sayols-Baixeras, S.; Fernández-Sanlés, A.; Prats-Uribe, A.; Subirana, I.; Plusquin, M.; Künzli, N.; Marrugat, J.; Basagaña, X.; Elosua, R. Association between Long-Term Air Pollution Exposure and DNA Methylation: The REGICOR Study. Environ. Res. 2019, 176, 108550. [Google Scholar] [CrossRef] [PubMed]
- Plusquin, M.; Guida, F.; Polidoro, S.; Vermeulen, R.; Raaschou-Nielsen, O.; Campanella, G.; Hoek, G.; Kyrtopoulos, S.A.; Georgiadis, P.; Naccarati, A.; et al. DNA Methylation and Exposure to Ambient Air Pollution in Two Prospective Cohorts. Environ. Int. 2017, 108, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Chi, G.C.; Liu, Y.; MacDonald, J.W.; Reynolds, L.M.; Enquobahrie, D.A.; Fitzpatrick, A.L.; Kerr, K.F.; Budoff, M.J.; Lee, S.-I.; Siscovick, D.; et al. Epigenome-Wide Analysis of Long-Term Air Pollution Exposure and DNA Methylation in Monocytes: Results from the Multi-Ethnic Study of Atherosclerosis. Epigenetics 2022, 17, 1900028. [Google Scholar] [CrossRef]
- Chi, G.C.; Liu, Y.; MacDonald, J.W.; Barr, R.G.; Donohue, K.M.; Hensley, M.D.; Hou, L.; McCall, C.E.; Reynolds, L.M.; Siscovick, D.S.; et al. Long-Term Outdoor Air Pollution and DNA Methylation in Circulating Monocytes: Results from the Multi-Ethnic Study of Atherosclerosis (MESA). Environ. Health 2016, 15, 119. [Google Scholar] [CrossRef]
- White, A.J.; Kresovich, J.K.; Keller, J.P.; Xu, Z.; Kaufman, J.D.; Weinberg, C.R.; Taylor, J.A.; Sandler, D.P. Air Pollution, Particulate Matter Composition and Methylation-Based Biologic Age. Environ. Int. 2019, 132, 105071. [Google Scholar] [CrossRef]
- Wang, C.; O’Brien, K.M.; Xu, Z.; Sandler, D.P.; Taylor, J.A.; Weinberg, C.R. Long-Term Ambient Fine Particulate Matter and DNA Methylation in Inflammation Pathways: Results from the Sister Study. Epigenetics 2020, 15, 524–535. [Google Scholar] [CrossRef]
- Fang, J.; Gao, Y.; Zhang, M.; Jiang, Q.; Chen, C.; Gao, X.; Liu, Y.; Dong, H.; Tang, S.; Li, T.; et al. Personal PM2.5 Elemental Components, Decline of Lung Function, and the Role of DNA Methylation on Inflammation-Related Genes in Older Adults: Results and Implications of the BAPE Study. Environ. Sci. Technol. 2022, 56, 15990–16000. [Google Scholar] [CrossRef]
- Shi, W.; Tang, S.; Fang, J.; Cao, Y.; Chen, C.; Li, T.; Gao, X.; Shi, X. Epigenetic Age Stratifies the Risk of Blood Pressure Elevation Related to Short-Term PM2.5 Exposure in Older Adults. Environ. Res. 2022, 212, 113507. [Google Scholar] [CrossRef]
- Tantoh, D.M.; Lee, K.-J.; Nfor, O.N.; Liaw, Y.-C.; Lin, C.; Chu, H.-W.; Chen, P.-H.; Hsu, S.-Y.; Liu, W.-H.; Ho, C.-C.; et al. Methylation at Cg05575921 of a Smoking-Related Gene (AHRR) in Non-Smoking Taiwanese Adults Residing in Areas with Different PM2.5 Concentrations. Clin. Epigenet. 2019, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Tantoh, D.M.; Wu, M.-C.; Chuang, C.-C.; Chen, P.-H.; Tyan, Y.S.; Nfor, O.N.; Lu, W.-Y.; Liaw, Y.-P. AHRR Cg05575921 Methylation in Relation to Smoking and PM2.5 Exposure among Taiwanese Men and Women. Clin. Epigenet. 2020, 12, 117. [Google Scholar] [CrossRef] [PubMed]
- Mostafavi, N.; Vermeulen, R.; Ghantous, A.; Hoek, G.; Probst-Hensch, N.; Herceg, Z.; Tarallo, S.; Naccarati, A.; Kleinjans, J.C.S.; Imboden, M.; et al. Acute Changes in DNA Methylation in Relation to 24 h Personal Air Pollution Exposure Measurements: A Panel Study in Four European Countries. Environ. Int. 2018, 120, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Eze, I.C.; Jeong, A.; Schaffner, E.; Rezwan, F.I.; Ghantous, A.; Foraster, M.; Vienneau, D.; Kronenberg, F.; Herceg, Z.; Vineis, P.; et al. Genome-Wide DNA Methylation in Peripheral Blood and Long-Term Exposure to Source-Specific Transportation Noise and Air Pollution: The SAPALDIA Study. Environ. Health Perspect. 2020, 128, 067003. [Google Scholar] [CrossRef] [PubMed]
- Baranyi, G.; Deary, I.J.; McCartney, D.L.; Harris, S.E.; Shortt, N.; Reis, S.; Russ, T.C.; Ward Thompson, C.; Vieno, M.; Cox, S.R.; et al. Life-Course Exposure to Air Pollution and Biological Ageing in the Lothian Birth Cohort 1936. Environ. Int. 2022, 169, 107501. [Google Scholar] [CrossRef]
- Monti, P.; Iodice, S.; Tarantini, L.; Sacchi, F.; Ferrari, L.; Ruscica, M.; Buoli, M.; Vigna, L.; Pesatori, A.C.; Bollati, V. Effects of PM Exposure on the Methylation of Clock Genes in A Population of Subjects with Overweight or Obesity. Int. J. Environ. Res. Public Health 2021, 18, 1122. [Google Scholar] [CrossRef]
- Bind, M.-A.; Baccarelli, A.; Zanobetti, A.; Tarantini, L.; Suh, H.; Vokonas, P.; Schwartz, J. Air Pollution and Markers of Coagulation, Inflammation, and Endothelial Function: Associations and Epigene-Environment Interactions in an Elderly Cohort. Epidemiology 2012, 23, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Alexeeff, S.E.; Coull, B.A.; Gryparis, A.; Suh, H.; Sparrow, D.; Vokonas, P.S.; Schwartz, J. Medium-Term Exposure to Traffic-Related Air Pollution and Markers of Inflammation and Endothelial Function. Environ. Health Perspect. 2011, 119, 481–486. [Google Scholar] [CrossRef]
- Bind, M.-A.; Peters, A.; Koutrakis, P.; Coull, B.; Vokonas, P.; Schwartz, J. Quantile Regression Analysis of the Distributional Effects of Air Pollution on Blood Pressure, Heart Rate Variability, Blood Lipids, and Biomarkers of Inflammation in Elderly American Men: The Normative Aging Study. Environ. Health Perspect. 2016, 124, 1189–1198. [Google Scholar] [CrossRef]
- Davis, E.; Malig, B.; Broadwin, R.; Ebisu, K.; Basu, R.; Gold, E.B.; Qi, L.; Derby, C.A.; Park, S.K.; Wu, X. Association between Coarse Particulate Matter and Inflammatory and Hemostatic Markers in a Cohort of Midlife Women. Environ. Health 2020, 19, 111. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Basu, R.; Malig, B.; Broadwin, R.; Ebisu, K.; Gold, E.B.; Qi, L.; Derby, C.; Green, R.S. Association between Gaseous Air Pollutants and Inflammatory, Hemostatic and Lipid Markers in a Cohort of Midlife Women. Environ. Int. 2017, 107, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Green, R.; Broadwin, R.; Malig, B.; Basu, R.; Gold, E.B.; Qi, L.; Sternfeld, B.; Bromberger, J.T.; Greendale, G.A.; Kravitz, H.M.; et al. Long-and Short-Term Exposure To Air Pollution and Inflammatory/Hemostatic Markers in Midlife Women. Epidemiology 2016, 27, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Lanki, T.; Hampel, R.; Tiittanen, P.; Andrich, S.; Beelen, R.; Brunekreef, B.; Dratva, J.; De Faire, U.; Fuks, K.B.; Hoffmann, B.; et al. Air Pollution from Road Traffic and Systemic Inflammation in Adults: A Cross-Sectional Analysis in the European ESCAPE Project. Environ. Health Perspect. 2015, 123, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Rückerl, R.; Hampel, R.; Breitner, S.; Cyrys, J.; Kraus, U.; Carter, J.; Dailey, L.; Devlin, R.B.; Diaz-Sanchez, D.; Koenig, W.; et al. Associations between Ambient Air Pollution and Blood Markers of Inflammation and Coagulation/Fibrinolysis in Susceptible Populations. Environ. Int. 2014, 70, 32–49. [Google Scholar] [CrossRef]
- Pilz, V.; Wolf, K.; Breitner, S.; Rückerl, R.; Koenig, W.; Rathmann, W.; Cyrys, J.; Peters, A.; Schneider, A. C-Reactive Protein (CRP) and Long-Term Air Pollution with a Focus on Ultrafine Particles. Int. J. Hyg. Environ. Health 2018, 221, 510–518. [Google Scholar] [CrossRef]
- Viehmann, A.; Hertel, S.; Fuks, K.; Eisele, L.; Moebus, S.; Möhlenkamp, S.; Nonnemacher, M.; Jakobs, H.; Erbel, R.; Jöckel, K.-H.; et al. Long-Term Residential Exposure to Urban Air Pollution, and Repeated Measures of Systemic Blood Markers of Inflammation and Coagulation. Occup. Environ. Med. 2015, 72, 656–663. [Google Scholar] [CrossRef]
- Hertel, S.; Viehmann, A.; Moebus, S.; Mann, K.; Bröcker-Preuss, M.; Möhlenkamp, S.; Nonnemacher, M.; Erbel, R.; Jakobs, H.; Memmesheimer, M.; et al. Influence of Short-Term Exposure to Ultrafine and Fine Particles on Systemic Inflammation. Eur. J. Epidemiol. 2010, 25, 581–592. [Google Scholar] [CrossRef]
- Li, W.; Dorans, K.S.; Wilker, E.H.; Rice, M.B.; Ljungman, P.L.; Schwartz, J.D.; Coull, B.A.; Koutrakis, P.; Gold, D.R.; Keaney, J.F.; et al. Short-Term Exposure to Ambient Air Pollution and Circulating Biomarkers of Endothelial Cell Activation: The Framingham Heart Study. Environ. Res. 2019, 171, 36–43. [Google Scholar] [CrossRef]
- Li, W.; Dorans, K.S.; Wilker, E.H.; Rice, M.B.; Ljungman, P.L.; Schwartz, J.D.; Coull, B.A.; Koutrakis, P.; Gold, D.R.; Keaney, J.F.; et al. Short-Term Exposure to Ambient Air Pollution and Biomarkers of Systemic Inflammation: The Framingham Heart Study. ATVB 2017, 37, 1793–1800. [Google Scholar] [CrossRef]
- Xu, H.; Wang, T.; Liu, S.; Brook, R.D.; Feng, B.; Zhao, Q.; Song, X.; Yi, T.; Chen, J.; Zhang, Y.; et al. Extreme Levels of Air Pollution Associated With Changes in Biomarkers of Atherosclerotic Plaque Vulnerability and Thrombogenicity in Healthy Adults: The Beijing AIRCHD Study. Circ. Res. 2019, 124, e30–e43. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Liu, C.; Yi, T.; Song, X.; Wang, Y.; Liu, S.; Chen, J.; Zhao, Q.; Zhang, Y.; Wang, T.; et al. Perturbation of Amino Acid Metabolism Mediates Air Pollution Associated Vascular Dysfunction in Healthy Adults. Environ. Res. 2021, 201, 111512. [Google Scholar] [CrossRef] [PubMed]
- Fiorito, G.; Vlaanderen, J.; Polidoro, S.; Gulliver, J.; Galassi, C.; Ranzi, A.; Krogh, V.; Grioni, S.; Agnoli, C.; Sacerdote, C.; et al. Oxidative Stress and Inflammation Mediate the Effect of Air Pollution on Cardio- and Cerebrovascular Disease: A Prospective Study in Nonsmokers: Effect of Air Pollution on Cardio- and Cerebrovascular Disease. Environ. Mol. Mutagen. 2018, 59, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Corlin, L.; Woodin, M.; Hart, J.E.; Simon, M.C.; Gute, D.M.; Stowell, J.; Tucker, K.L.; Durant, J.L.; Brugge, D. Longitudinal Associations of Long-Term Exposure to Ultrafine Particles with Blood Pressure and Systemic Inflammation in Puerto Rican Adults. Environ. Health 2018, 17, 33. [Google Scholar] [CrossRef]
- Azzouz, M.; Xu, Y.; Barregard, L.; Fagerberg, B.; Zöller, B.; Molnár, P.; Oudin, A.; Spanne, M.; Engström, G.; Stockfelt, L. Air Pollution and Biomarkers of Cardiovascular Disease and Inflammation in the Malmö Diet and Cancer Cohort. Environ. Health 2022, 21, 39. [Google Scholar] [CrossRef]
- Tripathy, S.; Marsland, A.L.; Kinnee, E.J.; Tunno, B.J.; Manuck, S.B.; Gianaros, P.J.; Clougherty, J.E. Long-Term Ambient Air Pollution Exposures and Circulating and Stimulated Inflammatory Mediators in a Cohort of Midlife Adults. Environ. Health Perspect. 2021, 129, 057007. [Google Scholar] [CrossRef]
- Elbarbary, M.; Oganesyan, A.; Honda, T.; Morgan, G.; Guo, Y.; Guo, Y.; Negin, J. Systemic Inflammation (C-Reactive Protein) in Older Chinese Adults Is Associated with Long-Term Exposure to Ambient Air Pollution. Int. J. Environ. Res. Public Health 2021, 18, 3258. [Google Scholar] [CrossRef]
- Iyer, H.S.; Hart, J.E.; Fiffer, M.R.; Elliott, E.G.; Yanosky, J.D.; Kaufman, J.D.; Puett, R.C.; Laden, F. Impacts of Long-Term Ambient Particulate Matter and Gaseous Pollutants on Circulating Biomarkers of Inflammation in Male and Female Health Professionals. Environ. Res. 2022, 214, 113810. [Google Scholar] [CrossRef]
- Dauchet, L.; Hulo, S.; Cherot-Kornobis, N.; Matran, R.; Amouyel, P.; Edmé, J.-L.; Giovannelli, J. Short-Term Exposure to Air Pollution: Associations with Lung Function and Inflammatory Markers in Non-Smoking, Healthy Adults. Environ. Int. 2018, 121, 610–619. [Google Scholar] [CrossRef]
- Mostafavi, N.; Vlaanderen, J.; Chadeau-Hyam, M.; Beelen, R.; Modig, L.; Palli, D.; Bergdahl, I.A.; Vineis, P.; Hoek, G.; Kyrtopoulos, S.A.; et al. Inflammatory Markers in Relation to Long-Term Air Pollution. Environ. Int. 2015, 81, 1–7. [Google Scholar] [CrossRef]
- Hajat, A.; Allison, M.; Diez-Roux, A.V.; Jenny, N.S.; Jorgensen, N.W.; Szpiro, A.A.; Vedal, S.; Kaufman, J.D. Long-Term Exposure to Air Pollution and Markers of Inflammation, Coagulation, and Endothelial Activation: A Repeat-Measures Analysis in the Multi-Ethnic Study of Atherosclerosis (MESA). Epidemiology 2015, 26, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Lane, K.J.; Levy, J.I.; Scammell, M.K.; Peters, J.L.; Patton, A.P.; Reisner, E.; Lowe, L.; Zamore, W.; Durant, J.L.; Brugge, D. Association of Modeled Long-Term Personal Exposure to Ultrafine Particles with Inflammatory and Coagulation Biomarkers. Environ. Int. 2016, 92–93, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Tsai, D.-H.; Amyai, N.; Marques-Vidal, P.; Wang, J.-L.; Riediker, M.; Mooser, V.; Paccaud, F.; Waeber, G.; Vollenweider, P.; Bochud, M. Effects of Particulate Matter on Inflammatory Markers in the General Adult Population. Part. Fibre Toxicol. 2012, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Sarnat, J.A.; Liu, K.H.; Hood, R.B.; Chang, C.-J.; Hu, X.; Tran, V.; Greenwald, R.; Chang, H.H.; Russell, A.; et al. Evaluation of the Use of Saliva Metabolome as a Surrogate of Blood Metabolome in Assessing Internal Exposures to Traffic-Related Air Pollution. Environ. Sci. Technol. 2022, 56, 6525–6536. [Google Scholar] [CrossRef]
- Liang, D.; Moutinho, J.L.; Golan, R.; Yu, T.; Ladva, C.N.; Niedzwiecki, M.; Walker, D.I.; Sarnat, S.E.; Chang, H.H.; Greenwald, R.; et al. Use of High-Resolution Metabolomics for the Identification of Metabolic Signals Associated with Traffic-Related Air Pollution. Environ. Int. 2018, 120, 145–154. [Google Scholar] [CrossRef]
- Tang, Z.; Sarnat, J.A.; Weber, R.J.; Russell, A.G.; Zhang, X.; Li, Z.; Yu, T.; Jones, D.P.; Liang, D. The Oxidative Potential of Fine Particulate Matter and Biological Perturbations in Human Plasma and Saliva Metabolome. Environ. Sci. Technol. 2022, 56, 7350–7361. [Google Scholar] [CrossRef]
- Nassan, F.L.; Kelly, R.S.; Kosheleva, A.; Koutrakis, P.; Vokonas, P.S.; Lasky-Su, J.A.; Schwartz, J.D. Metabolomic Signatures of the Long-Term Exposure to Air Pollution and Temperature. Environ. Health 2021, 20, 3. [Google Scholar] [CrossRef]
- Nassan, F.L.; Wang, C.; Kelly, R.S.; Lasky-Su, J.A.; Vokonas, P.S.; Koutrakis, P.; Schwartz, J.D. Ambient PM2.5 Species and Ultrafine Particle Exposure and Their Differential Metabolomic Signatures. Environ. Int. 2021, 151, 106447. [Google Scholar] [CrossRef]
- Liao, J.; Gheissari, R.; Thomas, D.C.; Gilliland, F.D.; Lurmann, F.; Islam, K.T.; Chen, Z. Transcriptomic and Metabolomic Associations with Exposures to Air Pollutants among Young Adults with Childhood Asthma History. Environ. Pollut. 2022, 299, 118903. [Google Scholar] [CrossRef]
- Chen, C.; Li, H.; Niu, Y.; Liu, C.; Lin, Z.; Cai, J.; Li, W.; Ge, W.; Chen, R.; Kan, H. Impact of Short-Term Exposure to Fine Particulate Matter Air Pollution on Urinary Metabolome: A Randomized, Double-Blind, Crossover Trial. Environ. Int. 2019, 130, 104878. [Google Scholar] [CrossRef]
- Yao, Y.; Schneider, A.; Wolf, K.; Zhang, S.; Wang-Sattler, R.; Peters, A.; Breitner, S. Longitudinal Associations between Metabolites and Long-Term Exposure to Ambient Air Pollution: Results from the KORA Cohort Study. Environ. Int. 2022, 170, 107632. [Google Scholar] [CrossRef] [PubMed]
- Ward-Caviness, C.K.; Breitner, S.; Wolf, K.; Cyrys, J.; Kastenmüller, G.; Wang-Sattler, R.; Schneider, A.; Peters, A. Short-Term NO 2 Exposure Is Associated with Long-Chain Fatty Acids in Prospective Cohorts from Augsburg, Germany: Results from an Analysis of 138 Metabolites and Three Exposures. Int. J. Epidemiol. 2016, 45, 1528–1538. [Google Scholar] [CrossRef] [PubMed]
- Jeong, A.; Fiorito, G.; Keski-Rahkonen, P.; Imboden, M.; Kiss, A.; Robinot, N.; Gmuender, H.; Vlaanderen, J.; Vermeulen, R.; Kyrtopoulos, S.; et al. Perturbation of Metabolic Pathways Mediates the Association of Air Pollutants with Asthma and Cardiovascular Diseases. Environ. Int. 2018, 119, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Ladva, C.N.; Golan, R.; Liang, D.; Greenwald, R.; Walker, D.I.; Uppal, K.; Raysoni, A.U.; Tran, V.; Yu, T.; Flanders, W.D.; et al. Particulate Metal Exposures Induce Plasma Metabolome Changes in a Commuter Panel Study. PLoS ONE 2018, 13, e0203468. [Google Scholar] [CrossRef]
- Liang, D.; Ladva, C.N.; Golan, R.; Yu, T.; Walker, D.I.; Sarnat, S.E.; Greenwald, R.; Uppal, K.; Tran, V.; Jones, D.P.; et al. Perturbations of the Arginine Metabolome Following Exposures to Traffic-Related Air Pollution in a Panel of Commuters with and without Asthma. Environ. Int. 2019, 127, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.I.; Lane, K.J.; Liu, K.; Uppal, K.; Patton, A.P.; Durant, J.L.; Jones, D.P.; Brugge, D.; Pennell, K.D. Metabolomic Assessment of Exposure to Near-Highway Ultrafine Particles. J. Expo. Sci. Environ. Epidemiol. 2019, 29, 469–483. [Google Scholar] [CrossRef]
- Hood, R.B.; Liang, D.; Tang, Z.; Kloog, I.; Schwartz, J.; Laden, F.; Jones, D.; Gaskins, A.J. Length of PM2.5 Exposure and Alterations in the Serum Metabolome among Women Undergoing Infertility Treatment. Environ. Epidemiol. 2022, 6, e191. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Han, Y.; Li, H.; Wang, Y.; Chen, X.; Chen, W.; Qiu, X.; Gong, J.; Li, W.; Zhu, T. Proinflammatory Lipid Signals Trigger the Health Effects of Air Pollution in Individuals with Prediabetes. Environ. Pollut. 2021, 290, 118008. [Google Scholar] [CrossRef]
- Menni, C.; Metrustry, S.J.; Mohney, R.P.; Beevers, S.; Barratt, B.; Spector, T.D.; Kelly, F.J.; Valdes, A.M. Circulating Levels of Antioxidant Vitamins Correlate with Better Lung Function and Reduced Exposure to Ambient Pollution. Am. J. Respir. Crit. Care Med. 2015, 191, 1203–1207. [Google Scholar] [CrossRef]
- Khan, A.T.; Gogarten, S.M.; McHugh, C.P.; Stilp, A.M.; Sofer, T.; Bowers, M.L.; Wong, Q.; Cupples, L.A.; Hidalgo, B.; Johnson, A.D.; et al. Recommendations on the Use and Reporting of Race, Ethnicity, and Ancestry in Genetic Research: Experiences from the NHLBI TOPMed Program. Cell Genom. 2022, 2, 100155. [Google Scholar] [CrossRef]
- Zhang, Z.; Chang, L.; Lau, A.K.; Chan, T.-C.; Chieh Chuang, Y.; Chan, J.; Lin, C.; Kai Jiang, W.; Dear, K.; Zee, B.C.; et al. Satellite-Based Estimates of Long-Term Exposure to Fine Particulate Matter Are Associated with C-Reactive Protein in 30,034 Taiwanese Adults. Int. J. Epidemiol. 2017, 46, 1126–1136. [Google Scholar] [CrossRef]
- Breitner, S.; Schneider, A.; Devlin, R.B.; Ward-Caviness, C.K.; Diaz-Sanchez, D.; Neas, L.M.; Cascio, W.E.; Peters, A.; Hauser, E.R.; Shah, S.H.; et al. Associations among Plasma Metabolite Levels and Short-Term Exposure to PM2.5 and Ozone in a Cardiac Catheterization Cohort. Environ. Int. 2016, 97, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Newgard, C.B.; Kim, J.S.; IIkayeva, O.; Alderete, T.L.; Thomas, D.C.; Berhane, K.; Breton, C.; Chatzi, L.; Bastain, T.M.; et al. Near-Roadway Air Pollution Exposure and Altered Fatty Acid Oxidation among Adolescents and Young Adults—The Interplay with Obesity. Environ. Int. 2019, 130, 104935. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Xu, W.; Ji, J.; Yang, Y.; Wang, S.-T.; Wang, J.; Chen, X.; Meng, S.; Tian, X.; Xu, K.-F. Lung Function and Systemic Inflammation Associated with Short-Term Air Pollution Exposure in Chronic Obstructive Pulmonary Disease Patients in Beijing, China. Environ. Health 2020, 19, 12. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Wang, X.; Gao, X.; Cardenas, A.; Baccarelli, A.A.; Guo, X.; Huang, J.; Wu, S. Association of DNA Methylation in Circulating CD4+T Cells with Short-Term PM2.5 Pollution Waves: A Quasi-Experimental Study of Healthy Young Adults. Ecotoxicol. Environ. Saf. 2022, 239, 113634. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Huang, J.; Zhao, Y.; Xue, L.; Li, H.; Liu, Q.; Cao, H.; Peng, W.; Guo, C.; Xie, Y.; et al. Inflammatory Cytokines and DNA Methylation in Healthy Young Adults Exposure to Fine Particulate Matter: A Randomized, Double-Blind Crossover Trial of Air Filtration. J. Hazard. Mater. 2020, 398, 122817. [Google Scholar] [CrossRef]
- Chen, R.; Qiao, L.; Li, H.; Zhao, Y.; Zhang, Y.; Xu, W.; Wang, C.; Wang, H.; Zhao, Z.; Xu, X.; et al. Fine Particulate Matter Constituents, Nitric Oxide Synthase DNA Methylation and Exhaled Nitric Oxide. Environ. Sci. Technol. 2015, 49, 11859–11865. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, W.; Niu, Y.; Xia, Y.; Lei, X.; Huo, J.; Zhao, Q.; Zhang, Y.; Duan, Y.; Cai, J.; et al. The Effects of Fine Particulate Matter Constituents on Exhaled Nitric Oxide and DNA Methylation in the Arginase–Nitric Oxide Synthase Pathway. Environ. Int. 2019, 131, 105019. [Google Scholar] [CrossRef]
- Tobaldini, E.; Bollati, V.; Prado, M.; Fiorelli, E.M.; Pecis, M.; Bissolotti, G.; Albetti, B.; Cantone, L.; Favero, C.; Cogliati, C.; et al. Acute Particulate Matter Affects Cardiovascular Autonomic Modulation and IFN-γ Methylation in Healthy Volunteers. Environ. Res. 2018, 161, 97–103. [Google Scholar] [CrossRef]
- Husby, A. On the Use of Blood Samples for Measuring DNA Methylation in Ecological Epigenetic Studies. Integr. Comp. Biol. 2020, 60, 1558–1566. [Google Scholar] [CrossRef]
- Dunnet, M.J.; Ortega-Recalde, O.J.; Waters, S.A.; Weeks, R.J.; Morison, I.M.; Hore, T.A. Leukocyte-Specific DNA Methylation Biomarkers and Their Implication for Pathological Epigenetic Analysis. Epigenet. Commun. 2022, 2, 5. [Google Scholar] [CrossRef]
- Koestler, D.C.; Christensen, B.C.; Karagas, M.R.; Marsit, C.J.; Langevin, S.M.; Kelsey, K.T.; Wiencke, J.K.; Houseman, E.A. Blood-Based Profiles of DNA Methylation Predict the Underlying Distribution of Cell Types: A Validation Analysis. Epigenetics 2013, 8, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Ramanathan, G.; Zhu, Y.; Yin, F.; Rea, N.D.; Lu, X.; Tseng, C.-H.; Faull, K.F.; Yoon, A.J.; Jerrett, M.; et al. Pro-Oxidative and Proinflammatory Effects after Traveling from Los Angeles to Beijing: A Biomarker-Based Natural Experiment. Circulation 2019, 140, 1995–2004. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Chen, R.; Wang, C.; Shi, J.; Zhao, Z.; Li, W.; Yan, B.; Chillrud, S.; Cai, J.; Kan, H. Personal Fine Particulate Matter Constituents, Increased Systemic Inflammation, and the Role of DNA Hypomethylation. Environ. Sci. Technol. 2019, 53, 9837–9844. [Google Scholar] [CrossRef]
- Midouhas, E.; Kokosi, T.; Flouri, E. Neighbourhood-Level Air Pollution and Greenspace and Inflammation in Adults. Health Place 2019, 58, 102167. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-Y.; Chan, C.-C.; Su, T.-C. Particulate and Gaseous Pollutants on Inflammation, Thrombosis, and Autonomic Imbalance in Subjects at Risk for Cardiovascular Disease. Environ. Pollut. 2017, 223, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Ryu, M.H.; Lau, K.S.-K.; Wooding, D.J.; Fan, S.; Sin, D.D.; Carlsten, C. Particle Depletion of Diesel Exhaust Restores Allergen-Induced Lung-Protective Surfactant Protein D in Human Lungs. Thorax 2020, 75, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Biagioni, B.J.; Tam, S.; Chen, Y.-W.R.; Sin, D.D.; Carlsten, C. Effect of Controlled Human Exposure to Diesel Exhaust and Allergen on Airway Surfactant Protein D, Myeloperoxidase and Club (Clara) Cell Secretory Protein 16. Clin. Exp. Allergy 2016, 46, 1206–1213. [Google Scholar] [CrossRef]
- Mookherjee, N.; Piyadasa, H.; Ryu, M.H.; Rider, C.F.; Ezzati, P.; Spicer, V.; Carlsten, C. Inhaled Diesel Exhaust Alters the Allergen-Induced Bronchial Secretome in Humans. Eur. Respir. J. 2018, 51, 1701385. [Google Scholar] [CrossRef]
- Espinosa, C.; Ali, S.M.; Khan, W.; Khanam, R.; Pervin, J.; Price, J.T.; Rahman, S.; Hasan, T.; Ahmed, S.; Raqib, R.; et al. Comparative Predictive Power of Serum vs Plasma Proteomic Signatures in Feto-Maternal Medicine. AJOG Glob. Rep. 2023, 3, 100244. [Google Scholar] [CrossRef]
- Paul, J.; Veenstra, T.D. Separation of Serum and Plasma Proteins for In-Depth Proteomic Analysis. Separations 2022, 9, 89. [Google Scholar] [CrossRef]
- Sotelo-Orozco, J.; Chen, S.-Y.; Hertz-Picciotto, I.; Slupsky, C.M. A Comparison of Serum and Plasma Blood Collection Tubes for the Integration of Epidemiological and Metabolomics Data. Front. Mol. Biosci. 2021, 8, 682134. [Google Scholar] [CrossRef]
- Yu, Z.; Kastenmüller, G.; He, Y.; Belcredi, P.; Möller, G.; Prehn, C.; Mendes, J.; Wahl, S.; Roemisch-Margl, W.; Ceglarek, U.; et al. Differences between Human Plasma and Serum Metabolite Profiles. PLoS ONE 2011, 6, e21230. [Google Scholar] [CrossRef]
- Huang, Q.; Hu, D.; Wang, X.; Chen, Y.; Wu, Y.; Pan, L.; Li, H.; Zhang, J.; Deng, F.; Guo, X.; et al. The Modification of Indoor PM2.5 Exposure to Chronic Obstructive Pulmonary Disease in Chinese Elderly People: A Meet-in-Metabolite Analysis. Environ. Int. 2018, 121, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, S.N.; Das, A.; Meena, R.; Nanda, R.K.; Rajamani, P. Biofluid Metabotyping of Occupationally Exposed Subjects to Air Pollution Demonstrates High Oxidative Stress and Deregulated Amino Acid Metabolism. Sci. Rep. 2016, 6, 35972. [Google Scholar] [CrossRef]
- Zhang, Y.; Chu, M.; Zhang, J.; Duan, J.; Hu, D.; Zhang, W.; Yang, X.; Jia, X.; Deng, F.; Sun, Z. Urine Metabolites Associated with Cardiovascular Effects from Exposure of Size-Fractioned Particulate Matter in a Subway Environment: A Randomized Crossover Study. Environ. Int. 2019, 130, 104920. [Google Scholar] [CrossRef]
- Zhang, Q.; Du, X.; Li, H.; Jiang, Y.; Zhu, X.; Zhang, Y.; Niu, Y.; Liu, C.; Ji, J.; Chillrud, S.N.; et al. Cardiovascular Effects of Traffic-Related Air Pollution: A Multi-Omics Analysis from a Randomized, Crossover Trial. J. Hazard. Mater. 2022, 435, 129031. [Google Scholar] [CrossRef] [PubMed]
- Gouveia-Figueira, S.; Karimpour, M.; Bosson, J.A.; Blomberg, A.; Unosson, J.; Sehlstedt, M.; Pourazar, J.; Sandström, T.; Behndig, A.F.; Nording, M.L. Mass Spectrometry Profiling Reveals Altered Plasma Levels of Monohydroxy Fatty Acids and Related Lipids in Healthy Humans after Controlled Exposure to Biodiesel Exhaust. Anal. Chim. Acta 2018, 1018, 62–69. [Google Scholar] [CrossRef]
- Cheng, W.; Duncan, K.E.; Ghio, A.J.; Ward-Caviness, C.; Karoly, E.D.; Diaz-Sanchez, D.; Conolly, R.B.; Devlin, R.B. Changes in Metabolites Present in Lung-Lining Fluid Following Exposure of Humans to Ozone. Toxicol. Sci. 2018, 163, 430–439. [Google Scholar] [CrossRef]
- Lam, M.P.Y.; Ping, P.; Murphy, E. Proteomics Research in Cardiovascular Medicine and Biomarker Discovery. J. Am. Coll. Cardiol. 2016, 68, 2819–2830. [Google Scholar] [CrossRef] [PubMed]
- Marshall, K.D.; Edwards, M.A.; Krenz, M.; Davis, J.W.; Baines, C.P. Proteomic Mapping of Proteins Released during Necrosis and Apoptosis from Cultured Neonatal Cardiac Myocytes. Am. J. Physiol.-Cell Physiol. 2014, 306, C639–C647. [Google Scholar] [CrossRef]
- Fang, M.; Ivanisevic, J.; Benton, H.P.; Johnson, C.H.; Patti, G.J.; Hoang, L.T.; Uritboonthai, W.; Kurczy, M.E.; Siuzdak, G. Thermal Degradation of Small Molecules: A Global Metabolomic Investigation. Anal. Chem. 2015, 87, 10935–10941. [Google Scholar] [CrossRef]
- Lu, W.; Su, X.; Klein, M.S.; Lewis, I.A.; Fiehn, O.; Rabinowitz, J.D. Metabolite Measurement: Pitfalls to Avoid and Practices to Follow. Annu. Rev. Biochem. 2017, 86, 277–304. [Google Scholar] [CrossRef]
- Kurbatov, I.; Dolgalev, G.; Arzumanian, V.; Kiseleva, O.; Poverennaya, E. The Knowns and Unknowns in Protein–Metabolite Interactions. Int. J. Mol. Sci. 2023, 24, 4155. [Google Scholar] [CrossRef]
- Cantone, L.; Tobaldini, E.; Favero, C.; Albetti, B.; Sacco, R.M.; Torgano, G.; Ferrari, L.; Montano, N.; Bollati, V. Particulate Air Pollution, Clock Gene Methylation, and Stroke: Effects on Stroke Severity and Disability. Int. J. Mol. Sci. 2020, 21, 3090. [Google Scholar] [CrossRef]
- Wang, C.; Chen, R.; Shi, M.; Cai, J.; Shi, J.; Yang, C.; Li, H.; Lin, Z.; Meng, X.; Liu, C.; et al. Possible Mediation by Methylation in Acute Inflammation Following Personal Exposure to Fine Particulate Air Pollution. Am. J. Epidemiol. 2018, 187, 484–493. [Google Scholar] [CrossRef]
- Chen, R.; Meng, X.; Zhao, A.; Wang, C.; Yang, C.; Li, H.; Cai, J.; Zhao, Z.; Kan, H. DNA Hypomethylation and Its Mediation in the Effects of Fine Particulate Air Pollution on Cardiovascular Biomarkers: A Randomized Crossover Trial. Environ. Int. 2016, 94, 614–619. [Google Scholar] [CrossRef]
- Wang, C.; Chen, R.; Cai, J.; Shi, J.; Yang, C.; Tse, L.A.; Li, H.; Lin, Z.; Meng, X.; Liu, C.; et al. Personal Exposure to Fine Particulate Matter and Blood Pressure: A Role of Angiotensin Converting Enzyme and Its DNA Methylation. Environ. Int. 2016, 94, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Bellavia, A.; Urch, B.; Speck, M.; Brook, R.D.; Scott, J.A.; Albetti, B.; Behbod, B.; North, M.; Valeri, L.; Bertazzi, P.A.; et al. DNA Hypomethylation, Ambient Particulate Matter, and Increased Blood Pressure: Findings from Controlled Human Exposure Experiments. JAHA 2013, 2, e000212. [Google Scholar] [CrossRef]
- Cantone, L.; Iodice, S.; Tarantini, L.; Albetti, B.; Restelli, I.; Vigna, L.; Bonzini, M.; Pesatori, A.C.; Bollati, V. Particulate Matter Exposure Is Associated with Inflammatory Gene Methylation in Obese Subjects. Environ. Res. 2017, 152, 478–484. [Google Scholar] [CrossRef]
- Barchitta, M.; Maugeri, A.; Quattrocchi, A.; Barone, G.; Mazzoleni, P.; Catalfo, A.; De Guidi, G.; Iemmolo, M.G.; Crimi, N.; Agodi, A. Mediterranean Diet and Particulate Matter Exposure Are Associated with LINE-1 Methylation: Results from a Cross-Sectional Study in Women. Front. Genet. 2018, 9, 514. [Google Scholar] [CrossRef]
- The BIOS Consortium; Lee, M.K.; Xu, C.-J.; Carnes, M.U.; Nichols, C.E.; Ward, J.M.; Kwon, S.O.; Kim, S.-Y.; Kim, W.J.; London, S.J. Genome-Wide DNA Methylation and Long-Term Ambient Air Pollution Exposure in Korean Adults. Clin. Epigenet. 2019, 11, 37. [Google Scholar] [CrossRef]
- Rabinovitch, N.; Jones, M.J.; Gladish, N.; Faino, A.V.; Strand, M.; Morin, A.M.; MacIsaac, J.; Lin, D.T.S.; Reynolds, P.R.; Singh, A.; et al. Methylation of Cysteinyl Leukotriene Receptor 1 Genes Associates with Lung Function in Asthmatics Exposed to Traffic-Related Air Pollution. Epigenetics 2021, 16, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Clifford, R.L.; Jones, M.J.; MacIsaac, J.L.; McEwen, L.M.; Goodman, S.J.; Mostafavi, S.; Kobor, M.S.; Carlsten, C. Inhalation of Diesel Exhaust and Allergen Alters Human Bronchial Epithelium DNA Methylation. J. Allergy Clin. Immunol. 2017, 139, 112–121. [Google Scholar] [CrossRef]
- Jiang, R.; Jones, M.J.; Sava, F.; Kobor, M.S.; Carlsten, C. Short-Term Diesel Exhaust Inhalation in a Controlled Human Crossover Study Is Associated with Changes in DNA Methylation of Circulating Mononuclear Cells in Asthmatics. Part. Fibre Toxicol. 2014, 11, 71. [Google Scholar] [CrossRef]
- Du, X.; Jiang, Y.; Li, H.; Zhang, Q.; Zhu, X.; Zhou, L.; Wang, W.; Zhang, Y.; Liu, C.; Niu, Y.; et al. Traffic-Related Air Pollution and Genome-Wide DNA Methylation: A Randomized, Crossover Trial. Sci. Total Environ. 2022, 850, 157968. [Google Scholar] [CrossRef]
- Gao, X.; Huang, J.; Cardenas, A.; Zhao, Y.; Sun, Y.; Wang, J.; Xue, L.; Baccarelli, A.A.; Guo, X.; Zhang, L.; et al. Short-Term Exposure of PM2.5 and Epigenetic Aging: A Quasi-Experimental Study. Environ. Sci. Technol. 2022, 56, 14690–14700. [Google Scholar] [CrossRef] [PubMed]
- Honkova, K.; Rossnerova, A.; Chvojkova, I.; Milcova, A.; Margaryan, H.; Pastorkova, A.; Ambroz, A.; Rossner, P.; Jirik, V.; Rubes, J.; et al. Genome-Wide DNA Methylation in Policemen Working in Cities Differing by Major Sources of Air Pollution. Int. J. Mol. Sci. 2022, 23, 1666. [Google Scholar] [CrossRef] [PubMed]
- Duan, R.; Niu, H.; Dong, F.; Yu, T.; Li, X.; Wu, H.; Zhang, Y.; Yang, T. Short-Term Exposure to Fine Particulate Matter and Genome-Wide DNA Methylation in Chronic Obstructive Pulmonary Disease: A Panel Study Conducted in Beijing, China. Front. Public Health 2023, 10, 1069685. [Google Scholar] [CrossRef]
- Li, H.; Chen, R.; Cai, J.; Cui, X.; Huang, N.; Kan, H. Short-Term Exposure to Fine Particulate Air Pollution and Genome-Wide DNA Methylation: A Randomized, Double-Blind, Crossover Trial. Environ. Int. 2018, 120, 130–136. [Google Scholar] [CrossRef]
- Su, C.-L.; Tantoh, D.M.; Chou, Y.-H.; Wang, L.; Ho, C.-C.; Chen, P.-H.; Lee, K.-J.; Nfor, O.N.; Hsu, S.-Y.; Liang, W.-M.; et al. Blood-Based SOX2-Promoter Methylation in Relation to Exercise and PM2.5 Exposure among Taiwanese Adults. Cancers 2020, 12, 504. [Google Scholar] [CrossRef] [PubMed]
- Pidsley, R.; Zotenko, E.; Peters, T.J.; Lawrence, M.G.; Risbridger, G.P.; Molloy, P.; Van Djik, S.; Muhlhausler, B.; Stirzaker, C.; Clark, S.J. Critical Evaluation of the Illumina MethylationEPIC BeadChip Microarray for Whole-Genome DNA Methylation Profiling. Genome Biol. 2016, 17, 208. [Google Scholar] [CrossRef] [PubMed]
- Moran, S.; Arribas, C.; Esteller, M. Validation of a DNA Methylation Microarray for 850,000 CpG Sites of the Human Genome Enriched in Enhancer Sequences. Epigenomics 2016, 8, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Mookherjee, N.; Ryu, M.H.; Hemshekhar, M.; Orach, J.; Spicer, V.; Carlsten, C. Defining the Effects of Traffic-Related Air Pollution on the Human Plasma Proteome Using an Aptamer Proteomic Array: A Dose-Dependent Increase in Atherosclerosis-Related Proteins. Environ. Res. 2022, 209, 112803. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liang, D.; Ye, D.; Chang, H.H.; Ziegler, T.R.; Jones, D.P.; Ebelt, S.T. Application of High-Resolution Metabolomics to Identify Biological Pathways Perturbed by Traffic-Related Air Pollution. Environ. Res. 2021, 193, 110506. [Google Scholar] [CrossRef]
- Walker, D.I.; Hart, J.E.; Patel, C.J.; Rudel, R.; Chu, J.; Garshick, E.; Pennell, K.D.; Laden, F.; Jones, D.P. Integrated Molecular Response of Exposure to Traffic-Related Pollutants in the US Trucking Industry. Environ. Int. 2022, 158, 106957. [Google Scholar] [CrossRef]
- Du, X.; Zhang, Q.; Jiang, Y.; Li, H.; Zhu, X.; Zhang, Y.; Liu, C.; Niu, Y.; Ji, J.; Jiang, C.; et al. Dynamic Molecular Choreography Induced by Traffic Exposure: A Randomized, Crossover Trial Using Multi-Omics Profiling. J. Hazard. Mater. 2022, 424, 127359. [Google Scholar] [CrossRef]
- Cruz, R.; Pasqua, L.; Silveira, A.; Damasceno, M.; Matsuda, M.; Martins, M.; Marquezini, M.V.; Lima-Silva, A.E.; Saldiva, P.; Bertuzzi, R. Traffic-Related Air Pollution and Endurance Exercise: Characterizing Non-Targeted Serum Metabolomics Profiling. Environ. Pollut. 2021, 291, 118204. [Google Scholar] [CrossRef]
- Wang, J.; Lin, L.; Huang, J.; Zhang, J.; Duan, J.; Guo, X.; Wu, S.; Sun, Z. Impact of PM2.5 Exposure on Plasma Metabolome in Healthy Adults during Air Pollution Waves: A Randomized, Crossover Trial. J. Hazard. Mater. 2022, 436, 129180. [Google Scholar] [CrossRef]
- Zhao, L.; Fang, J.; Tang, S.; Deng, F.; Liu, X.; Shen, Y.; Liu, Y.; Kong, F.; Du, Y.; Cui, L.; et al. PM2.5 and Serum Metabolome and Insulin Resistance, Potential Mediation by the Gut Microbiome: A Population-Based Panel Study of Older Adults in China. Environ. Health Perspect. 2022, 130, 027007. [Google Scholar] [CrossRef]
- Lin, Y.; Lu, X.; Qiu, X.; Yin, F.; Faull, K.F.; Tseng, C.-H.; Zhang, J.; Fiehn, O.; Zhu, T.; Araujo, J.A.; et al. Arachidonic Acid Metabolism and Inflammatory Biomarkers Associated with Exposure to Polycyclic Aromatic Hydrocarbons. Environ. Res. 2022, 212, 113498. [Google Scholar] [CrossRef]
- Mu, L.; Niu, Z.; Blair, R.H.; Yu, H.; Browne, R.W.; Bonner, M.R.; Fanter, T.; Deng, F.; Swanson, M. Metabolomics Profiling before, during, and after the Beijing Olympics: A Panel Study of Within-Individual Differences during Periods of High and Low Air Pollution. Environ. Health Perspect. 2019, 127, 057010. [Google Scholar] [CrossRef]
- Miller, D.B.; Ghio, A.J.; Karoly, E.D.; Bell, L.N.; Snow, S.J.; Madden, M.C.; Soukup, J.; Cascio, W.E.; Gilmour, M.I.; Kodavanti, U.P. Ozone Exposure Increases Circulating Stress Hormones and Lipid Metabolites in Humans. Am. J. Respir. Crit. Care Med. 2016, 193, 1382–1391. [Google Scholar] [CrossRef]
- Huan, S.; Jin, S.; Liu, H.; Xia, W.; Liang, G.; Xu, S.; Fang, X.; Li, C.; Wang, Q.; Sun, X.; et al. Fine Particulate Matter Exposure and Perturbation of Serum Metabolome: A Longitudinal Study in Baoding, China. Chemosphere 2021, 276, 130102. [Google Scholar] [CrossRef] [PubMed]
- Reisdorph, N.A.; Walmsley, S.; Reisdorph, R. A Perspective and Framework for Developing Sample Type Specific Databases for LC/MS-Based Clinical Metabolomics. Metabolites 2019, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Malinowska, J.M.; Viant, M.R. Confidence in Metabolite Identification Dictates the Applicability of Metabolomics to Regulatory Toxicology. Curr. Opin. Toxicol. 2019, 16, 32–38. [Google Scholar] [CrossRef]
- Yang, Y.; Lee, M.; Fairn, G.D. Phospholipid Subcellular Localization and Dynamics. J. Biol. Chem. 2018, 293, 6230–6240. [Google Scholar] [CrossRef] [PubMed]
- Boffa, M.B.; Koschinsky, M.L. Oxidized Phospholipids as a Unifying Theory for Lipoprotein(a) and Cardiovascular Disease. Nat. Rev. Cardiol. 2019, 16, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.Z.; Kar, N.S.; Podrez, E.A. Oxidized Phospholipids: Biomarker for Cardiovascular Diseases. Int. J. Biochem. Cell Biol. 2009, 41, 1241–1244. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Lv, S.; Liu, Y.; Li, Y. Biomarkers for the Adverse Effects on Respiratory System Health Associated with Atmospheric Particulate Matter Exposure. J. Hazard. Mater. 2022, 421, 126760. [Google Scholar] [CrossRef]
- Deutschman, D.H.; Carstens, J.S.; Klepper, R.L.; Smith, W.S.; Page, M.T.; Young, T.R.; Gleason, L.A.; Nakajima, N.; Sabbadini, R.A. Predicting Obstructive Coronary Artery Disease with Serum Sphingosine-1-Phosphate. Am. Heart J. 2003, 146, 62–68. [Google Scholar] [CrossRef]
- Yu, R.K.; Tsai, Y.-T.; Ariga, T.; Yanagisawa, M. Structures, Biosynthesis, and Functions of Gangliosides-an Overview. J. Oleo Sci. 2011, 60, 537–544. [Google Scholar] [CrossRef]
- Spijkers, L.J.A.; Van Den Akker, R.F.P.; Janssen, B.J.A.; Debets, J.J.; De Mey, J.G.R.; Stroes, E.S.G.; Van Den Born, B.-J.H.; Wijesinghe, D.S.; Chalfant, C.E.; MacAleese, L.; et al. Hypertension Is Associated with Marked Alterations in Sphingolipid Biology: A Potential Role for Ceramide. PLoS ONE 2011, 6, e21817. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Yu, J.; Shi, R.; Yan, L.; Yang, T.; Li, Y.; Zhang, Z.; Yu, G.; Bai, Y.; Schuchman, E.H.; et al. Elevation of Ceramide and Activation of Secretory Acid Sphingomyelinase in Patients with Acute Coronary Syndromes. Coron. Artery Dis. 2014, 25, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Strand, E.; Pedersen, E.R.; Svingen, G.F.T.; Olsen, T.; Bjørndal, B.; Karlsson, T.; Dierkes, J.; Njølstad, P.R.; Mellgren, G.; Tell, G.S.; et al. Serum Acylcarnitines and Risk of Cardiovascular Death and Acute Myocardial Infarction in Patients with Stable Angina Pectoris. JAHA 2017, 6, e003620. [Google Scholar] [CrossRef]
- Aitken-Buck, H.M.; Krause, J.; Zeller, T.; Jones, P.P.; Lamberts, R.R. Long-Chain Acylcarnitines and Cardiac Excitation-Contraction Coupling: Links to Arrhythmias. Front. Physiol. 2020, 11, 577856. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Canela, M.; Guasch-Ferré, M.; Razquin, C.; Toledo, E.; Hernández-Alonso, P.; Clish, C.B.; Li, J.; Wittenbecher, C.; Dennis, C.; Alonso-Gómez, Á.; et al. Plasma Acylcarnitines and Risk of Incident Heart Failure and Atrial Fibrillation: The Prevención Con Dieta Mediterránea Study. Rev. Española Cardiol. (Engl. Ed.) 2022, 75, 649–658. [Google Scholar] [CrossRef]
- Snodgrass, R.G.; Brüne, B. Regulation and Functions of 15-Lipoxygenases in Human Macrophages. Front. Pharmacol. 2019, 10, 719. [Google Scholar] [CrossRef]
- Burns, J.L.; Nakamura, M.T.; Ma, D.W.L. Differentiating the Biological Effects of Linoleic Acid from Arachidonic Acid in Health and Disease. Prostaglandins Leukot. Essent. Fat. Acids 2018, 135, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Sonnweber, T.; Pizzini, A.; Nairz, M.; Weiss, G.; Tancevski, I. Arachidonic Acid Metabolites in Cardiovascular and Metabolic Diseases. Int. J. Mol. Sci. 2018, 19, 3285. [Google Scholar] [CrossRef]
- Bulló, M.; Papandreou, C.; García-Gavilán, J.; Ruiz-Canela, M.; Li, J.; Guasch-Ferré, M.; Toledo, E.; Clish, C.; Corella, D.; Estruch, R.; et al. Tricarboxylic Acid Cycle Related-Metabolites and Risk of Atrial Fibrillation and Heart Failure. Metabolism 2021, 125, 154915. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.L.; Ruiz-Canela, M.; Razquin, C.; Clish, C.B.; Guasch-Ferré, M.; Babio, N.; Corella, D.; Gómez-Gracia, E.; Fiol, M.; Estruch, R.; et al. Circulating Citric Acid Cycle Metabolites and Risk of Cardiovascular Disease in the PREDIMED Study. Nutr. Metab. Cardiovasc. Dis. 2023, 33, 835–843. [Google Scholar] [CrossRef]
- Tran, D.H.; Wang, Z.V. Glucose Metabolism in Cardiac Hypertrophy and Heart Failure. JAHA 2019, 8, e012673. [Google Scholar] [CrossRef]
- Bell, S.M.; Burgess, T.; Lee, J.; Blackburn, D.J.; Allen, S.P.; Mortiboys, H. Peripheral Glycolysis in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 8924. [Google Scholar] [CrossRef] [PubMed]
- Kuspriyanti, N.P.; Ariyanto, E.F.; Syamsunarno, M.R.A.A. Role of Warburg Effect in Cardiovascular Diseases: A Potential Treatment Option. TOCMJ 2021, 15, 6–17. [Google Scholar] [CrossRef]
- Yang, Z.; Ming, X.-F. Functions of Arginase Isoforms in Macrophage Inflammatory Responses: Impact on Cardiovascular Diseases and Metabolic Disorders. Front. Immunol. 2014, 5, 533. [Google Scholar] [CrossRef]
- Hibbs, J.B.; Vavrin, Z.; Taintor, R.R. L-Arginine Is Required for Expression of the Activated Macrophage Effector Mechanism Causing Selective Metabolic Inhibition in Target Cells. J. Immunol. 1987, 138, 550–565. [Google Scholar] [CrossRef]
- Nathan, C.; Ding, A. Nonresolving Inflammation. Cell 2010, 140, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Mills, C. M1 and M2 Macrophages: Oracles of Health and Disease. Crit. Rev. Immunol. 2012, 32, 463–488. [Google Scholar] [CrossRef]
- Morris, S.M. Arginine Metabolism: Boundaries of Our Knowledge. J. Nutr. 2007, 137, 1602S–1609S. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Ming, X.-F. Arginase: The Emerging Therapeutic Target for Vascular Oxidative Stress and Inflammation. Front. Immunol. 2013, 4, 149. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, R.W.; Rodriguez, P.C.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.B. Arginase: A Multifaceted Enzyme Important in Health and Disease. Physiol. Rev. 2018, 98, 641–665. [Google Scholar] [CrossRef]
- Gauer, B.; Brucker, N.; Barth, A.; Arbo, M.D.; Gioda, A.; Thiesen, F.V.; Nardi, J.; Garcia, S.C. Are Metals and Pyrene Levels Additional Factors Playing a Pivotal Role in Air Pollution-Induced Inflammation in Taxi Drivers? Toxicol. Res. 2018, 7, 8–12. [Google Scholar] [CrossRef]
- Pope, C.A.; Bhatnagar, A.; McCracken, J.P.; Abplanalp, W.; Conklin, D.J.; O’Toole, T. Exposure to Fine Particulate Air Pollution Is Associated With Endothelial Injury and Systemic Inflammation. Circ. Res. 2016, 119, 1204–1214. [Google Scholar] [CrossRef] [PubMed]
- Siponen, T.; Yli-Tuomi, T.; Aurela, M.; Dufva, H.; Hillamo, R.; Hirvonen, M.-R.; Huttunen, K.; Pekkanen, J.; Pennanen, A.; Salonen, I.; et al. Source-Specific Fine Particulate Air Pollution and Systemic Inflammation in Ischaemic Heart Disease Patients. Occup. Environ. Med. 2015, 72, 277–283. [Google Scholar] [CrossRef]
- Rich, D.Q.; Kipen, H.M.; Huang, W.; Wang, G.; Wang, Y.; Zhu, P.; Ohman-Strickland, P.; Hu, M.; Philipp, C.; Diehl, S.R.; et al. Association between Changes in Air Pollution Levels during the Beijing Olympics and Biomarkers of Inflammation and Thrombosis in Healthy Young Adults. JAMA 2012, 307. [Google Scholar] [CrossRef]
- Ramesh, G.; MacLean, A.G.; Philipp, M.T. Cytokines and Chemokines at the Crossroads of Neuroinflammation, Neurodegeneration, and Neuropathic Pain. Mediat. Inflamm. 2013, 2013, 480739. [Google Scholar] [CrossRef] [PubMed]
- Borish, L.C.; Steinke, J.W. 2. Cytokines and Chemokines. J. Allergy Clin. Immunol. 2003, 111, S460–S475. [Google Scholar] [CrossRef]
- Clarke, R.; Valdes-Marquez, E.; Hill, M.; Gordon, J.; Farrall, M.; Hamsten, A.; Watkins, H.; Hopewell, J.C. Plasma Cytokines and Risk of Coronary Heart Disease in the PROCARDIS Study. Open Heart 2018, 5, e000807. [Google Scholar] [CrossRef]
- Tahir, A.; Martinez, P.J.; Ahmad, F.; Fisher-Hoch, S.P.; McCormick, J.; Gay, J.L.; Mirza, S.; Chaudhary, S.U. An Evaluation of Lipid Profile and Pro-Inflammatory Cytokines as Determinants of Cardiovascular Disease in Those with Diabetes: A Study on a Mexican American Cohort. Sci. Rep. 2021, 11, 2435. [Google Scholar] [CrossRef]
- Amin, M.N.; Siddiqui, S.A.; Ibrahim, M.; Hakim, M.L.; Ahammed, M.S.; Kabir, A.; Sultana, F. Inflammatory Cytokines in the Pathogenesis of Cardiovascular Disease and Cancer. SAGE Open Med. 2020, 8, 205031212096575. [Google Scholar] [CrossRef]
- Atamas, S.P.; Chapoval, S.P.; Keegan, A.D. Cytokines in Chronic Respiratory Diseases. F1000 Biol. Rep. 2013, 5, 3. [Google Scholar] [CrossRef]
- Sameer, A.S.; Nissar, S. Toll-Like Receptors (TLRs): Structure, Functions, Signaling, and Role of Their Polymorphisms in Colorectal Cancer Susceptibility. BioMed Res. Int. 2021, 2021, 1157023. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Ahmad, S.; Irshad, R.; Goyal, Y.; Rafat, S.; Siddiqui, N.; Dev, K.; Husain, M.; Ali, S.; Mohan, A.; et al. TLRs in Pulmonary Diseases. Life Sci. 2019, 233, 116671. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Cheong, Y.-K.; Kim, N.-H.; Chung, H.-T.; Kang, D.G.; Pae, H.-O. Mitogen-Activated Protein Kinases and Reactive Oxygen Species: How Can ROS Activate MAPK Pathways? J. Signal Transduct. 2011, 2011, 792639. [Google Scholar] [CrossRef] [PubMed]
- Takata, T.; Araki, S.; Tsuchiya, Y.; Watanabe, Y. Oxidative Stress Orchestrates MAPK and Nitric-Oxide Synthase Signal. Int. J. Mol. Sci. 2020, 21, 8750. [Google Scholar] [CrossRef]
- Muslin, A.J. MAPK Signalling in Cardiovascular Health and Disease: Molecular Mechanisms and Therapeutic Targets. Clin. Sci. 2008, 115, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Jubaidi, F.F.; Zainalabidin, S.; Taib, I.S.; Abdul Hamid, Z.; Mohamad Anuar, N.N.; Jalil, J.; Mohd Nor, N.A.; Budin, S.B. The Role of PKC-MAPK Signalling Pathways in the Development of Hyperglycemia-Induced Cardiovascular Complications. Int. J. Mol. Sci. 2022, 23, 8582. [Google Scholar] [CrossRef]
- Van Paridon, P.C.S.; Panova-Noeva, M.; Van Oerle, R.; Schulz, A.; Prochaska, J.H.; Arnold, N.; Schmidtmann, I.; Beutel, M.; Pfeiffer, N.; Münzel, T.; et al. Lower Levels of vWF Are Associated with Lower Risk of Cardiovascular Disease. Res. Pract. Thromb. Haemost. 2022, 6, e12797. [Google Scholar] [CrossRef]
- Spiel, A.O.; Gilbert, J.C.; Jilma, B. Von Willebrand Factor in Cardiovascular Disease: Focus on Acute Coronary Syndromes. Circulation 2008, 117, 1449–1459. [Google Scholar] [CrossRef]
- Hertle, E.; Van Greevenbroek, M.M.J.; Stehouwer, C.D.A. Complement C3: An Emerging Risk Factor in Cardiometabolic Disease. Diabetologia 2012, 55, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Olson, N.C.; Raffield, L.M.; Lange, L.A.; Lange, E.M.; Longstreth, W.T.; Chauhan, G.; Debette, S.; Seshadri, S.; Reiner, A.P.; Tracy, R.P. Associations of Activated Coagulation Factor VII and Factor VIIa-antithrombin Levels with Genome-wide Polymorphisms and Cardiovascular Disease Risk. J. Thromb. Haemost. 2018, 16, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Junker, R.; Heinrich, J.; Schulte, H.; Van De Loo, J.; Assmann, G. Coagulation Factor VII and the Risk of Coronary Heart Disease in Healthy Men. ATVB 1997, 17, 1539–1544. [Google Scholar] [CrossRef] [PubMed]
- Tofler, G.H.; Massaro, J.; O’Donnell, C.J.; Wilson, P.W.F.; Vasan, R.S.; Sutherland, P.A.; Meigs, J.B.; Levy, D.; D’Agostino, R.B. Plasminogen Activator Inhibitor and the Risk of Cardiovascular Disease: The Framingham Heart Study. Thromb. Res. 2016, 140, 30–35. [Google Scholar] [CrossRef]
- Stec, J.J.; Silbershatz, H.; Tofler, G.H.; Matheney, T.H.; Sutherland, P.; Lipinska, I.; Massaro, J.M.; Wilson, P.F.W.; Muller, J.E.; D’Agostino, R.B. Association of Fibrinogen with Cardiovascular Risk Factors and Cardiovascular Disease in the Framingham Offspring Population. Circulation 2000, 102, 1634–1638. [Google Scholar] [CrossRef]
- Zhao, J.V.; Schooling, C.M. Coagulation Factors and the Risk of Ischemic Heart Disease: A Mendelian Randomization Study. Circ. Genom. Precis. Med. 2018, 11, e001956. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Niu, Y.; Xia, Y.; Lei, X.; Wang, W.; Huo, J.; Zhao, Q.; Zhang, Y.; Duan, Y.; Cai, J.; et al. The Acute Effects of Fine Particulate Matter Constituents on Circulating Inflammatory Biomarkers in Healthy Adults. Sci. Total Environ. 2020, 707, 135989. [Google Scholar] [CrossRef]
- Hillis, G.S.; Flapan, A.D. Cell Adhesion Molecules in Cardiovascular Disease: A Clinical Perspective. Heart 1998, 79, 429–431. [Google Scholar] [CrossRef][Green Version]
- Zhou, Y.; Zhu, X.; Cui, H.; Shi, J.; Yuan, G.; Shi, S.; Hu, Y. The Role of the VEGF Family in Coronary Heart Disease. Front. Cardiovasc. Med. 2021, 8, 738325. [Google Scholar] [CrossRef]
- Camasão, D.B.; Mantovani, D. The Mechanical Characterization of Blood Vessels and Their Substitutes in the Continuous Quest for Physiological-Relevant Performances. A Critical Review. Mater. Today Bio 2021, 10, 100106. [Google Scholar] [CrossRef]
- Bevan, G.H.; Al-Kindi, S.G.; Brook, R.D.; Münzel, T.; Rajagopalan, S. Ambient Air Pollution and Atherosclerosis: Insights Into Dose, Time, and Mechanisms. ATVB 2021, 41, 628–637. [Google Scholar] [CrossRef]
- Schwartz, A.B.; Campos, O.A.; Criado-Hidalgo, E.; Chien, S.; Del Álamo, J.C.; Lasheras, J.C.; Yeh, Y.-T. Elucidating the Biomechanics of Leukocyte Transendothelial Migration by Quantitative Imaging. Front. Cell Dev. Biol. 2021, 9, 635263. [Google Scholar] [CrossRef] [PubMed]
- Gondalia, R.; Holliday, K.M.; Baldassari, A.; Justice, A.E.; Stewart, J.D.; Liao, D.; Yanosky, J.D.; Engel, S.M.; Jordahl, K.M.; Bhatti, P.; et al. Leukocyte Traits and Exposure to Ambient Particulate Matter Air Pollution in the Women’s Health Initiative and Atherosclerosis Risk in Communities Study. Environ. Health Perspect. 2020, 128, 017004. [Google Scholar] [CrossRef] [PubMed]
- Rezatabar, S.; Karimian, A.; Rameshknia, V.; Parsian, H.; Majidinia, M.; Kopi, T.A.; Bishayee, A.; Sadeghinia, A.; Yousefi, M.; Monirialamdari, M.; et al. RAS/MAPK Signaling Functions in Oxidative Stress, DNA Damage Response and Cancer Progression. J. Cell. Physiol. 2019, 234, 14951–14965. [Google Scholar] [CrossRef]
- Liu, D.; Xu, Y. P53, Oxidative Stress, and Aging. Antioxid. Redox Signal. 2011, 15, 1669–1678. [Google Scholar] [CrossRef]
- Liu, X.; Fan, L.; Lu, C.; Yin, S.; Hu, H. Functional Role of P53 in the Regulation of Chemical-Induced Oxidative Stress. Oxid. Med. Cell. Longev. 2020, 2020, 6039769. [Google Scholar] [CrossRef]
- Kannan, K.; Jain, S.K. Oxidative Stress and Apoptosis. Pathophysiology 2000, 7, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; McMillan-Ward, E.; Kong, J.; Israels, S.J.; Gibson, S.B. Oxidative Stress Induces Autophagic Cell Death Independent of Apoptosis in Transformed and Cancer Cells. Cell Death Differ. 2008, 15, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.R.; Rai, U.; Fanburg, B.L.; Cochran, B.H. Activation of the JAK-STAT Pathway by Reactive Oxygen Species. Am. J. Physiol.-Cell Physiol. 1998, 275, C1640–C1652. [Google Scholar] [CrossRef]
- Yun, H.R.; Jo, Y.H.; Kim, J.; Shin, Y.; Kim, S.S.; Choi, T.G. Roles of Autophagy in Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 3289. [Google Scholar] [CrossRef]
- Chan, G.H.-H.; Chan, E.; Kwok, C.T.-K.; Leung, G.P.-H.; Lee, S.M.-Y.; Seto, S.-W. The Role of P53 in the Alternation of Vascular Functions. Front. Pharmacol. 2022, 13, 981152. [Google Scholar] [CrossRef] [PubMed]
- Men, H.; Cai, H.; Cheng, Q.; Zhou, W.; Wang, X.; Huang, S.; Zheng, Y.; Cai, L. The Regulatory Roles of P53 in Cardiovascular Health and Disease. Cell. Mol. Life Sci. 2021, 78, 2001–2018. [Google Scholar] [CrossRef]
- Uddin, M.A.; Barabutis, N. P53 in the Impaired Lungs. DNA Repair. 2020, 95, 102952. [Google Scholar] [CrossRef] [PubMed]
- Seif, F.; Khoshmirsafa, M.; Aazami, H.; Mohsenzadegan, M.; Sedighi, G.; Bahar, M. The Role of JAK-STAT Signaling Pathway and Its Regulators in the Fate of T Helper Cells. Cell Commun. Signal. 2017, 15, 23. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.R.; Manzoor Syed, B.; Laghari, Z.; Pirzada, S. Analysis of Inflammatory Markers in Apparently Healthy Automobile Vehicle Drivers in Response to Exposure to Traffic Pollution Fumes: Inflammatory Markers in Healthy Automobile Vehicle Drivers. Pak. J. Med. Sci. 2020, 36, 657. [Google Scholar] [CrossRef] [PubMed]
- Laratta, C.R.; Kendzerska, T.; Carlsten, C.; Brauer, M.; Van Eeden, S.F.; Allen, A.J.M.H.; Fox, N.; Peres, B.U.; Ayas, N.T. Air Pollution and Systemic Inflammation in Patients with Suspected OSA Living in an Urban Residential Area. Chest 2020, 158, 1713–1722. [Google Scholar] [CrossRef]
- Mascareno, E.; El-Shafei, M.; Maulik, N.; Sato, M.; Guo, Y.; Das, D.K.; Siddiqui, M.A.Q. JAK/STAT Signaling Is Associated with Cardiac Dysfunction during Ischemia and Reperfusion. Circulation 2001, 104, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Kishore, R.; Verma, S.K. Roles of STATs Signaling in Cardiovascular Diseases. JAK-STAT 2012, 1, 118–124. [Google Scholar] [CrossRef]
- Purohit, M.; Gupta, G.; Afzal, O.; Altamimi, A.S.A.; Alzarea, S.I.; Kazmi, I.; Almalki, W.H.; Gulati, M.; Kaur, I.P.; Singh, S.K.; et al. Janus Kinase/Signal Transducers and Activator of Transcription (JAK/STAT) and Its Role in Lung Inflammatory Disease. Chem.-Biol. Interact. 2023, 371, 110334. [Google Scholar] [CrossRef]
- Yew-Booth, L.; Birrell, M.A.; Lau, M.S.; Baker, K.; Jones, V.; Kilty, I.; Belvisi, M.G. JAK–STAT Pathway Activation in COPD. Eur. Respir. J. 2015, 46, 843–845. [Google Scholar] [CrossRef]
- Kim, N.-H.; Kang, P.M. Apoptosis in Cardiovascular Diseases: Mechanism and Clinical Implications. Korean Circ. J. 2010, 40, 299. [Google Scholar] [CrossRef]
- Henson, P.M.; Tuder, R.M. Apoptosis in the Lung: Induction, Clearance and Detection. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2008, 294, L601–L611. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Gustafsson, Å.B. Role of Apoptosis in Cardiovascular Disease. Apoptosis 2009, 14, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.-J.; Zong, W.-X. The Cellular Decision between Apoptosis and Autophagy. Chin. J. Cancer 2013, 32, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Zhou, X.; Yang, T.; Wang, L.; Feng, L.; Wang, Z.; Xu, J.; Jing, W.; Wang, T.; Su, H.; et al. The Role of Autophagy in Cardiovascular Disease: Cross-Interference of Signaling Pathways and Underlying Therapeutic Targets. Front. Cardiovasc. Med. 2023, 10, 1088575. [Google Scholar] [CrossRef] [PubMed]
- Nishida, K.; Kyoi, S.; Yamaguchi, O.; Sadoshima, J.; Otsu, K. The Role of Autophagy in the Heart. Cell Death Differ. 2009, 16, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Bravo-San Pedro, J.M.; Kroemer, G.; Galluzzi, L. Autophagy and Mitophagy in Cardiovascular Disease. Circ. Res. 2017, 120, 1812–1824. [Google Scholar] [CrossRef] [PubMed]
- Lepeule, J.; Litonjua, A.A.; Coull, B.; Koutrakis, P.; Sparrow, D.; Vokonas, P.S.; Schwartz, J. Long-Term Effects of Traffic Particles on Lung Function Decline in the Elderly. Am. J. Respir. Crit. Care Med. 2014, 190, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Niu, Y.; Xia, Y.; Liu, C.; Lin, Z.; Wang, W.; Ge, Y.; Lei, X.; Wang, C.; Cai, J.; et al. Effects of Personal Nitrogen Dioxide Exposure on Airway Inflammation and Lung Function. Environ. Res. 2019, 177, 108620. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, Y.; Song, Y.; Xie, J.; Cui, X.; Zhang, B.; Shi, T.; Yuan, J.; Chen, W. Short-Term Effects of Outdoor Air Pollution on Lung Function among Female Non-Smokers in China. Sci. Rep. 2016, 6, 34947. [Google Scholar] [CrossRef]
- Bowatte, G.; Lodge, C.J.; Knibbs, L.D.; Erbas, B.; Perret, J.L.; Jalaludin, B.; Morgan, G.G.; Bui, D.S.; Giles, G.G.; Hamilton, G.S.; et al. Traffic Related Air Pollution and Development and Persistence of Asthma and Low Lung Function. Environ. Int. 2018, 113, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Bowatte, G.; Erbas, B.; Lodge, C.J.; Knibbs, L.D.; Gurrin, L.C.; Marks, G.B.; Thomas, P.S.; Johns, D.P.; Giles, G.G.; Hui, J.; et al. Traffic-Related Air Pollution Exposure over a 5-Year Period Is Associated with Increased Risk of Asthma and Poor Lung Function in Middle Age. Eur. Respir. J. 2017, 50, 1602357. [Google Scholar] [CrossRef]
- Schraufnagel, D.E.; Balmes, J.R.; Cowl, C.T.; De Matteis, S.; Jung, S.-H.; Mortimer, K.; Perez-Padilla, R.; Rice, M.B.; Riojas-Rodriguez, H.; Sood, A.; et al. Air Pollution and Noncommunicable Diseases. Chest 2019, 155, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Boogaard, H.; Patton, A.P.; Atkinson, R.W.; Brook, J.R.; Chang, H.H.; Crouse, D.L.; Fussell, J.C.; Hoek, G.; Hoffmann, B.; Kappeler, R.; et al. Long-Term Exposure to Traffic-Related Air Pollution and Selected Health Outcomes: A Systematic Review and Meta-Analysis. Environ. Int. 2022, 164, 107262. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, G.; Tian, L.; Guo, Q.; Pan, X. Short-Term Exposure to Air Pollution and Morbidity of COPD and Asthma in East Asian Area: A Systematic Review and Meta-Analysis. Environ. Res. 2016, 148, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Duan, R.-R.; Hao, K.; Yang, T. Air Pollution and Chronic Obstructive Pulmonary Disease. Chronic Dis. Transl. Med. 2020, 6, 260–269. [Google Scholar] [CrossRef]
- Duong-Quy, S. Clinical Utility of the Exhaled Nitric Oxide (NO) Measurement with Portable Devices in the Management of Allergic Airway Inflammation and Asthma. JAA 2019, 12, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Hancox, R.J.; Poulton, R.; Greene, J.M.; Filsell, S.; McLachlan, C.R.; Rasmussen, F.; Taylor, D.R.; Williams, M.J.A.; Williamson, A.; Sears, M.R. Systemic Inflammation and Lung Function in Young Adults. Thorax 2007, 62, 1064–1068. [Google Scholar] [CrossRef]
- Baines, K.J.; Backer, V.; Gibson, P.G.; Powel, H.; Porsbjerg, C.M. Impaired Lung Function Is Associated with Systemic Inflammation and Macrophage Activation. Eur. Respir. J. 2015, 45, 557–559. [Google Scholar] [CrossRef]
- Nerpin, E.; Jacinto, T.; Fonseca, J.A.; Alving, K.; Janson, C.; Malinovschi, A. Systemic Inflammatory Markers in Relation to Lung Function in NHANES. 2007–2010. Respir. Med. 2018, 142, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Kalhan, R.; Tran, B.T.; Colangelo, L.A.; Rosenberg, S.R.; Liu, K.; Thyagarajan, B.; Jacobs, D.R.; Smith, L.J. Systemic Inflammation in Young Adults Is Associated with Abnormal Lung Function in Middle Age. PLoS ONE 2010, 5, e11431. [Google Scholar] [CrossRef]
- Ward-Caviness, C.K. A Review of Gene-by-Air Pollution Interactions for Cardiovascular Disease, Risk Factors, and Biomarkers. Hum. Genet. 2019, 138, 547–561. [Google Scholar] [CrossRef]
- Al-Kindi, S.G.; Brook, R.D.; Biswal, S.; Rajagopalan, S. Environmental Determinants of Cardiovascular Disease: Lessons Learned from Air Pollution. Nat. Rev. Cardiol. 2020, 17, 656–672. [Google Scholar] [CrossRef]
- Xu, Z.; Wang, W.; Liu, Q.; Li, Z.; Lei, L.; Ren, L.; Deng, F.; Guo, X.; Wu, S. Association between Gaseous Air Pollutants and Biomarkers of Systemic Inflammation: A Systematic Review and Meta-Analysis. Environ. Pollut. 2022, 292, 118336. [Google Scholar] [CrossRef]
- Joshi, A.; Rienks, M.; Theofilatos, K.; Mayr, M. Systems Biology in Cardiovascular Disease: A Multiomics Approach. Nat. Rev. Cardiol. 2021, 18, 313–330. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.Y.; Pandey, A.K.; Maron, B.A.; Loscalzo, J. Network Medicine in Cardiovascular Research. Cardiovasc. Res. 2021, 117, 2186–2202. [Google Scholar] [CrossRef] [PubMed]
- Tahir, U.A.; Gerszten, R.E. Omics and Cardiometabolic Disease Risk Prediction. Annu. Rev. Med. 2020, 71, 163–175. [Google Scholar] [CrossRef]
- Usova, E.I.; Alieva, A.S.; Yakovlev, A.N.; Alieva, M.S.; Prokhorikhin, A.A.; Konradi, A.O.; Shlyakhto, E.V.; Magni, P.; Catapano, A.L.; Baragetti, A. Integrative Analysis of Multi-Omics and Genetic Approaches—A New Level in Atherosclerotic Cardiovascular Risk Prediction. Biomolecules 2021, 11, 1597. [Google Scholar] [CrossRef] [PubMed]
- Battaglini, D.; Al-Husinat, L.; Normando, A.G.; Leme, A.P.; Franchini, K.; Morales, M.; Pelosi, P.; Rocco, P.R. Personalized Medicine Using Omics Approaches in Acute Respiratory Distress Syndrome to Identify Biological Phenotypes. Respir. Res. 2022, 23, 318. [Google Scholar] [CrossRef]
- Wang, X.-W.; Wang, T.; Schaub, D.P.; Chen, C.; Sun, Z.; Ke, S.; Hecker, J.; Maaser-Hecker, A.; Zeleznik, O.A.; Zeleznik, R.; et al. Benchmarking Omics-Based Prediction of Asthma Development in Children. Respir. Res. 2023, 24, 63. [Google Scholar] [CrossRef]
- Nicora, G.; Vitali, F.; Dagliati, A.; Geifman, N.; Bellazzi, R. Integrated Multi-Omics Analyses in Oncology: A Review of Machine Learning Methods and Tools. Front. Oncol. 2020, 10, 1030. [Google Scholar] [CrossRef]
- Nativio, R.; Lan, Y.; Donahue, G.; Sidoli, S.; Berson, A.; Srinivasan, A.R.; Shcherbakova, O.; Amlie-Wolf, A.; Nie, J.; Cui, X.; et al. An Integrated Multi-Omics Approach Identifies Epigenetic Alterations Associated with Alzheimer’s Disease. Nat. Genet. 2020, 52, 1024–1035. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.; Mariani, L.H.; Kretzler, M. Integrated Multi-Omics Approaches to Improve Classification of Chronic Kidney Disease. Nat. Rev. Nephrol. 2020, 16, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Gillenwater, L.A.; Helmi, S.; Stene, E.; Pratte, K.A.; Zhuang, Y.; Schuyler, R.P.; Lange, L.; Castaldi, P.J.; Hersh, C.P.; Banaei-Kashani, F.; et al. Multi-Omics Subtyping Pipeline for Chronic Obstructive Pulmonary Disease. PLoS ONE 2021, 16, e0255337. [Google Scholar] [CrossRef] [PubMed]
- Smilde, A.K.; Næs, T.; Liland, K.H. Multiblock Data Fusion in Statistics and Machine Learning; John Wiley & Sons: Chichester, UK, 2022; ISBN 978-1-119-60096-1. [Google Scholar]
- McCullagh, P. Tensor Methods in Statistics; Dover Publications: Mineola, NY, USA, 2018. [Google Scholar]
- Cai, Z.; Poulos, R.C.; Liu, J.; Zhong, Q. Machine Learning for Multi-Omics Data Integration in Cancer. iScience 2022, 25, 103798. [Google Scholar] [CrossRef] [PubMed]
- Rappoport, N.; Shamir, R. Multi-Omic and Multi-View Clustering Algorithms: Review and Cancer Benchmark. Nucleic Acids Res. 2018, 46, 10546–10562. [Google Scholar] [CrossRef] [PubMed]
- Adossa, N.; Khan, S.; Rytkönen, K.T.; Elo, L.L. Computational Strategies for Single-Cell Multi-Omics Integration. Comput. Struct. Biotechnol. J. 2021, 19, 2588–2596. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casella, C.; Kiles, F.; Urquhart, C.; Michaud, D.S.; Kirwa, K.; Corlin, L. Methylomic, Proteomic, and Metabolomic Correlates of Traffic-Related Air Pollution in the Context of Cardiorespiratory Health: A Systematic Review, Pathway Analysis, and Network Analysis. Toxics 2023, 11, 1014. https://doi.org/10.3390/toxics11121014
Casella C, Kiles F, Urquhart C, Michaud DS, Kirwa K, Corlin L. Methylomic, Proteomic, and Metabolomic Correlates of Traffic-Related Air Pollution in the Context of Cardiorespiratory Health: A Systematic Review, Pathway Analysis, and Network Analysis. Toxics. 2023; 11(12):1014. https://doi.org/10.3390/toxics11121014
Chicago/Turabian StyleCasella, Cameron, Frances Kiles, Catherine Urquhart, Dominique S. Michaud, Kipruto Kirwa, and Laura Corlin. 2023. "Methylomic, Proteomic, and Metabolomic Correlates of Traffic-Related Air Pollution in the Context of Cardiorespiratory Health: A Systematic Review, Pathway Analysis, and Network Analysis" Toxics 11, no. 12: 1014. https://doi.org/10.3390/toxics11121014
APA StyleCasella, C., Kiles, F., Urquhart, C., Michaud, D. S., Kirwa, K., & Corlin, L. (2023). Methylomic, Proteomic, and Metabolomic Correlates of Traffic-Related Air Pollution in the Context of Cardiorespiratory Health: A Systematic Review, Pathway Analysis, and Network Analysis. Toxics, 11(12), 1014. https://doi.org/10.3390/toxics11121014