
Current Progress for Retrospective Identification of Nerve Agent Biomarkers in Biological Samples after Exposure


Abstract
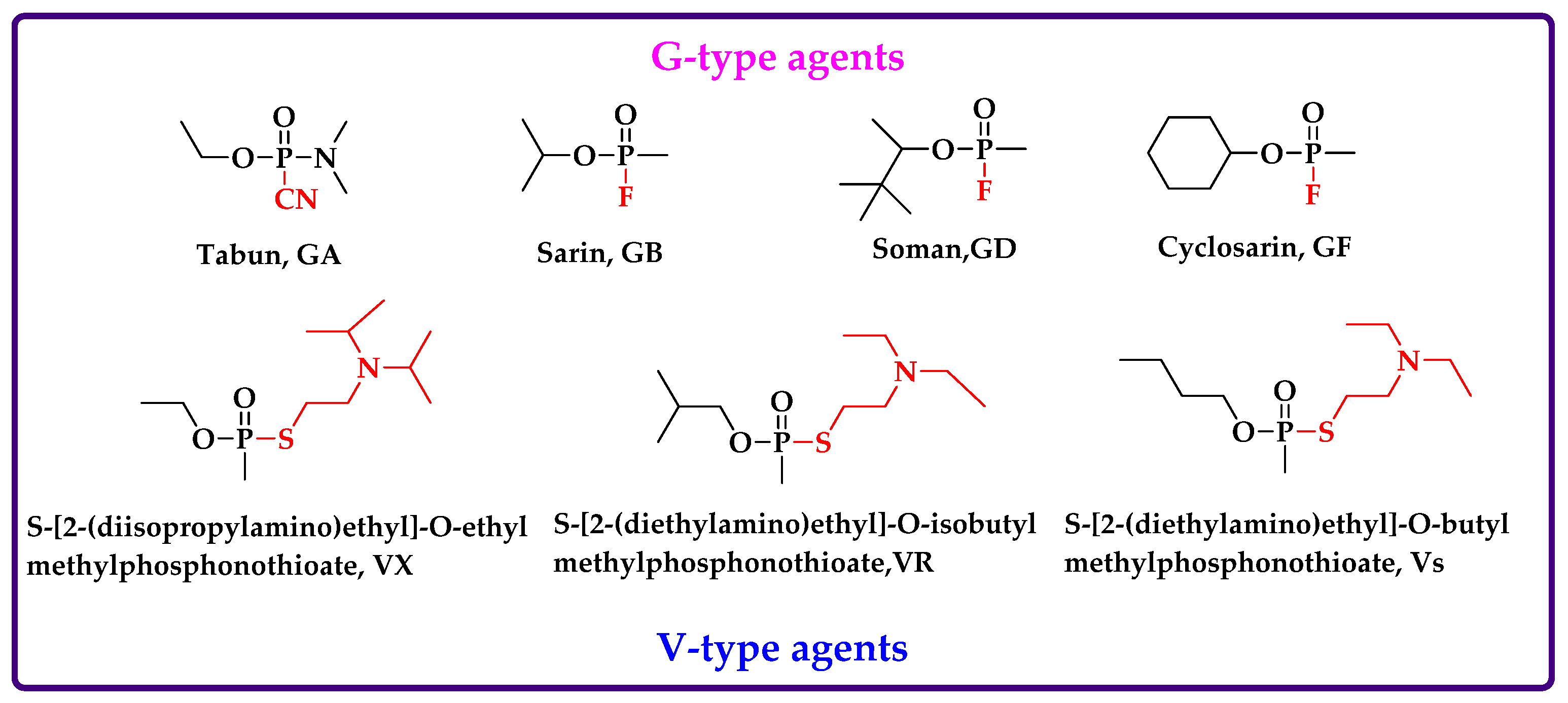
1. Introduction
2. Intact OPNAs
3. Degradation Products
4. Protein Adduct
4.1. Fluoride-Induced Reactivation
4.2. Digestive Peptides Analysis
4.2.1. AChE Adduct
4.2.2. BChE Adduct
4.2.3. Albumin Adduct
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Costanzi, S.; Machado, J.H.; Mitchell, M. Nerve Agents: What They Are, How They Work, How to Counter Them. ACS Chem. Neurosci. 2018, 9, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Weir, A.G.A.; Makin, S.; Breeze, J. Nerve agents: Emergency preparedness. BMJ Mil. Health 2020, 166, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Jett, D.A.; Sibrizzi, C.A.; Blain, R.B.; Hartman, P.A.; Lein, P.J.; Taylor, K.W.; Rooney, A.A. A national toxicology program systematic review of the evidence for long-term effects after acute exposure to sarin nerve agent. Crit. Rev. Toxicol. 2020, 50, 474–490. [Google Scholar] [CrossRef] [PubMed]
- Golime, R.; Singh, N.; Palit, M. Epigenetic and autophagic changes after nerve agent exposure in the rat piriform cortex and hippocampus. Toxicology 2019, 423, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, T.H.; Apland, J.P.; Braga, M.F.M.; Marini, A.M. Acute and long-term consequences of exposure to organophosphate nerve agents in humans. Epilepsia 2018, 59, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, J.M.; Chang, W.E.; Bah, M.J.; Wright, L.K.; Saviolakis, G.A.; Alagappan, A.; Robison, C.L.; Shah, J.D.; Meyerhoff, J.L.; Cerasoli, D.M.; et al. Repeated exposure to sublethal doses of the organophosphorus compound VX activates BDNF expression in mouse brain. Toxicol. Sci. 2012, 126, 497–505. [Google Scholar] [CrossRef]
- Wiercinski, A.; Jackson, J.P. Nerve Agents. In StatPearls; StatPearls Publishing LLC: Treasure Island, FL, USA, 2022. [Google Scholar]
- Greenfield, R.A.; Brown, B.R.; Hutchins, J.B.; Iandolo, J.J.; Jackson, R.; Slater, L.N.; Bronze, M.S. Microbiological, biological, and chemical weapons of warfare and terrorism. Am. J. Med. Sci. 2002, 323, 326–340. [Google Scholar] [CrossRef]
- Stone, R. Obscure Cold War nerve agents set to be banned. Science 2019, 366, 404–405. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, N.; Morita, H.; Nakajima, T. Sarin experiences in Japan: Acute toxicity and long-term effects. J. Neurol. Sci. 2006, 249, 76–85. [Google Scholar] [CrossRef]
- Butler, D. Attacks in UK and Syria highlight growing need for chemical-forensics expertise. Nature 2018, 556, 285–286. [Google Scholar] [CrossRef]
- John, H.; van der Schans, M.J.; Koller, M.; Spruit, H.E.T.; Worek, F.; Thiermann, H.; Noort, D. Fatal sarin poisoning in Syria 2013: Forensic verification within an international laboratory network. Forensic Toxicol. 2018, 36, 61–71. [Google Scholar] [CrossRef]
- Stone, R. U.K. attack puts nerve agent in the spotlight. Science 2018, 359, 1314–1315. [Google Scholar] [CrossRef] [PubMed]
- Halbrook, R.S.; Shugart, L.R.; Watson, A.P.; Munro, N.B.; Linnabary, R.D. Characterizing biological variability in livestock blood cholinesterase activity for biomonitoring organophosphate nerve agent exposure. J. Am. Vet. Med. Assoc. 1992, 201, 714–725. [Google Scholar] [PubMed]
- Munro, N.B.; Shugart, L.R.; Watson, A.P.; Halbrook, R.S. Cholinesterase activity in domestic animals as a potential biomonitor for nerve agent and other organophosphate exposure. J. Am. Vet. Med. Assoc. 1991, 199, 103–115. [Google Scholar] [PubMed]
- Valdez, C.A.; Leif, R.N. Analysis of organophosphorus-based nerve agent degradation products by gas chromatography-mass spectrometry (GC-MS): Current derivatization reactions in the analytical chemist’s toolbox. Molecules 2021, 26, 4631. [Google Scholar] [CrossRef] [PubMed]
- Blanca, M.; Shifrovitch, A.; Dachir, S.; Lazar, S.; Elgarisi, M.; Prihed, H.; Baranes, S.; Egoz, I.; Avraham, M.; Dekel Jaoui, H.; et al. Extended retrospective detection of regenerated sarin (GB) in rabbit blood and the IMPA metabolite in urine: A pharmacokinetics study. Arch. Toxicol. 2021, 95, 2403–2412. [Google Scholar] [CrossRef] [PubMed]
- Golime, R.; Chandra, B.; Palit, M.; Dubey, D.K. Adductomics: A promising tool for the verification of chemical warfare agents’ exposures in biological samples. Arch. Toxicol. 2019, 93, 1473–1484. [Google Scholar] [CrossRef]
- Black, R.M.; Read, R.W. Biological markers of exposure to organophosphorus nerve agents. Arch. Toxicol. 2013, 87, 421–437. [Google Scholar] [CrossRef]
- Elhanany, E.; Ordentlich, A.; Dgany, O.; Kaplan, D.; Segall, Y.; Barak, R.; Velan, B.; Shafferman, A. Resolving pathways of interaction of covalent inhibitors with the active site of acetylcholinesterases: MALDI-TOF/MS analysis of various nerve agent phosphyl adducts. Chem. Res. Toxicol. 2001, 14, 912–918. [Google Scholar] [CrossRef]
- Erhard, M.H.; Schmidt, P.; Kühlmann, R.; Lösch, U. Development of an ELISA for detection of an organophosphorus compound using monoclonal antibodies. Arch. Toxicol. 1989, 63, 462–468. [Google Scholar] [CrossRef]
- Loke, W.K.; Karlsson, B.; Waara, L.; Nyberg, A.G.; Cassel, G.E. Enzyme-based microassay for accurate determination of soman in blood samples. Anal. Biochem. 1998, 257, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Marder, D.; Dagan, S.; Yishai-Aviram, L.; Loewenthal, D.; Chapman, S.; Adani, R.; Lazar, S.; Weissberg, A.; Gura, S. Instantaneous monitoring of free sarin in whole blood by dry blood spot-thermal desorption-GC-FPD/MS analysis. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2020, 1136, 121911. [Google Scholar] [CrossRef] [PubMed]
- Blanca, M.; Shifrovitch, A.; Dachir, S.; Lazar, S.; Elgarisi, M.; Marder, D.; Shamai Yamin, T.; Baranes, S.; Avraham, M.; Dekel Jaoui, H.; et al. Highly sensitive retrospective determination of organophosphorous nerve agent biomarkers in human urine implemented in vivo in rabbit. Arch. Toxicol. 2020, 94, 3033–3044. [Google Scholar] [CrossRef] [PubMed]
- Young, S.A.; Capacio, B.R. The application of a single-column GC-MS-MS method for the rapid analysis of chemical warfare agents and breakdown products. J. Anal. Toxicol. 2019, 43, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Schulze, N.D.; Hamelin, E.I.; Winkeljohn, W.R.; Shaner, R.L.; Basden, B.J.; deCastro, B.R.; Pantazides, B.G.; Thomas, J.D.; Johnson, R.C. Evaluation of multiple blood matrices for assessment of human exposure to nerve agents. J. Anal. Toxicol. 2016, 40, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Rodin, I.; Braun, A.; Stavrianidi, A.; Baygildiev, T.; Shpigun, O.; Oreshkin, D.; Rybalchenko, I. ‘Dilute-and-shoot’ RSLC-MS-MS method for fast detection of nerve and vesicant chemical warfare agent metabolites in urine. J. Anal. Toxicol. 2015, 39, 69–74. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Otsuka, M.; Tsuge, K.; Seto, Y.; Miyaguchi, H.; Uchiyama, M. Analysis of degradation products of nerve agents via post-pentafluorobenzylation liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2018, 1577, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Yishai Aviram, L.; Magen, M.; Chapman, S.; Neufeld Cohen, A.; Lazar, S.; Dagan, S. Dry blood spot sample collection for post-exposure monitoring of chemical warfare agents—In vivo determination of phosphonic acids using LC-MS/MS. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1093–1094, 60–65. [Google Scholar] [CrossRef]
- Shaner, R.L.; Coleman, R.M.; Schulze, N.; Platanitis, K.; Brown, A.A.; Seymour, C.; Kaplan, P.; Perez, J.; Hamelin, E.I.; Johnson, R.C. Investigation of dried blood sampling with liquid chromatography tandem mass spectrometry to confirm human exposure to nerve agents. Anal. Chim. Acta 2018, 1033, 100–107. [Google Scholar] [CrossRef]
- Baygildiev, T.; Zatirakha, A.; Rodin, I.; Braun, A.; Stavrianidi, A.; Koryagina, N.; Rybalchenko, I.; Shpigun, O. Rapid IC-MS/MS determination of methylphosphonic acid in urine of rats exposed to organophosphorus nerve agents. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1058, 32–39. [Google Scholar] [CrossRef]
- Katagi, M.; Nishikawa, M.; Tatsuno, M.; Tsuchihashi, H. Determination of the main hydrolysis product of O-ethyl S-2-diisopropylaminoethyl methylphosphonothiolate, ethyl methylphosphonic acid, in human serum. J. Chromatogr. B Biomed. Sci. Appl. 1997, 689, 327–333. [Google Scholar] [CrossRef]
- Son, V.V.; Nakamura, H.; Imasaka, T.; Imasaka, T. Determination of nerve agent metabolites in human urine by femtosecond laser ionization mass spectrometry using 2-(bromomethyl)naphthalene as a derivatizing reagent. Anal. Chim. Acta 2019, 1069, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Rotenberg, J.S.; Newmark, J. Nerve agent attacks on children: Diagnosis and management. Pediatrics 2003, 112, 648–658. [Google Scholar] [CrossRef] [PubMed]
- McGarry, K.G.; Lalisse, R.F.; Moyer, R.A.; Johnson, K.M.; Tallan, A.M.; Winters, T.P.; Taris, J.E.; McElroy, C.A.; Lemmon, E.E.; Shafaat, H.S.; et al. A novel, modified human butyrylcholinesterase catalytically degrades the chemical warfare nerve agent, sarin. Toxicol. Sci. 2020, 174, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Fidder, A.; Hulst, A.G.; Noort, D.; de Ruiter, R.; van der Schans, M.J.; Benschop, H.P.; Langenberg, J.P. Retrospective detection of exposure to organophosphorus anti-cholinesterases: Mass spectrometric analysis of phosphylated human butyrylcholinesterase. Chem. Res. Toxicol. 2002, 15, 582–590. [Google Scholar] [CrossRef] [PubMed]
- van der Schans, M.J.; Polhuijs, M.; van Dijk, C.; Degenhardt, C.E.; Pleijsier, K.; Langenberg, J.P.; Benschop, H.P. Retrospective detection of exposure to nerve agents: Analysis of phosphofluoridates originating from fluoride-induced reactivation of phosphylated BuChE. Arch. Toxicol. 2004, 78, 508–524. [Google Scholar] [CrossRef]
- Lockridge, O. and L.M. Schopfer, Review of tyrosine and lysine as new motifs for organophosphate binding to proteins that have no active site serine. Chem. Biol. Interact. 2010, 187, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Adams, T.K.; Capacio, B.R.; Smith, J.R.; Whalley, C.E.; Korte, W.D. The application of the fluoride reactivation process to the detection of sarin and soman nerve agent exposures in biological samples. Drug Chem. Toxicol. 2004, 27, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Polhuijs, M.; Langenberg, J.P.; Benschop, H.P. New method for retrospective detection of exposure to organophosphorus anticholinesterases: Application to alleged sarin victims of Japanese terrorists. Toxicol. Appl. Pharmacol. 1997, 146, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Dabisch, P.A.; Davis, E.A.; Renner, J.A.; Jakubowski, E.M.; Mioduszewski, R.J.; Thomson, S.A. Biomarkers of low-level exposure to soman vapor: Comparison of fluoride regeneration to acetylcholinesterase inhibition. Inhal. Toxicol. 2008, 20, 149–156. [Google Scholar] [CrossRef] [PubMed]
- van der Meer, J.A.; Trap, H.C.; Noort, D.; van der Schans, M.J. Comprehensive gas chromatography with time of flight MS and large volume introduction for the detection of fluoride-induced regenerated nerve agent in biological samples. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2010, 878, 1320–1325. [Google Scholar] [CrossRef] [PubMed]
- McGuire, J.M.; Taylor, J.T.; Byers, C.E.; Jakubowski, E.M.; Thomson, S.M. Determination of VX-G analogue in red blood cells via gas chromatography-tandem mass spectrometry following an accidental exposure to VX. J. Anal. Toxicol. 2008, 32, 73–77. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, X.S.; Wu, J.N.; Yan, L.; Xing, Z.F.; Liu, C.C.; Chen, B.; Yuan, L.; Yang, Y. Fluoride reactivation-enabled sensitive quantification of tabun adducts on human serum albumin by GC-MS/MS via isotope dilution. Bioanalysis 2019, 11, 2145–2159. [Google Scholar] [CrossRef]
- Seto, Y.; Kanamori-Kataoka, M.; Komano, A.; Nagoya, T.; Sasano, R.; Matsuo, S. Gas chromatography-mass spectrometry with spiral large-volume injection for determination of fluoridated phosphonates produced by fluoride-mediated regeneration of nerve agent adduct in human serum. J. Chromatogr. A 2019, 1583, 108–116. [Google Scholar] [CrossRef]
- Meyer, M.J.; Bechtold, W.E. Protein adduct biomarkers:State of the art. Environ. Health Perspect. 1996, 104, 879–882. [Google Scholar] [PubMed]
- Noort, D.; Benschop, H.P.; Black, R.M. Biomonitoring of exposure to chemical warfare agents: A review. Toxicol. Appl. Pharmacol. 2002, 184, 116–126. [Google Scholar] [CrossRef]
- Lopachin, R.M.; Decaprio, A.P. Protein adduct formation as a molecular mechanism in neurotoxicity. Toxicol. Sci. 2005, 86, 214–225. [Google Scholar] [CrossRef]
- Peeples, E.S.; Schopfer, L.M.; Duysen, E.G.; Spaulding, R.; Voelker, T.; Thompson, C.M.; Lockridge, O. Albumin, a new biomarker of organophosphorus toxicant exposure, identified by mass spectrometry. Toxicol. Sci. 2005, 83, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Baygildiev, T.; Vokuev, M.F.; Braun, A.V.; Yashkir, V.A.; Rybalchenko, I.V.; Rodin, I.A. Identification of 2-(diethylamino)ethylthiol dipeptide (Cys-Pro) adduct as biomarker of nerve agents VR and CVX in human plasma using liquid chromatography-high-resolution tandem mass spectrometry. Anal. Bioanal. Chem. 2021, 413, 1905–1916. [Google Scholar] [CrossRef]
- Fu, F.; Liu, H.; Gao, R.; Zhao, P.; Lu, X.; Zhang, R.; Wang, L.; Wang, H.; Pei, C. Protein adduct binding properties of tabun-subtype nerve agents after exposure in vitro and in vivo. Toxicol. Lett. 2020, 321, 1–11. [Google Scholar] [CrossRef]
- Dafferner, A.J.; Schopfer, L.M.; Xiao, G.; Cashman, J.R.; Yerramalla, U.; Johnson, R.C.; Blake, T.A.; Lockridge, O. Immunopurification of acetylcholinesterase from red blood cells for detection of nerve agent exposure. Chem. Res. Toxicol. 2017, 30, 1897–1910. [Google Scholar] [CrossRef]
- Onder, S.; Schopfer, L.M.; Cashman, J.R.; Tacal, O.; Johnson, R.C.; Blake, T.A.; Lockridge, O. Use of Hupresin to capture red blood cell acetylcholinesterase for detection of soman exposure. Anal. Chem. 2018, 90, 974–979. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, C.E.; Pleijsier, K.; van der Schans, M.J.; Langenberg, J.P.; Preston, K.E.; Solano, M.I.; Maggio, V.L.; Barr, J.R. Improvements of the fluoride reactivation method for the verification of nerve agent exposure. J. Anal. Toxicol. 2004, 28, 364–371. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Holland, K.E.; Solano, M.I.; Johnson, R.C.; Maggio, V.L.; Barr, J.R. Modifications to the organophosphorus nerve agent-protein adduct refluoridation method for retrospective analysis of nerve agent exposures. J. Anal. Toxicol. 2008, 32, 116–124. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cerasoli, D.M.; Armstrong, S.J.; Reeves, T.E.; Hodgins, S.M.; Kasten, S.A.; Lee-Stubbs, R.B.; Cadieux, C.L.; Otto, T.C.; Capacio, B.R.; Lenz, D.E. Butyrylcholinesterase, a stereospecific in vivo bioscavenger against nerve agent intoxication. Biochem. Pharmacol. 2020, 171, 113670. [Google Scholar] [CrossRef] [PubMed]
- Carol-Visser, J.; van der Schans, M.; Fidder, A.; Hulst, A.G.; van Baar, B.L.; Irth, H.; Noort, D. Development of an automated on-line pepsin digestion-liquid chromatography-tandem mass spectrometry configuration for the rapid analysis of protein adducts of chemical warfare agents. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2008, 870, 91–97. [Google Scholar] [CrossRef] [PubMed]
- van der Schans, M.J.; Fidder, A.; van Oeveren, D.; Hulst, A.G.; Noort, D. Verification of exposure to cholinesterase inhibitors: Generic detection of OPCW Schedule 1 nerve agent adducts to human butyrylcholinesterase. J. Anal. Toxicol. 2008, 32, 125–130. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sporty, J.L.; Lemire, S.W.; Jakubowski, E.M.; Renner, J.A.; Evans, R.A.; Williams, R.F.; Schmidt, J.G.; van der Schans, M.J.; Noort, D.; Johnson, R.C. Immunomagnetic separation and quantification of butyrylcholinesterase nerve agent adducts in human serum. Anal. Chem. 2010, 82, 6593–6600. [Google Scholar] [CrossRef]
- Perez, J.W.; Pantazides, B.G.; Watson, C.M.; Thomas, J.D.; Blake, T.A.; Johnson, R.C. Enhanced stability of blood matrices using a dried sample spot assay to measure human butyrylcholinesterase activity and nerve agent adducts. Anal. Chem. 2015, 87, 5723–5729. [Google Scholar] [CrossRef]
- Tsuge, K.; Seto, Y. Detection of human butyrylcholinesterase-nerve gas adducts by liquid chromatography-mass spectrometric analysis after in gel chymotryptic digestion. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2006, 838, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.A.; Johnson, D.; Carter, M.D.; Stout, E.G.; Erol, H.A.; Isenberg, S.L.; Mathews, T.P.; Thomas, J.D.; Johnson, R.C. A high-throughput UHPLC-MS/MS method for the quantification of five aged butyrylcholinesterase biomarkers from human exposure to organophosphorus nerve agents. Biomed. Chromatogr. 2017, 31, e3830. [Google Scholar] [CrossRef] [PubMed]
- John, H.; Richter, A.; Siegert, M.; Eyer, F.; Thiermann, H. Evidence of exposure to organophosphorus toxicants by detection of the propionylated butyrylcholinesterase-derived nonapeptide-adduct as a novel biomarker. Forensic Sci. Int. 2021, 323, 110818. [Google Scholar] [CrossRef] [PubMed]
- Wieseler, S.; Schopfer, L.; Lockridge, O. Markers of organophosphate exposure in human serum. J. Mol. Neurosci. 2006, 30, 93–94. [Google Scholar] [CrossRef]
- Li, B.; Schopfer, L.M.; Hinrichs, S.H.; Masson, P.; Lockridge, O. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry assay for organophosphorus toxicants bound to human albumin at Tyr411. Anal. Biochem. 2007, 361, 263–272. [Google Scholar] [CrossRef] [PubMed]
- John, H.; Breyer, F.; Thumfart, J.O.; Höchstetter, H.; Thiermann, H. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) for detection and identification of albumin phosphylation by organophosphorus pesticides and G- and V-type nerve agents. Anal. Bioanal. Chem. 2010, 398, 2677–2691. [Google Scholar] [CrossRef]
- Black, R.M. History and perspectives of bioanalytical methods for chemical warfare agent detection. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2010, 878, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Dubrovskii, Y.A.; Podolskaya, E.P.; Murashko, E.A.; Babakov, V.; Nachon, F.; Masson, P.; Schopfer, L.M.; Lockridge, O. PHOS-select iron affinity beads enrich peptides for the detection of organophosphorus adducts on albumin. Chem. Res. Toxicol. 2013, 26, 1917–1925. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Duysen, E.G.; Froment, M.T.; Masson, P.; Nachon, F.; Jiang, W.; Schopfer, L.M.; Thiele, G.M.; Klassen, L.W.; Cashman, J.; et al. Polyclonal antibody to soman-tyrosine. Chem. Res. Toxicol. 2013, 26, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kim, C.; Lee, Y.H. Simultaneous time-concentration analysis of soman and VX adducts to butyrylcholinesterase and albumin by LC-MS-MS. J. Anal. Toxicol. 2018, 42, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Fu, F.; Sun, F.; Lu, X.; Song, T.; Ding, J.; Gao, R.; Wang, H.; Pei, C. A novel potential biomarker on Y263 site in human serum albumin poisoned by six nerve agents. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1104, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Ding, J.; Lu, X.; Gao, R.; Lu, X.; Shi, E.; Wang, H.; Pei, C. Mass spectral characterization of tabun-labeled lysine biomarkers in albumin. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1057, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Fu, F.; Liu, H.; Lu, X.; Zhang, R.; Li, L.; Gao, R.; Xie, J.; Wang, H.; Pei, C. Identification of S419 on human serum albumin as a novel biomarker for sarin and cyclosarin exposure. Rapid Commun. Mass Spectrom. 2020, 34, e8721. [Google Scholar] [CrossRef] [PubMed]
- Fu, F.; Guo, Y.; Lu, X.; Zhao, P.; Zou, S.; Wang, H.; Gao, R.; Pei, C. Forensic analysis of soman exposure using characteristic fragment ions from protein adducts. Hum. Exp. Toxicol. 2021, 40, 1519–1527. [Google Scholar] [CrossRef] [PubMed]
- Kranawetvogl, A.; Küppers, J.; Gütschow, M.; Worek, F.; Thiermann, H.; Elsinghorst, P.W.; John, H. Identification of novel disulfide adducts between the thiol containing leaving group of the nerve agent VX and cysteine containing tripeptides derived from human serum albumin. Drug Test. Anal. 2017, 9, 1192–1203. [Google Scholar] [CrossRef]
- Kranawetvogl, A.; Küppers, J.; Siegert, M.; Gütschow, M.; Worek, F.; Thiermann, H.; Elsinghorst, P.W.; John, H. Bioanalytical verification of V-type nerve agent exposure: Simultaneous detection of phosphonylated tyrosines and cysteine-containing disulfide-adducts derived from human albumin. Anal. Bioanal. Chem. 2018, 410, 1463–1474. [Google Scholar] [CrossRef] [PubMed]
- Fu, F.; Chen, J.; Zhao, P.; Lu, X.; Gao, R.; Chen, D.; Liu, H.; Wang, H.; Pei, C. Tracing and attribution of V-type nerve agents in human exposure by strategy of assessing the phosphonylated and disulfide adducts on ceruloplasmin. Toxicology 2020, 430, 152346. [Google Scholar] [CrossRef]
- Fu, F.; Gao, R.; Zhang, R.; Zhao, P.; Lu, X.; Li, L.; Wang, H.; Pei, C. Verification of soman-related nerve agents via detection of phosphonylated adducts from rabbit albumin in vitro and in vivo. Arch. Toxicol. 2019, 93, 1853–1863. [Google Scholar] [CrossRef]
- Kranawetvogl, A.; Worek, F.; Thiermann, H.; John, H. Modification of human serum albumin by the nerve agent VX: Microbore liquid chromatography/electrospray ionization high-resolution time-of-flight tandem mass spectrometry method for detection of phosphonylated tyrosine and novel cysteine containing disulfide adducts. Rapid Commun. Mass Spectrom. 2016, 30, 2191–2200. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Potential Biomarkers with High Reactivity | Other Signals | Sample | Protease | Analytical Method | Agents | References |
|---|---|---|---|---|---|---|
| VRY411TKKVPQVSTPTL LVRY411TKKVPQVSTPTL | Product ions | Human albumin and plasma | Pepsin | MALDI-TOF MS | GB | [65] |
| YKFQNALLVRY411TKKVPQVSTPTLVE IARRHPY148FY150APE LLFFAKRY161KAAFTE | Any sites | Pure HSA and plasma | Trypsin, pepsin, pronase | MALDI-TOF MS | GA, GB, GD, GF, VX, Vs, VR | [66] |
| VRY411TKKVPQVST LVRY411TKKVPQVST | Human plasma | Pepsin | MALDI-TOF/TOF MS | GD | [68] | |
| RY*GRK | Any sites | Albumin of human and mouse | Pepsin | MALDI-TOF-TOF MS | GD | [69] |
| VX and GD-tyrosines | Plasma of rhesus monkeys | Pepsin | LC–MS-MS SRM mode | GD, VX | [70] | |
| Y263ICENQDSISSK | Any sites | HSA | Trypsin | Q Exactive LC-MS/MS | GA, GB, GF, VX, ethyl GA, and propyl GA, | [71] |
| FPK224AEFVEVTK K524QTALVELLK AYK212AWALVR LDALK186EKALISAAQER K414VPQVSTPTLVEISR LC*AIPK79LR LDAVK186EK | Any sites and characteristic ions | Albumin of bovine, leporine, and rat | Trypsin | Orbitrap LC-MS/HRMS | GA | [72] |
| K414VPQVS419TPTLVESR | Any sites | HSA | Trypsin | ESI-Q-Orbitrap MS | GA, GB, GD, GF, VX, Vs, VR, MEGA, EEGA | [73] |
| DEAET-C34P | Product ions | Human plasma | Pronase | LC-MS/HR MS | VR, VX | [50] |
| K525QTALVELVK LK199CASLQK | Any lysine sites, Product ions for GAs-lysine | HSA, RSA in vitro and in vivo | Trypsin | ESI-Q-Orbitrap MS | GA | [51] |
| EK188ALISAAQER YK162AILTECCEAADK | Any sites | Rabbit albumin in vitro and in vivo | Trypsin | nLC-Q-Orbitrap-MS | GD | [78] |
| DPAET-Cys34Pro MetProCys448-DPAET AspIleCys514-DEAET | Product ions | Human plasma | Pronase | μLC-ESI MS/HRMS | VX, VR, Vs | [75] |
| PC34-DPAET MPC448-DPAET DIC514-DPAET | Product ions | HSA | Pronase | µLC-ESI MS/HR MS | VX | [76] |
| DPAET-C34P PC34-DPAET | Any sites | HSA | Pronase | µLC-ESI HR MS/MS | VX | [79] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Lu, X.; Gao, R.; Pei, C.; Wang, H. Current Progress for Retrospective Identification of Nerve Agent Biomarkers in Biological Samples after Exposure. Toxics 2022, 10, 439. https://doi.org/10.3390/toxics10080439
Wang J, Lu X, Gao R, Pei C, Wang H. Current Progress for Retrospective Identification of Nerve Agent Biomarkers in Biological Samples after Exposure. Toxics. 2022; 10(8):439. https://doi.org/10.3390/toxics10080439
Chicago/Turabian StyleWang, Jin, Xiaogang Lu, Runli Gao, Chengxin Pei, and Hongmei Wang. 2022. "Current Progress for Retrospective Identification of Nerve Agent Biomarkers in Biological Samples after Exposure" Toxics 10, no. 8: 439. https://doi.org/10.3390/toxics10080439
APA StyleWang, J., Lu, X., Gao, R., Pei, C., & Wang, H. (2022). Current Progress for Retrospective Identification of Nerve Agent Biomarkers in Biological Samples after Exposure. Toxics, 10(8), 439. https://doi.org/10.3390/toxics10080439