
Effects of Exogenous Chlorinated Amino Acetic Acid on Cadmium and Mineral Elements in Rice Seedlings




Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Locations
2.2. Cultivation and Processing Methods
2.3. Determination of Cd, K, Ca, Mg, Fe, Mn and Zn Concentrations
2.4. Determination of Subcellular Cadmium Content
2.5. Determination of Total Amino Acids
2.6. Data Statistics and Analysis
3. Results
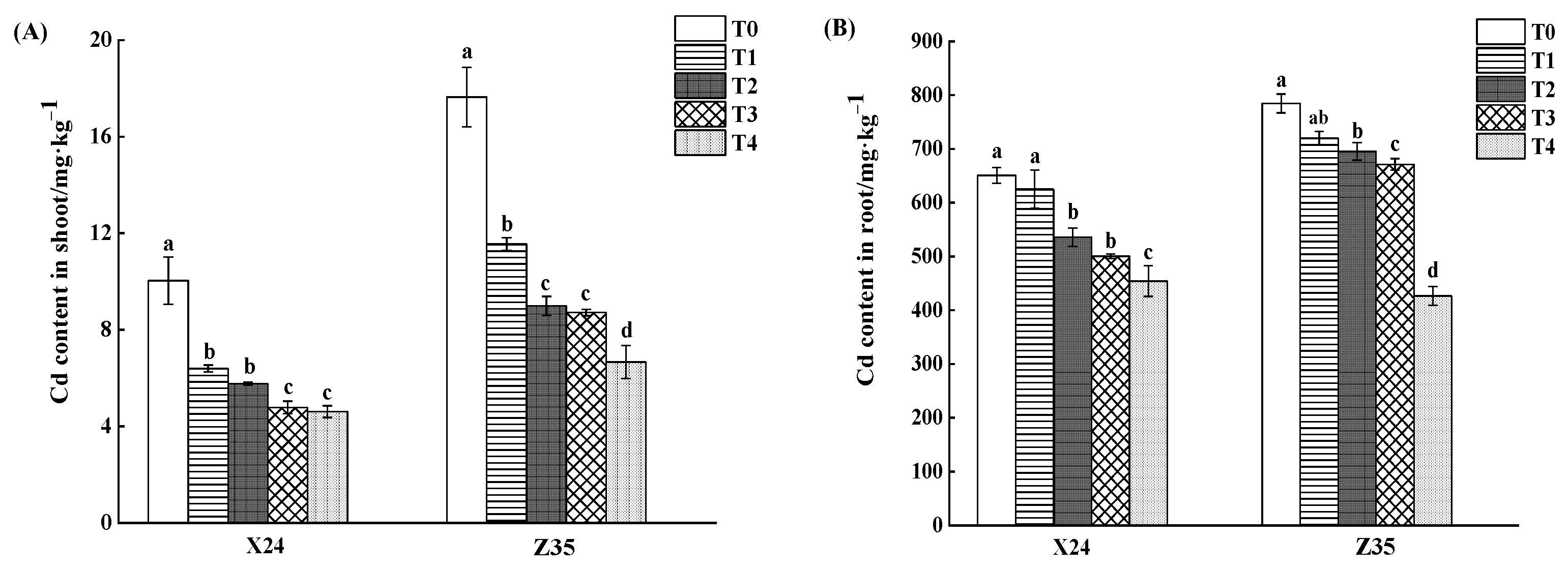
3.1. Effects of Chlorinated Amino Acetic Acid on Cadmium Content in Rice Seedlings
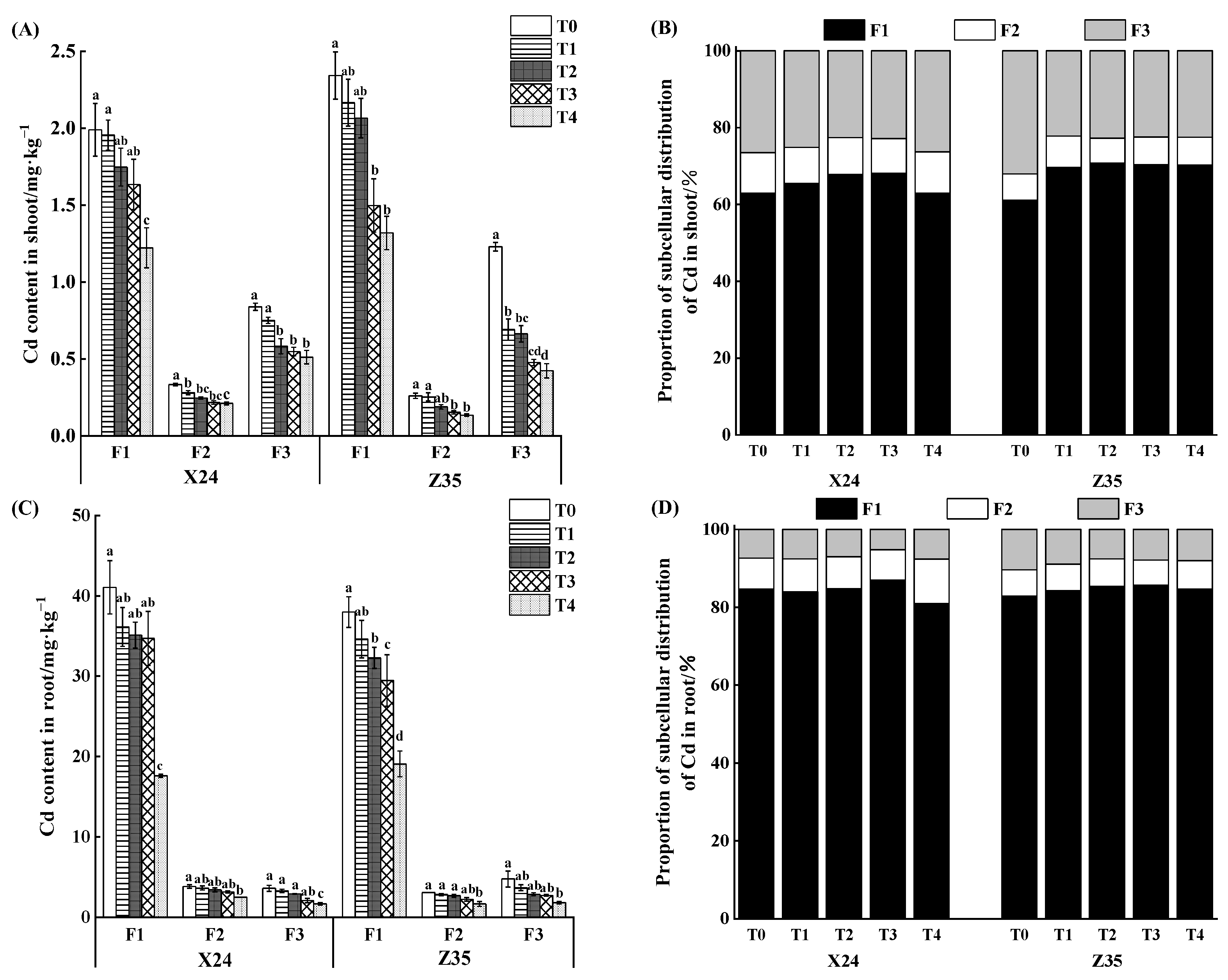
3.2. Effects of Chlorinated Amino Acetic Acid on Cadmium Content in Subcellular of Rice Seedlings
3.3. Effect of Chlorinated Amino Acetic Acid on Content of Mineral Elements in Rice Seedlings
3.4. Effect of Chlorinated Amino Acetic Acid on Total Amino Acid Content of Rice Seedlings
4. Discussion
4.1. Chlorinated Amino Acetic Acid Decreased Cadmium Uptake in Rice Seedlings
4.2. Chlorinated Amino Acetic Acid Promoted the Synthesis of Ca and Mn in Rice Seedlings
4.3. Chlorinated Amino Acetic Acid Promotes the Synthesis of Amino Acids in Cells
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vaverková, M.D.; Maxianová, A.; Winkler, J.; Adamcová, D.; Podlasek, A. Environmental consequences and the role of illegal waste dumps and their impact on land degradation. Land Use Pol. 2019, 89, 104–234. [Google Scholar] [CrossRef]
- Teng, Y.G.; Wu, J.; Lu, S.J.; Wang, Y.Y.; Jiao, X.D.; Song, L.T. Soil and soil environmental quality monitoring in China: A review. Environ. Int. 2014, 69, 177–199. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.J.; Ma, Y.B.; Zhu, Y.G.; Tang, Z.; McGrath, S.P. Soil contamination in China: Current status and mitigation strategies. Environ. Sci. Technol. 2015, 49, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.Q.; Yang, L.Y.; Chen, Y.L.; Jing, H.N.; Wu, P.; Yang, W.T. Selection of rice and maize varieties with low cadmium accumulation and derivation of soil environmental thresholds in karst. Ecotoxicol. Environ. Saf. 2022, 247, 114244. [Google Scholar] [CrossRef]
- Khan, Z.S.; Rizwan, M.; Hafeez, M. Effects of silicon nanoparticles on growth and physiology of wheat in cadmium contaminated soil under different soil moisture levels. Environ. Sci. Pollut. Res. 2020, 27, 4958–4968. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Jiang, J.; Li, S.; Li, M.; Tan, Y.Y.; Song, S.Y.; Shu, Q.Y.; Huang, J.Z. Glutamate alleviates cadmium toxicity in rice via suppressing cadmium uptake and translocation. J. Hazard. Mater. 2020, 384, 121319. [Google Scholar] [CrossRef]
- Song, Y.; Ma, Y.H.; Tang, X.W.; He, X. Research progress of cadmium transference pathway in plant. J. Environ. Manag. Coll. China 2019, 29, 56–59. [Google Scholar]
- Jali, P.; Pradhan, C.; Das, A.B. Effects of cadmium toxicity in plants: A review article. Scholars Acad. J. Biosci. 2016, 4, 1074–1081. [Google Scholar]
- He, J.L.; Li, H.; Luo, J.; Ma, C.F.; Li, S.J.; Qu, L.; Gai, Y.; Jiang, X.N.; Janz, D.; Polle, A.; et al. A transcriptomic network underlies microstructural and physiological responses to cadmium in Populus × canescens. Plant Physiol. 2013, 162, 424–439. [Google Scholar] [CrossRef]
- Hossain, Z.; Komatsu, S. Contribution of proteomic studies towards understanding plant heavy metal stress response. Front. Plant Sci. 2013, 3, 1–12. [Google Scholar] [CrossRef]
- Melandri, G.; AbdElgawad, H.; Riewe, D.; Hageman, J.A.; Asard, H.; Beemster, G.T.S.; Kadam, N.; Jagadish, K.; Altmann, T.; Spira, C.R.; et al. Bouwmeester Biomarkers for grain yield stability in rice under drought stress. J. Exp. Bot. 2020, 71, 669–683. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Khan, S.; Khan, M.A.; Qamar, Z.; Waqas, M. The uptake and bioaccumulation of heavy metals by food plants, their effects on plants nutrients, and associated health risk: A review. Environ. Sci. Pollut. Res. 2015, 22, 13772–13799. [Google Scholar] [CrossRef] [PubMed]
- Han, X.X.; Zhang, C.B.; Wang, C.R.; Huang, Y.C.; Liu, Z.Q. Gadolinium inhibits cadmium transport by blocking non-selective cation channels in rice seedlings. Ecotoxicol. Environ. Saf. 2019, 179, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.L.; Huang, Y.C.; Wang, C.R.; Liu, Z.Q.; Huang, Y.Z.; Zhang, C.B.; Wang, X.L. Mechanism of S-allyl-L-cysteine alleviating cadmium stress in seedling roots and buds of rice seedlings. Environ. Sci. 2020, 42, 3037–3045. [Google Scholar]
- Wang, B.S.; Huang, Y.Z.; Li, J.; Long, J.; Wang, N.; Huang, Y. Effects of different amendments on seed germination and cadmium uptake and accumulation in rice seedlings under cadmium stress. J. Agro-Environ. Sci. 2019, 38, 746–755. [Google Scholar]
- Ji, M.M.; Wang, X.X.; Ma, H.H.; Zhang, C.B.; Ruan, W.Q.; Ren, H.Y.; Deng, Y. Removal of Cd from contaminated soil using amino acid salt. J. Agro-Environ. Sci. 2021, 40, 329–337. [Google Scholar]
- Ma, H.H.; Beadham, I.; Ruan, W.Q.; Zhang, C.B.; Deng, Y. Enhancing rice straw compost with an amino acid-derived ionic liquid as additive. Bioresour. Technol. 2022, 345, 126387. [Google Scholar] [CrossRef]
- Zeilhofer, H.U.; Acuña, M.A.; Jacinthe, G.; Yévenes, G.E. Glycine receptors and glycine transporters: Targets for novel analgesics? Cell. Mol. Life Sci. 2018, 75, 447–465. [Google Scholar] [CrossRef] [PubMed]
- Amelio, I.; Cutruzzolá, F.; Antonov, A.; Agostini, M.; Melino, G. Serine and glycine metabolism in cancer. Trends Biochem. Sci. 2014, 39, 191–198. [Google Scholar] [CrossRef]
- Wang, M.; Wang, L.F.; Zhao, S.W.; Li, S.S.; Lei, X.Q.; Qin, Y.Y.; Sun, X.Y.; Chen, S.B. Manganese facilitates cadmium stabilization through physicochemical dynamics and amino acid accumulation in rice rhizosphere under flood-associated low pe+pH. J. Hazard. Mater. 2021, 416, 126079. [Google Scholar] [CrossRef]
- Liu, Z.Q. Research advance on the mechanism of cadmium transport in rice. Meteorol. Environ. Res. 2014, 5, 48–52. [Google Scholar]
- Zhang, C.J.; Yin, J.; Zhang, C.B.; Wang, J.A.; Liu, Z.Q. Effects of nonselective cation channels on accumulation and transfer of Cd in rice seedlings. J. Agro-Environ. Sci. 2015, 34, 1028–1033. [Google Scholar]
- Zhang, J.; Sun, W.C.; Li, Z.J.; Liang, Y.C.; Song, A. Cadmium fate and tolerance in rice cultivars. Agron. Sustain. Dev. 2009, 29, 483–490. [Google Scholar] [CrossRef]
- Gao, L.; Chang, J.D.; Chen, R.J.; Li, H.B.; Lu, H.F.; Tao, L.X.; Xiong, J. Comparison on cellular mechanisms of iron and cadmium accumulation in rice: Prospects for cultivating Fe-rich but Cd-free rice. Rice 2016, 9, 1–12. [Google Scholar] [CrossRef]
- Loix, C.; Huybrechts, M.; Vangronsveld, J.; Gielen, M.; Keunen, E.; Cuypers, A. Reciprocal interactions between cadmium-induced cell wall responses and oxidative stress in plants. Front. Plant Sci. 2017, 8, 1867. [Google Scholar] [CrossRef]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef]
- Li, H.; Luo, N.; Li, Y.W.; Cai, Q.Y.; Li, H.Y.; Mo, C.H.; Wong, M.H. Cadmium in rice: Transport mechanisms, influencing factors, and minimizing measures. Environ Pollut. 2017, 224, 622–630. [Google Scholar] [CrossRef]
- Liu, Z.Q.; Zhang, C.B. Advance in regulation of heavy metals on nonselective cation channels: A review. J. Agric. Resour. Environ. 2017, 34, 1–5. [Google Scholar]
- Zhang, W.H.; Skerrett, M.; Walker, N.A.; Patrick, J.W.; Tyerman, S.D. Nonselective currents and channels in plasma membranes of protoplasts from coats of developing seeds of bean. Plant Physiol. 2002, 128, 388–399. [Google Scholar] [CrossRef]
- Davenport, R.J.; Tester, M. A weakly voltage-dependent, nonselective cation channel mediates toxic sodium influx in wheat. Plant Physiol. 2000, 122, 823–834. [Google Scholar] [CrossRef]
- White, P.J. Calcium channels in higher plants. Biochim. Biophys. Acta Biomembr. 2000, 1465, 171–189. [Google Scholar] [CrossRef] [PubMed]
- Bouron, A.; Kiselyov, K.; Oberwinkler, J. Permeation, regulation and control of expression of TRP channels by trace metal ions. Pflugers Arch. 2015, 467, 1143–1164. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Ma, L.L.; Yang, F.; Wang, K.W.; Zheng, J. Divalent cations potentiate TRPV1 channel by lowering the heat activation threshold. J. Gen. Physiol. 2014, 143, 75–90. [Google Scholar] [CrossRef]
- Pinto, E.; Ferreira, I. Cation transporters/channels in plants: Tools for nutrient biofortification. J. Plant Physiol. 2015, 179, 64–82. [Google Scholar] [CrossRef]
- Shahid, M.; Pourrut, B.; Dumat, C.; Nadeem, M.; Aslam, M.; Pinelli, E. Heavy-metal-induced reactive oxygen species: Phytotoxicity and physicochemical changes in plants. Rev. Environ. Contam. Toxicol. 2014, 232, 1–44. [Google Scholar] [PubMed]
- Sarwar, N.; Saifullah; Malhi, S.S.; Zia, M.H.; Naeem, A.; Bibi, S.; Farid, G. Role of mineral nutrition in minimizing cadmium accumulation by plants. J. Sci. Food Agric. 2010, 90, 925–937. [Google Scholar] [CrossRef]
- Xue, W.J.; Wang, P.P.; Tang, L.; Zhang, C.B.; Wang, C.R.; Huang, Y.C.; Zhang, X.; Li, Y.K.; Zhao, B.R.; Liu, Z.Q. Citric acid inhibits Cd uptake by improving the preferential transport of Mn and triggering the defense response of amino acids in grains. Ecotoxicol. Environ. Saf. 2021, 211, 111921. [Google Scholar] [CrossRef] [PubMed]
- Pal’ove-Balang, P.; Kisová, A.; Pavlovkin, J.; Mistrík, I. Effect of manganeseon cadmium toxicity in maize seedlings. Plant Soil Environ. 2006, 52, 143–149. [Google Scholar] [CrossRef]
- Kupper, H.; Kupper, F.; Spiller, M. In situdetection of heavy metalsubstititution chlorophylls in water plants. Photosynth. Res. 1998, 58, 123–133. [Google Scholar] [CrossRef]
- Chen, J.; Chris Le, X.; Zhu, L.Z. Metabolomics and transcriptomics reveal defense mechanism of rice (Oryza sativa) grains under stress of 2,2′,4,4′-tetrabromodiphenyl ether. Environ. Int. 2019, 133, 105154. [Google Scholar] [CrossRef]
- Wang, H.J.; Xue, W.J.; Zhang, X.; Zhang, C.B.; Huang, Y.C.; Ren, X.H.; Liu, Z.Q. Effects of foliar application with malic acid on the accumulation of cadmium in rice. J. Agro-Environ. Sci. 2021, 40, 269–278. [Google Scholar]
- Zhang, S.; Lu, Z.Y.; Tang, Q.; Li, H.X.; Zhang, C.B.; Liu, Z.Q. Effects of foliar modulators on cadmium accumulation and transport of nutrient elements in rice. J. Agro-Environ. Sci. 2018, 37, 2507–2513. [Google Scholar]
- Zhao, Y.L.; Zhang, C.B.; Wang, C.R.; Huang, Y.C.; Liu, Z.Q. Increasing phosphate inhibits cadmium uptake in plants and promotes synthesis of amino acids in grains of rice. Environ. Pollut. 2020, 257, 113496. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Akbar, M.Z.; Shakoor, M.B.; Mahmood, A.; Ishaque, W.; Hussain, A. Foliar application of aspartic acid lowers cadmium uptake and Cd-induced oxidative stress in rice under Cd stress. Environ. Sci. Pollut. Res. 2017, 24, 21938–21947. [Google Scholar] [CrossRef]
- Zhao, L.J.; Huang, Y.X.; Paglia, K.; Vaniya, A.; Wancewicz, B.; Keller, A.A. Metabolomics reveals the molecular mechanisms of copper induced cucumber leaf (Cucumis sativus) senescence. Environ. Sci. Technol. 2018, 52, 7092–7100. [Google Scholar] [CrossRef]
- Yuan, K.; Wang, C.R.; Zhang, C.B.; Huang, Y.C.; Wang, P.P.; Liu, Z.Q. Rice grains alleviate cadmium toxicity by expending glutamate and increasing manganese in the cadmium contaminated farmland. Environ. Pollut. 2020, 262, 114236. [Google Scholar] [CrossRef]
- Na, G.; Salt, D.E. The role of sulfur assimilation and sulfur-containing compounds in trace element homeostasis in plants. Environ. Exp. Bot. 2011, 72, 18–25. [Google Scholar] [CrossRef]
- Rajab, H.; Khan, M.S.; Wirtz, M.; Malagoli, M.; Qahar, F.; Hell, R. Sulfur metabolic engineering enhances cadmium stress tolerance and root to shoot iron translocation in Brassica napus L. Plant Physiol. Biochem. 2020, 152, 32–43. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | T0 | T1 | T2 | T3 | T4 |
|---|---|---|---|---|---|
| Elements in shoots | |||||
| K (g/kg) | 49.6 ± 1.46 a | 50.1 ± 2.39 a | 44.6 ± 0.66 a | 46.1 ± 2.47 a | 47.4 ± 3.47 a |
| Mg (g/kg) | 4.16 ± 0.14 a | 4.38 ± 0.30 a | 4.19 ± 0.21 a | 4.25 ± 0.18 a | 4.49 ± 0.39 a |
| Ca (g/kg) | 5.53 ± 0.24 a | 5.43 ± 0.30 a | 5.79 ± 0.29 a | 6.21 ± 0.37 a | 6.20 ± 0.13 a |
| Fe (g/kg) | 0.29 ± 0.03 a | 0.22 ± 0.02 a | 0.40 ± 0.03 a | 0.28 ± 0.01 a | 0.36 ± 0.03 a |
| Mn (mg/kg) | 149.0 ± 0.27 b | 152.2 ± 7.13 b | 197.1 ± 5.13 a | 198.4 ± 19.14 a | 187.1 ± 7.78 ab |
| Zn (mg/kg) | 31.8 ± 1.04 a | 33.7 ± 0.43 a | 32.6 ± 2.82 a | 33.8 ± 0.23 a | 33.1 ± 0.47 a |
| Elements in roots | |||||
| K (g/kg) | 16.6 ± 0.61 a | 12.3 ± 1.37 b | 9.71 ± 1.68 bc | 8.99 ± 0.18 c | 13.5 ± 0.99 b |
| Mg (g/kg) | 0.70 ± 0.06 ab | 0.69 ± 0.06 ab | 0.68 ± 0.03 b | 0.66 ± 0.06 b | 0.82 ± 0.06 a |
| Ca (g/kg) | 2.92 ± 0.04 c | 2.77 ± 0.08 c | 5.17 ± 0.41 a | 3.58 ± 0.22 b | 3.66 ± 0.23 b |
| Fe (g/kg) | 4.63 ± 0.39 ab | 4.19 ± 0.05 b | 5.36 ± 0.38 ab | 6.18 ± 0.36 a | 5.39 ± 0.04 ab |
| Mn (mg/kg) | 137.8 ± 13.63 a | 555.3 ± 21.63 b | 624.6 ± 2.35 b | 804.1 ± 16.48 a | 758.9 ± 55.67 a |
| Zn (mg/kg) | 65.3 ± 3.94 b | 75.9 ± 4.47 ab | 69.7 ± 4.64 ab | 83.9 ± 4.97 a | 62.4 ± 2.05 b |
| Treatments | T0 | T1 | T2 | T3 | T4 |
|---|---|---|---|---|---|
| Elements in shoots | |||||
| K (g/kg) | 51.0 ± 1.54 a | 46.8 ± 4.00 ab | 46.7 ± 1.99 ab | 44.6 ± 3.40 b | 44.0 ± 3.86 b |
| Mg (g/kg) | 4.17 ± 0.18 ab | 4.61 ± 0.12 a | 4.25 ± 0.17 b | 4.20 ± 0.29 b | 3.70 ± 0.09 b |
| Ca (g/kg) | 6.51 ± 0.06 ab | 7.25 ± 0.44 a | 6.01 ± 0.01 b | 6.80 ± 0.43 ab | 5.74 ± 0.18 b |
| Fe (g/kg) | 0.23 ± 0.02 ab | 0.23 ± 0.02 ab | 0.24 ± 0.02 a | 0.22 ± 0.02 ab | 0.20 ± 0.02 ab |
| Mn (mg/kg) | 140.3 ± 12.63 bc | 185.9 ± 8.18 a | 150.8 ± 1.74 b | 170.6 ± 10.51 a | 121.7 ± 3.98 c |
| Zn (mg/kg) | 31.0 ± 1.75 bc | 31.2 ± 3.09 bc | 35.7 ± 3.12 a | 33.6 ± 1.85 ab | 29.4 ± 1.38 c |
| Elements in roots | |||||
| K (g/kg) | 21.9 ± 0.31 a | 18.2 ± 0.83 ab | 15.0 ± 1.10 b | 12.6 ± 0.47 b | 14.7 ± 0.75 b |
| Mg (g/kg) | 0.82 ± 0.04 a | 0.82 ± 0.03 a | 0.68 ± 0.01 b | 0.70 ± 0.01 ab | 0.78 ± 0.01 ab |
| Ca(g/kg) | 2.36 ± 0.13 c | 2.76 ± 0.24 bc | 2.74 ± 0.09 ab | 3.41 ± 0.18 ab | 3.45 ± 0.25 a |
| Fe (g/kg) | 3.91 ± 0.34 b | 4.97 ± 0.34 ab | 6.72 ± 0.18 a | 6.90 ± 0.07 ab | 6.20 ± 0.09 a |
| Mn (mg/kg) | 145.3 ± 7.45 d | 785.5 ± 35.69 bc | 982.4 ± 51.55 a | 936.4 ± 72.72 | 962.0 ± 33.99 ab |
| Zn (mg/kg) | 64.5 ± 5.60 d | 74.3 ± 3.02 c | 90.6 ± 2.20 a | 82.2 ± 3.73 b | 59.7 ± 3.24 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Fu, L.; Zhang, C.; Deng, J.; Xue, W.; Deng, Y. Effects of Exogenous Chlorinated Amino Acetic Acid on Cadmium and Mineral Elements in Rice Seedlings. Toxics 2023, 11, 71. https://doi.org/10.3390/toxics11010071
Liu S, Fu L, Zhang C, Deng J, Xue W, Deng Y. Effects of Exogenous Chlorinated Amino Acetic Acid on Cadmium and Mineral Elements in Rice Seedlings. Toxics. 2023; 11(1):71. https://doi.org/10.3390/toxics11010071
Chicago/Turabian StyleLiu, Shuangyue, Lin Fu, Changbo Zhang, Jiawei Deng, Weijie Xue, and Yun Deng. 2023. "Effects of Exogenous Chlorinated Amino Acetic Acid on Cadmium and Mineral Elements in Rice Seedlings" Toxics 11, no. 1: 71. https://doi.org/10.3390/toxics11010071
APA StyleLiu, S., Fu, L., Zhang, C., Deng, J., Xue, W., & Deng, Y. (2023). Effects of Exogenous Chlorinated Amino Acetic Acid on Cadmium and Mineral Elements in Rice Seedlings. Toxics, 11(1), 71. https://doi.org/10.3390/toxics11010071