
Remediation Agents Drive Bacterial Community in a Cd-Contaminated Soil



Abstract
1. Introduction
2. Materials and Methods
2.1. Soil, Plants and Amendments
2.2. Experimental Design and Sample Collection
2.3. Analysis of Soil and Plant Samples
2.4. Analysis of Soil Bacterial Communities
2.5. Data Analysis
3. Results
3.1. Soil Properties and Cd Content in Plants
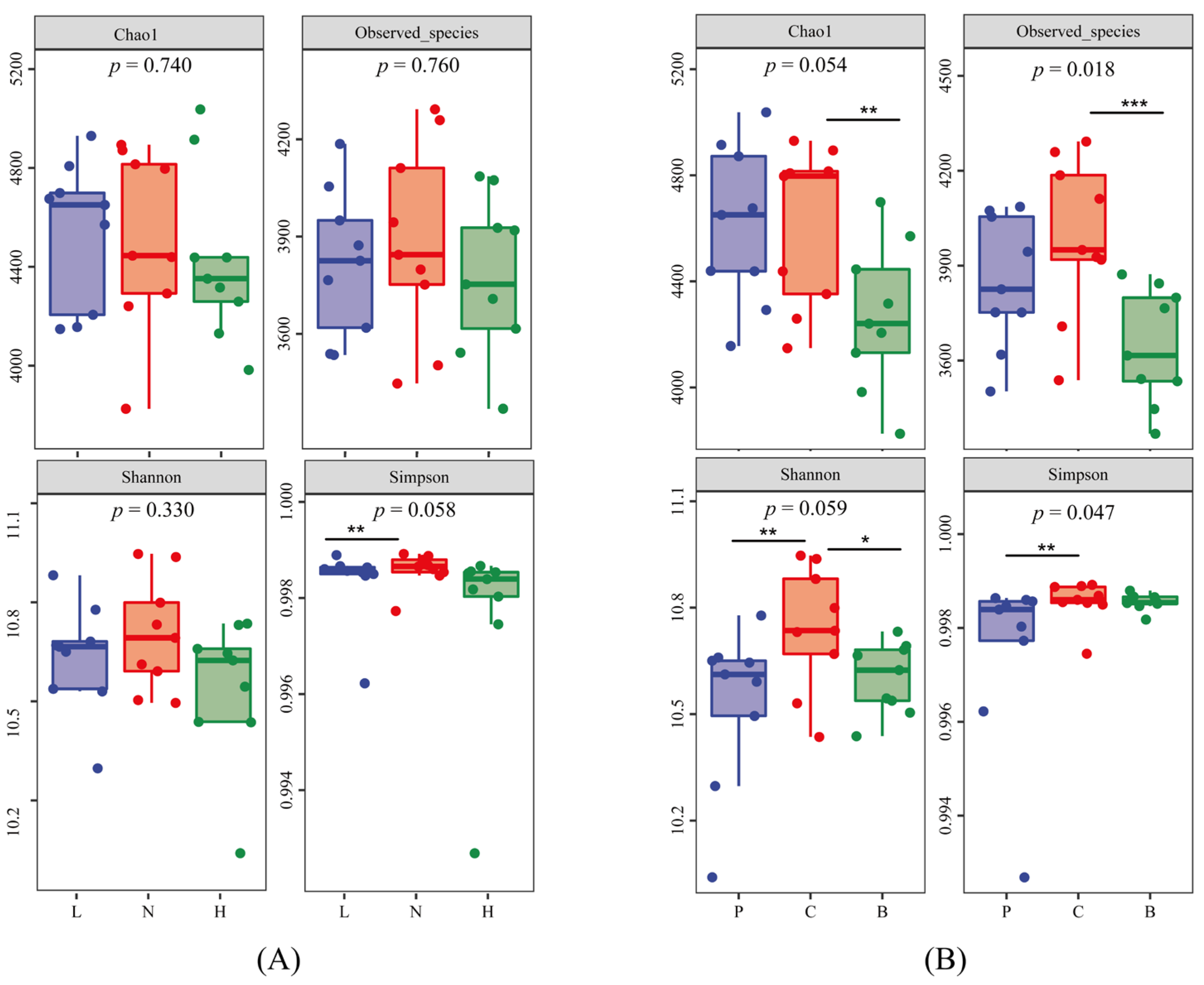
3.2. Soil Bacterial Communities Structure and Diversity
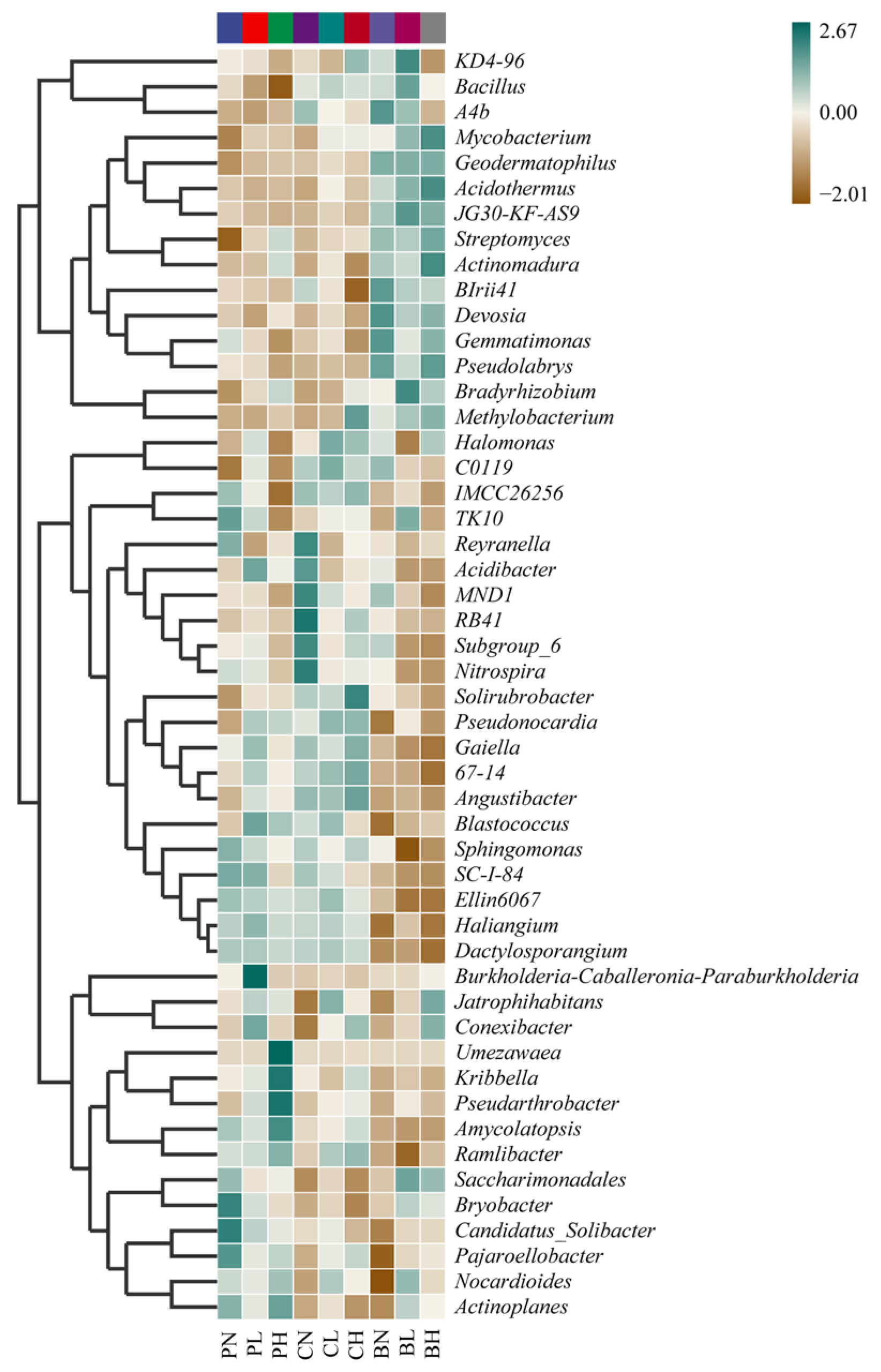
3.3. Effects of SRAs and Plant Cultivation on Bacterial Community Composition
3.4. MetagenomeSeq Analysis
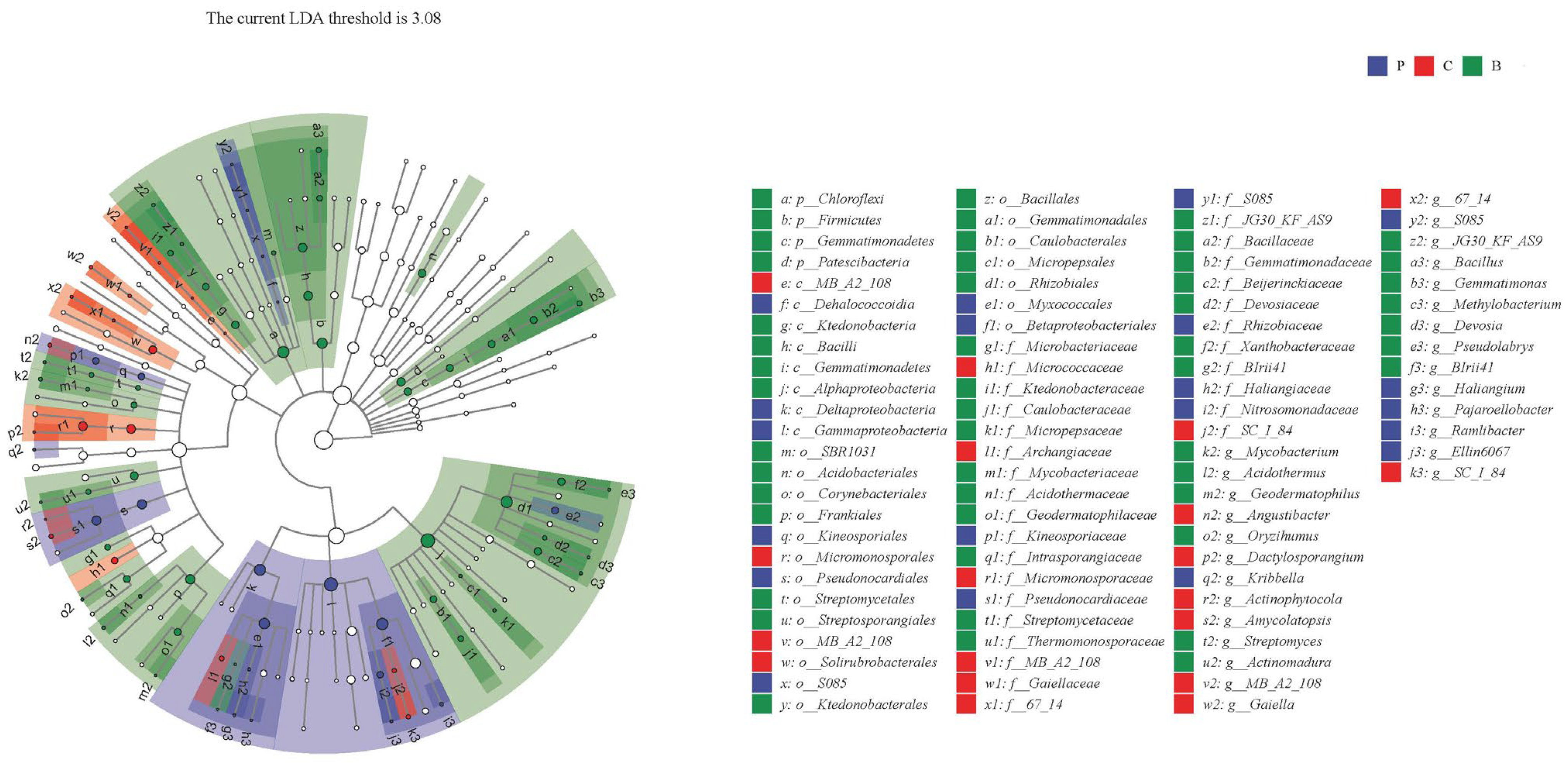
3.5. LEfSe Analysis
3.6. Correlations between Soil Properties and Bacterial Abundance
4. Discussion
4.1. Effects of Biochar and HAP on Bacterial Communities by Altering Soil Properties
4.2. Effects of Planting Density on Soil Bacterial Communities
4.3. Effects of SRAs and Plant Cultivation on Specific Bacteria
4.4. Potential Functions of Marker Species in Soil C and P Cycling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, F.-J.; Wang, P. Arsenic and cadmium accumulation in rice and mitigation strategies. Plant Soil 2019, 446, 1–21. [Google Scholar] [CrossRef]
- Wang, P.; Chen, H.; Kopittke, P.M.; Zhao, F.J. Cadmium contamination in agricultural soils of China and the impact on food safety. Environ. Pollut. 2019, 249, 1038–1048. [Google Scholar] [CrossRef]
- Ayangbenro, A.S.; Babalola, O.O. A new strategy for heavy metal polluted environments: A review of microbial biosorbents. Int. J. Environ. Res. Public Health 2017, 14, 94. [Google Scholar] [CrossRef]
- Chen, D.; Liu, X.Y.; Bian, R.J.; Cheng, K.; Zhang, X.H.; Zheng, J.F.; Joseph, S.; Crowley, D.; Pan, G.X.; Li, L.Q. Effects of biochar on availability and plant uptake of heavy metals—A meta-analysis. J. Environ. Manag. 2018, 222, 76–85. [Google Scholar] [CrossRef]
- He, L.; Zhong, H.; Liu, G.; Dai, Z.; Brookes, P.C.; Xu, J. Remediation of heavy metal contaminated soils by biochar: Mechanisms, potential risks and applications in China. Environ. Pollut. 2019, 252, 846–855. [Google Scholar] [CrossRef]
- Li, H.; Liu, Y.; Tang, S.; Yu, Z.; Cai, X.; Xu, S.; Chen, Y.; Wang, M.; Wang, G. Mechanisms for potential Pb immobilization by hydroxyapatite in a soil-rice system. Sci. Total Environ. 2021, 783, 147037. [Google Scholar] [CrossRef]
- Cui, H.; Bao, B.; Cao, Y.; Zhang, S.; Shi, J.; Zhou, J.; Zhou, J. Combined application of ferrihydrite and hydroxyapatite to immobilize soil copper, cadmium, and phosphate under flooding-drainage alternations. Environ. Pollut. 2021, 292, 118323. [Google Scholar] [CrossRef]
- Cui, H.; Shi, Y.; Zhou, J.; Chu, H.; Cang, L.; Zhou, D. Effect of different grain sizes of hydroxyapatite on soil heavy metal bioavailability and microbial community composition. Agric. Ecosys. Environ. 2018, 267, 165–173. [Google Scholar] [CrossRef]
- Hamid, Y.; Tang, L.; Sohail, M.I.; Cao, X.R.; Hussain, B.; Aziz, M.Z.; Usman, M.; He, Z.L.; Yang, X.E. An explanation of soil amendments to reduce cadmium phytoavailability and transfer to food chain. Sci. Total Environ. 2019, 660, 80–96. [Google Scholar] [CrossRef]
- Gholizadeh, M.; Hu, X. Removal of heavy metals from soil with biochar composite: A critical review of the mechanism. J. Environ. Chem. Eng. 2021, 9, 105830. [Google Scholar] [CrossRef]
- Ghorbani, M.; Konvalina, P.; Neugschwandtner, R.W.; Kopecky, M.; Amirahmadi, E.; Moudry, J., Jr.; Mensik, L. Preliminary findings on cadmium bioaccumulation and photosynthesis in rice (Oryza sativa L.) and maize (Zea mays L.) using biochar made from C3- and C4-originated straw. Plants 2022, 11, 1424. [Google Scholar] [CrossRef]
- Mignardi, S.; Corami, A.; Ferrini, V. Evaluation of the effectiveness of phosphate treatment for the remediation of mine waste soils contaminated with Cd, Cu, Pb, and Zn. Chemosphere 2012, 86, 354–360. [Google Scholar] [CrossRef]
- Lee, H.H.; Owens, V.N.; Park, S.; Kim, J.; Hong, C.O. Adsorption and precipitation of cadmium affected by chemical form and addition rate of phosphate in soils having different levels of cadmium. Chemosphere 2018, 206, 369–375. [Google Scholar] [CrossRef]
- Sun, R.-J.; Chen, J.-H.; Fan, T.-T.; Zhou, D.-M.; Wang, Y.-J. Effect of nanoparticle hydroxyapatite on the immobilization of Cu and Zn in polluted soil. Environ. Sci. Pollut. Res. 2018, 25, 73–80. [Google Scholar] [CrossRef]
- Zeng, G.; Wan, J.; Huang, D.; Hu, L.; Huang, C.; Cheng, M.; Xue, W.; Gong, X.; Wang, R.; Jiang, D. Precipitation, adsorption and rhizosphere effect: The mechanisms for phosphate-induced Pb immobilization in soils—A review. J. Hazard. Mater. 2017, 339, 354–367. [Google Scholar] [CrossRef]
- Dai, Z.; Xiong, X.; Zhu, H.; Xu, H.; Leng, P.; Li, J.; Tang, C.; Xu, J. Association of biochar properties with changes in soil bacterial, fungal and fauna communities and nutrient cycling processes. Biochar 2021, 3, 239–254. [Google Scholar] [CrossRef]
- Sun, D.; Meng, J.; Xu, E.G.; Chen, W. Microbial community structure and predicted bacterial metabolic functions in biochar pellets aged in soil after 34 months. Appl. Soil Ecol. 2016, 100, 135–143. [Google Scholar] [CrossRef]
- Khodadad, C.L.M.; Zimmerman, A.R.; Green, S.J.; Uthandi, S.; Foster, J.S. Taxa-specific changes in soil microbial community composition induced by pyrogenic carbon amendments. Soil Biol. Biochem. 2011, 43, 385–392. [Google Scholar] [CrossRef]
- Yu, L.; Bai, J.; Huang, L.; Zhang, G.; Wang, W.; Wang, X.; Yu, Z. Carbon-rich substrates altered microbial communities with indication of carbon metabolism functional shifting in a degraded salt marsh of the Yellow River Delta, China. J. Clean. Prod. 2022, 331, 129898. [Google Scholar] [CrossRef]
- Mackie, K.A.; Marhan, S.; Ditterich, F.; Schmidt, H.P.; Kandeler, E. The effects of biochar and compost amendments on copper immobilization and soil microorganisms in a temperate vineyard. Agric. Ecosyst. Environ. 2015, 201, 58–69. [Google Scholar] [CrossRef]
- Meschewski, E.; Holm, N.; Sharma, B.K.; Spokas, K.; Minalt, N.; Kelly, J.J. Pyrolysis biochar has negligible effects on soil greenhouse gas production, microbial communities, plant germination, and initial seedling growth. Chemosphere 2019, 228, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Meng, J.; Liang, H.; Yang, E.; Huang, Y.; Chen, W.; Jiang, L.; Lan, Y.; Zhang, W.; Gao, J. Effect of volatile organic compounds absorbed to fresh biochar on survival of Bacillus mucilaginosus and structure of soil microbial communities. J. Soils Sediments 2014, 15, 271–281. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota–a review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dynamics of Gemmatimonadetes in soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Yi, Q.; Yang, X.; Wang, X.; Wu, H.; Zhou, J. Effects of hydroxyapatite on leaching of cadmium and phosphorus and their availability under simulated acid rain. J. Environ. Chem. Eng. 2017, 5, 3773–3779. [Google Scholar] [CrossRef]
- Cheng, J.; Li, Y.; Gao, W.; Chen, Y.; Pan, W.; Lee, X.; Tang, Y. Effects of biochar on Cd and Pb mobility and microbial community composition in a calcareous soil planted with tobacco. Biol. Fertil. Soils 2018, 54, 373–383. [Google Scholar] [CrossRef]
- Bourhane, Z.; Lanzén, A.; Cagnon, C.; Said, O.B.; Mahmoudi, E.; Coulon, F.; Atai, E.; Borja, A.; Cravo-Laureau, C.; Duran, R. Microbial diversity alteration reveals biomarkers of contamination in soil-river-lake continuum. J. Hazard. Mater. 2022, 421, 126789. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, B.; Zhu, L.; Xing, B. Effects and mechanisms of biochar-microbe interactions in soil improvement and pollution remediation: A review. Environ. Pollut. 2017, 227, 98–115. [Google Scholar] [CrossRef]
- Ennis, C.J.; Evans, A.G.; Islam, M.; Ralebitso-Senior, T.K.; Senior, E. Biochar: Carbon sequestration, land remediation, and impacts on soil microbiology. Crit. Rev. Environ. Sci. Technol. 2012, 42, 2311–2364. [Google Scholar] [CrossRef]
- Palansooriya, K.N.; Wong, J.T.F.; Hashimoto, Y.; Huang, L.; Rinklebe, J.; Chang, S.X.; Bolan, N.; Wang, H.; Ok, Y.S. Response of microbial communities to biochar-amended soils: A critical review. Biochar 2019, 1, 3–22. [Google Scholar] [CrossRef]
- Feng, Y.; Yang, J.; Liu, W.; Yan, Y.; Wang, Y. Hydroxyapatite as a passivator for safe wheat production and its impacts on soil microbial communities in a Cd-contaminated alkaline soil. J. Hazard. Mater. 2021, 404, 124005. [Google Scholar] [CrossRef]
- Qin, Y.; Shi, X.; Wang, Z.; Pei, C.; Cao, M.; Luo, J. Influence of planting density on the phytoremediation efficiency of Festuca arundinacea in Cd-polluted soil. Bull. Environ. Contam. Toxicol. 2021, 107, 154–159. [Google Scholar] [CrossRef]
- Wang, L.; Hou, D.; Shen, Z.; Zhu, J.; Jia, X.; Ok, Y.S.; Tack, F.M.; Rinklebe, J. Field trials of phytomining and phytoremediation: A critical review of influencing factors and effects of additives. Crit. Rev. Environ. Sci. Technol. 2020, 50, 2724–2774. [Google Scholar] [CrossRef]
- Wang, F.; Cheng, P.; Zhang, S.; Zhang, S.; Sun, Y. Contribution of arbuscular mycorrhizal fungi and soil amendments to remediation of heavy metal-contaminated soil using sweet sorghum. Pedosphere 2022, 32, 844–855. [Google Scholar] [CrossRef]
- Dar, R.A.; Dar, E.A.; Kaur, A.; Phutela, U.G. Sweet sorghum-a promising alternative feedstock for biofuel production. Renew. Sustain. Energy Rev. 2018, 82, 4070–4090. [Google Scholar] [CrossRef]
- Zhang, G.; Gong, Z. Soil Survey Laboratory Methods; Science Press: Beijing, China, 2012. [Google Scholar]
- Chang, E.; Chiang, P.-C.; Lu, P.-H.; Ko, Y.-W. Comparisons of metal leachability for various wastes by extraction and leaching methods. Chemosphere 2001, 45, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Quilliam, R.S.; Glanville, H.C.; Wade, S.C.; Jones, D.L. Life in the ‘charosphere’–Does biochar in agricultural soil provide a significant habitat for microorganisms? Soil Biol. Biochem. 2013, 65, 287–293. [Google Scholar] [CrossRef]
- Gul, S.; Whalen, J.K.; Thomas, B.W.; Sachdeva, V.; Deng, H. Physico-chemical properties and microbial responses in biochar-amended soils: Mechanisms and future directions. Agric. Ecosyst. Environ. 2015, 206, 46–59. [Google Scholar] [CrossRef]
- Rawat, J.; Saxena, J.; Sanwal, P. Biochar: A Sustainable Approach for Improving Plant Growth and Soil Properties. In Biochar—An Imperative Amendment for Soil and the Environment; Abrol, V., Sharma, P., Eds.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Obayomi, O.; Seyoum, M.M.; Ghazaryan, L.; Tebbe, C.C.; Murase, J.; Bernstein, N.; Gillor, O. Soil texture and properties rather than irrigation water type shape the diversity and composition of soil microbial communities. Appl. Soil Ecol. 2021, 161, 103834. [Google Scholar] [CrossRef]
- Tripathi, B.M.; Stegen, J.C.; Kim, M.; Dong, K.; Adams, J.M.; Lee, Y.K. Soil pH mediates the balance between stochastic and deterministic assembly of bacteria. ISME J. 2018, 12, 1072–1083. [Google Scholar] [CrossRef]
- Rousk, J.; Baath, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Xu, D.; Yuan, H.; Wu, B.; Chen, B.; Tan, Y.; Lin, J.; Guo, D. Investigation on the association of soil microbial populations with ecological and environmental factors in the Pearl River estuary. J. Geosci. Environ. Prot. 2018, 6, 8–14. [Google Scholar] [CrossRef][Green Version]
- Zeng, T.; Wang, L.; Zhang, X.; Song, X.; Li, J.; Yang, J.; Chen, S.; Zhang, J. Characterization of microbial communities in wastewater treatment plants containing heavy metals located in chemical industrial zones. Int. J. Environ. Res. Public Health 2022, 19, 6529. [Google Scholar] [CrossRef] [PubMed]
- Zhalnina, K.; De Quadros, P.D.; Gano, K.A.; Davis-Richardson, A.; Fagen, J.R.; Brown, C.T.; Giongo, A.; Drew, J.C.; Sayavedra-Soto, L.A.; Arp, D.J.; et al. Ca. Nitrososphaera and Bradyrhizobium are inversely correlated and related to agricultural practices in long-term field experiments. Front. Microbiol. 2013, 4, 104. [Google Scholar] [CrossRef]
- Xu, M.; Hao, X.; Xiong, Z.; Liao, H.; Wang, L.; Zhang, T.; Luo, X.; Chen, W.; Huang, Q. Soil amendments change bacterial functional genes more than taxonomic structure in a cadmium-contaminated soil. Soil Biol. Biochem. 2021, 154, 108126. [Google Scholar] [CrossRef]
- Nguyen, M.K.; Lin, C.; Hoang, H.G.; Sanderson, P.; Dang, B.T.; Bui, X.T.; Nguyen, N.S.H.; Vo, D.-V.N.; Tran, H.T. Evaluate the role of biochar during the organic waste composting process: A critical review. Chemosphere 2022, 299, 134488. [Google Scholar] [CrossRef]
- Yang, W.; Li, C.; Wang, S.; Zhou, B.; Mao, Y.; Rensing, C.; Xing, S. Influence of biochar and biochar-based fertilizer on yield, quality of tea and microbial community in an acid tea orchard soil. Appl. Soil Ecol. 2021, 166, 104005. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, S.; Cheng, P.; Zhang, S.; Sun, Y. Effects of soil amendments on heavy metal immobilization and accumulation by maize grown in a multiple-metal-contaminated soil and their potential for safe crop production. Toxics 2020, 8, 102. [Google Scholar] [CrossRef]
- Chintala, R.; Mollinedo, J.; Schumacher, T.E.; Malo, D.D.; Julson, J.L. Effect of biochar on chemical properties of acidic soil. Arch. Agron. Soil Sci. 2014, 60, 393–404. [Google Scholar] [CrossRef]
- Freddo, A.; Cai, C.; Reid, B.J. Environmental contextualisation of potential toxic elements and polycyclic aromatic hydrocarbons in biochar. Environ. Pollut. 2012, 171, 18–24. [Google Scholar] [CrossRef]
- Fabbri, D.; Rombolà, A.G.; Torri, C.; Spokas, K.A. Determination of polycyclic aromatic hydrocarbons in biochar and biochar amended soil. J. Analyt. Appl. Pyrolysis 2013, 103, 60–67. [Google Scholar] [CrossRef]
- Kusmierz, M.; Oleszczuk, P.; Kraska, P.; Palys, E.; Andruszczak, S. Persistence of polycyclic aromatic hydrocarbons (PAHs) in biochar-amended soil. Chemosphere 2016, 146, 272–279. [Google Scholar] [CrossRef]
- Odinga, E.S.; Gudda, F.O.; Waigi, M.G.; Wang, J.; Gao, Y. Occurrence, formation and environmental fate of polycyclic aromatic hydrocarbons in biochars. Fundam. Res. 2021, 1, 296–305. [Google Scholar] [CrossRef]
- Dürre, P. Clostridia. In Encyclopedia of Life Sciences; Wiley: Hoboken, NJ, USA, 2007. [Google Scholar] [CrossRef]
- Sall, S.N.; Masse, D.; Ndour, N.Y.B.; Chotte, J.-L. Does cropping modify the decomposition function and the diversity of the soil microbial community of tropical fallow soil? Appl. Soil Ecol. 2006, 31, 211–219. [Google Scholar] [CrossRef]
- Zong, N.; Shi, P. Soil properties rather than plant production strongly impact soil bacterial community diversity along a desertification gradient on the Tibetan Plateau. Grassl. Sci. 2020, 66, 197–206. [Google Scholar] [CrossRef]
- Warnock, D.D.; Lehmann, J.; Kuyper, T.W.; Rillig, M.C. Mycorrhizal responses to biochar in soil—concepts and mechanisms. Plant Soil 2007, 300, 9–20. [Google Scholar] [CrossRef]
- Chen, J.; Liu, X.; Zheng, J.; Zhang, B.; Lu, H.; Chi, Z.; Pan, G.; Li, L.; Zheng, J.; Zhang, X.; et al. Biochar soil amendment increased bacterial but decreased fungal gene abundance with shifts in community structure in a slightly acid rice paddy from Southwest China. Appl. Soil Ecol. 2013, 71, 33–44. [Google Scholar] [CrossRef]
- Jezequel, K.; Perrin, J.; Lebeau, T. Bioaugmentation with a Bacillus sp. to reduce the phytoavailable Cd of an agricultural soil: Comparison of free and immobilized microbial inocula. Chemosphere 2005, 59, 1323–1331. [Google Scholar] [CrossRef]
- Maity, J.P.; Chen, G.-S.; Huang, Y.-H.; Sun, A.-C.; Chen, C.-Y. Ecofriendly heavy metal stabilization: Microbial induced mineral precipitation (MIMP) and biomineralization for heavy metals within the contaminated soil by indigenous bacteria. Geomicrobiol. J. 2019, 36, 612–623. [Google Scholar] [CrossRef]
- Liu, X.; Chen, C.; Wang, J.; Zou, S.; Long, X. Phosphorus solubilizing bacteria Bacillus thuringiensis and Pantoea ananatis simultaneously promote soil inorganic phosphate dissolution and soil Pb immobilization. Rhizosphere 2021, 20, 100448. [Google Scholar] [CrossRef]
- Seneviratne, M.; Gunaratne, S.; Bandara, T.; Weerasundara, L.; Rajakaruna, N.; Seneviratne, G.; Vithanage, M. Plant growth promotion by Bradyrhizobium japonicum under heavy metal stress. S. Afr. J. Bot. 2016, 105, 19–24. [Google Scholar] [CrossRef]
- Ali, A.; Guo, D.; Li, Y.; Shaheen, S.M.; Wahid, F.; Antoniadis, V.; Abdelrahman, H.; Al-Solaimani, S.G.; Li, R.; Tsang, D.C.W.; et al. Streptomyces pactum addition to contaminated mining soils improved soil quality and enhanced metals phytoextraction by wheat in a green remediation trial. Chemosphere 2021, 273, 129692. [Google Scholar] [CrossRef]
- Zloch, M.; Thiem, D.; Gadzala-Kopciuch, R.; Hrynkiewicz, K. Synthesis of siderophores by plant-associated metallotolerant bacteria under exposure to Cd2+. Chemosphere 2016, 156, 312–325. [Google Scholar] [CrossRef]
- Madhaiyan, M.; Poonguzhali, S.; Sa, T. Metal tolerating methylotrophic bacteria reduces nickel and cadmium toxicity and promotes plant growth of tomato (Lycopersicon esculentum L.). Chemosphere 2007, 69, 220–228. [Google Scholar] [CrossRef]
- Cui, H.; Liu, L.L.; Dai, J.R.; Yu, X.N.; Guo, X.; Yi, S.J.; Zhou, D.Y.; Guo, W.H.; Du, N. Bacterial community shaped by heavy metals and contributing to health risks in cornfields. Ecotoxicol. Environ. Saf. 2018, 166, 259–269. [Google Scholar] [CrossRef]
- Feng, X.; Wang, Q.; Sun, Y.; Zhang, S.; Wang, F. Microplastics change soil properties, heavy metal availability and bacterial community in a Pb-Zn-contaminated soil. J. Hazard. Mater. 2022, 424, 127364. [Google Scholar] [CrossRef]
- Wu, B.; Luo, S.; Luo, H.; Huang, H.; Xu, F.; Feng, S.; Xu, H. Improved phytoremediation of heavy metal contaminated soils by Miscanthus floridulus under a varied rhizosphere ecological characteristic. Sci. Total Environ. 2022, 808, 151995. [Google Scholar] [CrossRef]
- Gołębiewski, M.; Deja-Sikora, E.; Cichosz, M.; Tretyn, A.; Wróbel, B. 16S rDNA pyrosequencing analysis of bacterial community in heavy metals polluted soils. Microb. Ecol. 2014, 67, 635–647. [Google Scholar] [CrossRef]
- Wang, H.; Zeng, Y.; Guo, C.; Bao, Y.; Lu, G.; Reinfelder, J.R.; Dang, Z. Bacterial, archaeal, and fungal community responses to acid mine drainage-laden pollution in a rice paddy soil ecosystem. Sci. Total Environ. 2018, 616, 107–116. [Google Scholar] [CrossRef]
- Li, B.; Xu, R.; Sun, X.; Han, F.; Xiao, E.; Chen, L.; Qiu, L.; Sun, W. Microbiome–environment interactions in antimony-contaminated rice paddies and the correlation of core microbiome with arsenic and antimony contamination. Chemosphere 2021, 263, 128227. [Google Scholar] [CrossRef]
- Bian, F.; Zhong, Z.; Zhang, X.; Li, Q.; Huang, Z. Bamboo-based agroforestry changes phytoremediation efficiency by affecting soil properties in rhizosphere and non-rhizosphere in heavy metal-polluted soil (Cd/Zn/Cu). J. Soils Sediments 2022, 23, 368–378. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, Q.; Du, W.; Lin, R.; Li, J.; Ai, F.; Yin, Y.; Ji, R.; Wang, X.; Guo, H. In-situ immobilization of cadmium-polluted upland soil: A ten-year field study. Ecotoxicol. Environ. Saf. 2021, 207, 111275. [Google Scholar] [CrossRef]
- Li, Y.; Gong, X.; Xiong, J.; Sun, Y.; Shu, Y.; Niu, D.; Lin, Y.; Wu, L.; Zhang, R. Different dissolved organic matters regulate the bioavailability of heavy metals and rhizosphere microbial activity in a plant-wetland soil system. J. Environ. Chem. Eng. 2021, 9, 106823. [Google Scholar] [CrossRef]
- Singh, K.N.; Narzary, D. Heavy metal tolerance of bacterial isolates associated with overburden strata of an opencast coal mine of Assam (India). Environ. Sci. Pollut. Res. 2021, 28, 63111–63126. [Google Scholar] [CrossRef]
- Desale, P.; Patel, B.; Singh, S.; Malhotra, A.; Nawani, N. Plant growth promoting properties of Halobacillus sp. and Halomonas sp. in presence of salinity and heavy metals. J. Basic Microbiol. 2014, 54, 781–791. [Google Scholar] [CrossRef]
- Větrovský, T.; Baldrian, P. An in-depth analysis of actinobacterial communities shows their high diversity in grassland soils along a gradient of mixed heavy metal contamination. Biol. Fertil. Soils 2015, 51, 827–837. [Google Scholar] [CrossRef]
- Luo, Z.; Ma, J.; Chen, F.; Li, X.; Zhang, S. Effects of Pb smelting on the soil bacterial community near a secondary lead plant. Int. J. Environ. Res. Public Health 2018, 15, 1030. [Google Scholar] [CrossRef]
- Zhang, S.; Fang, Y.; Luo, Y.; Li, Y.; Ge, T.; Wang, Y.; Wang, H.; Yu, B.; Song, X.; Chen, J.; et al. Linking soil carbon availability, microbial community composition and enzyme activities to organic carbon mineralization of a bamboo forest soil amended with pyrogenic and fresh organic matter. Sci. Total Environ. 2021, 801, 149717. [Google Scholar] [CrossRef]
- Zheng, Y.; Maruoka, M.; Nanatani, K.; Hidaka, M.; Abe, N.; Kaneko, J.; Sakai, Y.; Abe, K.; Yokota, A.; Yabe, S. High cellulolytic potential of the Ktedonobacteria lineage revealed by genome-wide analysis of CAZymes. J. Biosci. Bioengin. 2021, 131, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.; Lin, Y.; Liu, D.; Chen, Z.; Luo, J.; Bolan, N.; Fan, J.; Ding, W. Long-term application of manure over plant residues mitigates acidification, builds soil organic carbon and shifts prokaryotic diversity in acidic Ultisols. Appl. Soil Ecol. 2019, 133, 24–33. [Google Scholar] [CrossRef]
- Li, F.; Chen, L.; Zhang, J.; Yin, J.; Huang, S. Bacterial community structure after long-term organic and inorganic fertilization reveals important associations between soil nutrients and specific taxa involved in nutrient transformations. Front. Microbiol. 2017, 8, 187. [Google Scholar] [CrossRef]
- Berry, A.; Normand, P. Acidothermaceae. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2020. [Google Scholar] [CrossRef]
- Asadu, C.; Aneke, N.; Agulanna, A.; Eze, C. Degradation of sawdust by thermo tolerant microorganisms for bio fertilizer synthesis. Asian J. Biotechnol. Bioresour. Technol. 2018, 2, 1–7. [Google Scholar] [CrossRef]
- Bräuer, S.; Harbison, A.; Ueki, A. Micropepsales. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2018. [Google Scholar] [CrossRef]
- Kim, D.U.; Kim, S.G.; Lee, H.; Park, A.Y.; Ka, J.O. Oryzihumus soli sp. nov., isolated from soil and emended description of the genus Oryzihumus. Int. J. System. Evol. Microbiol. 2017, 67, 3960–3964. [Google Scholar] [CrossRef]
- Bergkemper, F.; Scholer, A.; Engel, M.; Lang, F.; Kruger, J.; Schloter, M.; Schulz, S. Phosphorus depletion in forest soils shapes bacterial communities towards phosphorus recycling systems. Environ. Microbiol. 2016, 18, 1988–2000. [Google Scholar] [CrossRef] [PubMed]
- Tchuisseu Tchakounte, G.V.; Berger, B.; Patz, S.; Fankem, H.; Ruppel, S. Data on molecular identification, phylogeny and in vitro characterization of bacteria isolated from maize rhizosphere in Cameroon. Data Brief 2018, 19, 1410–1417. [Google Scholar] [CrossRef] [PubMed]
- Chatli, A.S.; Beri, V.; Sidhu, B. Isolation and characterisation of phosphate solubilising microorganisms from the cold desert habitat of Salix alba Linn. in trans Himalayan region of Himachal Pradesh. Indian J. Microbiol. 2008, 48, 267–273. [Google Scholar] [CrossRef]
- Wang, G.; Jin, Z.; Wang, X.; George, T.S.; Feng, G.; Zhang, L. Simulated root exudates stimulate the abundance of Saccharimonadales to improve the alkaline phosphatase activity in maize rhizosphere. Appl. Soil Ecol. 2022, 170, 104274. [Google Scholar] [CrossRef]
- Zhang, W.H.; Sun, R.B.; Xu, L.; Liang, J.N.; Zhou, J. Assessment of bacterial communities in Cu-contaminated soil immobilized by a one-time application of micro-/nano-hydroxyapatite and phytoremediation for 3 years. Chemosphere 2019, 223, 240–249. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | NH4+-N | NO3−-N | SOC | Available K | Available P | Total Cd | Total Zn |
|---|---|---|---|---|---|---|---|
| mg kg−1 | mg kg−1 | g kg−1 | mg kg−1 | mg kg−1 | mg kg−1 | mg kg−1 | |
| 5.04 ± 0.02 | 0.43 ± 0.19 | 3.84 ± 0.20 | 16.15 ± 0.63 | 130.83 ± 6.08 | 48.32 ± 15.28 | 0.47 ± 0.06 | 32.34 ± 7.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, W.; Liu, Y.; Li, W.; Pei, L.; Xu, S.; Sun, Y.; Liu, J.; Wang, F. Remediation Agents Drive Bacterial Community in a Cd-Contaminated Soil. Toxics 2023, 11, 53. https://doi.org/10.3390/toxics11010053
Cui W, Liu Y, Li W, Pei L, Xu S, Sun Y, Liu J, Wang F. Remediation Agents Drive Bacterial Community in a Cd-Contaminated Soil. Toxics. 2023; 11(1):53. https://doi.org/10.3390/toxics11010053
Chicago/Turabian StyleCui, Wenzhi, Yingying Liu, Wenguang Li, Lei Pei, Shuang Xu, Yuhuan Sun, Jianbo Liu, and Fayuan Wang. 2023. "Remediation Agents Drive Bacterial Community in a Cd-Contaminated Soil" Toxics 11, no. 1: 53. https://doi.org/10.3390/toxics11010053
APA StyleCui, W., Liu, Y., Li, W., Pei, L., Xu, S., Sun, Y., Liu, J., & Wang, F. (2023). Remediation Agents Drive Bacterial Community in a Cd-Contaminated Soil. Toxics, 11(1), 53. https://doi.org/10.3390/toxics11010053