
Assessing the Effect of Modified Clay on the Toxicity of Karenia mikimotoi Using Marine Medaka (Oryzias melastigma) as a Model Organism

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Organisms Culture and MC Preparation
2.2. Experimental Design
2.2.1. Effect of K. mikimotoi on the Marine Medaka
2.2.2. Determination of Hemolytic Toxicity of K. mikimotoi
2.2.3. Effect of MC on the Toxicity of K. mikimotoi
2.3. Histological Morphology Examination
2.4. Measurement of the Biochemical and Physiological Indexes
2.5. RNA Extraction, Library Construction and RNA Sequencing (RNA-Seq)
2.6. Functional Annotation of Genes
2.7. Quantitative Real-Time PCR
2.8. Statistical Analysis
3. Results
3.1. Effects of MC on K. mikimotoi
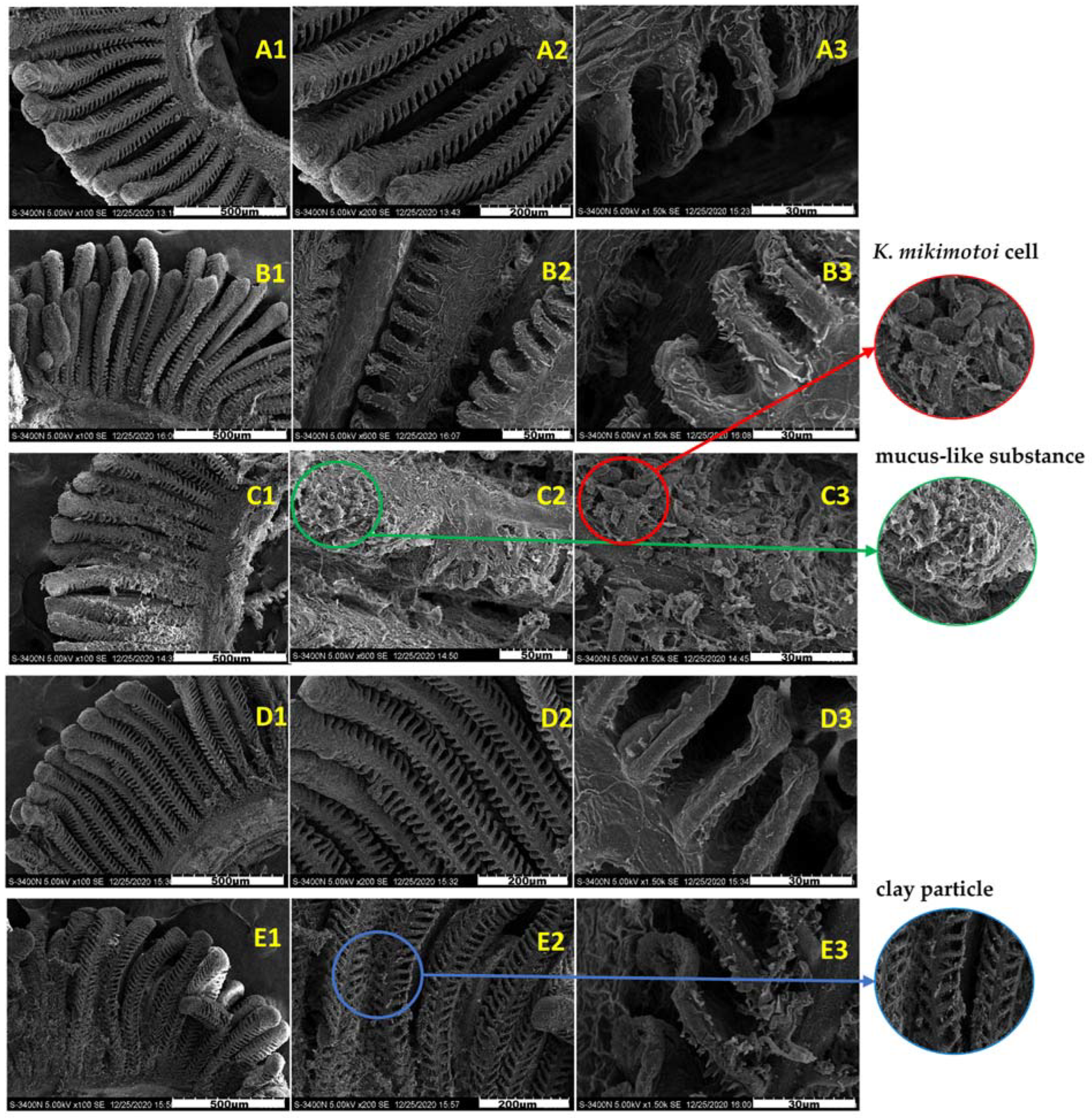
3.1.1. Effects of MC on K. mikimotoi Cell Morphology
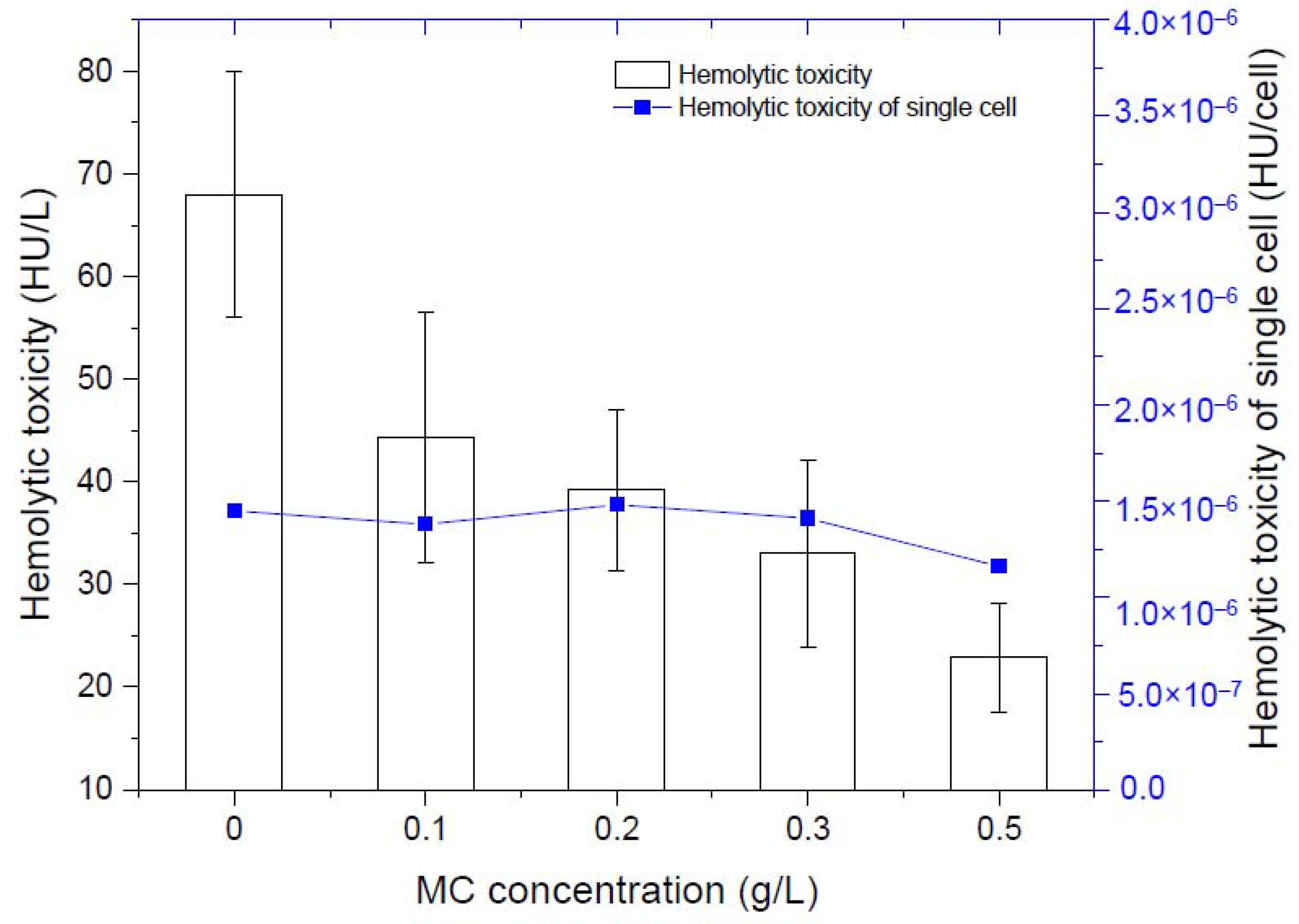
3.1.2. Effect of MC on the Hemolytic Toxicity of K. mikimotoi
3.2. Marine Medaka Survival
3.3. Fish Gill Surface Morphology
3.4. Changes in the Gill Biochemical and Physiological Indexes
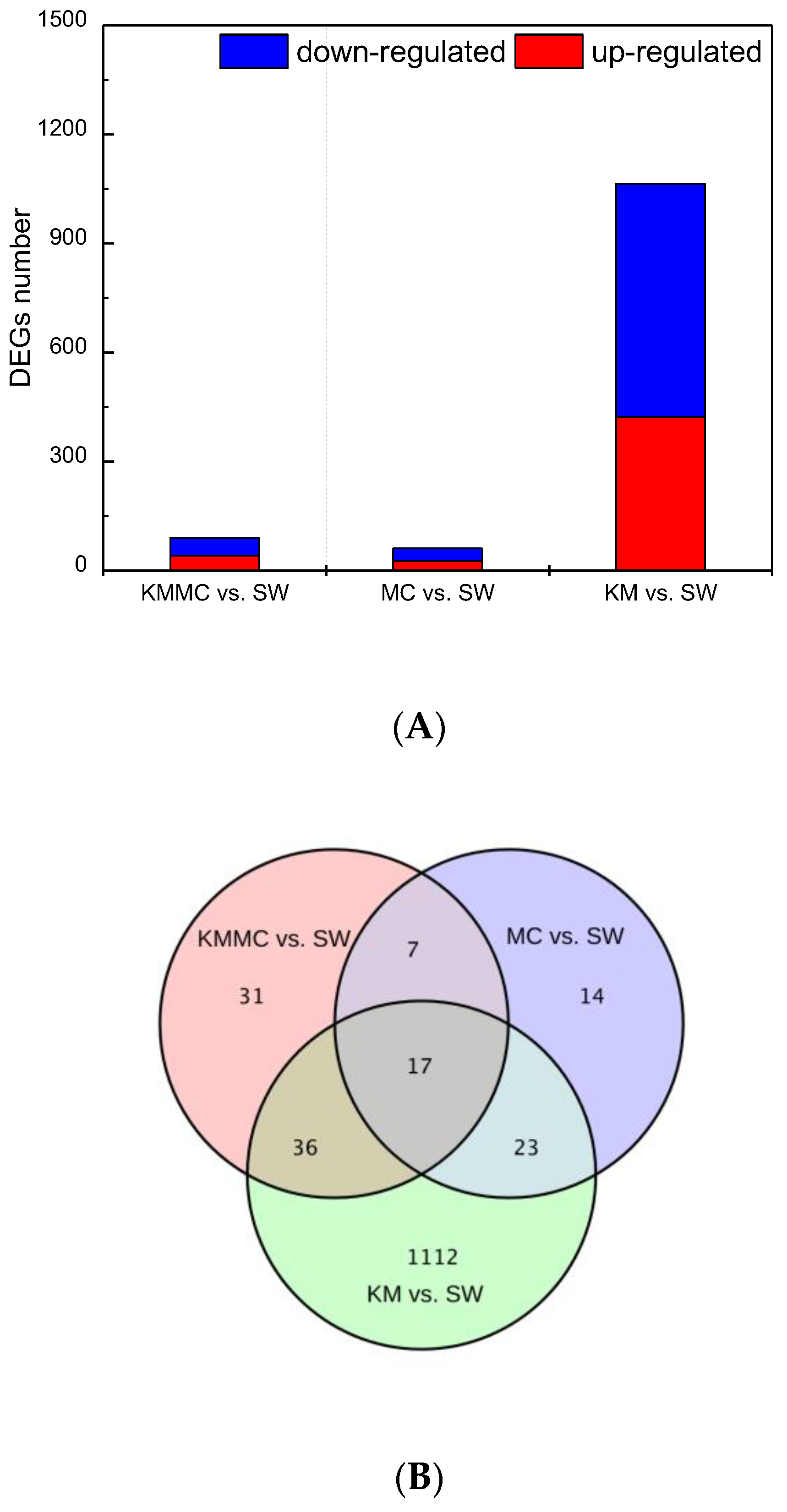
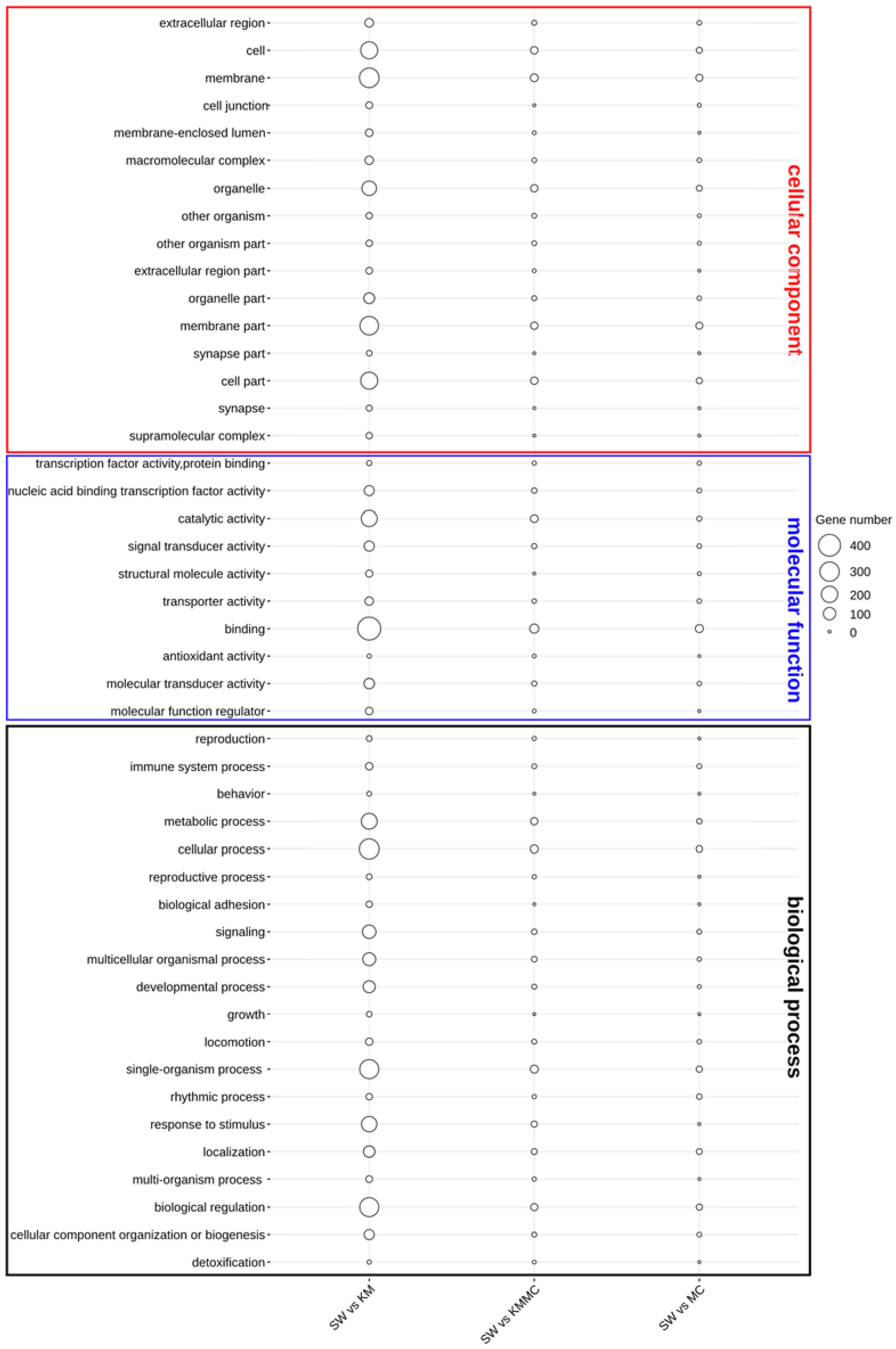
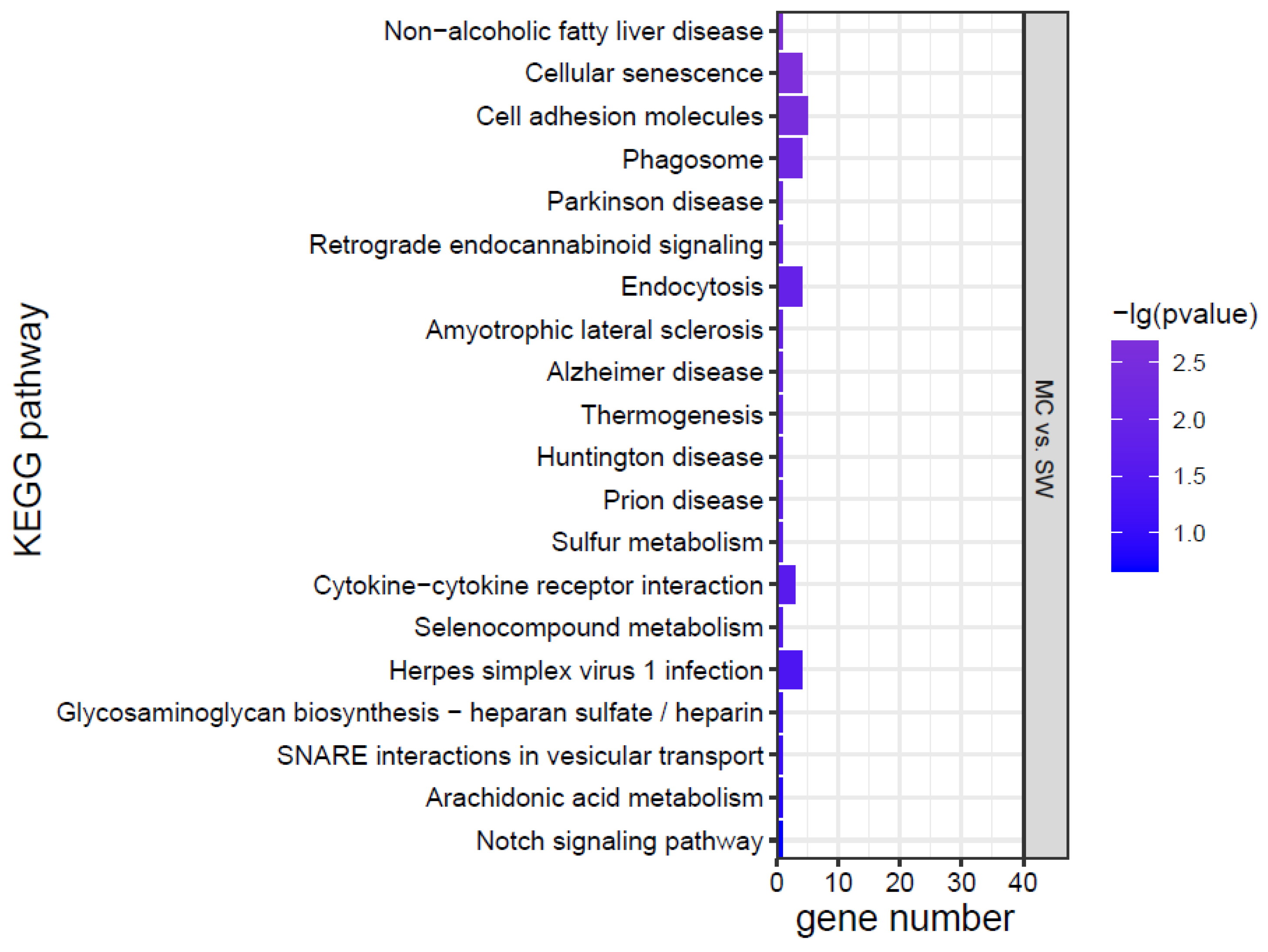
3.5. Gene Expression
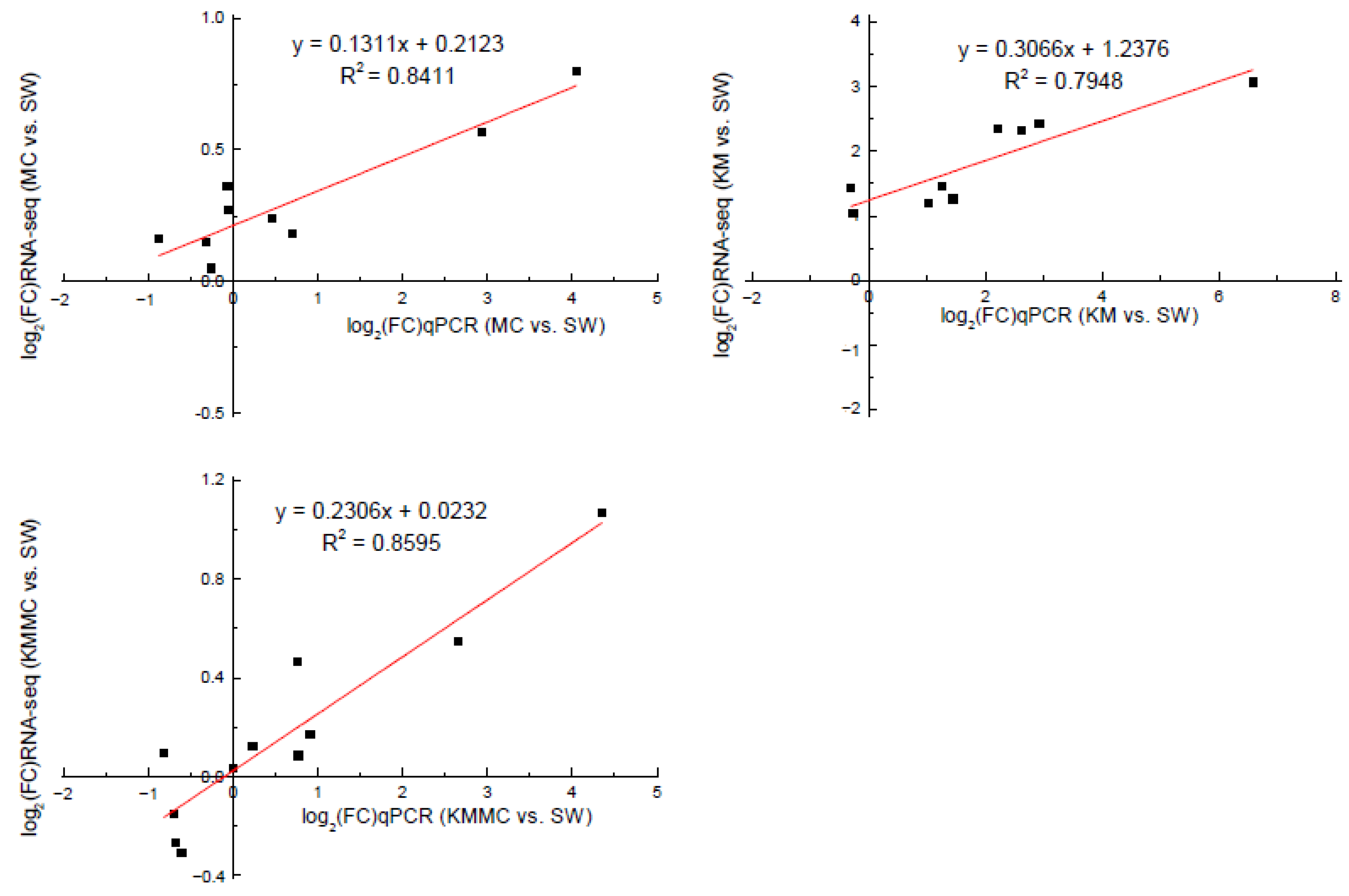
3.6. Data Validation by qRT-PCR
4. Discussion
4.1. Marine Medaka Survival
4.2. The External Morphology and Osmotic Adjustment of Gills
4.3. Oxidative Stress in Fish Gills
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, X.; Yan, T.; Yu, R.; Zhou, M. A review of karenia mikimotoi: Bloom events, physiology, toxicity and toxic mechanism. Harmful Algae 2019, 90, 101702. [Google Scholar] [CrossRef]
- Mitchell, S.; Rodger, H. Pathology of wild and cultured fish affected by a Karenia mikimotoi bloom in Ireland, 2005. Bull. Eur. Assoc. Fish Pathol. 2007, 27, 39–42. [Google Scholar]
- Davidson, K.; Miller, P.; Wilding, T.A.; Shutler, J.; Bresnan, E.; Kennington, K.; Swan, S. A large and prolonged bloom of Karenia mikimotoi in Scottish waters in 2006. Harmful Algae 2009, 8, 349–361. [Google Scholar] [CrossRef]
- Shimada, H.; Kanamori, M.; Yoshida, H.; Imai, I. First record of red tide due to the harmful dinoflagellate Karenia mikimotoi in Hakodate Bay, southern Hokkaido, in autumn 2015. Nippon Suisan Gakkaishi 2016, 82, 934–938. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yan, T.; Lin, J.; Yu, R.; Zhou, M. Detrimental impacts of the dinoflagellate Karenia mikimotoi in Fujian coastal waters on typical marine organisms. Harmful Algae 2017, 61, 1–12. [Google Scholar] [CrossRef]
- Lin, J.; Yan, T.; Zhang, Q.; Wang, Y.; Liu, Q.; Zhou, M. Effects of Karenia mikimotoi blooms on antioxidant enzymes in gastropod abalone, Haliotis discus hannai. Mar. Sci. 2016, 40, 17–22. [Google Scholar]
- Zhang, T.; Yan, T.; Zhang, Q.; Li, X.; Zhou, M. Acute effect of four typical bloom forming algae on abalone Haliotis discus hannai and its antioxidant enzymes system. Mar. Environ. Sci. 2018, 37, 207–214. [Google Scholar]
- Mooney, B.D.; de Salas, M.; Hallegraeff, G.M.; Place, A.R. Survey for karlotoxin production in 15 species of gymnodinioid dinoflagellates (Kareniaceae, Dinophyta). J. Phycol. 2009, 45, 164–175. [Google Scholar] [CrossRef]
- Brand, L.E.; Campbell, L.; Bresnan, E. Karenia: The biology and ecology of a toxic genus. Harmful Algae 2012, 14, 156–178. [Google Scholar] [CrossRef]
- Shi, F.; McNabb, P.; Rhodes, L.; Holland, P.; Webb, S.; Adamson, J.; Immers, A.; Gooneratne, R.; Holland, J. The toxic effects of three dinoflagellate species from the genus Karenia on invertebrate larvae and finfish. N. Z. J. Mar. Freshw. Res. 2012, 46, 149–165. [Google Scholar] [CrossRef]
- Neely, T.; Campbell, L. A modified assay to determine hemolytic toxin variability among Karenia clones isolated from the Gulf of Mexico. Harmful Algae 2006, 5, 592–598. [Google Scholar] [CrossRef]
- Zou, Y.; Yamasaki, Y.; Matsuyama, Y.; Yamaguchi, K.; Honjo, T.; Oda, T. Possible involvement of hemolytic activity in the contact-dependent lethal effects of the dinoflagellate Karenia mikimotoi on the rotifer Brachionus plicatilis. Harmful Algae 2010, 9, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Satake, M.; Shoji, M.; Oshima, Y.; Naoki, H.; Yasumoto, T. Gymnocin-A, a cytotoxic polyether from the notorious red tide dinoflagellate, Gymnodinium mikimotoi. Cheminform 2002, 43, 5829–5832. [Google Scholar] [CrossRef]
- Satake, M.; Tanaka, Y.; Ishikura, Y.; Oshima, Y.; Yasumoto, T. Gymnocin-B with the largest contiguous polyether rings from the red tide dinoflagellate, Karenia (formerly Gymnodinium) mikimotoi. Tetrahedron Lett. 2005, 46, 3537–3540. [Google Scholar] [CrossRef]
- Mooney, B.D.; Nichols, P.D.; Salas, M.; Hallegraeff, G.M. Lipid, fatty acid, and sterol composition of eight species of Kareniaceae (Dinophyta): Chemotaxonomy and putative lipid phycotoxins. J. Phycol. 2007, 43, 101–111. [Google Scholar] [CrossRef]
- Kong, R.Y.C.; Giesy, J.P.; Wu, R.S.S.; Chen, E.X.H.; Chiang, M.W.L.; Lim, P.L.; Yuen, B.B.H.; Yip, B.W.P.; Mok, H.O.L.; Au, D.W.T. Development of a marine fish model for studying in vivo molecular responses in ecotoxicology. Aquat. Toxicol. 2008, 86, 131–141. [Google Scholar] [CrossRef]
- Dong, S.; Kang, M.; Wu, X.; Ye, T. Development of a Promising Fish Model (Oryzias melastigma) for Assessing Multiple Responses to Stresses in the Marine Environment. Biomed Res. Int. 2014, 2014, 563131. [Google Scholar] [CrossRef] [Green Version]
- Kostic, J.; Kolarevic, S.; Kracun-Kolarevic, M.; Aborgiba, M.; Gacic, Z.; Paunovic, M.; Visnjic-Jeftic, Z.; Raskovic, B.; Poleksic, V.; Lenhardt, M.; et al. The impact of multiple stressors on the biomarkers response in gills and liver of freshwater breams during different seasons. Sci. Total Environ. 2017, 601, 1670–1681. [Google Scholar] [CrossRef]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef]
- Jones, K.; Ayres, P.; Bullock, A.; Roberts, R.; Tett, P. A Red Tide of Gyrod Inium Aurelum in Sea Lochs of the Firth of Clyde and Associated Mortality of Pond-Reared Salmon. J. Mar. Biol. Assoc. U. K. 1982, 62, 771–782. [Google Scholar] [CrossRef]
- Sola, F.; Masoni, A.; Fossat, B.; Porthe-Nibelle, J.; Gentien, P.; Bodennec, G. Toxicity of fatty acid 18: 5n3 from Gymnodinium cf. mikimotoi: I. Morphological and biochemical aspects on Dicentrarchus labrax gills and intestine. J. Appl. Toxicol. 1999, 19, 279–284. [Google Scholar] [CrossRef]
- Oda, M. The red tide of Gymnodinium mikimotoi Miyake et Kominami n. sp. (MS) and the influence of copper sulfate on the red tide. Zool. Mag. 2021, 47, 35–48. [Google Scholar]
- Anderson, D.M.; Andersen, P.; Bricelj, V.M.; Cullen, J.J.; Rensel, J. Monitoring and Management Strategies for Harmful Algal Blooms in Coastal Waters; UNESCO: Paris, France, 2001. [Google Scholar]
- Song, X.; Zhang, Y.; Yu, Z. An eco-environmental assessment of harmful algal bloom mitigation using modified clay. Harmful Algae 2021, 107, 102067. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Song, X.; Li, J.; Zhang, Y.; Shen, H.; Zhang, P.; Yu, Z. Toxin remained in residual Alexandrium Pacificum after flocculation with modified clay. Oceanol. Limnol. Sin. 2021, 52, 917–924. [Google Scholar]
- Liu, S.; Yu, Z.; Song, X.; Cao, X. Physiological and photosynthetic responses of Karenia mikimotoi to the modified clay mitigation method. Mar. Pollut. Bull. 2018, 133, 491–499. [Google Scholar] [CrossRef]
- Zhu, J.; Yu, Z.; He, L.; Cao, X.; Liu, S.; Song, X. Molecular mechanism of modified clay controlling the brown tide organism, Aureococcus anophagefferens revealed by transcriptome analysis. Environ. Sci. Technol. 2018, 52, 7006–7014. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, X.; Yu, Z.; Zhang, P.; Cao, X.; Yuan, Y. Impact assessment of modifi ed clay on embryo-larval stages of turbot Scophthalmus maximus L. Chin. J. Oceanol. Limnol. 2019, 37, 1051–1061. [Google Scholar] [CrossRef]
- Lee, Y.C.; Jin, E.S.; Jung, S.W.; Kim, Y.M.; Chang, K.S.; Yang, J.W.; Kim, S.W.; Kim, Y.O.; Shin, H.J. Utilizing the algicidal activity of aminoclay as a practical treatment for toxic red tides. Sci. Rep. 2013, 3, 1292. [Google Scholar] [CrossRef] [Green Version]
- Orizar, I.S.; Rivera, P.P.L.; San Diego-McGlone, M.L.; Azanza, R.V. Harmful Algal Bloom (HAB) mitigation using ball clay: Effect on non-target organisms. J. Environ. Sci. Manag. 2013, 16, 36–43. [Google Scholar]
- Zhang, P.; Song, X.; Li, J.; Yu, Z. Effects of modified clay on Atlantic salmon (Salmo Salar). Oceanol. Limnol. Sin. 2019, 50, 216–223. [Google Scholar]
- Zhang, Y.; Song, X.; Shen, H.; Cao, X.; Yuan, Y.; Wu, Z.; Yu, Z. The Effects of Modified Clay on Abalone (Haliotis discus hannai) Based on Laboratory and Field Experiments. Environ. Toxicol. Chem. 2020, 39, 2065–2075. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yu, Z.; Song, X.; Xihua, C.; Liu, K. Impact of modified clay on the growth of the infant Apostichopus Japonicas Selenka in HABs controling. Oceanol. Limnol. Sin. 2014, 45, 233–238. [Google Scholar]
- Gordon, D.C. The influence of dilute clay suspensions on sea scallop (Placopecten magellanicus) feeding activity and tissue growth. Neth. J. Sea Res. 1992, 30, 107–120. [Google Scholar]
- Shumway, S.E.; Frank, D.M.; Ewart, L.M.; Evan Ward, J. Effect of yellow loess on clearance rate in seven species of benthic, filter-feeding invertebrates. Aquac. Res. 2003, 34, 1391–1402. [Google Scholar] [CrossRef]
- Pierce, R.H.; Henry, M.S.; Higham, C.J.; Blum, P.; Sengco, M.R.; Anderson, D.M. Removal of harmful algal cells (Karenia brevis) and toxins from seawater culture by clay flocculation. Harmful Algae 2004, 3, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Sengco, M.R.; Hagström, J.A.; Granéli, E.; Anderson, D.M. Removal of Prymnesium parvum (Haptophyceae) and its toxins using clay minerals. Harmful Algae 2005, 4, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Morri, R.J.; Williams, D.E.; Luu, H.A.; Holmes, C.; Andersen, R.J.; Calvert, S.E. The adsorption of microcystin-LR by natural clay particles. Toxicon 2000, 38, 303–308. [Google Scholar] [CrossRef]
- Yu, Z.; Zou, J.; Ma, X. A new method to improve the capability of clays for removing red tide organisms. Oceanol. Limnol. Sin. 1994, 25, 226–232. [Google Scholar]
- Zhang, P.; Song, X.; Zhang, Y.; Shen, H.; Dong, X.; Li, J.; Yu, Z. Effect of Modified Clay on the Growth Dynamics and Physio—Biochemical Response of Newly Hatched Larvae of the Marine Medaka (Oryzias melastigma). J. Mar. Sci. Eng. 2021, 9, 822. [Google Scholar] [CrossRef]
- Yuan, J.; Wang, X.; Gu, Z.; Zhang, Y.; Wang, Z. Activity and Transcriptional Responses of Hepatopancreatic Biotransformation and Antioxidant Enzymes in the Oriental River Prawn Macrobrachium nipponense Exposed to Microcystin-LR. Toxins 2015, 7, 4006–4022. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yu, Z.; Song, X.; Cao, X. Effects of modified clay on the infant of Patinopecten yessoensisfor HABs control. Mar. Environ. Sci. 2014, 33, 817–821. [Google Scholar]
- Gao, Y.; Yu, Z.; Song, X.; Cao, X. Impact of Modified Clays on the Infant Oyster(Crassostrea gigas). Mar. Sci. Bull. 2007, 26, 8. [Google Scholar]
- Wang, Z.; Song, X.; Zhang, Y.; Yu, Z.; Tang, X. Effects of modified clay on Mercenaria Mercenaria. Oceanol. Limnol. Sin. 2019, 50, 692–699. [Google Scholar]
- Helm, M.M.; Hepper, B.T.; Spencer, B.E.; Walne, P.R. Lugworm mortalities and a bloom of Gyrodinium aureolum Hulburt in the eastern Irish Sea, Autumn 1971. J. Mar. Biol. Assoc. U. K. 1974, 54, 857–869. [Google Scholar] [CrossRef]
- Shi, Z.; Liu, J.; Li, H.; Cui, W.; Yang, W. Effects of some membrane lipids on the hemolysis induced by hemolytic toxin from Karenia mikimotoi. J. Hyg. Res. 2011, 40, 308–311. [Google Scholar]
- Zhu, J.; Yu, Z.; He, L.; Cao, X.; Song, X. Physiological response dynamics of the brown tide organism Aureococcus anophagefferens treated with modified clay. Harmful Algae 2019, 86, 1–9. [Google Scholar] [CrossRef]
- Guh, Y.-J.; Hwang, P.-P. Insights into molecular and cellular mechanisms of hormonal actions on fish ion regulation derived from the zebrafish model. Gen. Comp. Endocrinol. 2017, 251, 12–20. [Google Scholar] [CrossRef]
- Zhang, X.; Wen, H.; Qi, X.; Zhang, K.; Liu, Y.; Fan, H.; Yu, P.; Tian, Y.; Li, Y. Na+-K+-ATPase and nka genes in spotted sea bass (Lateolabrax maculatus) and their involvement in salinity adaptation. Comp. Biochem. Physiol. a-Mol. Integr. Physiol. 2019, 235, 69–81. [Google Scholar] [CrossRef]
- Agrahari, S.; Gopal, K. Inhibition of Na+-K+-ATPase in different tissues of freshwater fish Channa punctatus (Bloch) exposed to monocrotophos. Pestic. Biochem. Physiol. 2008, 92, 57–60. [Google Scholar] [CrossRef]
- Li, X.; Yan, T.; Zhang, Q.; Yu, R.; Zhou, M. Inhibition to crucial enzymes in the lethal effects of the dinoflagellate Karenia mikimotoi on the rotifer Brachionus plicatilis. Mar. Environ. Res. 2020, 157, 104866. [Google Scholar] [CrossRef] [PubMed]
- Chasiotis, H.; Kolosov, D.; Bui, P.; Kelly, S.P. Tight junctions, tight junction proteins and paracellular permeability across the gill epithelium of fishes: A review. Respir. Physiol. Neurobiol. 2012, 184, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, X.; Cheng, J.; He, Y.; Wang, X.; Wang, Z.; Qi, J.; Yu, H.; Zhang, Q. Transcriptome profiling provides gene resources for understanding gill immune responses in Japanese flounder (Paralichthys olivaceus) challenged with Edwardsiella tarda. Fish Shellfish Immunol. 2018, 72, 593–603. [Google Scholar] [CrossRef]
- Pinho, G.; Rosa, C.; Maciel, F.E.; Bianchini, A.; Yunes, J.S.; Proenca, L.A.O.; Monserrat, J.M. Antioxidant responses and oxidative stress after microcystin exposure in the hepatopancreas of an estuarine crab species. Ecotoxicol. Environ. Saf. 2005, 61, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Jian, L.I.; Jitao, L.I.; Tan, Z.; Ren, H.; Zhao, F. Toxic dinoflagellate Alexandrium tamarense induces oxidative stress and apoptosis in hepatopancreas of shrimp (Fenneropenaeus chinensis). J. Ocean. Univ. China 2014, 13, 7. [Google Scholar] [CrossRef]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Franco, R.; Sánchez-Olea, R.; Reyes-Reyes, E.M.; Panayiotidis, M.I. Environmental toxicity, oxidative stress and apoptosis: Ménage à Trois. Mutat. Res. 2009, 674, 3–22. [Google Scholar] [CrossRef]
- Xue, T.; Luo, P.; Zhu, H.; Zhao, Y.; Wu, H.; Gai, R.; Wu, Y.; Yang, B.; Yang, X.; He, Q. Oxidative stress is involved in Dasatinib-induced apoptosis in rat primary hepatocytes. Toxicol. Appl. Pharmacol. 2012, 261, 280–291. [Google Scholar] [CrossRef]
- Schattenberg, J.M.; Worns, M.A.; Zimmermann, T.; He, Y.W.; Galle, P.R.; Schuchmann, M. The role of death effector domain-containing proteins in acute oxidative cell injury in hepatocytes. Free. Radic. Biol. Med. 2012, 52, 1911–1917. [Google Scholar] [CrossRef] [Green Version]
- Savan, R.; Sakai, M. Genomics of fish cytokines. Comp. Biochem. Physiol. D-Genom. Proteom. 2006, 1, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Cao, J.; Qin, X.; Qiu, W.; Mei, J.; Xie, J. Toxic Effects on Bioaccumulation, Hematological Parameters, Oxidative Stress, Immune Responses and Tissue Structure in Fish Exposed to Ammonia Nitrogen: A Review. Animals 2021, 11, 3304. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Correlations | Algal Cell Density | Hemolytic Toxicity | |
|---|---|---|---|
| Algal cell density | Pearson Correlation | 1 | 0.974 ** |
| Sig.(2-tailed) | 0.000 | ||
| N | 18 | 18 | |
| Time (h) | Intact Algal Cell Density (Cells/mL) | 3000 Cells/mL | ||||
|---|---|---|---|---|---|---|
| 1000 | 3000 | 5000 | 10000 | The Cell-Free Culture Supernatant | The Ruptured Cell Suspension | |
| 1 |  |  | |  | | |
| 3 | | | | | | |
| 24 | | | | | | |
| 48 | | | | | | |
| 96 | | | | | | |
indicates all were alive, indicates some were alive, indicates all were dead.Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Song, X.; Zhang, Y.; Zhu, J.; Shen, H.; Yu, Z. Assessing the Effect of Modified Clay on the Toxicity of Karenia mikimotoi Using Marine Medaka (Oryzias melastigma) as a Model Organism. Toxics 2022, 10, 105. https://doi.org/10.3390/toxics10030105
Zhang P, Song X, Zhang Y, Zhu J, Shen H, Yu Z. Assessing the Effect of Modified Clay on the Toxicity of Karenia mikimotoi Using Marine Medaka (Oryzias melastigma) as a Model Organism. Toxics. 2022; 10(3):105. https://doi.org/10.3390/toxics10030105
Chicago/Turabian StyleZhang, Peipei, Xiuxian Song, Yue Zhang, Jianan Zhu, Huihui Shen, and Zhiming Yu. 2022. "Assessing the Effect of Modified Clay on the Toxicity of Karenia mikimotoi Using Marine Medaka (Oryzias melastigma) as a Model Organism" Toxics 10, no. 3: 105. https://doi.org/10.3390/toxics10030105
APA StyleZhang, P., Song, X., Zhang, Y., Zhu, J., Shen, H., & Yu, Z. (2022). Assessing the Effect of Modified Clay on the Toxicity of Karenia mikimotoi Using Marine Medaka (Oryzias melastigma) as a Model Organism. Toxics, 10(3), 105. https://doi.org/10.3390/toxics10030105