
Combine Effect of ZnO NPs and Bacteria on Protein and Gene’s Expression Profile of Rice (Oryza sativa L.) Plant

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Water Sampling
2.2. Characteristics of ZnO NPs
2.3. Rice Plant
2.4. Bio-Priming of Seeds
2.5. Hydroponic Culture Experiment
2.6. Growth Analysis and Partitioning of Biomass
2.7. Total Nitrogen-Protein
2.8. Total Structural Protein
2.8.1. Protein Extraction from a Plant Sample
2.8.2. Bradford Assay
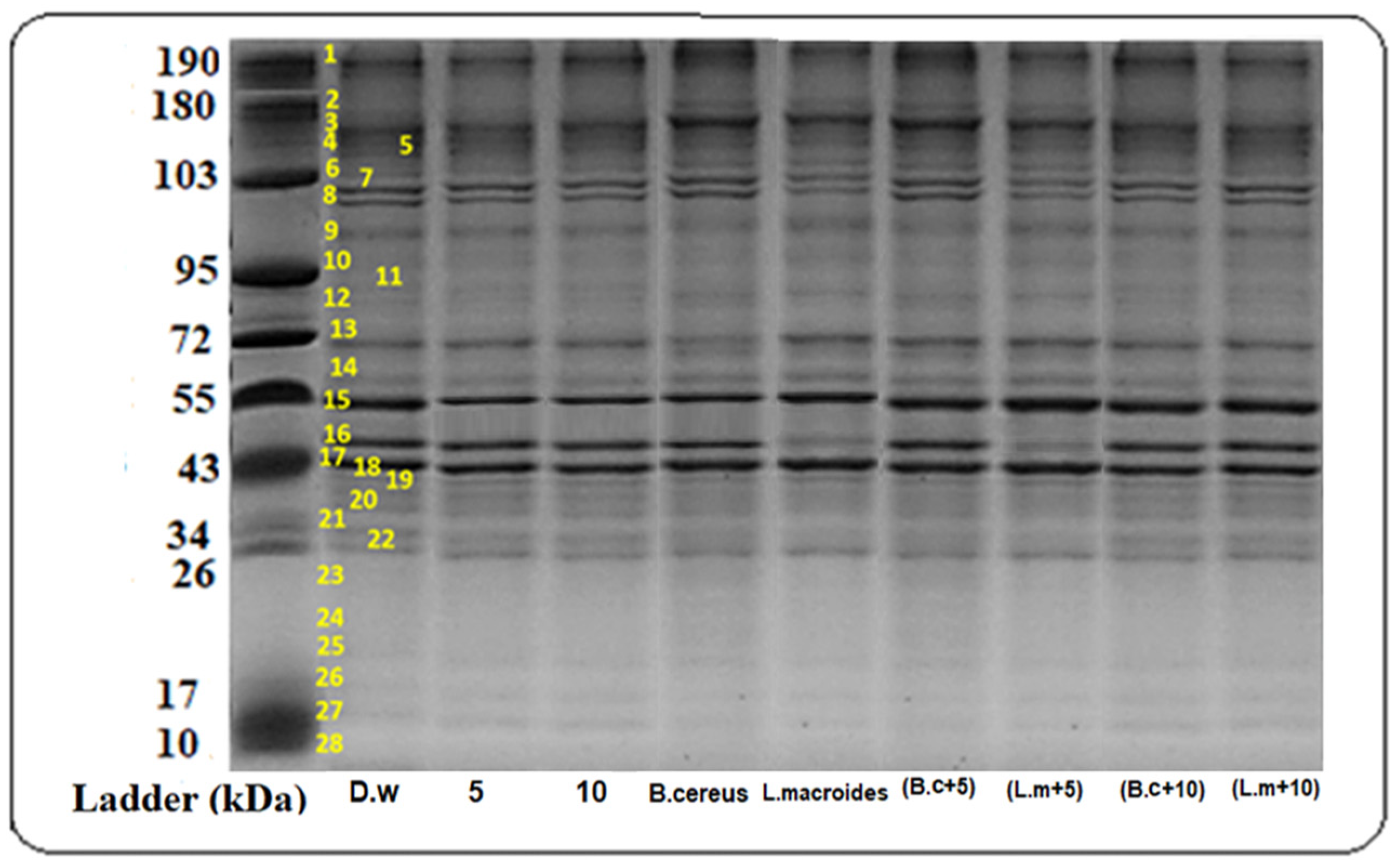
2.8.3. SDS-PAGE, (Sodium, Dodecyl, Sulfate, Polyacrylamide Gel Electrophoresis) Analysis
2.9. Molecular Studies
2.9.1. RNA Extraction
2.9.2. cDNA Synthesis
2.9.3. Polymerase Chain Reaction (PCR)
2.10. Statistical Analysis
3. Results
3.1. Physicochemical Properties of Wastewater
3.2. Total, Nitrogen and Protein Content
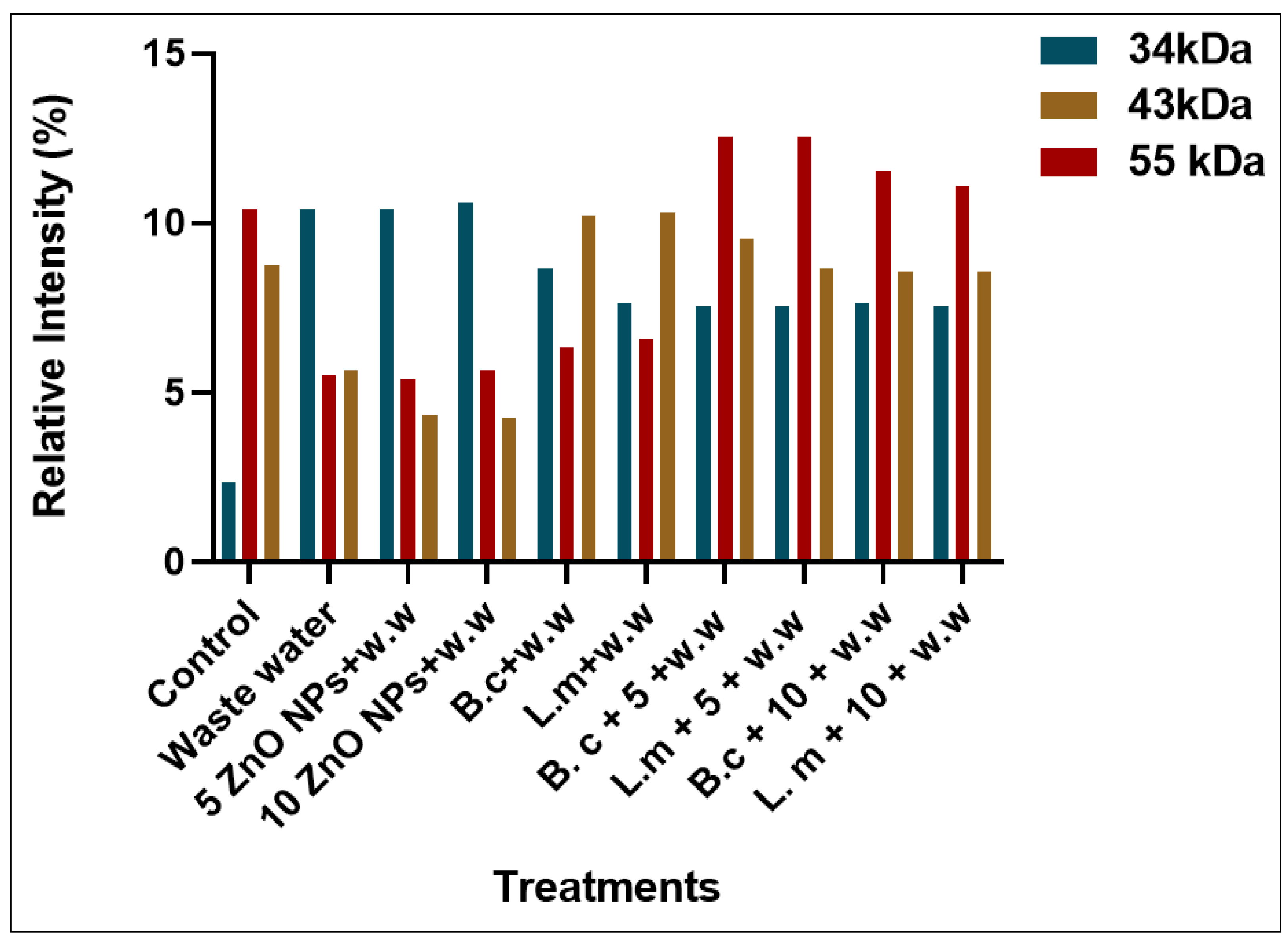
3.3. Total Structural Protein (SDS-PAGE)
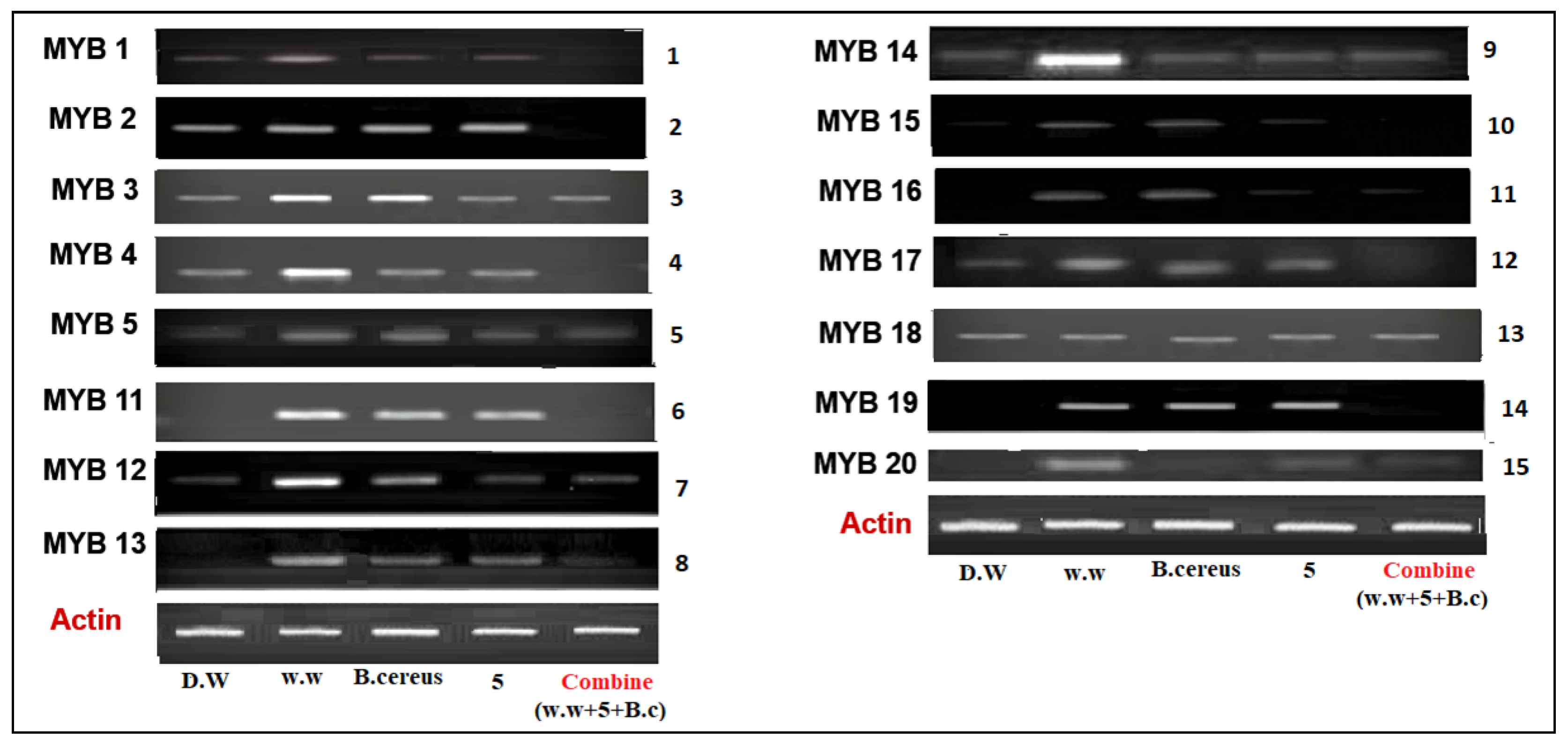
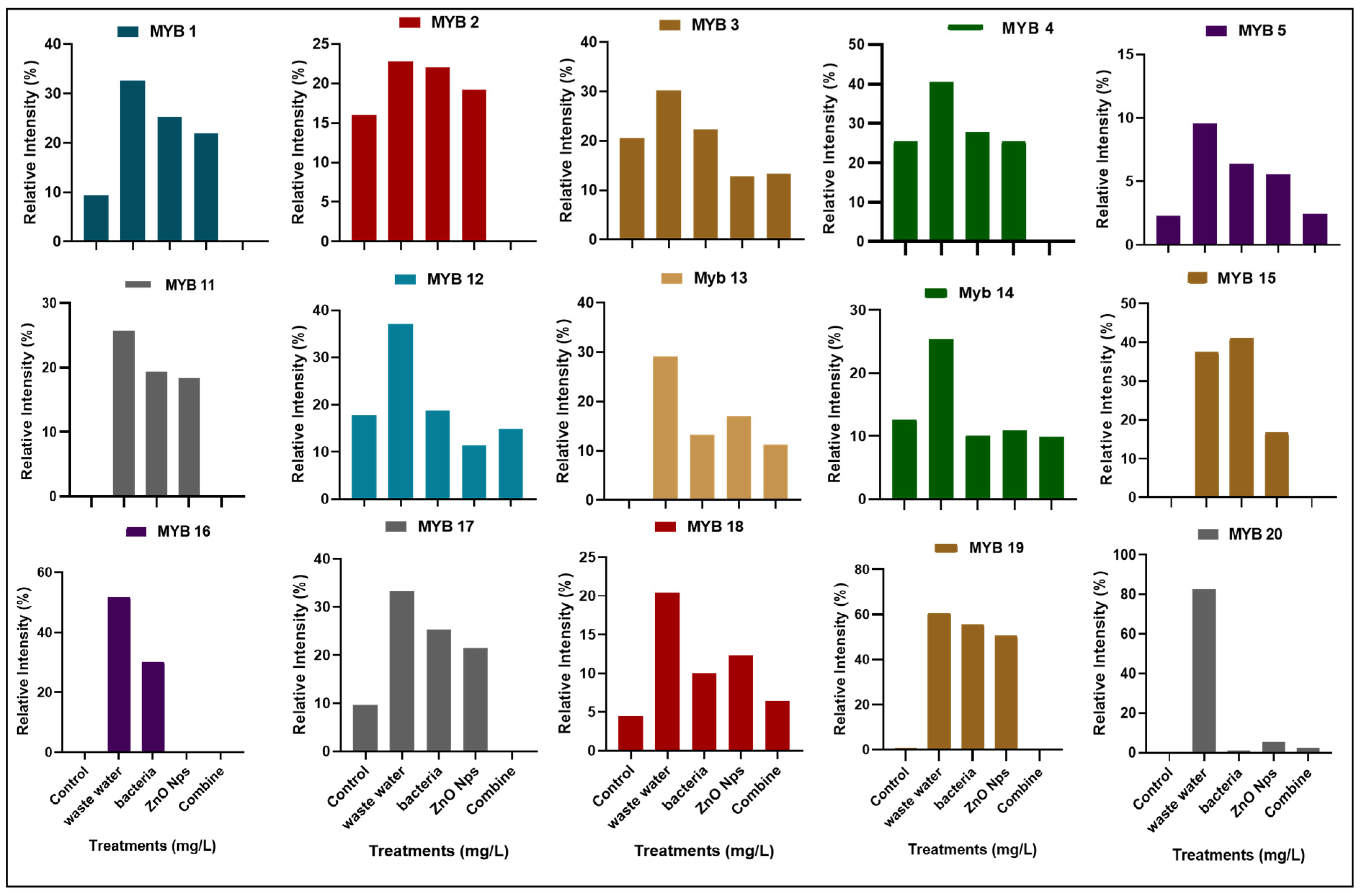
3.4. Molecular Analysis
3.4.1. Expression of Myeloblastosis (Myb) Gene
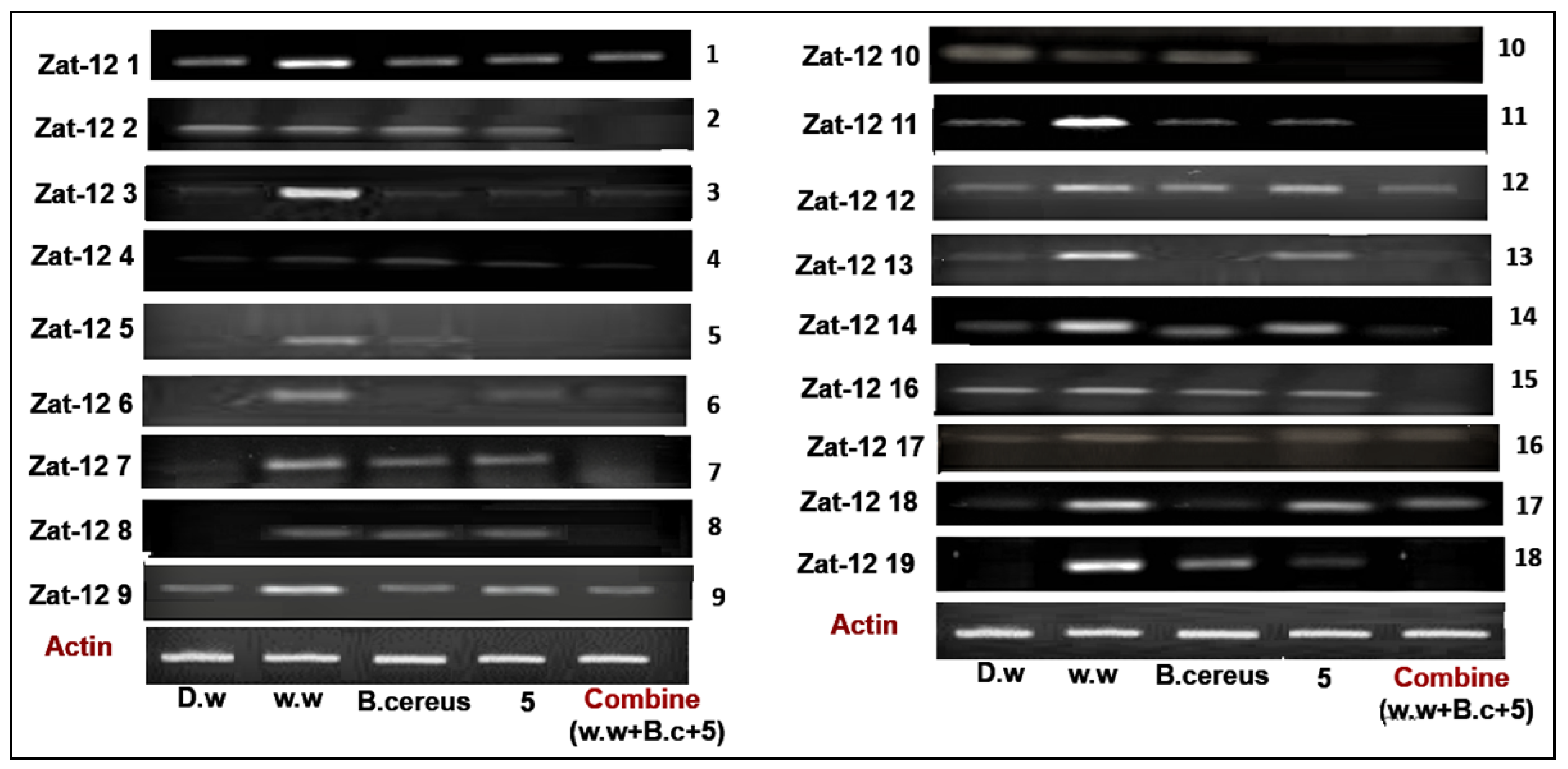
3.4.2. Expression of Zinc Finger Protein (Zat-12) Gene Family
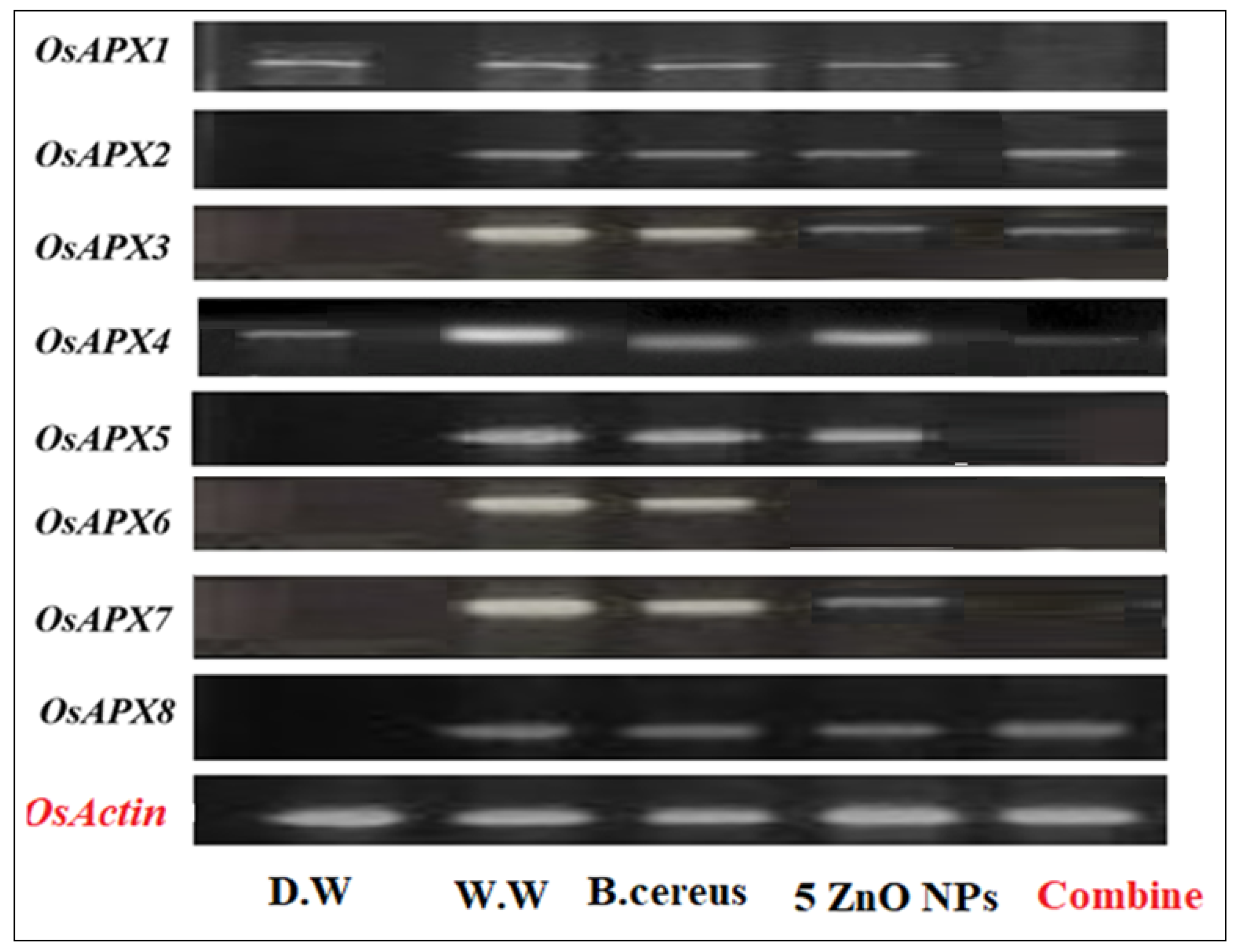
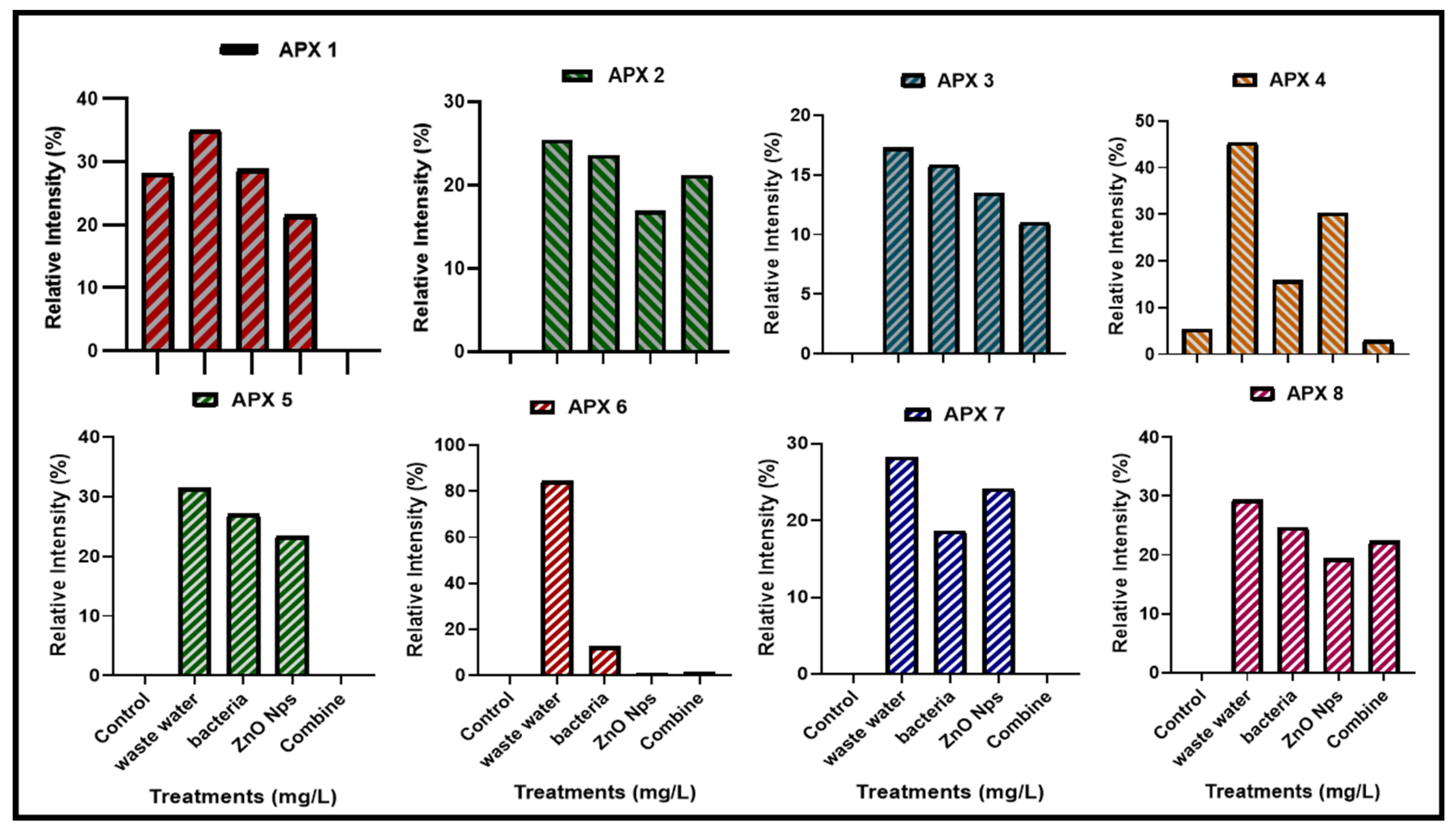
3.4.3. Expression of Ascorbate Peroxidase (Apx) Gene Family
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wiszniewska, A. Priming strategies for benefiting plant performance under toxic trace metal exposure. Plants 2021, 10, 623. [Google Scholar] [CrossRef]
- Zakaria, Z.; Zulkafflee, N.S.; Mohd Redzuan, N.A.; Selamat, J.; Ismail, M.R.; Praveena, S.M.; Abdull Razis, A.F. Understanding potential heavy metal contamination, absorption, translocation and accumulation in rice and human health risks. Plants 2021, 10, 1070. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Khan, S.; Rehman, S.U.; Rehman, Z.U.; Rehman, Z.U.M.; Rha, E.S.; Jamil, M. Zinc oxide nanoparticles enhance the tolerance and remediation potential of Bacillus spp. against heavy metals stress. Adsorpt. Sci. Technol. 2021, 9, 110–115. [Google Scholar] [CrossRef]
- Kaur, R.; Das, S.; Bansal, S.; Singh, G.; Sardar, S.; Dhar, H.; Ram, H. Heavy metal stress in rice: Uptake, transport, signaling, and tolerance mechanisms. Physiol. Plant. 2021, 173, 430–448. [Google Scholar] [CrossRef]
- Angulo-Bejarano, P.I.; Puente-Rivera, J.; Cruz-Ortega, R. Metal and metalloid toxicity in plants: An overview on molecular aspects. Plants 2021, 10, 635. [Google Scholar] [CrossRef] [PubMed]
- Altaf, R.; Altaf, S.; Hussain, M.; Shah, R.U.; Ullah, R.; Ullah, M.I.; Datta, R. Heavy metal accumulation by roadside vegetation and implications for pollution control. PLoS ONE 2021, 16, e0249147. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Rolly, N.K.; Al Azzawi, T.N.I.; Imran, M.; Mun, B.G.; Lee, I.J.; Yun, B.W. Lead (Pb)-induced oxidative stress alters the morphological and physio-biochemical properties of rice (Oryza sativa L.). Agronomy 2021, 11, 409. [Google Scholar] [CrossRef]
- Lee, S. Recent Advances on nitrogen use efficiency in rice. Agronomy 2021, 11, 753. [Google Scholar] [CrossRef]
- Tariq, F.; Wang, X.; Saleem, M.H.; Khan, Z.I.; Ahmad, K.; Saleem Malik, I.; Ali, S. Risk assessment of heavy metals in basmati rice: Implications for public health. Sustainability 2021, 13, 8513. [Google Scholar] [CrossRef]
- Kerchev, P.; van der Meer, T.; Sujeeth, N.; Verlee, A.; Stevens, C.V.; Van Breusegem, F.; Gechev, T. Molecular priming as an approach to induce tolerance against abiotic and oxidative stresses in crop plants. Biotechnol. Adv. 2020, 40, 107–117. [Google Scholar] [CrossRef]
- Muzammil, M.; Zahid, A.; Breuer, L. Water resources management strategies for irrigated agriculture in the Indus basin of Pakistan. Water 2020, 12, 1429. [Google Scholar] [CrossRef]
- Zhou, P.; Adeel, M.; Shakoor, N.; Guo, M.; Hao, Y.; Azeem, I.; Rui, Y. Application of nanoparticles alleviates heavy metals stress and promotes plant growth: An overview. Nanomaterials 2021, 11, 26. [Google Scholar] [CrossRef] [PubMed]
- Jaskulak, M.; Rorat, A.; Grobelak, A.; Chaabene, Z.; Kacprzak, M.; Vandenbulcke, F. Bioaccumulation, antioxidative response, and metallothionein expression in Lupinus luteus L. exposed to heavy metals and silver nanoparticles. Environ. Sci. Pollut. 2019, 26, 16040–16052. [Google Scholar] [CrossRef] [PubMed]
- Javaid, S.; Zaman, Q.; Sultan, K.; Riaz, U.; Aslam, A.; Saba Sharif, N.E.; Ibraheem, S. Heavy metals stress, mechanism and remediation techniques in rice (Oryza sativa L.): A review. Pure Appl. Biol. 2020, 9, 403–426. [Google Scholar] [CrossRef]
- Mitra, D.; Mondal, R.; Khoshru, B.; Shadangi, S.; Mohapatra, P.K.D.; Panneerselvam, P. Rhizobacteria mediated seed bio-priming triggers the resistance and plant growth for sustainable crop production. Curr. Res. Microb. Sci. 2021, 2, 100071. [Google Scholar] [CrossRef]
- Wang, S.; Liu, T.; Xiao, X.; Luo, S. Advances in microbial remediation for heavy metal treatment: A mini review. J. Leather Sci. Eng. 2021, 3, 1–10. [Google Scholar] [CrossRef]
- Radulescu, C.; Stihi, C.; Ionita, I.; Dulama, I.D.; Chilian, A.; Necula, C.; Chelarescu, E.D. Determination of heavy metal levels in water and therapeutic mud by atomic absorption spectrometry. Rom. J. Phys. 2014, 59, 1057–1059. [Google Scholar]
- Hossain, S.; Hossain, S.; Islam, M.R.; Kabir, M.H.; Ali, S.; Islam, M.S.; Mahmud, Z.H. Bioremediation of hexavalent chromium by chromium resistant bacteria reduces phytotoxicity. Int. J. Environ. Res. Public Health 2020, 17, 6013. [Google Scholar] [CrossRef]
- Amara, U.; Shad, S.; Ilyas, N.; Manaf, A.; Raja, N.I. In vitro germination and biochemical profiling of Brassica napus in response to biosynthesised zinc nanoparticles. IET Nanobiotechnology 2018, 13, 46–51. [Google Scholar]
- Khattak, M.R.; Shah, G.; Naeem, R.; Fayyaz, M.; Rehman, M.; Anees, M.; Rha, E.S.; Jamil, M. Assessment of heavy metal tolerance and reduction potential of bacillus sp. isolated from the soils contaminated with industrial effluents. Fresenius Environ. Bull. 2021, 30, 4391–4401. [Google Scholar]
- Ashraf, I.; Ahmad, F.; Sharif, A.; Altaf, A.R.; Teng, H. Heavy metals assessment in water, soil, vegetables and their associated health risks via consumption of vegetables, District Kasur, Pakistan. SN Appl. Sci. 2021, 3, 552. [Google Scholar] [CrossRef]
- Gu, M.; Hao, L.; Wang, Y.; Li, X.; Chen, Y.; Li, W.; Jiang, L. The selective heavy metal ions adsorption of zinc oxide nanoparticles from dental wastewater. Chem. Phys. 2020, 534, 110750. [Google Scholar] [CrossRef]
- Delrue, F.; Ribeiro de Jesus Cerqueira, M.; Compadre, A.; Alvarez, P.; Fleury, G.; Escoffier, C.; Sassi, J.F. Hydroponic farm wastewater treatment using an indigenous consortium. Processes 2021, 9, 519. [Google Scholar] [CrossRef]
- Ahmad, P.; Alyemeni, M.N.; Al-Huqail, A.A.; Alqahtani, M.A.; Wijaya, L.; Ashraf, M.; Bajguz, A. Zinc oxide nanoparticles application alleviates arsenic (As) toxicity in soybean plants by restricting the uptake of as and modulating key biochemical attributes, antioxidant enzymes, ascorbate-glutathione cycle and glyoxalase system. Plants 2020, 9, 825. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, P.; Jayaraj, M.; Manikandan, R.; Geetha, N.; Rene, E.R.; Sharma, N.C.; Sahi, S.V. Zinc oxide nanoparticles (ZnONPs) alleviate heavy metal-induced toxicity in Leucaena leucocephala seedlings: A physiochemical analysis. Plant Physiol. Biochem. 2017, 110, 59–69. [Google Scholar] [CrossRef]
- Maluin, F.N.; Hussein, M.Z.; Nik Ibrahim, N.N.L.; Wayayok, A.; Hashim, N. Some emerging opportunities of nanotechnology development for soilless and microgreen farming. Agronomy 2021, 11, 1213. [Google Scholar] [CrossRef]
- Sáez-Plaza, P.; Michałowski, T.; Navas, M.J.; Asuero, A.G.; Wybraniec, S. An overview of the Kjeldahl method of nitrogen determination. Part I. Early history, chemistry of the procedure, and titrimetric finish. Crit. Rev. Anal. Chem. 2013, 43, 178–223. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Deepa, K.; Sheeja, T.E.; Santhi, R.; Sasikumar, B.; Cyriac, A.; Deepesh, P.V.; Prasath, D. A simple and efficient protocol for isolation of high quality functional RNA from different tissues of turmeric (Curcuma longa L.). Physiol. Mol. Biol. Plants 2014, 20, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Bao, J.; Wang, T.; Moryani, H.T.; Kang, W.; Zheng, J.; Xiao, W. Hazardous heavy metals accumulation and health risk assessment of different vegetable species in contaminated soils from a typical mining city, central china. Int. J. Environ. Res. Public Health 2021, 18, 2617. [Google Scholar] [CrossRef]
- Li, J.; Zhang, M.; Sun, J.; Mao, X.; Wang, J.; Liu, H.; Zou, D. Heavy metal stress-associated proteins in rice and arabidopsis: Genome-wide identification, phylogenetics, duplication, and expression profiles analysis. Front. Genet. 2020, 11, 477–485. [Google Scholar] [CrossRef]
- Awan, S.A.; Ilyas, N.; Khan, I.; Raza, M.A.; Rehman, A.U.; Rizwan, M.; Brestic, M. Bacillus siamensis reduces cadmium accumulation and improves growth and antioxidant defense system in two wheat (Triticum aestivum L.) varieties. Plants 2020, 9, 878. [Google Scholar] [CrossRef]
- Priyanka, N.; Geetha, N.; Manish, T.; Sahi, S.V.; Venkatachalam, P. Zinc oxide nanocatalyst mediates cadmium and lead toxicity tolerance mechanism by differential regulation of photosynthetic machinery and antioxidant enzymes level in cotton seedlings. Toxicol. Rep. 2021, 8, 295–302. [Google Scholar]
- Fan, Y.; Zhang, Y.; Rui, C.; Xu, N.; Zhang, H.; Wang, J.; Ye, W. Zinc finger transcription factor zat family genes confer multi-tolerances in gossypium hirsutum L. J. Cotton Res. 2021, 4, 24. [Google Scholar] [CrossRef]
- Rossi, L.; Sharifan, H.; Zhang, W.; Schwab, A.P.; Ma, X. Mutual effects and in planta accumulation of co-existing cerium oxide nanoparticles and cadmium in hydroponically grown soybean (Glycine max (L.) Merr.). Environ. Sci. Nanotechnol. 2018, 5, 150–157. [Google Scholar]
- Mahmud, U.; Salam, M.T.B.; Khan, A.S.; Rahman, M.M. Ecological risk of heavy metal in agricultural soil and transfer to rice grains. Discov. Mater. 2021, 1, 10. [Google Scholar] [CrossRef]
- Carrouel, F.; Viennot, S.; Ottolenghi, L.; Gaillard, C.; Bourgeois, D. Nanoparticles as anti-microbial, anti-inflammatory, and remineralizing agents in oral care cosmetics: A review of the current situation. Nanomaterials 2020, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Vannini, C.; Domingo, G.; Fiorilli, V.; Seco, D.G.; Novero, M.; Marsoni, M.; Bonfante, P. Proteomic analysis reveals how pairing of a Mycorrhizal fungus with plant growth-promoting bacteria modulates growth and defense in wheat. Plant Cell Environ. 2021, 44, 1946–1960. [Google Scholar] [CrossRef]
- Tiwari, V.; Mishra, N.; Gadani, K.; Solanki, P.S.; Shah, N.A.; Tiwari, M. Mechanism of anti-bacterial activity of zinc oxide nanoparticle against Carbapenem-Resistant Acinetobacter baumannii. Front. Microbiol. 2018, 9, 1218–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, S.; Habib-Ur-Rehman, M.; Khanam, T.; Sheer, A.; Kebin, Z.; Jianjun, Y. Health risk assessment of different heavy metals dissolved in drinking water. Int. J. Environ. Res. Public Health 2019, 16, 1737. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, H.; Menon, S.; Venkat Kumar, S.; Rajeshkumar, S. Mechanistic study on antibacterial action of zinc oxide nanoparticles synthesized using green route. Chem. Biol. Interact. 2018, 286, 60–70. [Google Scholar] [CrossRef]
- Fiol, D.F.; Terrile, M.C.; Frik, J.; Mesas, F.A.; Álvarez, V.A.; Casalongué, C.A. Nanotechnology in plants: Recent advances and challenges. J. Chem. Technol. Biotechnol. 2021, 96, 2095–2108. [Google Scholar] [CrossRef]
- Kasahun, M.; Alemayhu Yadate, A.B.; Belay, Z.; Ramalingam, M. Antimicrobial activity of chemical, thermal and green route-derived zinc oxide nanoparticles: A comparative analysis. Nanoparticles Biomed. Eng. 2020, 12, 47–56. [Google Scholar] [CrossRef]
- Sabir, S.; Zahoor, M.A.; Waseem, M.; Siddique, M.H.; Shafique, M.; Imran, M.; Muzammil, S. Biosynthesis of ZnO nanoparticles using bacillus subtilis: Characterization and nutritive significance for promoting plant growth in Zea mays L. Dose-Response 2020, 18, 1559325820958911. [Google Scholar] [CrossRef]
- Elemike, E.E.; Uzoh, I.M.; Onwudiwe, D.C.; Babalola, O.O. The role of nanotechnology in the fortification of plant nutrients and improvement of crop production. Appl. Sci. 2019, 9, 499. [Google Scholar] [CrossRef] [Green Version]
- Gudkov, S.V.; Burmistrov, D.E.; Serov, D.A.; Rebezov, M.B.; Semenova, A.A.; Lisitsyn, A.B. A mini review of antibacterial properties of ZnO nanoparticles. Front. Phys. 2021, 9, 641–681. [Google Scholar] [CrossRef]
- Gonzalez, P.J.; Lozano, M.; Lascano, H.R.; Lagares, A.; Melchiorre, M.N. Increased oxidative stress tolerance by superoxide dismutase overexpression in mesorhizobium loti. BioRxiv 2018, 1, 363556. [Google Scholar]
- Khande, R.; Sharma, S.K.; Ramesh, A.; Sharma, M.P. Zinc solubilizing Bacillus strains that modulate growth, yield and zinc biofortification of soybean and wheat. Rhizosphere 2017, 4, 126–138. [Google Scholar] [CrossRef]
- Hong, S.H.; Tripathi, B.N.; Chung, M.S.; Cho, C.; Lee, S.; Kim, J.H.; Lee, S.S. Functional switching of ascorbate peroxidase 2 of rice (OsAPX2) between peroxidase and molecular chaperone. Sci. Rep. 2018, 8, 9171. [Google Scholar] [CrossRef]
- Gupta, P.; Diwan, B. Bacterial Exopolysaccharide mediated heavy metal removal: A Review on biosynthesis, mechanism and remediation strategies. Biotechnol. Rep. 2017, 13, 58–71. [Google Scholar] [CrossRef]
- Gupta, S.D.; Agarwal, A.; Pradhan, S. Phytostimulatory effect of silver nanoparticles (AgNPs) on rice seedling growth: An insight from antioxidative enzyme activities and gene expression patterns. Ecotoxicol. Environ. Saf. 2018, 161, 624–633. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Running Gel Composition | Stacking Gel Composition | ||
|---|---|---|---|
| Components | Quantity | Components | Quantity |
| 1.5 M Tris-HCl (pH 8.8) | 2.6 mL | 0.5 M Tris-HCl (pH 6.8) | 2.50 mL |
| Distilled water (DW) | 3.2 mL | Distilled water (DW) | 3.975 mL |
| 30% of Acrylamide | 4 mL | 30% of Acrylamide | 600 µL |
| 20% of SDS | 100 µL | 10% of APS | 50 µL |
| 10% of APS | 100 µL | 20% of SDS | 100 µL |
| TEMED | 7–10 µL | TEMED | 5–7 µL |
| Physico-Chemical Parameters | HIE Wastewater | Standard (NEQS) |
|---|---|---|
| Temperature | 24.5 °C | 40.0 °C |
| pH | 7.23 pH | 6–10 pH |
| EC | 682 (µS/m) | 500 (µS/m) |
| Odure | Bad | Bad |
| TSS | 400 mg/L | 150 mg/L |
| TDS | 4485 mg/L | 3500 mg/L |
| BOD | 250 mg/L | 80 mg/L |
| COD | 400 mg/L | 150 mg/L |
| Lead | 2.84 mg/L | 0.5 mg/L |
| Cadmium | 2.26 mg/L | 1.0 mg/L |
| Chromium | 2.40 mg/L | 1.0 mg/L |
| Iron | 1.19 mg/L | 0.2 mg/L |
| Manganese | 1.36 mg/L | 1.5 mg/L |
| Nickle | 1.83 mg/L | 1.0 mg/L |
| Total Nitrogen (µg/g) | Total Protein (µg/g) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatments | Leaf | Shoot | Root | Leaf | Shoot | Root | ||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| D.W | 5.62 | 0.211 a | 4.23 | 0.029 ab | 3.11 | 0.023 a | 45.43 | 0.026 de | 40.43 | 0.01 a | 32.22 | 0.028 ab |
| Wastewater | 2.21 | 0.029 a | 1.62 | 0.064 cde | 1.12 | 0.052 a | 15.32 | 0.031 ad | 9.23 | 0.054 a | 6.21 | 0.01 bc |
| Bacillus cereus | 12.8 | 0.034 ab | 9.31 | 0.021 ab | 6.12 | 0.028 ab | 60.23 | 0.015 cd | 53.23 | 0.023 ab | 47.31 | 0.078 bc |
| (B. cereus + w.w) | 14.32 | 0.039 abc | 10.52 | 0.083 cdf | 8.23 | 0.011 ab | 23.21 | 0.023 ab | 16.43 | 0.063 ab | 10.23 | 0.016 a |
| Lysinibacillus macroides | 10.2 | 0.020 a | 7.22 | 0.037 ac | 5.43 | 0.078 bc | 55.41 | 0.012 ab | 47.41 | 0.021 abc | 35.41 | 0.078 b |
| (L. macroides + w.w) | 12.13 | 0.018 ab | 8.32 | 0.094 abc | 6.43 | 0.024 ab | 21.12 | 0.01 ab | 14.12 | 0.034 b | 9.23 | 0.025 c |
| 5 mg/L ZnO Nps | 15.21 | 0.018 a | 10.31 | 0.022 a | 8.26 | 0.037 def | 62.21 | 0.013 ab | 47.32 | 0.012 ab | 38.23 | 0.030 a |
| (5 mg/L + w.w) | 12.23 | 0.017 bc | 7.21 | 0.032 cdf | 5.46 | 0.015 ac | 22.34 | 0.021 a | 16.54 | 0.051 bc | 10.78 | 0.052 c |
| 10 mg/L ZnO Nps | 20.31 | 0.039 d | 15.21 | 0.025 bc | 11.61 | 0.030 ab | 67.12 | 0.022 ab | 50.21 | 0.043 ab | 42.12 | 0.017 c |
| (10 mg/L + w.w) | 17.21 | 0.039 bcd | 12.42 | 0.074 ab | 9.62 | 0.027 ab | 34.21 | 0.021 ab | 28.23 | 0.012 ab | 23.21 | 0.017 b |
| (B. cereus +5) | 20.23 | 0.037 b | 18.31 | 0.01 ab | 14.81 | 0.091 bc | 70.32 | 0.024 ab | 63.21 | 0.011 abc | 56.21 | 0.052 ab |
| (B. cereus +5 + w.w) | 25.32 | 0.013 bcd | 20.43 | 0.073 abd | 17.62 | 0.032 ab | 32.21 | 0.034 b | 26.54 | 0.032 c | 16.43 | 0.032 a |
| (L. macroides +5) | 19.9 | 0.023 ab | 15.41 | 0.016 ae | 12.13 | 0.056 be | 66.41 | 0.021 ab | 52.31 | 0.032 a | 47.32 | 0.015 ab |
| (L. macroides +5 + w.w) | 22.65 | 0.022 abc | 17.22 | 0.047 abe | 15.51 | 0.016 ab | 30.22 | 0.043 ab | 22.21 | 0.021 bd | 14.21 | 0.016 c |
| (B. cereus +w.w) | 27.52 | 0.024 bc | 22.12 | 0.062 bc | 19.72 | 0.033 ab | 77.22 | 0.03 ab | 45.32 | 0.024 b | 49.32 | 0.013 cd |
| (B. cereus + 10 + w.w) | 21.42 | 0.031 abc | 18.32 | 0.026 ab | 17.73 | 0.019 ac | 42.54 | 0.04 ab | 31.21 | 0.024 ab | 29.43 | 0.013 bc |
| (L. macroides +w.w) | 26.24 | 0.025 ab | 20.32 | 0.022 ab | 15.52 | 0.094 ab | 70.31 | 0.06 ab | 39.12 | 0.031 bc | 53.23 | 0.022 b |
| (L. macroides +10 + w.w) | 22.31 | 0.034 abd | 13.32 | 0.072 ab | 15.62 | 0.017 ab | 40.21 | 0.03 ab | 30.42 | 0.032 c | 26.43 | 0.022 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akhtar, N.; Khan, S.; Jamil, M.; Rehman, S.U.; Rehman, Z.U.; Rha, E.S. Combine Effect of ZnO NPs and Bacteria on Protein and Gene’s Expression Profile of Rice (Oryza sativa L.) Plant. Toxics 2022, 10, 305. https://doi.org/10.3390/toxics10060305
Akhtar N, Khan S, Jamil M, Rehman SU, Rehman ZU, Rha ES. Combine Effect of ZnO NPs and Bacteria on Protein and Gene’s Expression Profile of Rice (Oryza sativa L.) Plant. Toxics. 2022; 10(6):305. https://doi.org/10.3390/toxics10060305
Chicago/Turabian StyleAkhtar, Nazneen, Sehresh Khan, Muhammad Jamil, Shafiq Ur Rehman, Zia Ur Rehman, and Eui Shik Rha. 2022. "Combine Effect of ZnO NPs and Bacteria on Protein and Gene’s Expression Profile of Rice (Oryza sativa L.) Plant" Toxics 10, no. 6: 305. https://doi.org/10.3390/toxics10060305
APA StyleAkhtar, N., Khan, S., Jamil, M., Rehman, S. U., Rehman, Z. U., & Rha, E. S. (2022). Combine Effect of ZnO NPs and Bacteria on Protein and Gene’s Expression Profile of Rice (Oryza sativa L.) Plant. Toxics, 10(6), 305. https://doi.org/10.3390/toxics10060305