
RETRACTED: Combined Effects of Potassium Perchlorate and a Neonicotinoid on Zebrafish Larvae (Danio rerio)

 ,
,  ,
, 
 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Maintenance and Embryo Collection
2.2. Survival Rate, Hatching Rate and Morphology Score
2.3. Alcian Blue PAS
2.4. Gene Expression Analysis
2.5. Lipid Peroxidation
2.6. Statistical Evaluation
3. Results
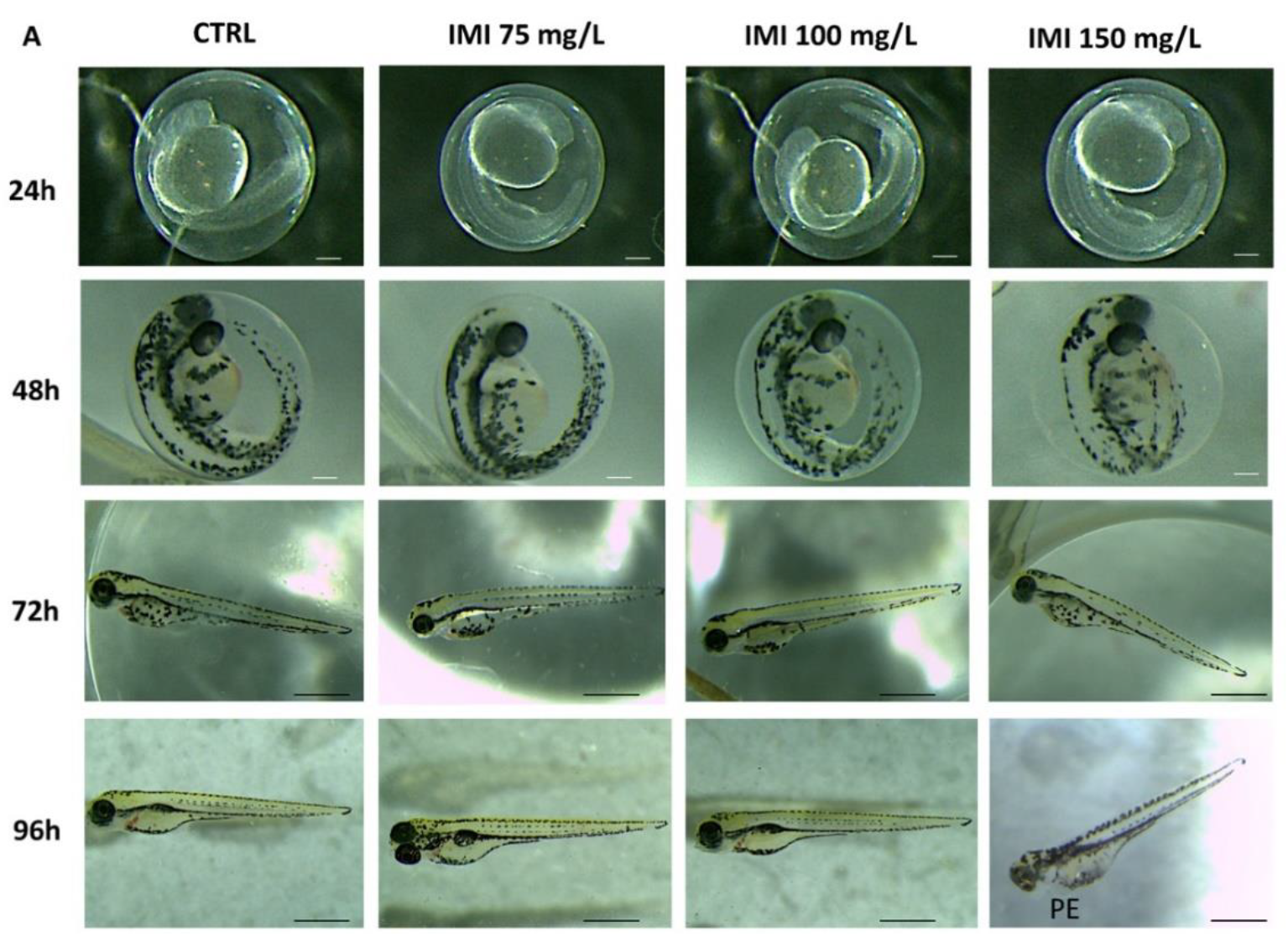
3.1. Morphology and Survival Rate
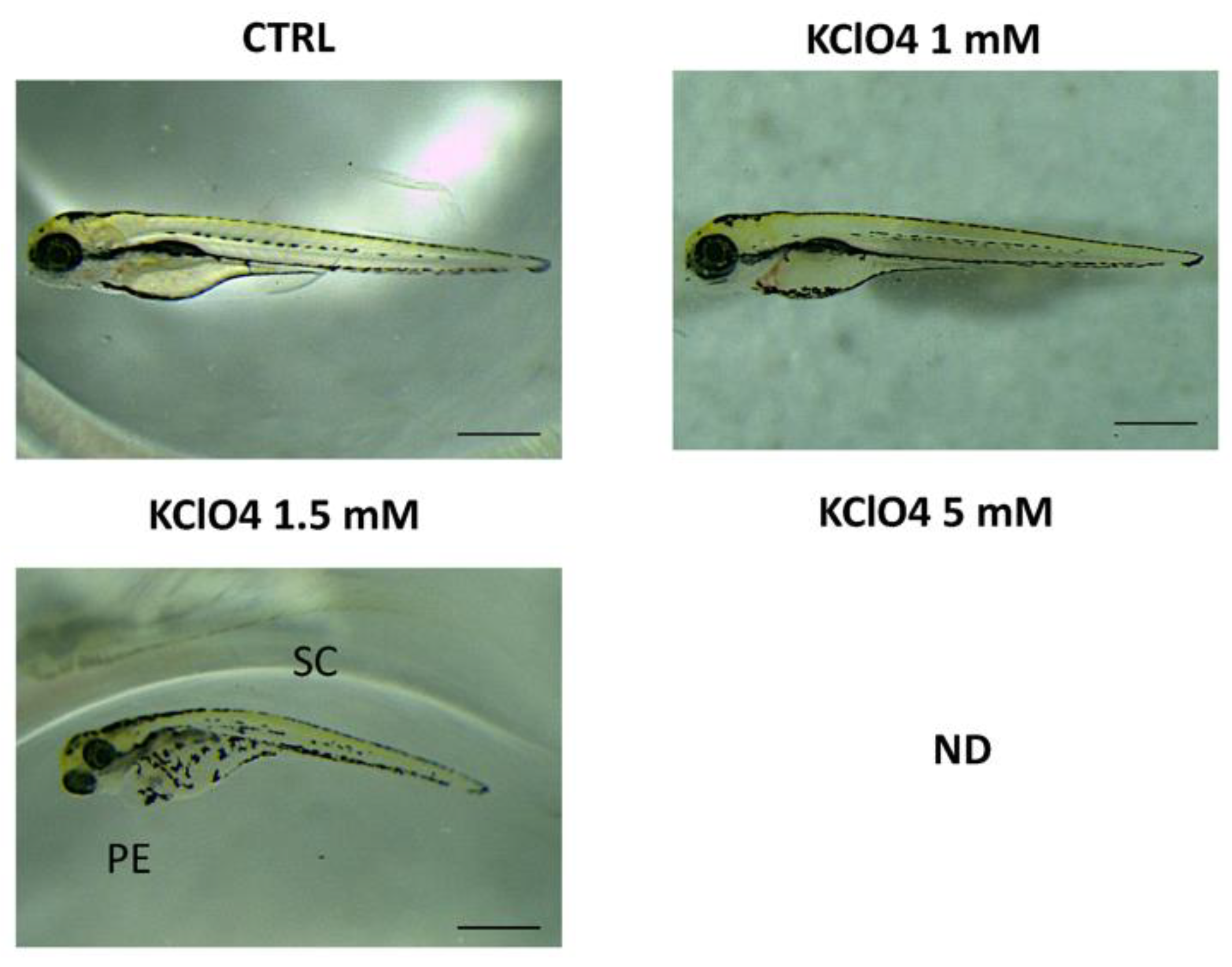
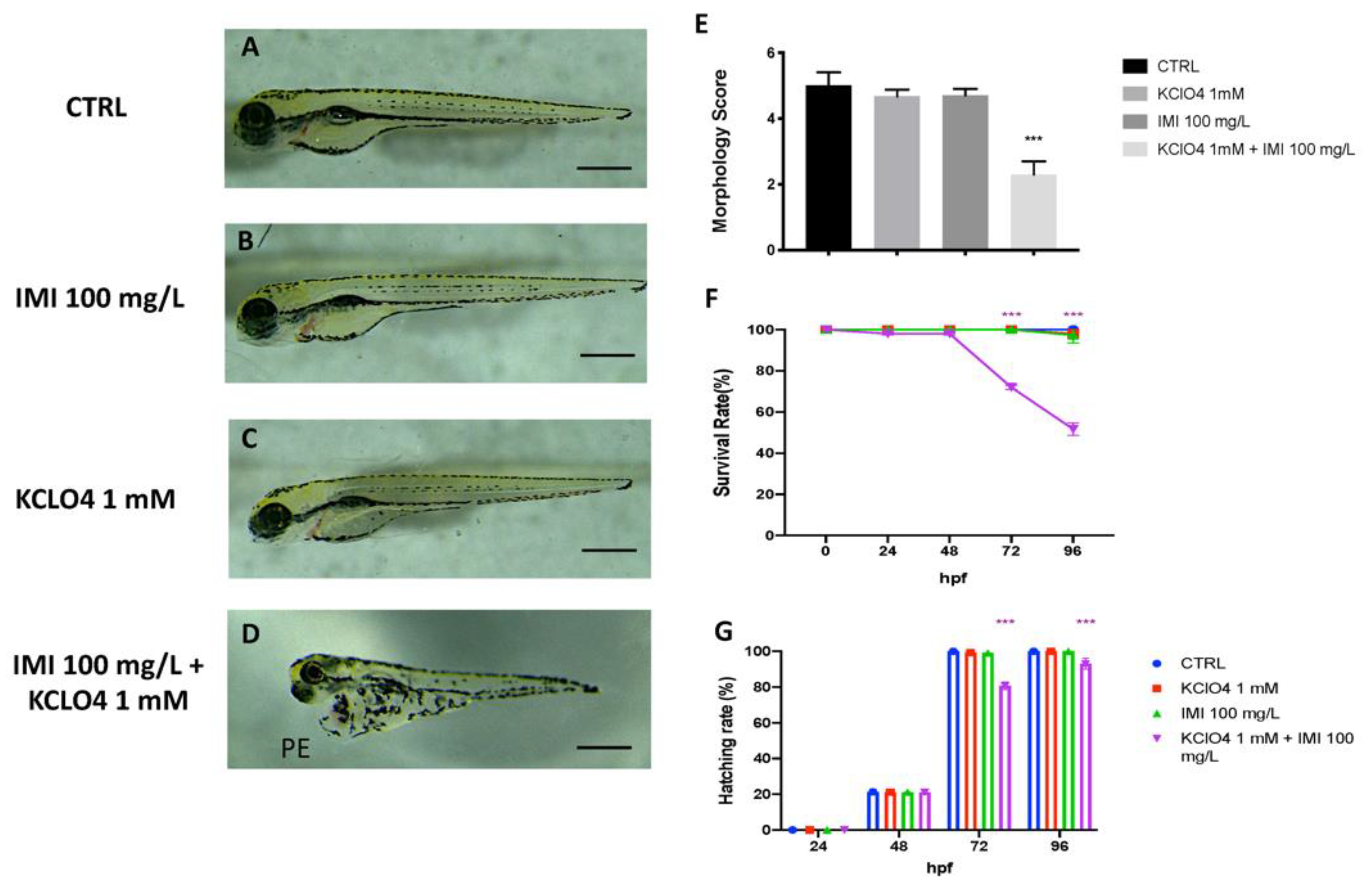
3.2. Toxic Effect of Combined Exposure to IMI and KClO4 on Malformation, Survival and Hatching
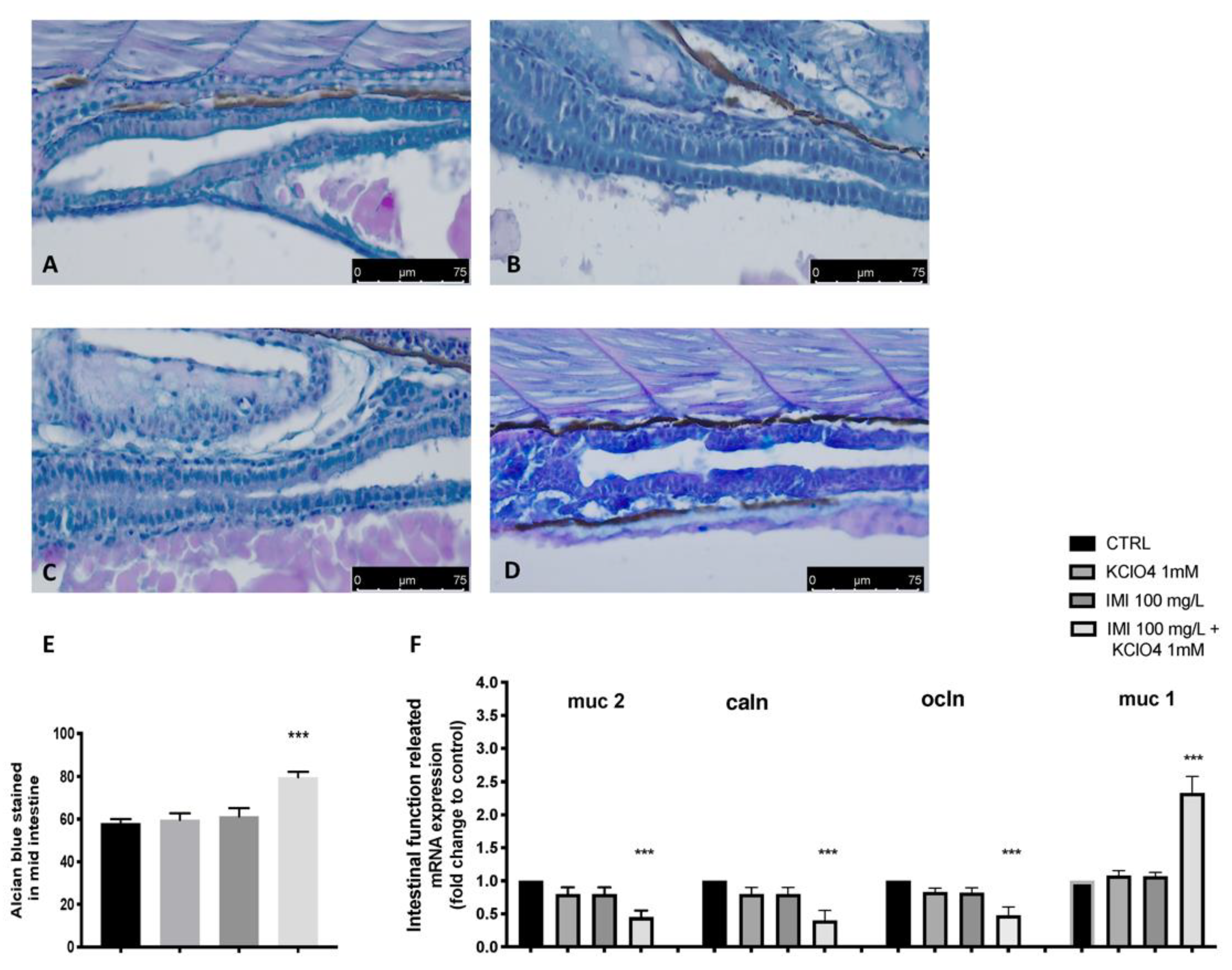
3.3. Intestinal Effect of Co-Exposure to IMI and KClO4
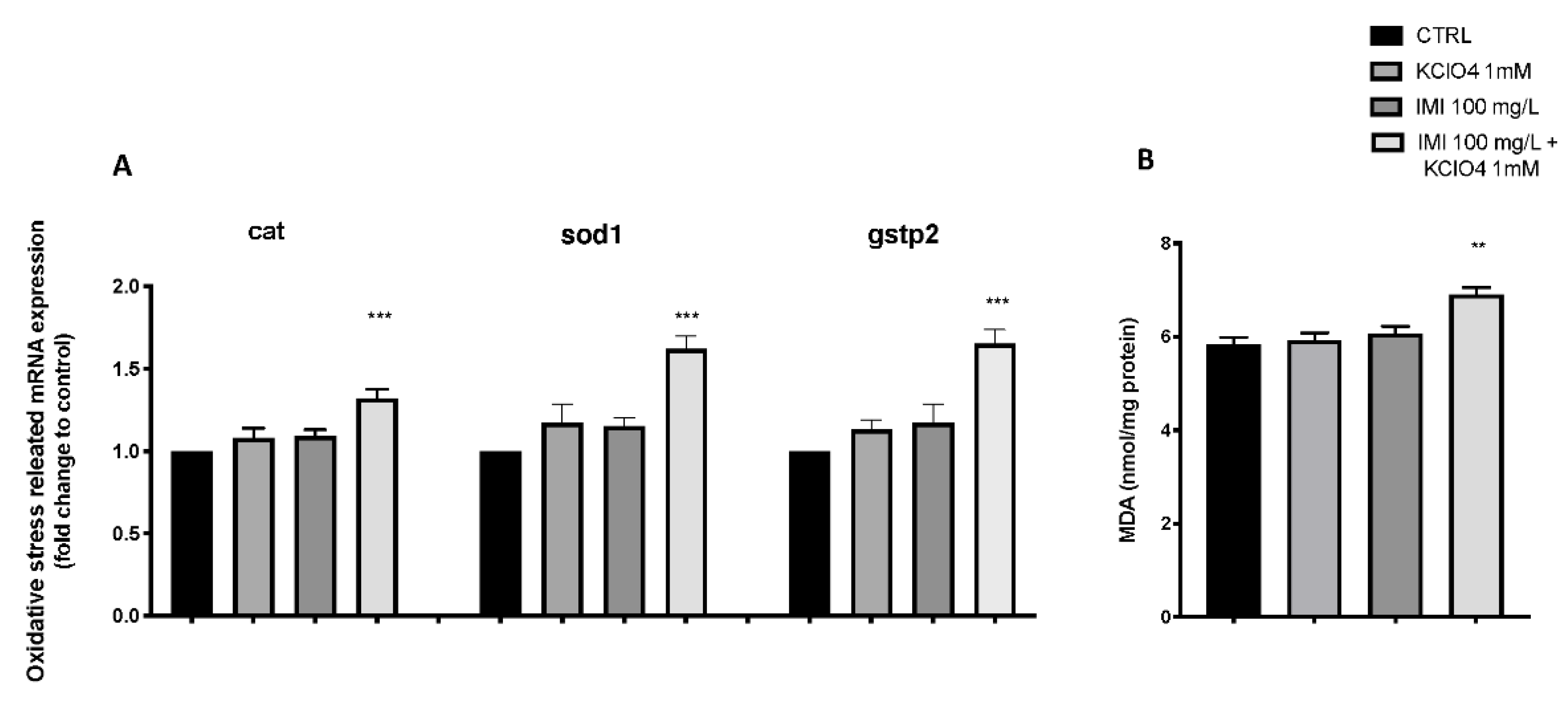
3.4. Effect of IMI and KClO4 on Antioxidant Pathway and Lipid Peroxidation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Dijk, T.C.; Van Staalduinen, M.A.; Van der Sluijs, J.P. Macro-invertebrate decline in surface water polluted with imidacloprid. PLoS ONE 2013, 8, e62374. [Google Scholar] [CrossRef]
- Simon-Delso, N.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.-M.; Chagnon, M.; Downs, C.; Furlan, L.; Gibbons, D.W.; Giorio, C.; Girolami, V. Systemic insecticides (neonicotinoids and fipronil): Trends, Uses, Mode of Action and Metabolites. Environ. Sci. Pollut. Res. 2015, 22, 5–34. [Google Scholar] [CrossRef]
- Bonmatin, J.-M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.A. Environmental fate and exposure; neonicotinoids and fipronil. Environ. Sci. Pollut. Res. 2015, 22, 35–67. [Google Scholar] [CrossRef] [PubMed]
- Moschet, C.; Wittmer, I.; Simovic, J.; Junghans, M.; Piazzoli, A.; Singer, H.; Stamm, C.; Leu, C.; Hollender, J. How a complete pesticide screening changes the assessment of surface water quality. Environ. Sci. Technol. 2014, 48, 5423–5432. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, C.A.; Mineau, P.; Devries, J.H.; Sanchez-Bayo, F.; Liess, M.; Cavallaro, M.C.; Liber, K. Neonicotinoid contamination of global surface waters and associated risk to aquatic invertebrates: A Review. Environ. Int. 2015, 74, 291–303. [Google Scholar] [CrossRef]
- Anderson, J.; Dubetz, C.; Palace, V. Neonicotinoids in the Canadian aquatic environment: A Literature Review on Current Use Products with a Focus on Fate, Exposure, and Biological Effects. Sci. Total Environ. 2015, 505, 409–422. [Google Scholar] [CrossRef]
- Hayasaka, D.; Korenaga, T.; Suzuki, K.; Saito, F.; Sánchez-Bayo, F.; Goka, K. Cumulative ecological impacts of two successive annual treatments of imidacloprid and fipronil on aquatic communities of paddy mesocosms. Ecotoxicol. Environ. Saf. 2012, 80, 355–362. [Google Scholar] [CrossRef]
- Gibbons, D.; Morrissey, C.; Mineau, P. A review of the direct and indirect effects of neonicotinoids and fipronil on vertebrate wildlife. Environ. Sci. Pollut. Res. 2015, 22, 103–118. [Google Scholar] [CrossRef]
- Wu, S.; Li, X.; Liu, X.; Yang, G.; An, X.; Wang, Q.; Wang, Y. Joint toxic effects of triazophos and imidacloprid on zebrafish (Danio rerio). Environ. Pollut. 2018, 235, 470–481. [Google Scholar] [CrossRef] [PubMed]
- Key, P.; Chung, K.; Siewicki, T.; Fulton, M. Toxicity of three pesticides individually and in mixture to larval grass shrimp (Palaemonetes pugio). Ecotoxicol. Environ. Saf. 2007, 68, 272–277. [Google Scholar] [CrossRef]
- Grung, M.; Lin, Y.; Zhang, H.; Steen, A.O.; Huang, J.; Zhang, G.; Larssen, T. Pesticide levels and environmental risk in aquatic environments in China—A review. Environ. Int. 2015, 81, 87–97. [Google Scholar] [CrossRef]
- Von Burg, R. Perchlorates. J. Appl. Toxicol. 1995, 15, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Urbansky, E.; Brown, S.; Magnuson, M.; Kelty, C. Perchlorate levels in samples of sodium nitrate fertilizer derived from Chilean caliche. Environ. Pollut. 2001, 112, 299–302. [Google Scholar] [CrossRef]
- Dasgupta, P.K.; Martinelango, P.K.; Jackson, W.A.; Anderson, T.A.; Tian, K.; Tock, R.W.; Rajagopalan, S. The origin of naturally occurring perchlorate: The Role of Atmospheric Processes. Environ. Sci. Technol. 2005, 39, 1569–1575. [Google Scholar] [CrossRef]
- Urbansky, E.T. Perchlorate chemistry: Implications for Analysis and Remediation. Bioremediation J. 1998, 2, 81–95. [Google Scholar] [CrossRef]
- Feitsma, H.; Cuppen, E. Zebrafish as a cancer model. Mol. Cancer Res. 2008, 6, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef]
- Yang, L.; Ho, N.Y.; Alshut, R.; Legradi, J.; Weiss, C.; Reischl, M.; Mikut, R.; Liebel, U.; Müller, F.; Strähle, U. Zebrafish embryos as models for embryotoxic and teratological effects of chemicals. Reprod. Toxicol. 2009, 28, 245–253. [Google Scholar] [CrossRef]
- Langheinrich, U. Zebrafish: A New Model on the Pharmaceutical Catwalk. Bioessays 2003, 25, 904–912. [Google Scholar] [CrossRef]
- Wilson, B.D.; Ii, M.; Park, K.W.; Suli, A.; Sorensen, L.K.; Larrieu-Lahargue, F.; Urness, L.D.; Suh, W.; Asai, J.; Kock, G.A. Netrins promote developmental and therapeutic angiogenesis. Science 2006, 313, 640–644. [Google Scholar] [CrossRef]
- Page, L.M. Zebrafish as developmental models. Science 1990, 250, 1320. [Google Scholar] [CrossRef] [PubMed]
- Roush, W. Zebrafish embryology builds better model vertebrate. Science 1996, 272, 1103. [Google Scholar] [CrossRef] [PubMed]
- Lele, Z.; Krone, P. The zebrafish as a model system in developmental, toxicological and transgenic research. Biotechnol. Adv. 1996, 14, 57–72. [Google Scholar] [CrossRef]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef]
- Sipes, N.S.; Padilla, S.; Knudsen, T.B. Zebrafish—As an integrative model for twenty-first century toxicity testing. Birth Defects Res. Part C Embryo Today Rev. 2011, 93, 256–267. [Google Scholar] [CrossRef]
- Pei, D.-S.; Strauss, P.R. Zebrafish as a model system to study DNA damage and repair. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2013, 743, 151–159. [Google Scholar] [CrossRef]
- Pei, D.-S.; Yang, X.-J.; Liu, W.; Guikema, J.E.; Schrader, C.E.; Strauss, P.R. A novel regulatory circuit in base excision repair involving AP endonuclease 1, Creb1 and DNA polymerase β. Nucleic Acids Res. 2011, 39, 3156–3165. [Google Scholar] [CrossRef]
- Konantz, M.; Balci, T.B.; Hartwig, U.F.; Dellaire, G.; André, M.C.; Berman, J.N.; Lengerke, C. Zebrafish xenografts as a tool for in vivo studies on human cancer. Ann. N. Y. Acad. Sci. 2012, 1266, 124–137. [Google Scholar] [CrossRef]
- Berger, J.; Currie, P.D. Zebrafish models flex their muscles to shed light on muscular dystrophies. Dis. Models Mech. 2012, 5, 726–732. [Google Scholar] [CrossRef]
- Li, Y.-j.; Hu, B. Establishment of multi-site infection model in zebrafish larvae for studying Staphylococcus aureus infectious disease. J. Genet. Genom. 2012, 39, 521–534. [Google Scholar] [CrossRef]
- Goolish, E.M.; Okutake, K.; Johnson, P. The behavioral response of zebrafish to hypergravity conditions. J. Gravit. Physiol. A J. Int. Soc. Gravit. Physiol. 2000, 7, P99–P100. [Google Scholar]
- MIZEll, M.; Romig, E. The aquatic vertebrate embryo as a sentinel for toxins: Zebrafish Embryo Dechorionation and Perivitelline Space Microinjection. Int. J. Dev. Biol. 2002, 41, 411–423. [Google Scholar]
- Crosby, E.B.; Bailey, J.M.; Oliveri, A.N.; Levin, E.D. Neurobehavioral impairments caused by developmental imidacloprid exposure in zebrafish. Neurotoxicology Teratol. 2015, 49, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Tišler, T.; Jemec, A.; Mozetič, B.; Trebše, P. Hazard identification of imidacloprid to aquatic environment. Chemosphere 2009, 76, 907–914. [Google Scholar] [CrossRef]
- Scheil, V.; Köhler, H.-R. Influence of nickel chloride, chlorpyrifos, and imidacloprid in combination with different temperatures on the embryogenesis of the zebrafish Danio rerio. Arch. Environ. Contam. Toxicol. 2009, 56, 238–243. [Google Scholar] [CrossRef]
- Brugman, S. The zebrafish as a model to study intestinal inflammation. Dev. Comp. Immunol. 2016, 64, 82–92. [Google Scholar] [CrossRef]
- Di Paola, D.; Capparucci, F.; Lanteri, G.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Cuzzocrea, S. Combined Toxicity of Xenobiotics Bisphenol A and Heavy Metals on Zebrafish Embryos (Danio rerio). Toxics 2021, 9, 344. [Google Scholar] [CrossRef]
- Di Paola, D.; Capparucci, F.; Abbate, J.M.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Genovese, T.; Impellizzeri, D. Environmental Risk Assessment of Oxaliplatin Exposure on Early Life Stages of Zebrafish (Danio rerio). Toxics 2022, 10, 81. [Google Scholar] [CrossRef]
- Di Paola, D.; Natale, S.; Gugliandolo, E.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Cuzzocrea, S. Assessment of 2-Pentadecyl-2-oxazoline Role on Lipopolysaccharide-Induced Inflammation on Early Stage Development of Zebrafish (Danio rerio). Life 2022, 12, 128. [Google Scholar] [CrossRef]
- Di Paola, D.; Natale, S.; Iaria, C.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Cuzzocrea, S. Intestinal Disorder in Zebrafish Larvae (Danio rerio): The Protective Action of N-Palmitoylethanolamide-oxazoline. Life 2022, 12, 125. [Google Scholar] [CrossRef]
- Di Paola, D.; Iaria, C.; Lanteri, G.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Cuzzocrea, S. Sensitivity of Zebrafish Embryogenesis to Risk of Fotemustine Exposure. Fishes 2022, 7, 67. [Google Scholar] [CrossRef]
- Buschmann, J. The OECD guidelines for the testing of chemicals and pesticides. Methods Mol. Biol 2013, 947, 37–56. [Google Scholar] [CrossRef] [PubMed]
- Parenti, C.C.; Ghilardi, A.; Della Torre, C.; Magni, S.; Del Giacco, L.; Binelli, A. Evaluation of the infiltration of polystyrene nanobeads in zebrafish embryo tissues after short-term exposure and the related biochemical and behavioural effects. Environ. Pollut. 2019, 254, 112947. [Google Scholar] [CrossRef] [PubMed]
- Kuder, R.S.; Gundala, H.P. Developmental toxicity of deltamethrin and 3-phenoxybenzoic acid in embryo-larval stages of zebrafish (Danio rerio). Toxicol. Mech. Methods 2018, 28, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.; Liu, K.; He, Q.; Sun, C.; Han, J.; Han, L.; Tian, Q. Xiaoaiping induces developmental toxicity in zebrafish embryos through activation of ER stress, apoptosis and the Wnt pathway. Front. Pharmacol. 2018, 9, 1250. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, E.; Cordaro, M.; Fusco, R.; Peritore, A.F.; Siracusa, R.; Genovese, T.; D’Amico, R.; Impellizzeri, D.; Di Paola, R.; Cuzzocrea, S. Protective effect of snail secretion filtrate against ethanol-induced gastric ulcer in mice. Sci Rep. 2021, 11, 3638. [Google Scholar] [CrossRef]
- Cordaro, M.; Siracusa, R.; Fusco, R.; D’Amico, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; Scuto, M.; Crupi, R.; Mandalari, G. Cashew (Anacardium occidentale L.) nuts counteract oxidative stress and inflammation in an acute experimental model of Carrageenan-induced Paw edema. Antioxidants 2020, 9, 660. [Google Scholar] [CrossRef]
- Jin, Y.; Chen, R.; Liu, W.; Fu, Z. Effect of endocrine disrupting chemicals on the transcription of genes related to the innate immune system in the early developmental stage of zebrafish (Danio rerio). Fish Shellfish. Immun. 2010, 28, 854–861. [Google Scholar] [CrossRef]
- Varela, M.; Dios, S.; Novoa, B.; Figueras, A. Characterisation, expression and ontogeny of interleukin-6 and its receptors in zebrafish (Danio rerio). Dev. Comp. Immunol. 2012, 37, 97–106. [Google Scholar] [CrossRef]
- Zhang, Y.; Takagi, N.; Yuan, B.; Zhou, Y.; Si, N.; Wang, H.; Yang, J.; Wei, X.; Zhao, H.; Bian, B. The protection of indolealkylamines from LPS-induced inflammation in zebrafish. J. Ethnopharmacol. 2019, 243, 112122. [Google Scholar] [CrossRef]
- Petrosino, S.; Schiano Moriello, A.; Verde, R.; Allarà, M.; Imperatore, R.; Ligresti, A.; Mahmoud, A.M.; Peritore, A.F.; Iannotti, F.A.; Di Marzo, V. Palmitoylethanolamide counteracts substance P-induced mast cell activation in vitro by stimulating diacylglycerol lipase activity. J. Neuroinflammation 2019, 16, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Steenbergen, P.J.; Bardine, N. Antinociceptive effects of buprenorphine in zebrafish larvae: An Alternative for Rodent Models to Study Pain and Nociception? Appl. Anim. Behav. Sci. 2014, 152, 92–99. [Google Scholar] [CrossRef]
- Fusco, R.; Gugliandolo, E.; Siracusa, R.; Scuto, M.; Cordaro, M.; D’Amico, R.; Evangelista, M.; Peli, A.; Peritore, A.F.; Impellizzeri, D. Formyl peptide receptor 1 signaling in acute inflammation and neural differentiation induced by traumatic brain injury. Biology 2020, 9, 238. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.F.; Gil, F.; Lacasaña, M. Toxicological interactions of pesticide mixtures: An Update. Arch. Toxicol. 2017, 91, 3211–3223. [Google Scholar] [CrossRef]
- González-Alzaga, B.; Lacasaña, M.; Aguilar-Garduño, C.; Rodríguez-Barranco, M.; Ballester, F.; Rebagliato, M.; Hernández, A. A systematic review of neurodevelopmental effects of prenatal and postnatal organophosphate pesticide exposure. Toxicol. Lett. 2014, 230, 104–121. [Google Scholar] [CrossRef]
- Mokarizadeh, A.; Faryabi, M.R.; Rezvanfar, M.A.; Abdollahi, M. A comprehensive review of pesticides and the immune dysregulation: Mechanisms, Evidence and Consequences. Toxicol. Mech. Methods 2015, 25, 258–278. [Google Scholar] [CrossRef]
- Ntzani, E.E.; Ntritsos, G.C.M.; Evangelou, E.; Tzoulaki, I. Literature review on epidemiological studies linking exposure to pesticides and health effects. EFSA Supporting Publ. 2013, 10, 497E. [Google Scholar] [CrossRef]
- Parrón, T.; Requena, M.; Hernández, A.F.; Alarcón, R. Association between environmental exposure to pesticides and neurodegenerative diseases. Toxicol. Appl. Pharmacol. 2011, 256, 379–385. [Google Scholar] [CrossRef]
- Sreenivasa Rao, A.; Pillala, R.R. The concentration of pesticides in sediments from Kolleru Lake in India. Pest. Manag. Sci. Former. Pestic. Sci. 2001, 57, 620–624. [Google Scholar] [CrossRef]
- Singh, S.K.; Singh, S.K.; Yadav, R.P. Toxicological and biochemical alterations of Cypermethrin (synthetic pyrethroids) against freshwater teleost Fish Colisa fasciatus at different season. World J. Zool. 2010, 5, 25–32. [Google Scholar]
- Kosygin, L.; Dhamendra, H.; Gyaneshwari, R. Pollution status and conservation strategies of Moirang river, Manipur with a note on its aquatic bio-resources. J. Environ. Biol. 2007, 28, 669–673. [Google Scholar] [PubMed]
- Murthy, K.S.; Kiran, B.; Venkateshwarlu, M. A review on toxicity of pesticides in Fish. Int. J. Open Sci. Res. 2013, 1, 15–36. [Google Scholar]
- Wang, S.; Hong, H.; Wang, X. Bioenergetic responses in green lipped mussels (Perna viridis) as indicators of pollution stress in Xiamen coastal waters, China. Mar. Pollut. Bull. 2005, 51, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Trumpolt, C.W.; Crain, M.; Cullison, G.D.; Flanagan, S.J.; Siegel, L.; Lathrop, S. Perchlorate: Sources, Uses, and Occurrences in the Environment. Remediat. J. Environ. Cleanup Costs Technol. Tech. 2005, 16, 65–89. [Google Scholar] [CrossRef]
- Esposito, E.; Campolo, M.; Casili, G.; Lanza, M.; Franco, D.; Filippone, A.; Peritore, A.F.; Cuzzocrea, S. Protective effects of xyloglucan in association with the polysaccharide gelose in an experimental model of gastroenteritis and urinary tract infections. Int. J. Mol. Sci. 2018, 19, 1844. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, G.; Dai, D.; Xu, Z.; Cai, L.; Wang, Q.; Yu, Y. Individual and mixture effects of five agricultural pesticides on zebrafish (Danio rerio) larvae. Environ. Sci. Pollut. Res. 2017, 24, 4528–4536. [Google Scholar] [CrossRef]
- Jin, C.; Luo, T.; Zhu, Z.; Pan, Z.; Yang, J.; Wang, W.; Fu, Z.; Jin, Y. Imazalil exposure induces gut microbiota dysbiosis and hepatic metabolism disorder in zebrafish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 202, 85–93. [Google Scholar] [CrossRef]
- Jin, Y.; Xia, J.; Pan, Z.; Yang, J.; Wang, W.; Fu, Z. Polystyrene microplastics induce microbiota dysbiosis and inflammation in the gut of adult zebrafish. Environ. Pollut. 2018, 235, 322–329. [Google Scholar] [CrossRef]
- Graziani, C.; Talocco, C.; De Sire, R.; Petito, V.; Lopetuso, L.; Gervasoni, J.; Persichilli, S.; Franceschi, F.; Ojetti, V.; Gasbarrini, A. Intestinal permeability in physiological and pathological conditions: Major Determinants and Assessment Modalities. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 795–810. [Google Scholar]
- Çavaş, T.; Çinkılıç, N.; Vatan, Ö.; Yılmaz, D.; Coşkun, M. In vitro genotoxicity evaluation of acetamiprid in CaCo-2 cells using the micronucleus, comet and γH2AX foci assays. Pestic. Biochem. Physiol. 2012, 104, 212–217. [Google Scholar] [CrossRef]
- Brunet, J.-L.; Maresca, M.; Fantini, J.; Belzunces, L.P. Human intestinal absorption of imidacloprid with Caco-2 cells as enterocyte model. Toxicol. Appl. Pharmacol. 2004, 194, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.; Kolosov, D.; O’Donnell, M.J.; Erlandson, M.A.; McNeil, J.N.; Donly, C. The effect of diet on midgut and resulting changes in infectiousness of AcMNPV baculovirus in Trichoplusia ni. Front. Physiol. 2018, 9, 1348. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Furuse, M.; Fujimoto, K.; Tsukita, S. Claudin multigene family encoding four-transmembrane domain protein components of tight junction strands. Proc. Natl. Acad. Sci. USA 1999, 96, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; van Passel, M.W.; van de Bovenkamp, J.H.; Schipper, R.; de Vos, W.; Dekker, J. Mucin-bacterial interactions in the human oral cavity and digestive tract. Gut Microbes 2010, 1, 254–268. [Google Scholar] [CrossRef]
- Yang, G.; Yuan, X.; Jin, C.; Wang, D.; Wang, Y.; Miao, W.; Jin, Y. Imidacloprid disturbed the gut barrier function and interfered with bile acids metabolism in mice. Environ. Pollut. 2020, 266, 115290. [Google Scholar] [CrossRef]
- Zhou, L.; Limbu, S.M.; Shen, M.; Zhai, W.; Qiao, F.; He, A.; Du, Z.-Y.; Zhang, M. Environmental concentrations of antibiotics impair zebrafish gut health. Environ. Pollut. 2018, 235, 245–254. [Google Scholar] [CrossRef]
- Wang, X.; Shen, M.; Zhou, J.; Jin, Y. Chlorpyrifos disturbs hepatic metabolism associated with oxidative stress and gut microbiota dysbiosis in adult zebrafish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 216, 19–28. [Google Scholar] [CrossRef]
- Ge, W.; Yan, S.; Wang, J.; Zhu, L.; Chen, A.; Wang, J. Oxidative stress and DNA damage induced by imidacloprid in zebrafish (Danio rerio). J. Agric. Food Chem. 2015, 63, 1856–1862. [Google Scholar] [CrossRef]
- Vieira, C.E.D.; Pérez, M.R.; Acayaba, R.D.A.; Raimundo, C.C.M.; dos Reis Martinez, C.B. DNA damage and oxidative stress induced by imidacloprid exposure in different tissues of the Neotropical fish Prochilodus lineatus. Chemosphere 2018, 195, 125–134. [Google Scholar] [CrossRef]
- Özdemir, S.; Altun, S.; Arslan, H. Imidacloprid exposure cause the histopathological changes, activation of TNF-α, iNOS, 8-OHdG biomarkers, and alteration of caspase 3, iNOS, CYP1A, MT1 gene expression levels in common carp (Cyprinus carpio L.). Toxicol. Rep. 2018, 5, 125–133. [Google Scholar] [CrossRef]
- Dong, M.; Zhu, L.; Shao, B.; Zhu, S.; Wang, J.; Xie, H.; Wang, J.; Wang, F. The effects of endosulfan on cytochrome P450 enzymes and glutathione S-transferases in zebrafish (Danio rerio) livers. Ecotoxicol. Environ. Saf. 2013, 92, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Mu, X.; Wang, K.; Chai, T.; Yang, Y.; Qiu, L.; Wang, C. Cyhalofop-butyl has the potential to induce developmental toxicity, oxidative stress and apoptosis in early life stage of zebrafish (Danio rerio). Environ. Pollut. 2015, 203, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, W.; Hua, J.; Hu, C.; Lok-Shun Lai, N.; Qian, P.-Y.; Lam, P.K.; Lam, J.C.; Zhou, B. Dysregulation of intestinal health by environmental pollutants: Involvement of the Estrogen Receptor and Aryl Hydrocarbon Receptor. Environ. Sci. Technol. 2018, 52, 2323–2330. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yu, K.; Shi, X.; Wang, J.; Lam, P.K.; Wu, R.S.; Zhou, B. Induction of oxidative stress and apoptosis by PFOS and PFOA in primary cultured hepatocytes of freshwater tilapia (Oreochromis niloticus). Aquat. Toxicol. 2007, 82, 135–143. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Orientation | Nucleotide Sequence |
|---|---|---|
| b-actin | forward | 5′- CCATCGGCAATGAGCGTTTC -3′ |
| reverse | 5′- CAAGATTCCATACCCAGGAAGGA -3′ | |
| Muc2.1 | forward | 5′- CAACATCGATGGCTGCTTCTG -3′ |
| reverse | 5′- CTGACAGTAACATTCTTCCTCGC -3′ | |
| Muc.2.2 | forward | 5′- ACACGCTCAAGTAATCGCACAGTC -3′ |
| reverse | 5′- TCAGCGAGTGTTTGGCTCACTT -3′ | |
| Ocln | forward | 5′- CAAAATCAGGCAAAGGCTTC -3′ |
| reverse | 5′- AACAATAGTGGCGATGAGCA -3′ | |
| Caln | forward | 5′- GTACCCTCCGCAAAGTCGTA -3′ |
| reverse | 5′- CTTTCAAGGAAAGACTGACAGC -3′ | |
| sod1 | forward | 5′- GGCCAACCGATAGTGTTAGA -3′ |
| reverse | 5′- CCAGCGTTGCCAGTTTTTAG -3′ | |
| cat | forward | 5′- AGGGCAACTGGGATCTTACA -3′ |
| reverse | 5′- TTTATGGGACCAGACCTTGG -3′ | |
| gstp2 | forward | 5′- CACAGACCTCGCTTTTCACAC -3′ |
| reverse | 5′- GAGAGAAGCCTCACAGTCGT -3′ |
| Survival | ||||
|---|---|---|---|---|
| 24 h | 48 h | 72 h | 96 h | |
| CTRL | 100 ± 0 | 100 ± 0 | 100 ± 0 | 100 ± 0 |
| IMI 75 mg/L | 100 ± 0.57 | 100 ± 0.57 | 100 ± 0.57 | 100 ± 0.57 |
| IMI 100 mg/L | 100 ± 0.57 | 100 ± 0.57 | 100 ± 0.57 | 100 ± 0.57 |
| IMI 150 mg/L | 99.67 ± 0.33 | 95.00 ± 2.08 | 82.33 ± 1.20 | 51.67 ± 1.76 |
| KClO4 1 mM | 100 ± 0.57 | 100 ± 0.57 | 100 ± 0.57 | 98 ± 2 |
| KClO4 1.5 mM | 99.67 ± 0.33 | 97.00 ± 1.15 * | 83.67 ± 2.02 *** | 48.00 ± 2.08 *** |
| KClO4 5 mM | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paola, D.D.; Capparucci, F.; Natale, S.; Crupi, R.; Cuzzocrea, S.; Spanò, N.; Gugliandolo, E.; Peritore, A.F. RETRACTED: Combined Effects of Potassium Perchlorate and a Neonicotinoid on Zebrafish Larvae (Danio rerio). Toxics 2022, 10, 203. https://doi.org/10.3390/toxics10050203
Paola DD, Capparucci F, Natale S, Crupi R, Cuzzocrea S, Spanò N, Gugliandolo E, Peritore AF. RETRACTED: Combined Effects of Potassium Perchlorate and a Neonicotinoid on Zebrafish Larvae (Danio rerio). Toxics. 2022; 10(5):203. https://doi.org/10.3390/toxics10050203
Chicago/Turabian StylePaola, Davide Di, Fabiano Capparucci, Sabrina Natale, Rosalia Crupi, Salvatore Cuzzocrea, Nunziacarla Spanò, Enrico Gugliandolo, and Alessio Filippo Peritore. 2022. "RETRACTED: Combined Effects of Potassium Perchlorate and a Neonicotinoid on Zebrafish Larvae (Danio rerio)" Toxics 10, no. 5: 203. https://doi.org/10.3390/toxics10050203
APA StylePaola, D. D., Capparucci, F., Natale, S., Crupi, R., Cuzzocrea, S., Spanò, N., Gugliandolo, E., & Peritore, A. F. (2022). RETRACTED: Combined Effects of Potassium Perchlorate and a Neonicotinoid on Zebrafish Larvae (Danio rerio). Toxics, 10(5), 203. https://doi.org/10.3390/toxics10050203