
Histological and Chemical Analysis of Heavy Metals in Kidney and Gills of Boops boops: Melanomacrophages Centers and Rodlet Cells as Environmental Biomarkers

 ,
,  ,
,  ,
,  ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Tissue Preparation for Histological Evaluation
2.3. Histology and Histochemistry
2.4. Immunohistochemistry
2.5. Chemical Analysis-Instrumentation
2.6. Chemicals and Standard Solutions
2.7. Sample Preparation
2.8. Sample Analysis by ICP-OES
2.9. Sample Analysis by ICP-MS
2.10. Statistical Analysis
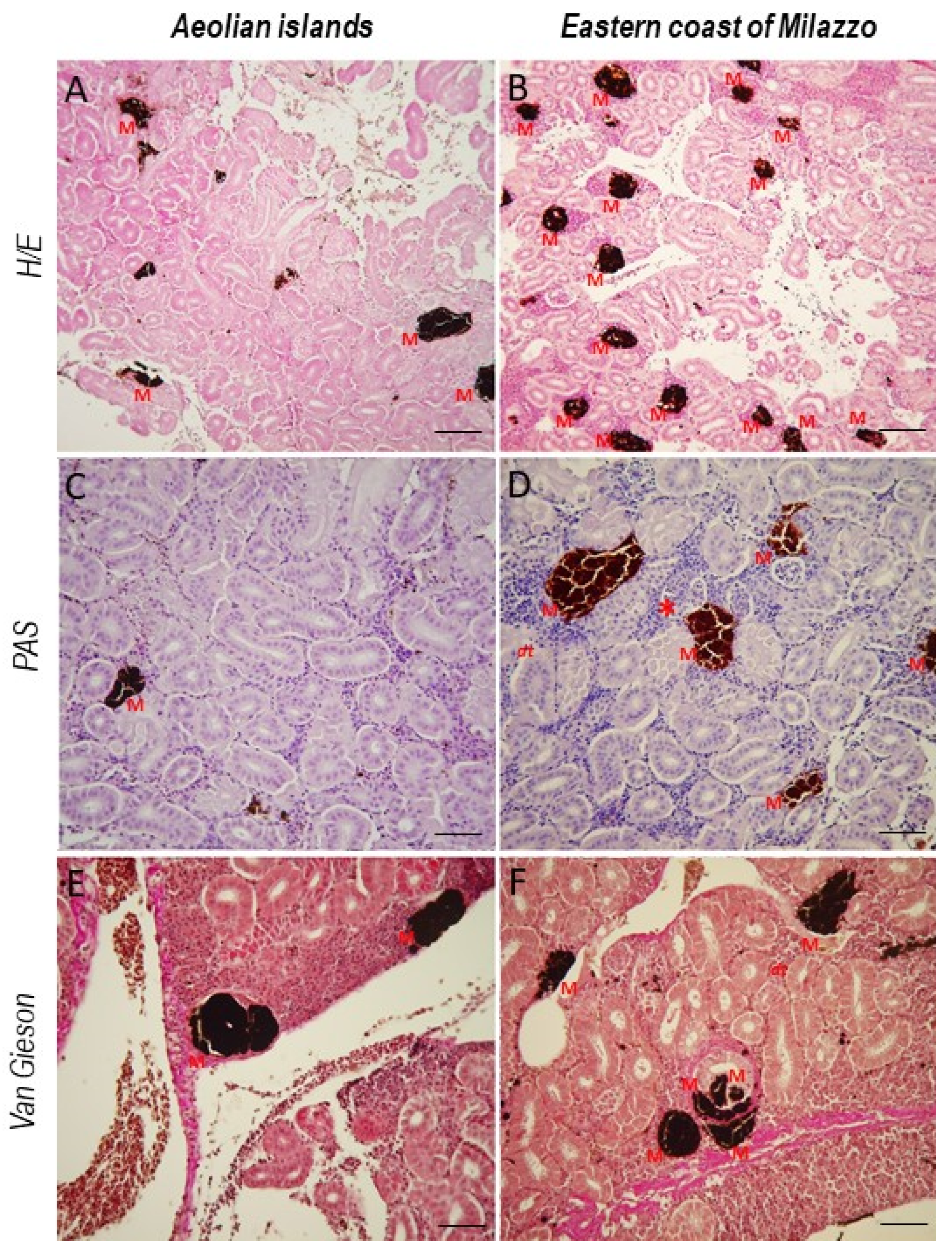
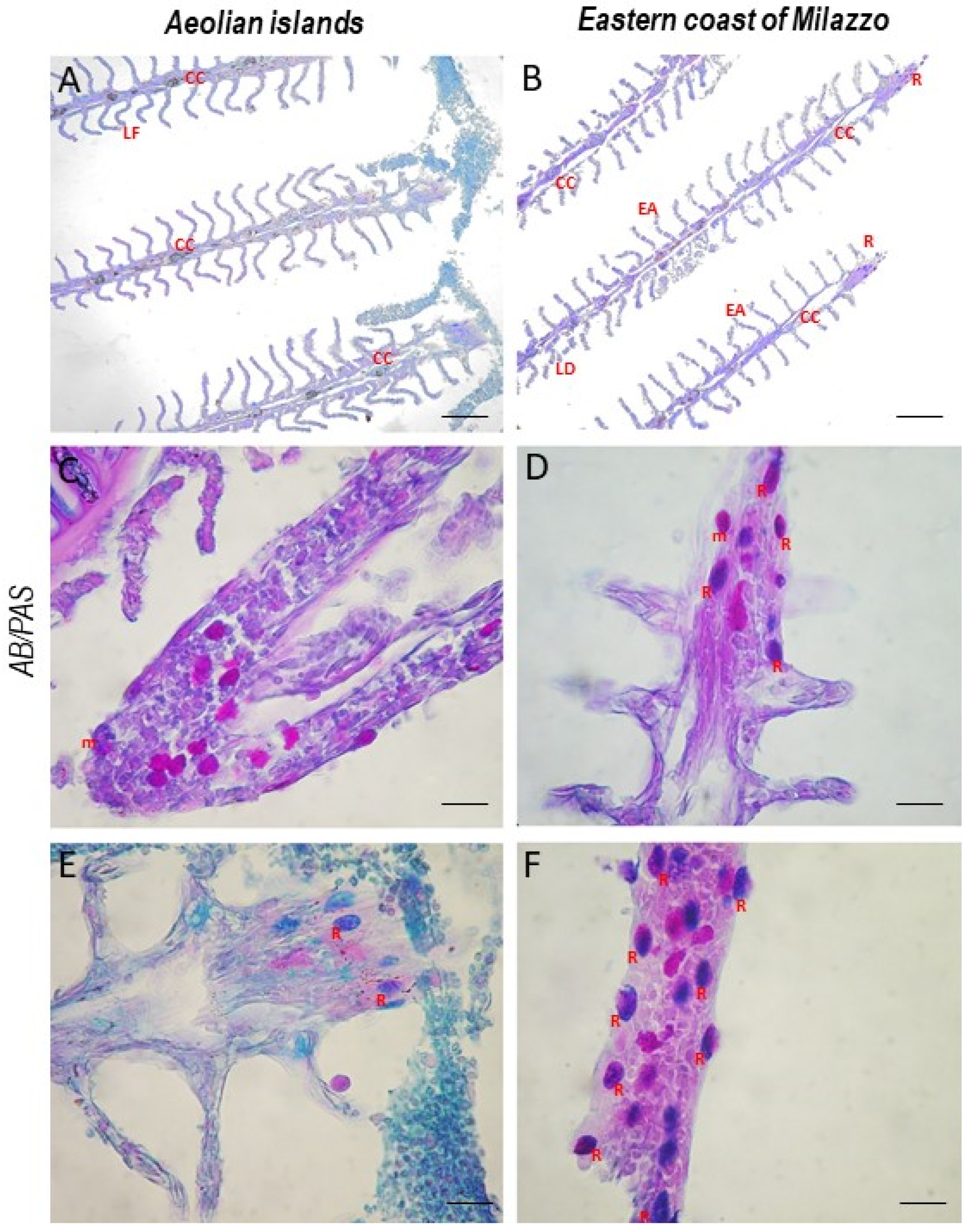
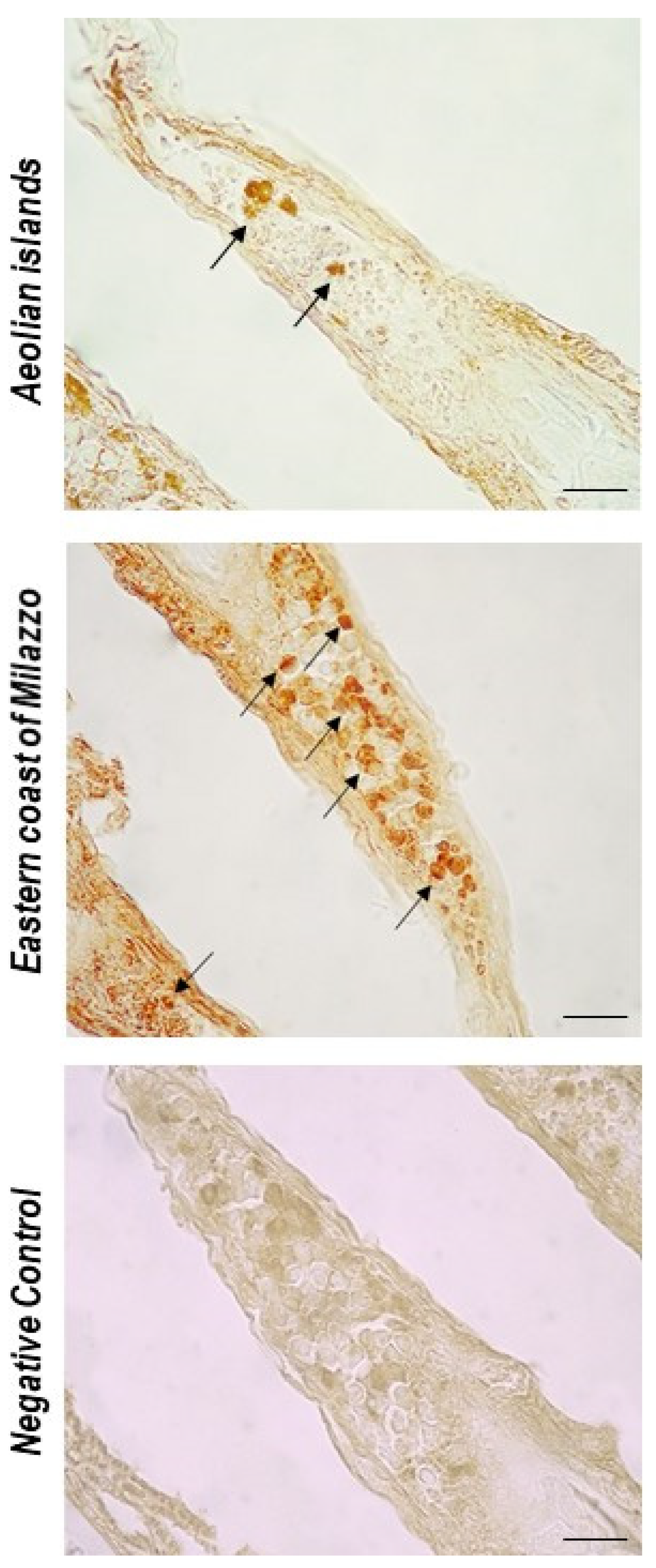
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Khan, M.S.; Javed, M.; Rehman, T.; Urooj, M.; Ahmad, I. Heavy metal pollution and risk assessment by the battery of toxicity tests. Sci. Rep. 2020, 10, 16593. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Catalini, C.; Goldfarb, A.; NET Institute Working paper, no. The Geography of Crowdfunding; NET Institute Working paper no. 10-08; National Bureau of Economic Research: Cambridge, MA, USA, 2010. [Google Scholar] [CrossRef]
- Chouvelon, T.; Spitz, J.; Caurant, F.; Mèndez-Fernandez, P.; Autier, J.; Lassus-Débat, A.; Chappuis, A.; Bustamante, P. Enhanced bioaccumulation of mercury in deep-sea fauna from the Bay of Biscay (North-East Atlantic) in relation to trophic positions identified by analysis of carbon and nitrogen stable isotopes. Deep Sea Res. Part I Oceanogr. Res. Pap. 2012, 65, 113–124. [Google Scholar] [CrossRef]
- Rajfur, M. Algae—Heavy Metals Biosorbent/Glony—Biosorbent Metali Ciężkich. Ecol. Chem. Eng. S 2013, 20, 23–40. [Google Scholar] [CrossRef][Green Version]
- Mormede, S.; Davies, I.M. Horizontal and vertical distribution of organic contaminants in deep-sea fish species. Chemosphere 2003, 50, 563–574. [Google Scholar] [CrossRef]
- Kamble, G.B.; Muley, D.V. Effect of Acute Exposure of Endusolfan and Chlor. Indian J. Environ. Sci. 2000, 4, 97–102. [Google Scholar]
- Javed, H.; Islam, A.; Chauhan, A.; Kumar, S.; Kumar, S. Efficacy of engineered GO Amberlite XAD-16 picolylamine sorbent for the trace determination of Pb (II) and Cu (II) in fishes by solid phase extraction column coupled with inductively coupled plasma optical emission spectrometry. Sci. Rep. 2018, 8, 17560. [Google Scholar] [CrossRef]
- Nriagu, J.O.; Pacyna, J.M. Quantitative assessment of worldwide contamination of air, water and soils by trace metals. Nature 1988, 333, 134–139. [Google Scholar] [CrossRef]
- Lenntech, K. Water Treatment and Air Purification; Lenntech: Rotterdam, The Netherlands, 2004. [Google Scholar]
- Adeboyejo, A.O.; Clarke, E.O.; Hammed, A.M.; Adaramoye, R.O. Haematological and Hepatic Responses of the African Catfish Clarias gariepinus to Sublethal Exposure of Industrial Effluents from Ologe Lagoon Environs, Lagos, Nigeria. J. Food Sci. Eng. 2018, 8, 198–209. [Google Scholar] [CrossRef][Green Version]
- Di Bella, G.; Potortì, A.G.; Turco, V.L.; Bua, D.; Licata, P.; Cicero, N.; Dugo, G. Trace elements inThunnus thynnusfrom Mediterranean Sea and benefit–risk assessment for consumers. Food Addit. Contam. Part B 2015, 8, 175–181. [Google Scholar] [CrossRef]
- Naccari, C.; Cicero, N.; Ferrantelli, V.; Giangrosso, G.; Vella, A.; Macaluso, A.; Naccari, F.; Dugo, G. Toxic Metals in Pelagic, Benthic and Demersal Fish Species from Mediterranean FAO Zone 37. Bull. Environ. Contam. Toxicol. 2015, 95, 567–573. [Google Scholar] [CrossRef]
- Afonso, C.; Lourenço, H.M.; Pereira, C.; Martins, M.F.; Carvalho, M.L.; Castro, M.; Nunes, M.L. Total and organic mercury, selenium and α-tocopherol in some deep-water fish species. J. Sci. Food Agric. 2008, 88, 2543–2550. [Google Scholar] [CrossRef]
- Salvo, A.; Cicero, N.; Vadalà, R.; Mottese, A.F.; Bua, D.; Mallamace, D.; Giannetto, C.; Dugo, G. Toxic and essential metals determination in commercial seafood: Paracentrotus lividus by ICP-MS. Nat. Prod. Res. 2016, 30, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Cammilleri, G.; Vazzana, M.; Arizza, V.; Giunta, F.; Vella, A.; Dico, G.L.; Giaccone, V.; Giofrè, S.V.; Giangrosso, G.; Cicero, N.; et al. Mercury in fish products: What’s the best for consumers between bluefin tuna and yellowfin tuna? Nat. Prod. Res. 2018, 32, 457–462. [Google Scholar] [CrossRef]
- Graci, S.; Collura, R.; Cammilleri, G.; Buscemi, M.D.; Giangrosso, G.; Principato, D.; Gervasi, T.; Cicero, N.; Ferrantelli, V. Mercury accumulation in Mediterranean Fish and Cephalopods Species of Sicilian coasts: Correlation between pollution and the presence of Anisakis parasites. Nat. Prod. Res. 2017, 31, 1156–1162. [Google Scholar] [CrossRef] [PubMed]
- Morcillo, P.; Esteban, M.; Cuesta, A. Heavy metals produce toxicity, oxidative stress and apoptosis in the marine teleost fish SAF-1 cell line. Chemosphere 2016, 144, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Feist, S.; Stentiford, G.; Kent, M.; Santos, A.R.; Lorance, P. Histopathological assessment of liver and gonad pathology in continental slope fish from the northeast Atlantic Ocean. Mar. Environ. Res. 2015, 106, 42–50. [Google Scholar] [CrossRef] [PubMed]
- La Torre, G.L.; Cicero, N.; Bartolomeo, G.; Rando, R.; Vadalà, R.; Santini, A.; Durazzo, A.; Lucarini, M.; Dugo, G.; Salvo, A. Assessment and Monitoring of Fish Quality from a Coastal Ecosystem under High Anthropic Pressure: A Case Study in Southern Italy. Int. J. Environ. Res. Public Health 2020, 17, 3285. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, F.; Nava, V.; Giarratana, F.; Gervasi, T.; Cicero, N. Assessment of heavy- and semi-metals contamination in edible seaweed and dried fish sold in ethnic food stores on the Italian market. J. Food Compos. Anal. 2021, 104, 104150. [Google Scholar] [CrossRef]
- Reish, D.L.; Oshida, P.S. (Eds.) Manual of Methods in Aquatic Environment Research. 10: Short-Term Static Bioassays/by Donald L. Reish and Philip S. Oshida; FAO Fisheries Technical Paper: Rome, Italy, 1987; ISBN 978-92-5-102517-8. [Google Scholar]
- Kaur, S.; Khera, K.S.; Kondal, J.K. Heavy Metal Induced Histopathological Alterations in Liver, Muscle and Kidney of Freshwater Cyprinid, Labeo Rohita (Hamilton). J. Entomol. Zool. Stud. 2018, 6, 2137–2144. [Google Scholar]
- Adams, S.M. Biological Indicators of Aquatic Ecosystem Stress; American Fisheries Society: New York, NY, USA, 2002. [Google Scholar]
- Gernhöfer, M.; Pawert, M.; Schramm, M.; Müller, E.; Triebskorn, R. Ultrastructural biomarkers as tools to characterize the health status of fish in contaminated streams. J. Aquat. Ecosyst. Stress Recover. 2001, 8, 241–260. [Google Scholar] [CrossRef]
- Camargo, M.M.P.; Martinez, C.B.R. Histopathology of gills, kidney and liver of a Neotropical fish caged in an urban stream. Neotrop. Ichthyol. 2007, 5, 327–336. [Google Scholar] [CrossRef]
- Thophon, S.; Kruatrachue, M.; Upatham, E.; Pokethitiyook, P.; Sahaphong, S.; Jaritkhuan, S. Histopathological alterations of white seabass, Lates calcarifer, in acute and subchronic cadmium exposure. Environ. Pollut. 2003, 121, 307–320. [Google Scholar] [CrossRef]
- Mabrouk, D.B.M. Aspects of Manzalah Lake Pollution on Mugil Species. Ph.D. Thesis, Ain Shams University, El Cairo, Egypt, 2004. [Google Scholar]
- Shah, N.; Khan, A.; Ali, R.; Marimuthu, K.; Uddin, M.N.; Rizwan, M.; Rahman, K.U.; Alam, M.; Adnan, M.; Muhammad; et al. Monitoring Bioaccumulation (in Gills and Muscle Tissues), Hematology, and Genotoxic Alteration in Ctenopharyngodon idella Exposed to Selected Heavy Metals. BioMed Res. Int. 2020, 2020, 6185231. [Google Scholar] [CrossRef] [PubMed]
- Viana, H.; Jesus, W.; Silva, S.; Jorge, M.; Santos, D.; Neta, R.C. Aggregation of hepatic melanomacrophage centers in S. herzbergii (Pisces, Ariidae) as indicators of environmental change and well-being. Arq. Bras. Med. Vet. Zootec. 2021, 73, 868–876. [Google Scholar] [CrossRef]
- Fishelson, L. Cytomorphological alterations of the thymus, spleen, head-kidney, and liver in cardinal fish (Apogonidae, Teleostei) as bioindicators of stress. J. Morphol. 2006, 267, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.; Younes, H.A. Melanomacrophage centers in Clarias gariepinus as an immunological biomarker for toxicity of silver nanoparticles. J. Microsc. Ultrastruct. 2017, 5, 97–104. [Google Scholar] [CrossRef]
- Naguib, M.; Mahmoud, U.M.; Mekkawy, I.A.; Sayed, A.E.-D.H. Hepatotoxic effects of silver nanoparticles on Clarias gariepinus; Biochemical, histopathological, and histochemical studies. Toxicol. Rep. 2020, 7, 133–141. [Google Scholar] [CrossRef]
- Manrique, W.G.; Claudiano, G.D.S.; Petrillo, T.R.; De Castro, M.P.; Figueiredo, M.; Belo, M.A.D.A.; De Moraes, J.R.E.; de Moraes, F.R. Response of splenic melanomacrophage centers of Oreochromis niloticus (Linnaeus, 1758) to inflammatory stimuli by BCG and foreign bodies. J. Appl. Ichthyol. 2014, 30, 1001–1006. [Google Scholar] [CrossRef]
- Dang, T.S.M. Melanomacrophage Centers and Mucus Cells of Shorthorn Sculpin (Myoxocephalus scorpius) as Biomarkers of Contaminants. Ph.D. Thesis, University of Tasmania, Hobart, Australia, 2021. [Google Scholar]
- Dang, M.; Nørregaard, R.; Sonne, C.; Bach, L.; Stride, M.; Jantawongsri, K.; Nowak, B. Splenic and renal melanomacrophage centers in shorthorn sculpins (Myoxocephalus scorpius) in Nuuk harbor, West Greenland. Polar Biol. 2021, 44, 2011–2021. [Google Scholar] [CrossRef]
- Leino, R.L. The Contractile Mechanism and ‘“Holocrine”’Secretion in Teleost Rodlet Cells. In Proceedings of the 53rd Annual Proceeding of the Scandinavian Society for Electron Microscopy, Tampere, Finland, 12–15 June 2002. [Google Scholar]
- Walaa, E.; Marwa, F.; Zeinab, A.-A. Responses of the rodlet cells to metacercarial infections in oreochromis niloticus. Assiut Vet. Med. J. 2019, 65, 59–71. [Google Scholar] [CrossRef]
- Vigliano, F.; Bermúdez, R.; Nieto, J.; Quiroga, M. Development of rodlet cells in the gut of turbot (Psetta maxima L.): Relationship between their morphology and S100 protein immunoreactivity. Fish Shellfish Immunol. 2009, 26, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Pergolizzi, S.; Capillo, G.; Cascio, P.L.; Lauriano, E.R. Rodlet cells in kidney of goldfish (Carassius auratus, Linnaeus 1758): A light and confocal microscopy study. Acta Histochem. 2022, 124, 151876. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Lauriano, E.R.; Aragona, M.; Capillo, G.; Pergolizzi, S. Marking vertebrates langerhans cells, from fish to mammals. Acta Histochem. 2020, 122, 151622. [Google Scholar] [CrossRef] [PubMed]
- De Souza Araujo, N.; Borges, J.C.S. Rodlet Cells Changes in Oreochromis Niloticus in Response to Organophosphate Pesticide and Their Relevance as Stress Biomarker in Teleost Fishes. Int. J. Aquat. Biol. 2016, 3, 398–408. [Google Scholar]
- Matisz, C.E.; Goater, C.P.; Bray, D. Density and maturation of rodlet cells in brain tissue of fathead minnows (Pimephales promelas) exposed to trematode cercariae. Int. J. Parasitol. 2010, 40, 307–312. [Google Scholar] [CrossRef]
- De Pasquale, J.A. Tyrosine phosphatase inhibitor triggers rodlet cell discharge in sunfish scale epidermis cultures. Acta Zool. 2014, 95, 209–219. [Google Scholar] [CrossRef]
- Manera, M.; Dezfuli, B.S. Rodlet cells in teleosts: A new insight into their nature and functions. J. Fish Biol. 2004, 65, 597–619. [Google Scholar] [CrossRef]
- Kramer, C.R.; Hongach, S.W.; Antonov, K.; Qama, E. Are rodlet cells reliable biomarkers in Fundulus heteroclitus (L.)? J. Morphol. Sci. 2016, 33, 205–214. [Google Scholar] [CrossRef]
- Giari, L.; Simoni, E.; Manera, M.; Dezfuli, B.S. Histo-cytological responses of Dicentrarchus labrax (L.) following mercury exposure. Ecotoxicol. Environ. Saf. 2008, 70, 400–410. [Google Scholar] [CrossRef]
- Poltronieri, C.; Laurà, R.; Bertotto, D.; Negrato, E.; Simontacchi, C.; Guerrera, M.C.; Radaelli, G. Effects of exposure to overcrowding on rodlet cells of the teleost fish Dicentrarchus labrax (L.). Vet. Res. Commun. 2009, 33, 619–629. [Google Scholar] [CrossRef]
- Panda, A.; Patri, M. Histopathological Changes and Presence of Rodlet Cells in Different Organs of Common Finfish Liza Tade in Brackish Water of the Chilika Lagoon. Int. J. Fish. Aquat. Stud. 2020, 8, 186–196. [Google Scholar]
- Zaccone, G.; Lauriano, E.; Silvestri, G.; Kenaley, C.; Icardo, J.M.; Pergolizzi, S.; Alesci, A.; Sengar, M.; Kuciel, M.; Gopesh, A. Comparative neurochemical features of the innervation patterns of the gut of the basal actinopterygian, Lepisosteus oculatus, and the euteleost, Clarias batrachus. Acta Zool. 2015, 96, 127–139. [Google Scholar] [CrossRef]
- Lauriano, E.R.; Żuwała, K.; Kuciel, M.; Budzik, K.A.; Capillo, G.; Alesci, A.; Pergolizzi, S.; Dugo, G.; Zaccone, G. Confocal immunohistochemistry of the dermal glands and evolutionary considerations in the caecilian, Typhlonectes natans (Amphibia: Gymnophiona). Acta Zool. 2016, 97, 154–164. [Google Scholar] [CrossRef]
- Pergolizzi, S.; Rizzo, G.; Favaloro, A.; Alesci, A.; Pallio, S.; Melita, G.; Cutroneo, G.; Lauriano, E.R. Expression of VAChT and 5-HT in Ulcerative colitis dendritic cells. Acta Histochem. 2021, 123, 151715. [Google Scholar] [CrossRef] [PubMed]
- Pergolizzi, S.; Alesci, A.; Centofanti, A.; Aragona, M.; Pallio, S.; Magaudda, L.; Cutroneo, G.; Lauriano, E.R. Role of Serotonin in the Maintenance of Inflammatory State in Crohn’s Disease. Biomedicines 2022, 10, 765. [Google Scholar] [CrossRef] [PubMed]
- Lauriano, E.R.; Icardo, J.M.; Zaccone, D.; Kuciel, M.; Satora, L.; Alesci, A.; Alfa, M.; Zaccone, G. Expression patterns and quantitative assessment of neurochemical markers in the lung of the gray bichir, Polypterus senegalus (Cuvier, 1829). Acta Histochem. 2015, 117, 738–746. [Google Scholar] [CrossRef]
- Zaccone, G.; Lauriano, E.R.; Kuciel, M.; Capillo, G.; Pergolizzi, S.; Alesci, A.; Ishimatsu, A.; Ip, Y.K.; Icardo, J.M. Identification and distribution of neuronal nitric oxide synthase and neurochemical markers in the neuroepithelial cells of the gill and the skin in the giant mudskipper, Periophthalmodon schlosseri. Zoology 2017, 125, 41–52. [Google Scholar] [CrossRef]
- Alesci, A.; Cicero, N.; Salvo, A.; Palombieri, D.; Zaccone, D.; Dugo, G.; Bruno, M.; Vadalà, R.; Lauriano, E.R.; Pergolizzi, S. Extracts deriving from olive mill waste water and their effects on the liver of the goldfish Carassius auratus fed with hypercholesterolemic diet. Nat. Prod. Res. 2014, 28, 1343–1349. [Google Scholar] [CrossRef]
- Alesci, A.; Salvo, A.; Lauriano, E.R.; Gervasi, T.; Palombieri, D.; Bruno, M.; Pergolizzi, S.; Cicero, N. Production and extraction of astaxanthin from Phaffia rhodozyma and its biological effect on alcohol-induced renal hypoxia in Carassius auratus. Nat. Prod. Res. 2015, 29, 1122–1126. [Google Scholar] [CrossRef]
- Alessio, A.; Pergolizzi, S.; Gervasi, T.; Aragona, M.; Cascio, P.L.; Cicero, N.; Lauriano, E.R. Biological effect of astaxanthin on alcohol-induced gut damage in Carassius auratus used as experimental model. Nat. Prod. Res. 2021, 35, 5737–5743. [Google Scholar] [CrossRef]
- Alesci, A.; Pergolizzi, S.; Fumia, A.; Calabrò, C.; Cascio, P.L.; Lauriano, E.R. Mast cells in goldfish (Carassius auratus) gut: Immunohistochemical characterization. Acta Zool. 2022; in press. [Google Scholar] [CrossRef]
- Bua, D.G.; Annuario, G.; Albergamo, A.; Cicero, N.; Dugo, G. Heavy metals in aromatic spices by inductively coupled plasma-mass spectrometry. Food Addit. Contam. Part B 2016, 9, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Turco, V.L.; Di Bella, G.; Furci, P.; Cicero, N.; Pollicino, G.; Dugo, G. Heavy metals content by ICP-OES in Sarda sarda, Sardinella aurita and Lepidopus caudatus from the Strait of Messina (Sicily, Italy). Nat. Prod. Res. 2013, 27, 518–523. [Google Scholar] [CrossRef]
- Salvo, A.; Potorti, A.G.; Cicero, N.; Bruno, M.; Turco, V.L.; Di Bella, G.; Dugo, G. Statistical characterisation of heavy metal contents in Paracentrotus lividusfrom Mediterranean Sea. Nat. Prod. Res. 2014, 28, 718–726. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Abah, J.; Mashebe, P.; Sylvanus, O.A. Preliminary Assessment of Some Heavy Metals Pollution Status of Lisikili River Water in Zambezi Region, Namibia. Int. J. Environ. Pollut. Res. 2016, 4, 13–30. [Google Scholar]
- Iqbal, F.; Qureshi, I.Z.; Ali, M. Histopathological Changes in the Liver of a Farmed Cyprinid Fish, Cyprinus Carpio, Following Exposure to Nitrate. Pak. J. Zool. 2005, 37, 297–300. [Google Scholar]
- Gupta, P.; Srivastava, N. Effects of Sub-Lethal Concentrations of Zinc on Histological Changes and Bioaccumulation of Zinc by Kidney of Fish Channa Punctatus (Bloch). J. Environ. Biol. 2006, 27, 211–215. [Google Scholar]
- Younis, E.; Abdel-Warith, A.-W.; Al-Asgah, N.; Ebaid, H. Histopathological alterations in the liver and intestine of Nile tilapia Oreochromis niloticus exposed to long-term sublethal concentrations of cadmium chloride. Chin. J. Oceanol. Limnol. 2015, 33, 846–852. [Google Scholar] [CrossRef]
- Rana, M.A.; Jabeen, F.; Shabbir, S.; Naureen, A.; Sultana, K.; Ahmad, I.; Shabnam, M. Histopathological Study of Liver and Kidney in Common Carp (Cyprinus Carpio) Exposed to Different Doses of Potassium Dichromate. Int. J. Biosci. 2015, 6, 108–116. [Google Scholar]
- Al-Bairuty, G.; Shaw, B.J.; Handy, R.D.; Henry, T. Histopathological effects of waterborne copper nanoparticles and copper sulphate on the organs of rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2013, 126, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Hermenean, A.; Damache, G.; Albu, P.; Ardelean, A.; Ardelean, G.; Ardelean, D.P.; Horge, M.; Nagy, T.; Braun, M.; Zsuga, M.; et al. Histopatological alterations and oxidative stress in liver and kidney of Leuciscus cephalus following exposure to heavy metals in the Tur River, North Western Romania. Ecotoxicol. Environ. Saf. 2015, 119, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Hermenean, A.; Gheorghiu, G.; Stan, M.S.; Herman, H.; Onita, B.; Ardelean, D.P.; Ardelean, A.; Braun, M.; Zsuga, M.; Kéki, S.; et al. Biochemical, Histopathological and Molecular Responses in Gills of Leuciscus cephalus Exposed to Metals. Arch. Environ. Contam. Toxicol. 2017, 73, 607–618. [Google Scholar] [CrossRef]
- Raju, K.V.; Somashekar, R.K.; Prakash, K.L. Metal Concentration in Fresh Water Fish Organs. Open J. Met. 2013, 3, 23–28. [Google Scholar] [CrossRef]
- Fonseca, A.; Fernandes, L.F.S.; Fontaínhas-Fernandes, A.; Monteiro, S.M.; Pacheco, F. The impact of freshwater metal concentrations on the severity of histopathological changes in fish gills: A statistical perspective. Sci. Total Environ. 2017, 599–600, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Mladin, B.O.; Albu, P.; Herman, H.; Balta, C.; Lazar, V.; Fulop, A.; Baranyai, E.; Harangi, S.; Keki, S.; Nagy, L.; et al. Correlation between Heavy Metal-Induced Histopathological Changes and Trophic Interactions between Different Fish Species. Appl. Sci. 2021, 11, 3760. [Google Scholar] [CrossRef]
- Ayoola, S.O.; Alajabo, O.T. Acute Toxicity and Histopathological Effects of Engine Oil on Sarotherodon Melanotheron (Black Jaw Tilapia). Am.-Eurasian J. Toxicol. Sci. 2012, 4, 48–55. [Google Scholar]
- Abdelhafeez, H.; Soliman, S. Origin of Rodlet Cells and Mapping Their Distribution in Ruby-Red-Fin Shark (Rainbow Shark) Epalzeorhynchos frenatum (Teleostei: Cyprinidae): Light, Immunohistochemistry and Ultrastructure Study. J. Cytol. Histol. 2016, 7, 1000435. [Google Scholar] [CrossRef]
- Marenholz, I.; Heizmann, C.W.; Fritz, G. S100 proteins in mouse and man: From evolution to function and pathology (including an update of the nomenclature). Biochem. Biophys. Res. Commun. 2004, 322, 1111–1122. [Google Scholar] [CrossRef]
- Mokhtar, D.; Abdelhafeez, H. Light- and electron-microscopic studies of olfactory organ of Red-tail shark, Epalzeorhynchos bicolor (Teleostei: Cyprinidae). J. Microsc. Ultrastruct. 2014, 2, 182. [Google Scholar] [CrossRef]
- Reite, O.B.; Evensen, Ø. Inflammatory cells of teleostean fish: A review focusing on mast cells/eosinophilic granule cells and rodlet cells. Fish Shellfish Immunol. 2006, 20, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.C.; Smith, S.A.; Jortner, B.S.; McMaster, M.E. Putative Rodlet Cell Neoplasms in the Livers of Two White Suckers (Catostomus commersonii). J. Comp. Pathol. 2018, 164, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Stosik, M.P.; Tokarz-Deptuła, B.; Deptuła, W. Melanomacrophages and melanomacrophage centres in Osteichthyes. Cent. Eur. J. Immunol. 2019, 44, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Silphaduang, U.; Colorni, A.; Noga, E.J. Evidence for widespread distribution of piscidin antimicrobial peptides in teleost fish. Dis. Aquat. Org. 2006, 72, 241–252. [Google Scholar] [CrossRef]
- Zaccone, D.; Icardo, J.M.; Kuciel, M.; Alesci, A.; Pergolizzi, S.; Satora, L.; Lauriano, E.R.; Zaccone, G. Polymorphous granular cells in the lung of the primitive fish, the bichir Polypterus senegalus. Acta Zool. 2017, 98, 13–19. [Google Scholar] [CrossRef]
- Maina, J.N.; Icardo, J.M.; Zaccone, G.; Aragona, M.; Lauriano, E.R.; Alesci, A.; Albano, M.; Guerrera, M.C.; Germana, A.; Fernandes, J.M.O.; et al. Immunohistochemical and ultrastructural study of the immune cell system and epithelial surfaces of the respiratory organs in the bimodally breathing African sharptooth catfish (Clarias gariepinus Burchell, 1822). Anat. Rec. 2022, 130, 6–18. [Google Scholar] [CrossRef]
- Zaccone, G.; Capillo, G.; Fernandes, J.M.O.; Kiron, V.; Lauriano, E.R.; Alesci, A.; Cascio, P.L.; Guerrera, M.C.; Kuciel, M.; Zuwala, K.; et al. Expression of the Antimicrobial Peptide Piscidin 1 and Neuropeptides in Fish Gill and Skin: A Potential Participation in Neuro-Immune Interaction. Mar. Drugs 2022, 20, 145. [Google Scholar] [CrossRef]
- Lauriano, E.; Silvestri, G.; Kuciel, M.; Żuwała, K.; Zaccone, D.; Palombieri, D.; Alesci, A.; Pergolizzi, S. Immunohistochemical localization of Toll-like receptor 2 in skin Langerhans’ cells of striped dolphin (Stenella coeruleoalba). Tissue Cell 2014, 46, 113–121. [Google Scholar] [CrossRef]
- Lauriano, E.R.; Pergolizzi, S.; Capillo, G.; Kuciel, M.; Alesci, A.; Faggio, C. Immunohistochemical characterization of Toll-like receptor 2 in gut epithelial cells and macrophages of goldfish C arassius auratus fed with a high-cholesterol diet. Fish Shellfish Immunol. 2016, 59, 250–255. [Google Scholar] [CrossRef]
- Marino, A.; Pergolizzi, S.; Lauriano, E.R.; Santoro, G.; Spataro, F.; Cimino, F.; Speciale, A.; Nostro, A.; Bisignano, G. TLR2 activation in corneal stromal cells by Staphylococcus aureus-induced keratitis. APMIS 2015, 123, 163–168. [Google Scholar] [CrossRef]
- Marino, A.; Pergolizzi, S.; Cimino, F.; Lauriano, E.R.; Speciale, A.; D’Angelo, V.; Sicurella, M.; Argnani, R.; Manservigi, R.; Marconi, P. Role of Herpes Simplex Envelope Glycoprotein B and Toll-Like Receptor 2 in Ocular Inflammation: An ex vivo Organotypic Rabbit Corneal Model. Viruses 2019, 11, 819. [Google Scholar] [CrossRef] [PubMed]
- Lauriano, E.R.; Aragona, M.; Alesci, A.; Cascio, P.L.; Pergolizzi, S. Toll-Like Receptor 2 and α-Smooth Muscle Actin expressed in the tunica of a urochordate, Styela plicata. Tissue Cell 2021, 71, 101584. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Pergolizzi, S.; Cascio, P.L.; Fumia, A.; Lauriano, E.R. Neuronal regeneration: Vertebrates comparative overview and new perspectives for neurodegenerative diseases. Acta Zool. 2021, 103, 129–140. [Google Scholar] [CrossRef]
- Alesci, A.; Pergolizzi, S.; Cascio, P.L.; Capillo, G.; Lauriano, E.R. Localization of vasoactive intestinal peptide and toll-like receptor 2 immunoreactive cells in endostyle of urochordate Styela plicata (Lesueur, 1823). Microsc. Res. Technol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Miller, A.; Tardugno, R.; Pergolizzi, S. Chemical analysis, biological and therapeutic activities of Olea europaea L. extracts. Nat. Prod. Res. 2021, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Aragona, M.; Cicero, N.; Lauriano, E.R. Can nutraceuticals assist treatment and improve COVID-19 symptoms? Nat. Prod. Res. 2021, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Fumia, A.; Cascio, P.L.; Miller, A.; Cicero, N. Immunostimulant and Antidepressant Effect of Natural Compounds in the Management of COVID-19 Symptoms. J. Am. Coll. Nutr. 2021, 1–15. [Google Scholar] [CrossRef]
- Fumia, A.; Cicero, N.; Gitto, M.; Nicosia, N.; Alesci, A. Role of nutraceuticals on neurodegenerative diseases: Neuroprotective and immunomodulant activity. Nat. Prod. Res. 2021, 1–18. [Google Scholar] [CrossRef]
- Alesci, A.; Nicosia, N.; Fumia, A.; Giorgianni, F.; Santini, A.; Cicero, N. Resveratrol and Immune Cells: A Link to Improve Human Health. Molecules 2022, 27, 424. [Google Scholar] [CrossRef]
- Alesci, A.; Lauriano, E.R.; Fumia, A.; Irrera, N.; Mastrantonio, E.; Vaccaro, M.; Gangemi, S.; Santini, A.; Cicero, N.; Pergolizzi, S. Relationship between Immune Cells, Depression, Stress, and Psoriasis: Could the Use of Natural Products Be Helpful? Molecules 2022, 27, 1953. [Google Scholar] [CrossRef]
- Sayed, A.E.-D.H.; Hamed, H.S. Induction of apoptosis and DNA damage by 4-nonylphenol in African catfish (Clarias gariepinus) and the antioxidant role of Cydonia oblonga. Ecotoxicol. Environ. Saf. 2017, 139, 97–101. [Google Scholar] [CrossRef]
- Agius, C.; Roberts, R.J. Melano-macrophage centres and their role in fish pathology. J. Fish Dis. 2003, 26, 499–509. [Google Scholar] [CrossRef]
- Malik, M.S.; Bjørgen, H.; Nyman, I.B.; Wessel, Ø.; Koppang, E.O.; Dahle, M.K.; Rimstad, E. PRV-1 Infected Macrophages in Melanized Focal Changes in White Muscle of Atlantic Salmon (Salmo salar) Correlates with a Pro-Inflammatory Environment. Front. Immunol. 2021, 12, 664624. [Google Scholar] [CrossRef]
- Barst, B.D.; Bridges, K.; Korbas, M.; Roberts, A.P.; Van Kirk, K.; McNeel, K.; Drevnick, P.E. The role of melano-macrophage aggregates in the storage of mercury and other metals: An example from yelloweye rockfish (Sebastes ruberrimus): Melano-Macrophages and Metal Storage. Environ. Toxicol. Chem. 2015, 34, 1918–1925. [Google Scholar] [CrossRef]
- Parolini, M.; Iacobuzio, R.; Bassano, B.; Pennati, R.; Saino, N. Melanin-Based Skin Coloration Predicts Antioxidant Capacity in the Brown Trout (Salmo trutta). Physiol. Biochem. Zool. 2018, 91, 1026–1035. [Google Scholar] [CrossRef]
- Fiocchi, E.; Civettini, M.; Carbonara, P.; Zupa, W.; Lembo, G.; Manfrin, A. Development of molecular and histological methods to evaluate stress oxidative biomarkers in sea bass (Dicentrarchus labrax). Fish Physiol. Biochem. 2020, 46, 1577–1588. [Google Scholar] [CrossRef]
- Mustafa, S.A. Histopathology and heavy metal bioaccumulation in some tissues of Luciobarbus xanthopterus collected from Tigris River of Baghdad, Iraq. Egypt. J. Aquat. Res. 2020, 46, 123–129. [Google Scholar] [CrossRef]
- Ullah, S.; Li, Z.; Hassan, S.; Ahmad, S.; Guo, X.; Wanghe, K.; Nabi, G. Heavy metals bioaccumulation and subsequent multiple biomarkers based appraisal of toxicity in the critically endangered Tor putitora. Ecotoxicol. Environ. Saf. 2021, 228, 113032. [Google Scholar] [CrossRef]
- Authman, M.M. Use of Fish as Bio-indicator of the Effects of Heavy Metals Pollution. J. Aquac. Res. Dev. 2015, 6, 1000328. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Heavy Metal | As | Cd | Cr | Pb | Mn | Zn | Cu |
| Fish Permissible Limits (mg/Kg) | 0.5 | 1.0 | 12.0–13.0 | 1.5 | 1.0 | 100.0 | 30.0 |
| Heavy Metals in Muscle | As | Cd | Cr | Pb | Mn | Zn | Cu |
| Marine Area of Milazzo | 0.05 | 0.09 | 0.9 | 0.08 | 0.07 | 11.6 | 2.8 |
| Marine Area of Aeolian Islands | 0.02 | 0.05 | 0.04 | 0.06 | 0.04 | 10.4 | 1.9 |
| Fish Permissible Limits (on 200 g) | 0.1 | 0.2 | 2.5 | 0.3 | 0.2 | 20.0 | 6.0 |
| Heavy Metals in Muscle | As | Cd | Cr | Pb | Mn | Zn | Cu |
| Marine Area of Milazzo | 0.09 | 0.7 | 3.4 | 0.25 | 0.08 | 14.7 | 3.7 |
| Marine Area of Aeolian Islands | 0.06 | 0.09 | 1.0 | 0.1 | 0.05 | 11.8 | 2.2 |
| Fish Permissible Limits (on 200 g) | 0.1 | 0.2 | 2.5 | 0.3 | 0.2 | 20.0 | 6.0 |
| Heavy Metals in Gills | As | Cd | Cr | Pb | Mn | Zn | Cu |
| Marine Area of Milazzo | 0.08 | 0.68 | 3.7 | 0.27 | 0.14 | 18.9 | 5.7 |
| Marine Area of Aeolian Islands | 0.04 | 0.19 | 2.3 | 0.12 | 0.08 | 13.7 | 3.6 |
| Fish Permissible Limits (on 200 g) | 0.1 | 0.2 | 2.5 | 0.3 | 0.2 | 20.0 | 6.0 |
| MMCs in Kidneys | RCs in Gills | |
|---|---|---|
| Fish from the Aeolian Island coast | 36.21 ± 4.37 * | 53.39 ± 7.48 * |
| Fish from the eastern coast of Milazzo | 198.54 ± 35.32 ** | 210.67 ± 12.78 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alesci, A.; Cicero, N.; Fumia, A.; Petrarca, C.; Mangifesta, R.; Nava, V.; Lo Cascio, P.; Gangemi, S.; Di Gioacchino, M.; Lauriano, E.R. Histological and Chemical Analysis of Heavy Metals in Kidney and Gills of Boops boops: Melanomacrophages Centers and Rodlet Cells as Environmental Biomarkers. Toxics 2022, 10, 218. https://doi.org/10.3390/toxics10050218
Alesci A, Cicero N, Fumia A, Petrarca C, Mangifesta R, Nava V, Lo Cascio P, Gangemi S, Di Gioacchino M, Lauriano ER. Histological and Chemical Analysis of Heavy Metals in Kidney and Gills of Boops boops: Melanomacrophages Centers and Rodlet Cells as Environmental Biomarkers. Toxics. 2022; 10(5):218. https://doi.org/10.3390/toxics10050218
Chicago/Turabian StyleAlesci, Alessio, Nicola Cicero, Angelo Fumia, Claudia Petrarca, Rocco Mangifesta, Vincenzo Nava, Patrizia Lo Cascio, Sebastiano Gangemi, Mario Di Gioacchino, and Eugenia Rita Lauriano. 2022. "Histological and Chemical Analysis of Heavy Metals in Kidney and Gills of Boops boops: Melanomacrophages Centers and Rodlet Cells as Environmental Biomarkers" Toxics 10, no. 5: 218. https://doi.org/10.3390/toxics10050218
APA StyleAlesci, A., Cicero, N., Fumia, A., Petrarca, C., Mangifesta, R., Nava, V., Lo Cascio, P., Gangemi, S., Di Gioacchino, M., & Lauriano, E. R. (2022). Histological and Chemical Analysis of Heavy Metals in Kidney and Gills of Boops boops: Melanomacrophages Centers and Rodlet Cells as Environmental Biomarkers. Toxics, 10(5), 218. https://doi.org/10.3390/toxics10050218