Abstract
The rainbow trout (Oncorhynchus mykiss) is one of the most commercially sought-after freshwater fish species and one of the most farmed in the world. On the other hand, aquaculture breeding frequently results in outbreaks of infectious diseases and pests, and compromises the production and welfare of fish. Arthrospira platensis (known as “Spirulina”) has been used as a supplement in diets to enhance fish welfare in recent years because of its beneficial properties. This study aimed to assess the possible protective effects of Arthrospira platensis on rainbow trout specimens exposed to three different doses of the toxicant CdCl2. The experiment was carried out using five experimental treatments of 40 individuals each: control group; group II (0.2 mg CdCl2 per kg of commercial fish feed); group III (0.2 mg Kg−1 of CdCl2 plus 2.5 g per kg of A. platensis); group IV (0.2 mg Kg−1 of CdCl2 plus 5 g per kg of A. platensis); group V (0.2 mg Kg−1 of CdCl2 plus 10 g per kg of A. platensis). During the experiment, dietary supplementation of A. platensis normalized all serum and blood parameters altered by the presence of CdCl2. A. platensis also had a protective effect on markers of oxidative stress.
1. Introduction
One of the freshwater fish species that is most commercially in demand and one of the most widely farmed worldwide is the rainbow trout (Oncorhynchus mykiss). The global production of rainbow trout in 2016 was roughly 0.8 million tonnes, accounting for 2% of all fish aquaculture worldwide production [1,2,3]. However, intensive production systems are susceptible to infectious diseases and pests, which generate stressful conditions in fish and interfere with their well-being.
The application of immunostimulants through food inclusion has been suggested as a possible approach to improving health and preventing illness [4]; it is also of great relevance for maintaining the quality of the fish and increasing their yield [5]. Numerous immunostimulants, including vitamins, chitin, glucans, microbes, and other readily accessible by-products, have been shown to have favorable effects [6,7]. The fact that medicinal plants and their derivatives include a variety of active ingredients makes them suitable as supplements to the nutritional diets of aquatic organisms such as rainbow trout. Indeed, these substances often result in beneficial effects on such organisms. [7,8,9]. Numerous recent studies indicate the benefits of these natural product constituents on the health of numerous aquatic animals. These compounds appear to be able to directly interact with the immune response [10,11,12] by boosting defensive activity, and they also have an antioxidant impact, protecting animals from oxidative stress [13,14]. Among these beneficial and natural sources of immunostimulants, there is Arthrospira platensis, a blue-green microalga (cyanobacteria) that has been utilized for generations as a food source. It has a spiral structure that varies in number and narrowness and is typically 0.1 mm in diameter [15]. The United Nations World Food Conference has put algae on its list of healthy foods due to its nutritional properties. Arthrospira platensis is thought to contain antiviral, antibacterial, antioxidant, anti-diabetic, anti-cancer, and anti-inflammatory properties in addition to its nutritional properties. As a result, this microalga is regarded as a superfood [16] and seems to also be effective in fish farming, since it appears to boost fish development [17], stress tolerance, and resistance to hunger [18,19]. It can also positively influence intestinal flora, lipid digestion, and coloration [20]. An interesting study [21] shows that in some species such as Oreochromis niloticus, the use of feed consisting of these algae improved antioxidant biomarkers, particularly in the gills and liver. Due to its positive contributions and in particular its protein component, A. platensis has been adopted as a supplement within diets in recent years [22]. For example, it is also used to reduce oxidative damage [23].
Indeed, fish in aquaculture facilities are exposed to many stressors daily (crowding stress, handling stress, pathogens, parasites, etc.), all of which affect fish health, growth, and performance. Heavy metals are the biggest threat to the health of aquatic organisms. Metals usually penetrate the surface and underground waters through industrial sewage contamination, mining activities, and municipal and agricultural wastewater. Cadmium (Cd) is one of the most known transition metals and is widely used in the electronics, plastics, and battery industries [24]. Therefore, Cd can be found in various aquatic ecosystems due to direct industrial discharges into waters or watersheds. Much of the cadmium that reaches aquatic systems tends to accumulate in sediments, and under certain conditions, re-enters the water column. Thus, finfish and shellfish may be exposed to water-borne Cd through feed or water. Although fish can eliminate a significant amount of Cd from their bodies, exposure to Cd may lead to biochemical, physiological, and behavioral disorders in these animals [25,26,27].
In this study, a stress condition was simulated by exposing rainbow trout to CdCl2, a known toxicant resulting from anthropogenic pollution and in particular from industry [28,29,30]. Furthermore, due to its poor excretion, Cd is toxic even at low doses [31,32] and has a negative impact on several metabolic processes in fish tissues and organs, and it has therefore been highly studied in several model organisms [33] and fish species [34,35,36,37,38]. It was then decided to evaluate whether the inclusion of A. platensis in the diet could contribute to increasing the robustness of the fish and alleviate the negative effects of stress. For this reason, this study aimed to assess the protective effects of A. platensis on rainbow trout specimens subjected to three different CdCl2 concentrations. Using A. platensis, it might be possible to reduce the side-effects of Cd in aquatic animals by increasing the efficiency of the detoxification system.
2. Materials and Methods
2.1. Sampling of the Specimens
The present study was conducted at the Fish Farm, Almas-Dime Village, Koohrang, Charmahal and Bakhtiari Province, in Iran, during the period from April to July 2016. According to the National Ethical Framework for Animal Research in Iran, two hundred animals belonging to the rainbow trout (Oncorhynchus mykiss) species were used for the performed analyses. Each of the specimens weighed 200 ± 10 g.
2.2. Experimental Design and Diet
The experiment lasted for 21 days, during which a total of 200 juvenile rainbow trout (Oncorhynchus mykiss) were randomly distributed in 5 concrete gullies (10,000 L) and acclimatized in aerated freshwater (16 ± 2 °C; pH, 7.4 ± 0.2; 100% water exchange/day; a natural photoperiod) for a fortnight prior to the experiment. During the acclimatization period, the fish were fed twice daily with commercial feed from the “Faradaneh Company, Sharkourd”, produced in Iran (Table 1). Moreover, CdCl2 (99% purity, Merck Co., Darmstadt, Germany) was orally administered to fish and the Arthrospira platensis treatment. Dry A. platensis powder was obtained from Sinamicroalgae Co. (Qeshm, Iran).

Table 1.
A proximate composition of the experimental diets for the rainbow trout.
The experiment was carried out using a completely randomized model with five experimental treatments of 40 individuals each:
- Group I, or the control group, was fed on a basal diet without any treatment;
- Group II was fed with a basal diet, and 0.2 mg CdCl2 per kg of commercial fish feed;
- Group III was fed simultaneously with 0.2 mg Kg−1 of CdCl2 and supplemented with 2.5 g A. platensis;
- Group IV was fed simultaneously with 0.2 mg Kg−1 CdCl2 and supplemented with 5 g A. platensis per kg commercial fish feed;
- Group V was fed simultaneously with 0.2 mg Kg−1 CdCl2 and supplemented with 10 g A. platensis per kg commercial fish feed.
According to the information provided on brochure of commercial diet purchased from Faradaneh Co. (Sharkourd, Iran), the proximate analysis of basal diet indicated 39–40% crude protein, 10.5–11% crude lipid, and 2–2.5% fiber [39].
2.3. Preparing Samples
Twelve fish from each group were collected after 21 days of CdCl2 exposure and Arthrospira platensis (AP) treatment and anaesthetized with clove powder (150 mg L−1). Next, blood samples were collected from the caudal vein with a 2.5 cc syringe and poured into a microtube (2 mL). After centrifuging the blood samples (4 °C, 15 min, 6000 rpm), the supernatant was separated to measure blood biochemical parameters using biochemical reagents obtained from ParsAzmun Co., Iran (Baharstan Industrial Township, Karaj, Alborz, Iran). Then, the fish were euthanized, and their livers were extracted, rinsed with a physiological solution, and homogenized in a cold phosphate buffer solution for 2 min (pH: 4.7). The resulting homogenized solution was centrifuged at 150,000 rpm for 15 min at 4 °C. The supernatant was collected for biochemical parameter measurements and kept at −70 °C until biochemical analysis [40].
2.4. Blood Biochemical Parameter Analyses Performed
Relevant blood parameters were evaluated, such as the glucose, cholesterol, triglyceride, total protein, albumin, creatinine contents and aspartate aminotransferase, alanine aminotransferase, alkaline phosphatase, lactate dehydrogenase, creatinine phosphokinase, gamma-glutamyltransferase, and butyrylcholinesterase activities. These parameters were measured using biochemical reagents purchased from ParsAzmun Co. (A producer of clinical chemistry and immunoturbidimetric reagents, in Iran). All serum parameters were determined strictly according to the manufacturer’s instructions. Briefly, biochemical reagents that included buffer and substrate or enzyme were added to a certain amount of sample and then incubated. Finally, changes in wavelength absorption were measured with a spectrophotometer, and concentrations of analytes or activities of enzymes were calculated following the manufacturer’s instructions.
Glucose contents in serum were determined using glucose oxidase [41]. Triglyceride and cholesterol concentrations were assessed in the presence of lipase and cholesterol esterase, respectively [42]. Pyric acid was used to measure creatinine [43]. Total protein and albumin were estimated using bromocresol green and cupric ion, respectively. Globulin content was also calculated by subtracting albumin from total protein [44].
The level of the aspartate aminotransferase (AST) enzyme was determined using a paired reaction with malate dehydrogenase in the presence of NADH. The enzyme can react with alanine and a-ketoglutarate to produce glutamate and pyruvate during the alanine aminotransferase (ALT) assay. The AST and ALT activities were determined by measuring the absorbance variation in 3 min at 340 nm. The lactate dehydrogenase (LDH) activity was evaluated by measuring the conversion of pyruvate in L-lactate and monitoring NADH oxidation for 3 min at 340 nm. The activity of alkaline phosphatase (ALP) was determined by measuring the conversion of p-nitrophenol phosphate to nitrophenol at 405 nm in an alkaline buffer. CPK (creatinin phosphokinase) activity was measured using creatinine phosphate and adenosine diphosphate (ADP) as substrates at 340 nm [45]. BChE activity was determined using butyrylcholine at 405 nm [46]. GGT activity was evaluated using glutamic acid at 405 nm [45]. Total immunoglobulin (Ig) levels were detected using polyethylene glycol following the technique described by Banaee et al. [47], at 540 nm.
Using the FRAP reagent, total antioxidant capacity was determined based on the plasma’s ferric-reductive ability. The freshly made FRAP reagent consisted of 5 mL of 10 mmol/L TPTZ (2,4,6-tripyridyl-s-triazine) solution in 40 mmol/L HCl, 5 mL of 20 mmol/L FeCl3, and 50 mL of acetate buffer (0.3 mol/L, pH = 3.6). Following that, 3 mL of FRAP reagent was combined with 100 µL of supernatant aliquots. As the conversion rate of the ferric tri-pyridyl-s-triazine complex (Fe3+-TPTZ) to ferrous tri-pyridyl-s-triazine complex (Fe2+-TPTZ) at pH 3.6 and 25 °C is directly proportional to the total antioxidant concentration in the sample, through this method it was possible to obtain the data. The Fe2+-TPTZ complex has a strong blue color that may be detected using a UV/VIS spectrophotometer for 5 min at 593 nm. Furthermore, calculations were performed using a calibration curve of FeSO4-7H2O (100 to 1000 µM/L) [48].
Malondialdehyde (MDA) level was expressed as mol/g tissue and was estimated by using the modulated thiobarbituric acid test. In this context, 500 µL of supernatant was mixed with 2500 µL of trichloroacetic acid (20%) and 1000 µL of thiobarbituric acid (67%), in a Pyrex tube. Tubes were then placed in hot water at 100 °C for 15 min. After boiling, the organic phase of the chromogenic substrate was extracted with 1000 µL of distilled water and 5000 µL of n-butanol:pyridine (15:1). The mixture was subsequently centrifuged at 2000 rpm for 15 min at 4 °C. The reaction resulted in a pink-colored complex that was measured using a spectrophotometer at 532 nm to detect MDA levels, and its concentration was quantified using the MDA standard, which was prepared and synthesized from tetraethoxypropane and absolute ethanol [49].
Catalase (CAT) activity was determined by making some variations to the above-mentioned kits. Indeed, the hydrogen peroxidase assay was used, which is based on the formation of a stable complex by the addition of ammonium molybdate. The reaction produced a yellow complex, whose concentration was measured at 405 nm [50].
Blank1 included 1.0 mL substrate, 1.0 mL molybdate, and 0.2 mL distilled water; blank2 included 1.0 mL substrate, 1.0 mL molybdate, and 0.2 mL buffer; and blank3 included 1.0 mL buffer, 1.0 mL molybdate, and 0.2 mL buffer.
Superoxide dismutase, glutathione reductase, glutathione peroxidase, and glucose 6 phosphate dehydrogenase activities were tested in supernatant obtained from liver tissue homogenate using biochemical reagents purchased from Biorex-Fars Co. (Shiraz, Iran). SOD activity was assayed using xanthine oxidase and xanthine to produce superoxide anions. GPx activity was estimated utilizing reduced glutathione (GSH) and cumene hydroperoxide. GR activity was detected using NADPH and oxidized glutathione (GSSG) [51]. G6PDH activity was measured using glucose 6-phosphate as substrate [52].
All biochemical endpoints above-mentioned were measured using a UV/VIS spectrophotometer (Biochrom Libra S22 model, Waterbeach Cambridge, CB25 9PE, UK)
2.5. Measurement of Cadmium Bioaccumulation
After the autopsy, muscle, skin, gills, and liver samples were cut and dried in the oven. Then, 1 g of each tissue was mixed with 5 mL of H2O2 and kept overnight at room temperature (25 °C). Next, samples were blended with 15 mL of acid mixture concentrated HNO3/HCl (3:1) and digested at 150 °C for 12 h. Digested samples were cooled and filtered using a Whatman filter (0.22 μm) and diluted in deionized water to a final volume of 25 mL. Finally, cadmium concentrations in the samples were estimated using inductively coupled plasma optical emission spectrometry (ICP–OES spectrometer provided by SPECTRO Analytical Instruments GmbH Boschstr. 10, 47533 Kleve, Germany) [53].
2.6. Data Analyses
To verify the normality of data, the Kolmogorov–Smirnov normality test was performed using SPSS, version 22 (Chicago, IL, USA). One-way ANOVA was used to analyze the data. The significant differences between experimental groups were calculated with Tukey’s post hoc test at both p < 0.05 and p < 0.01.
3. Results
3.1. Comments on the Clinical Status
Neither mortality nor clinical indications were documented in either the control or treatment groups during the experiment. In the CdCl2 exposed group, however, only fast opercular movement was seen, indicating increased respiration.
3.2. Serum Biochemical Parameters
Regarding all test statistics, degrees of freedom, and p-values (p < 0.01) and (p < 0.05) of all ANOVAs conducted, please refer to the tables in the Supplementary Materials.
After exposure to CdCl2, the amount of total protein decreased compared to the control; the difference was significant (p < 0.01) for the first group. In contrast, all groups that received the AP supplement had a progressive increase in the total amount of protein compared to the control group, as opposed to exposure to Cd alone. As for albumin levels, these remained superimposable to those of the control group (Table 2).
Table 2.
Results regarding analysis of serum biochemical parameters.
The values of globulins decreased following exposure to cadmium (p < 0.05); treatment with AP, especially in the group exposed to the highest AP concentrations, brought the values back to a level similar to that of the control. In this case, the values were not significantly different. Total immunoglobulin levels significantly (p < 0.01) increased in fish exposed to 0.2 mg CdCl2, relative to controls. In contradistinction, the use of CdCl2 feed was responsible for significant (p < 0.01) increases in certain blood values—in particular, glucose, cholesterol, and triglycerides. Again, the administration of AP resulted in an average drop in values in the groups with AP concentrations of 2.5 g (group III) and 5.0 g (group IV). The use of a greater dose of 10.0 mg in the final experimental group resulted in a decrease in the values compared to the control (Table 2).
The creatinine level in group I (0.2 mg CdCl2 alone) was significantly (p < 0.01) higher in comparison to the control group. Experimental groups treated with increasing concentrations of AP showed a trend directly proportional to the administered AP concentrations. Indeed, fish belonging to group III, treated with 2.5 g of AP, showed significant divergence in creatinine level compared to the control group, despite the amount of creatinine administered being less than in group II. Fish belonging to groups IV and V treated with 5 and 10 g of AP showed creatinine levels comparable to that of the control group (Table 2).
In fish fed with 0.2 mg CdCl2 (II group), AST activity in serum was significantly (p < 0.01) higher compared to the control group (I group). The AST activity in fish treated with the highest concentration of AP was similar to the AST activity in the control group. In the groups treated with 2.5 g (group III) and 5 g (group IV) of AP, the divergence with the control group was still higher, while at the highest administered concentration of AP (V group), AST activity decreased compared to the control group, despite the value is not statistically significant (Table 3).
Table 3.
Results on serum biochemical parameters.
Fish which were CdCl2-treated, belonging to group II (0.2 mg CdCl2), showed a significant (p < 0.01) increase in ALT activity in serum. Group III fish, treated with 2.5 g SP, showed a significant decreasing trend in ALT activity. In groups IV (5 g AP) and V (10.0 g AP), ALT values in comparison to the control group were maintained (Table 3).
CdCl2 treatment of group-II fish (0.2 mg CdCl2 alone) resulted in a relevant significant (p < 0.01) increase in LDH activity compared to the control group. The group-III fish, treated with 2.5 g of AP, also showed a trend of LDH activity significantly higher than the control. The 5 g AP-treated group (group III) had less high LDH activity than group II but still higher than the control group. A non-significant (p > 0.05) LDH activity difference with respect to the control group was highlighted in the group (V group) treated with 10 g of AP (Table 3).
The results revealed that the activity of ALP significantly (p < 0.01) increased in the serum of fish orally exposed to 0.2 mg CdCl2 without AP. In comparison, no significant changes were observed between fish treated with 0.2 mg CdCl2 plus AP (5 and 10 mg AP per kg feed) and the control group.
GGT activity in the sera of group II (0.2 mg CdCl2 without AP) was significantly (p < 0.01) higher compared to the control group. The same was found for the 2.5 g AP-treated group (group III), and the trend in GGT activity was decreased in the experimental group treated with 10 g AP (group V) compared to the control, though the difference was non-significant (Table 3).
In CdCl2-treated fish (group II—0.2 mg CdCl2 without AP), enzymatic BchE activity in serum showed a significant decrease compared to the control group. In groups treated with various concentrations of AP, BchE activity was significantly (p < 0.01) decreased compared to the control group, and the trend was directly proportional to the AP concentration administrated (Table 3).
Group II (0.2 mg CdCl2 alone) showed a significant (p < 0.01) increase in CPK activity in serum, whereas the groups treated with higher concentrations of AP (III, IV, and V groups) maintained comparable enzymatic CPK activities compared to the control group (Table 3).
3.3. Tissue Antioxidant and Oxidative Stress Markers
Tissue homogenate of the livers of Oncorhynchus mykiss was prepared to investigate the antioxidant status and oxidative stress markers in the CdCl2-treated group of fish and the preventive role of AP (Figure 1).
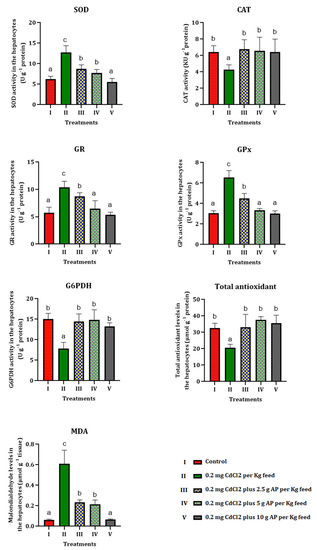
Figure 1.
Results on tissue antioxidant values and oxidative stress markers. Different letters show significant changes in values (p < 0.05) and (p < 0.01), and the same letter shows there was no significant difference between the experimental groups. One-way ANOVA was used to analyze the data. Duncan’s test was used for the comparison, with confidence levels of 95% (p = 0.05) and 99% (p = 0.01).
The level of total cellular antioxidants in the homogenized liver tissue of fish exposed to CdCl2 decreased significantly (p < 0.01) compared to the control. The administration of 2.5, 5, and 10 g of AP, in contrast, led to an increase in the total antioxidant level, which recovered to the value of the control in group V (basal diet with 0.2 mg CdCl2, plus 10 g AP).
The MDA levels in the livers of the fish treated with CdCl2 alone (group II) were significantly (p < 0.01) higher compared to the control group; and the gradual inclusion of AP in the diet of the fish (2.5, 5.0, 10.0 g AP) had a beneficial effect, bringing the MDA values back to the same level as the control group, thereby eliminating the alteration produced by CdCl2. A significant (p < 0.05) decrease in CAT values was found in group II (0.2 mg CdCl2 without AP), which started to gradually increase with the addition of AP in groups III (2.5 g AP), IV (5 g AP) and V (10.0 g AP), reaching, in the group with the highest AP concentration, the control group’s values. In the group exposed to CdCl2 alone, a significant (p < 0.01) increase in hepatic SOD was observed, which gradually subsided with AP supplementation of 2.5, 5, and 10.0 g. Dietary supplementation of AP resulted in almost full recovery and normalization of hepatic SOD values (Figure 1).
Glutathione reductase (GR) levels also increased significantly (p < 0.01) in group II, and then decreased with increasing AP supplementation in the fish’s diet. Similar results were observed in the analyses of the hepatic glutathione peroxidase (GPx), which was significantly (p < 0.01) elevated in the group exposed to CdCl2 alone, and then gradually decreased to levels overlapping those of the control in the group fed the highest concentration of AP (group V). Glucose-6-phosphate dehydrogenase values (p < 0.01) decreased in the presence of the pollutant alone (group II) and then returned to the initial values in groups III (2.5 g AP), IV (5 g AP), and V (10.0 g AP), i.e., those in which AP was present in the diet (Figure 1).
3.4. Bioaccumulation of Cadmium
Results showed that feeding fish with a polluted diet by Cd increased its bioaccumulation levels in various tissues. Although administration of AP could mitigate Cd bioaccumulation, there was a significant difference between Cd contents in the different tissues of the experimental fish and control group (Figure 2).
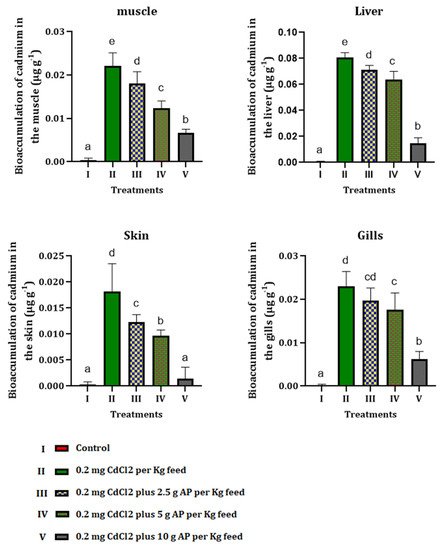
Figure 2.
Bioaccumulation of cadmium in different tissues of fish orally exposed to CdCl2 and Arthrospira platensis. Different letters show significant changes in values (p < 0.05) and (p < 0.01), and the same letter by different numbers shows there was no significant difference between the experimental groups.
4. Discussion
Cadmium is one of the most toxic heavy metals. It has a wide distribution in the environment. It is a non-essential heavy metal that can bioaccumulate and be toxic to organisms even at small concentrations [54]. It has also been demonstrated to induce free-radical formation, resulting in oxidative damage to lipids, proteins, and DNA [55,56]. The toxic effects of Cd are numerous, including stunted development and growth [57], disturbances in liver function [58], and pathological alterations in certain tissues and organs [59]. A 2018 study [60] assessed the acute sensitivity of the larval stage of Nothobranchius furzeri to cadmium in combination with a 4 °C temperature increase. Cadmium was found to be highly toxic, with 100% mortality being achieved very rapidly at the two highest concentrations. In addition, a recent study [61] showed that exposure to cadmium can also affect the maturation time and reproductive performance of fish. In this study, cadmium delayed maturation in females (Nothobranchius furzeri) and reduced adult mass and fecundity.
Furthermore, as the metal concentration rises and exceeds the capacity of the organisms’ detoxification mechanisms, a variety of deleterious consequences and increased mortality emerge [62,63]. The various reactions to Cd exposure are undoubtedly connected to oxidative stress, which is characterized as an imbalance between oxidant fluxes and antioxidant defenses [64], which in turn determines an increase in the activities of antioxidant enzymes. These findings are also in agreement with data obtained by Al-Asgah et al. in 2015 [65].
Arthrospira platensis is described as a powerful tonic for the immune system and for boosting animals’ growth and development. Due to these properties, Arthrospira is utilized as a supplemental ingredient in fish feeds, and increasingly, as a protein and vitamin supplement in aquarium feeds [16].
Interestingly, CdCl2 reduced the concentration of serum protein in exposed fish—in particular, total protein and globulins. As for albumin levels, these remained superimposable with those of the control group. Serum-globulin deficiency obviously suggests liver failure. Furthermore, it is suggested that the decrease in serum protein content is due to an increase in stressor levels in exposed fish [66,67]. Dietary supplementation of AP alleviates the toxicity induced by CdCl2, and the concentration of serum protein is restored to its normal level, reestablishing normal hepatic function.
During cadmium exposure to the rainbow trout specimens in the study, serum glucose and cholesterol were significantly elevated in the group exposed to CdCl2. This increase is a clear response to the damage that cadmium can cause in fish organs and is considered an indicator of acute stress due to the action of the hormone cortisol, which stimulates increases in glycogenesis and gluconeogenesis. Furthermore, a rise in serum cholesterol is thought to be a sign of cell-structure degradation in the membranes of kidney and muscle cells [68]. Thus, increases in cholesterol levels are good indicators of environmental stress in fish [69]. Serum cholesterol and glucose levels reverted to normal in the AP-treated groups, confirming Arthrospira’s anti-stress action against xenobiotic toxicity and its protective impact on liver tissue [58].
Triglycerides have the main function of supplying energy to cells and can be used as indicators of nutritional status. The present study showed a slight increase in serum triglyceride concentration in the cadmium-exposed group, which returned to the control level after AP treatment. This could be related to the liver failure of the fish [16]. Triglyceride levels in metal exposed Perca flavescens [70] were also altered in the same way. Furthermore, rises in triglyceride and cholesterol levels in fish plasma may be a physiological response to supply enough energy to mitigate the harmful effects of this contaminant [71].
AST and ALT are non-organ-specific enzymes and are found in two different cytosolic and mitochondrial isoforms in all animal tissues. The activity of AST and ALT enzymes in the blood can be used as an indicator of stress [65]. Increased AST and ALT values in fish reveal the exportation of enzymes from the liver to the bloodstream [72], which may indicate hepatocellular, mitochondrial, or cell membrane damage. AST plays a crucial role in both glutathione biosynthesis and gluconeogenesis in hepatocytes. Therefore, an increase in plasma AST activity may be indicative of oxidative stress and tissue damage in rainbow trout specimens after CdCl2 exposure. These findings agree with those of Shalaby et al. [71] from a study conducted in 2007, in which they found that sub-lethal concentrations of Cd caused significant increases in AST and ALT in O. niloticus. There is a similar result also in a study from 2015 [65]. In this research, dietary supplementation with AP improved liver function by reducing the activity of both hepatic transaminases and alkaline phosphatase. The antioxidant compounds in AP, including ß-carotene, vitamins, and minerals, are thought to contribute significantly to protecting hepatic tissues from xenobiotic damage. Indeed, Arthrospira platensis showed a hepatoprotective effect in CdCl2-exposed fish by dramatically lowering serum transaminase and ALP activities.
Regarding LDH values in fish exposed to cadmium, there was a highly significant increase compared to the control group. LDH plays an essential role in the conversion of lactate to pyruvate, NAD+ to NADH, and vice versa in both cases. Hypoxic conditions, dysfunction of mitochondrial oxidation, necrosis, or cell death can increase LDH activity in the blood [73,74,75]. Similar results were observed in the blood of carp (Cyprinus carpio) exposed to microplastics and cadmium in a 2019 study [47]. The influence of the Arthrospira treatment, on the other hand, helped to bring the values back to normal, again confirming its protective and beneficial effect.
An increase in GGT activity was also identified during the investigation. GGT is a membrane-bound enzyme involved in glutathione reformation and biodegradation and xenobiotic detoxification. Therefore, GGT activity is essential to provide amino acids for the synthesis and renewal of intracellular glutathione [76]. Increased GGT activity indicates depletion of cellular glutathione, especially in hepatocytes, resulting in oxidative stress [77].
BChE is found in all tissues and cells, including erythrocytes [78]. It is involved in, among other things, the transmission of cellular signals. Oxidative stress and lipid peroxidation could be responsible for a significant decrease in BChE activity [77] in fish exposed to CdCl2. In any case, the Arthrospira-treated groups appear to restore normal values, acting in opposition to oxidative stress.
CPK is present in high concentrations in animal muscle cells, heart tissue, gills, kidneys, and the brain and can be released into circulation because of cell injury [79]. Increased CPK activity in the serum of fish can be due to muscle or renal damage.
An increase in creatinine levels was also found in the cadmium-treated fish compared to the control fish. Creatinine is metabolically produced by the breakdown of creatine phosphate during muscle and protein metabolism. The kidneys take care of its excretion. Therefore, if the kidneys are not functioning properly, or have any injuries, the creatinine level in the blood increases. In fact, the blood creatinine level is an excellent biomarker for assessing the glomerular filtration rate [47]. By reversing CPK and creatinine elevation, Arthrospira platensis could play an important role in the prevention and treatment of liver and kidney diseases, especially those mediated by oxidative stress.
The SOD-CAT system is a key component of the antioxidant defense that can exert a protective effect against oxidative stress by converting hydrogen peroxide into oxygen and water [80]. During the experiment, there was a decrease in CAT activity, which may be due to the direct effect of the metal [81]. In general, the inhibition of CAT activity is related to the binding of metal ions to the enzyme’s -SH groups, which increases the H2O2 or superoxide radical concentrations [82]. The decrease in CAT can also be linked to overproduction of ROS or altered gene expression. Similar results were also obtained in other ecotoxicology studies on fish [83,84]. The high activity of SOD resulting from exposure to CdCl2 may indicate high production of superoxide anion radicals [85,86].
The potential of oxidative stress formation increases as overall antioxidant capability decreases [87,88,89,90]. Total antioxidant levels were shown to be lower in fish exposed to cadmium. The decrease in total antioxidants, along with the rise in MDA, implies that CdCl2 exposure may lead to oxidative stress in fish. MDA levels rose considerably after metal exposure compared to the control group. Peroxidation of essential macromolecules such as lipids and proteins can result in the formation of metabolites such as MDA and protein carbonyl derivatives [88,89,90]. As a result, elevated MDA can be a good biomarker for detecting oxidative stress and the failure of the antioxidant defense mechanism.
Results showed a significant increase in the GPx activity of hepatocytes after exposure to cadmium. This increase in GPx activity may hasten the conversion of H2O2 and other proxide radicals to H2O and O2 and may minimize the formation of ROS in tissues [77,88].
As G6PD is a regulatory enzyme for NADPH-dependent biotransformation and defense against oxidative stress, a reduction in G6PD activity may lead to a decrease in NADPH production. NADPH is essential to maintaining glutathione in the reduced form, which reduces peroxides and protects cells from oxidative damage in the course [89,90].
A significant decrease in the Cd bioaccumulation may be related to phytochemical compounds of AP, especially flavonoids, which can inhibit the absorption of heavy metals in the digestive system. Panche et al. [91] showed that flavonoids could decrease the metal accumulation rate in the biological system. Moreover, AP antioxidants can also play an essential role in the cadmium detoxification system and remove it from the fish’s body. Bhattacharya [92] found that the administration of A. platensis can alleviate the toxicity effects of heavy metals through increased cellular antioxidant capacity.
During the experiment, the role of Arthrospira platensis became evident in bringing all values altered by the presence of CdCl2 back to normal, compared to the control. In the case of oxidative stress markers, therefore, one can see how A. platensis exerted a protective effect.
5. Conclusions
According to the experiment conducted on Oncorhynchus mykiss, exposure to CdCl2 induced an alteration in serum biochemical parameters, alterations in liver function biochemical parameters, reductions in antioxidant enzyme activities, and increases in markers of oxidative stress. The study found that Arthrospira platensis supplementation, because of its beneficial multi-properties, provided nearly total protection, minimizing or eliminating the detrimental effects caused by the heavy metal utilized in the study. Additionally, these results suggest that inclusion of Arthrospira platensis into the diet of farmed fishes may help to boost their robustness to stress, which may positively affect the wellbeing, quality, and yield of fish in aquacultural production systems.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/toxics10120731/s1. Table S1: One-way ANOVA test of serum biochemical parameters—part a. Table S2: One-way ANOVA test of serum biochemical parameters—part b. Table S3: One-way ANOVA test of tissue antioxidant values and oxidative stress markers.
Author Contributions
Conceptualization, H.E.-Z.S. and M.B.; validation, G.P. and C.F.; investigation, H.E.-Z.S. and M.B.; data curation, M.B.; writing—original draft preparation, F.I.; writing—review and editing, F.I. and M.B.; supervision, G.P. and C.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Agriculture Organization of the United Nations. The State of World Fisheries and Aquacuture; Fisheries Department, Agriculture Organization of the United Nations: Rome, Italy, 2000. [Google Scholar]
- Rashidian, G.; Gorji, S.B.; Farsani, M.N.; Prokić, M.D.; Faggio, C. The oak (Quercus brantii) acorn as a growth promotor for rainbow trout (Oncorhynchus mykiss): Growth performance, body composition, liver enzymes activity and blood biochemical parameters. Nat. Prod. Res. 2020, 34, 2413–2423. [Google Scholar] [CrossRef] [PubMed]
- Vaclavik, J.; Sehonova, P.; Hodkovicova, N.; Vecerkova, L.; Blahova, J.; Franc, A.; Marsalek, P.; Mares, J.; Tichy, F.; Svobodova, Z.; et al. The effect of foodborne sertraline on rainbow trout (Oncorhynchus mykiss). Sci. Total. Environ. 2020, 708, 135082. [Google Scholar] [CrossRef] [PubMed]
- Jeyavani, J.; Sibiya, A.; Sivakamavalli, J.; Divya, M.; Preetham, E.; Vaseeharan, B.; Faggio, C. Phytotherapy and combined nanoformulations as a promising disease management in aquaculture: A review. Aquac. Int. 2022, 30, 1071–1086. [Google Scholar] [CrossRef]
- Guardiola, F.A.; Porcino, C.; Cerezuela, R.; Cuesta, A.; Faggio, C.; Esteban, M. Impact of date palm fruits extracts and probiotic enriched diet on antioxidant status, innate immune response and immune-related gene expression of European seabass (Dicentrarchus labrax). Fish Shellfish Immunol. 2016, 52, 298–308. [Google Scholar] [CrossRef]
- Tukmechi, A.; Andani, H.R.R.; Manaffar, R.; Sheikhzadeh, N. Dietary administration of beta-mercapto-ethanol treated Saccharomyces cerevisiae enhanced the growth, innate immune response and disease resistance of the rainbow trout, Oncorhynchus mykiss. Fish Shellfish Immunol. 2011, 30, 923–928. [Google Scholar] [CrossRef] [PubMed]
- Rashidian, G.; Shahin, K.; Elshopakey, G.E.; Mahboub, H.H.; Fahim, A.; Elabd, H.; Prokić, M.D.; Faggio, C. The Dietary Effects of Nutmeg (Myristica fragrans) Extract on Growth, Hematological Parameters, Immunity, Antioxidant Status, and Disease Resistance of Common Carp (Cyprinus carpio) against Aeromonas hydrophila. J. Mar. Sci. Eng. 2022, 10, 325. [Google Scholar] [CrossRef]
- Rashidian, G.; Boldaji, J.T.; Rainis, S.; Prokić, M.; Faggio, C. Oregano (Origanum vulgare) Extract Enhances Zebrafish (Danio rerio) Growth Performance, Serum and Mucus Innate Immune Responses and Resistance against Aeromonas hydrophila Challenge. Animals 2021, 11, 299. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Dawood, M.A.O.; Alagawany, M.; Faggio, C.; Nowosad, J.; Kucharczyk, D. Health benefits and potential applications of fucoidan (FCD) extracted from brown seaweeds in aquaculture: An updated review. Fish Shellfish Immunol. 2022, 122, 115–130. [Google Scholar] [CrossRef]
- Rashidian, G.; Kajbaf, K.; Prokić, M.D.; Faggio, C. Extract of common mallow (Malvae sylvestris) enhances growth, immunity, and resistance of rainbow trout (Oncorhynchus mykiss) fingerlings against Yersinia ruckeri infection. Fish Shellfish Immunol. 2020, 96, 254–261. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Shakouri, M.; Yousefi, S.; Van Doan, H.; Shafiei, S.; Yousefi, M.; Mazandarani, M.; Mozanzadeh, M.T.; Tulino, M.G.; Faggio, C. Humoral and skin mucosal immune parameters, intestinal immune related genes expression and antioxidant defense in rainbow trout (Oncorhynchus mykiss) fed olive (Olea europea L.) waste. Fish Shellfish Immunol. 2020, 100, 171–178. [Google Scholar] [CrossRef]
- Chakraborty, S.B.; Horn, P.; Hancz, C. Application of phytochemicals as growth-promoters and endocrine modulators in fish culture. Rev. Aquac. 2014, 6, 1–19. [Google Scholar] [CrossRef]
- Moure, A.; Cruz, J.M.; Franco, D.; Domínguez, J.; Sineiro, J.; Domínguez, H.; Núñez, M.J.; Parajó, J. Natural antioxidants from residual sources. Food Chem. 2001, 72, 145–171. [Google Scholar] [CrossRef]
- Vazirzadeh, A.; Marhamati, A.; Rabiee, R.; Faggio, C. Immunomodulation, antioxidant enhancement and immune genes up-regulation in rainbow trout (Oncorhynchus mykiss) fed on seaweeds included diets. Fish Shellfish Immunol. 2020, 106, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Saranraj, P.; Sivasakthi, S. Spirulina platensis—Food For Future: A Review. Asian J. Pharm. Sci. Technol. 2014, 4, 26–33. [Google Scholar]
- Jung, F.; Krüger-Genge, A.; Waldeck, P.; Küpper, J.-H. Spirulina platensis, a super food? J. Cell. Biotechnol. 2019, 5, 43–54. [Google Scholar] [CrossRef]
- Roohani, A.M.; Kenari, A.A.; Kapoorchali, M.F.; Borani, M.S.; Zoriezahra, S.J.; Smiley, A.H.; Esmaeili, M.; Rombenso, A.N. Effect of spirulina Spirulina platensis as a complementary ingredient to reduce dietary fish meal on the growth performance, whole-body composition, fatty acid and amino acid profiles, and pigmentation of Caspian brown trout (Salmo trutta caspius) juveniles. Aquac. Nutr. 2019, 25, 633–645. [Google Scholar] [CrossRef]
- Nandeesha, M.C.; Gangadhar, B.; Varghese, T.J.; Keshavanath, P. Effect of feeding Spirulina platensis on the growth, proximate composition and organoleptic quality of common carp, Cyprinus carpio L. Aquac. Res. 1998, 29, 305–312. [Google Scholar] [CrossRef]
- Kumar, A.; Ramamoorthy, D.; Verma, D.K.; Kumar, A.; Kumar, N.; Kanak, K.R.; Marwein, B.M.; Mohan, K. Antioxidant and phytonutrient activities of Spirulina platensis. Energy Nexus 2022, 6, 100070. [Google Scholar] [CrossRef]
- Mohammadiazarm, H.; Maniat, M.; Ghorbanijezeh, K.; Ghotbeddin, N. Effects of spirulina powder (Spirulina platensis) as a dietary additive on Oscar fish, Astronotus ocellatus: Assessing growth performance, body composition, digestive enzyme activity, immune-biochemical parameters, blood indices and total pigmentation. Aquac. Nutr. 2021, 27, 252–260. [Google Scholar] [CrossRef]
- Sheikhzadeh, N.; Mousavi, S.; Oushani, A.K.; Firouzamandi, M.; Mardani, K. Spirulina platensis in rainbow trout (Oncorhynchus mykiss) feed: Effects on growth, fillet composition, and tissue antioxidant mechanisms. Aquac. Int. 2019, 27, 1613–1623. [Google Scholar] [CrossRef]
- Zhang, F.; Man, Y.B.; Mo, W.Y.; Wong, M.H. Application of Spirulina in aquaculture: A review on wastewater treatment and fish growth. Rev. Aquac. 2020, 12, 582–599. [Google Scholar] [CrossRef]
- Mahmoud, Y.I.; Shehata, A.M.M.; Fares, N.H.; Mahmoud, A.A. Spirulina inhibits hepatocellular carcinoma through activating p53 and apoptosis and suppressing oxidative stress and angiogenesis. Life Sci. 2021, 265, 118827. [Google Scholar] [CrossRef] [PubMed]
- McRae, N.K.; Gaw, S.; Glover, C.N. Effects of waterborne cadmium on metabolic rate, oxidative stress, and ion regulation in the freshwater fish, inanga (Galaxias maculatus). Aquat. Toxicol. 2018, 194, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sierra-Marquez, L.; Espinosa-Araujo, J.; Atencio-Garcia, V.; Olivero-Verbel, J. Effects of cadmium exposure on sperm and larvae of the neotropical fish Prochilodus magdalenae. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 225, 108577. [Google Scholar] [CrossRef]
- Vicentini, M.; Fernandes, L.D.S.P.; Marques, A.E.M.L.; Osório, F.H.T.; Baika, L.M.; Risso, W.E.; Martinez, C.B.D.R.; Grassi, M.T.; Fávaro, L.F.; Mela, M.; et al. Effects of cadmium on the female reproductive axis of a Neotropical fish. Chemosphere 2022, 286, 131639. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-C.; Choi, Y.J.; Kim, J.-H. Toxic effects of waterborne cadmium exposure on hematological parameters, oxidative stress, neurotoxicity, and heat shock protein 70 in juvenile olive flounder, Paralichthys olivaceus. Fish Shellfish Immunol. 2022, 122, 476–483. [Google Scholar] [CrossRef]
- Pagano, M.; Savoca, S.; Impellitteri, F.; Albano, M.; Capillo, G.; Faggio, C. Toxicological Evaluation of Acetylsalicylic Acid in Non-Target Organisms: Chronic Exposure on Mytilus galloprovincialis (Lamarck, 1819). Front. Physiol. 2022, 13, 1165. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy Metal Toxicity and the Environment. Mol. Clin. Environ. Toxicol. 2012, 101, 133–164. [Google Scholar] [CrossRef]
- Pagano, M.; Porcino, C.; Briglia, M.; Fiorino, E.; Vazzana, M.; Silvestro, S.; Faggio, C. The Influence of Exposure of Cadmium Chloride and Zinc Chloride on Haemolymph and Digestive Gland Cells from Mytilus galloprovincialis. Int. J. Environ. Res. 2017, 11, 207–216. [Google Scholar] [CrossRef]
- Banni, M.; Chouchene, L.; Saïd, K.; Kerkeni, A.; Messaoudi, I. Mechanisms underlying the protective effect of zinc and selenium against cadmium-induced oxidative stress in zebrafish Danio rerio. BioMetals 2011, 24, 981–992. [Google Scholar] [CrossRef]
- Torre, A.; Trischitta, F.; Faggio, C. Effect of CdCl2 on Regulatory Volume Decrease (RVD) in Mytilus galloprovincialis digestive cells. Toxicol. Vitr. 2013, 27, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Curpan, A.-S.; Impellitteri, F.; Plavan, G.; Ciobica, A.; Faggio, C. Review: Mytilus galloprovincialis: An essential, low-cost model organism for the impact of xenobiotics on oxidative stress and public health. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 256, 109302. [Google Scholar] [CrossRef] [PubMed]
- Slobodskova, V.V.; Solodova, E.E.; Slinko, E.N.; Chelomin, V.P. Evaluation of the genotoxicity of cadmium in gill cells of the clam Corbicula japonica using the comet assay. Russ. J. Mar. Biol. 2010, 36, 311–315. [Google Scholar] [CrossRef]
- DeiviArunachalam, K.; Kuruva, J.K.; Pradhoshini, K.P.; Musthafa, M.S.; Faggio, C. Antioxidant and antigenotoxic potential of Morinda tinctoria Roxb. leaf extract succeeding cadmium exposure in Asian catfish, Pangasius sutchi. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 249, 109149. [Google Scholar] [CrossRef] [PubMed]
- Mohiseni, M.; Sepidnameh, M.; Bagheri, D.; Banaee, M.; Haghi, B.N. Comparative effects of Shirazi thyme and vitamin E on some growth and plasma biochemical changes in common carp (Cyprinus carpio) during cadmium exposure. Aquac. Res. 2017, 48, 4811–4821. [Google Scholar] [CrossRef]
- Banaee, M.; Mohammadipour, S.; Madhani, S. Effects of sublethal concentrations of permethrin on bioaccumulation of cadmium in zebra cichlid (Cichlasoma nigrofasciatum). Toxicol. Environ. Chem. 2015, 97, 200–207. [Google Scholar] [CrossRef]
- Shiry, N.; Derakhshesh, N.; Gholamhosseini, A.; Pouladi, M.; Faggio, C. Heavy Metal Concentrations in Cynoglossus arel (Bloch & Schneider, 1801) and Sediment in the Chabahar Bay, Iran. Int. J. Environ. Res. 2021, 15, 773–784. [Google Scholar] [CrossRef]
- Gholamhosseini, A.; Hosseinzadeh, S.; Soltanian, S.; Banaee, M.; Sureda, A.; Rakhshaninejad, M.; Heidari, A.A.; Anbazpour, H. Effect of dietary supplements of Artemisia dracunculus extract on the haemato-immunological and biochemical response, and growth performance of the rainbow trout (Oncorhynchus mykiss). Aquac. Res. 2021, 52, 2097–2109. [Google Scholar] [CrossRef]
- Soleimany, V.; Banaee, M.; Mohiseni, M.; Hagi, B.N.; Dehmourdi, L.M. Evaluation of pre-clinical safety and toxicology of Althaea officinalis extracts as naturopathic medicine for common carp (Cyprinus carpio). Iran. J. Fish. Sci. 2016, 15, 613–629. [Google Scholar] [CrossRef]
- Sacks, D.B. Carbohydrates. In Tietz Textbook of Clinical Chemistry, 3rd ed.; Burtis, C.A., Ashwood, E.R., Eds.; W.B. Saunders Company: Philadelphia, PA, USA, 1999; pp. 766–785. [Google Scholar]
- Rifai, N.; Bachorik, P.S.; Albers, J.J. Lipids, lipoproteins and apolipoproteins. In Tietz Textbook of Clinical Chemistry, 3rd ed.; Burtis, C.A., Ashwood, E.R., Eds.; W.B. Saunders Company: Philadelphia, PA, USA, 1999; pp. 809–861. [Google Scholar]
- Foster-Swanson, A.; Swartzentruber, M.; Roberts, P. Refrence interval studies of the rate-blancked creatinine, Jaffe method on BM/Hitachi Systems in Six U.S. Laboratories. Clin. Chem. 1994, 40, 361. [Google Scholar]
- Johnson, A.M.; Rohlfs, E.M.; Silverman, L.M. Proteins. In Tietz Textbook of Clinical Chemistry, 3rd ed.; Burtis, C.A., Ashwood, E.R., Eds.; W.B. Saunders Company: Philadelphia, PA, USA, 1999; pp. 477–540. [Google Scholar]
- Moss, D.V.; Henderson, A.R. Clinical enzymology. In Tietz Textbook of Clinical Chemistry, 3rd ed.; Burtis, C.A., Ashwood, E.R., Eds.; W.B. Saunders Company: Philadelphia, PA, USA, 1999; pp. 617–677. [Google Scholar]
- Chuiko, G.M. Comparative study of acetylcholinesterase and butyrylcholinesterase in brain and serum of several freshwater fish: Specific activities and in vitro inhibition by DDVP, an organophosphorus pesticide. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 2000, 127, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Banaee, M.; Soltanian, S.; Sureda, A.; Gholamhosseini, A.; Haghi, B.N.; Akhlaghi, M.; Derikvandy, A. Evaluation of single and combined effects of cadmium and micro-plastic particles on biochemical and immunological parameters of common carp (Cyprinus carpio). Chemosphere 2019, 236, 124335. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Placer, Z.A.; Cushman, L.L.; Johnson, B.C. Estimation of product of lipid peroxidation (malonyl dialdehyde) in biochemical systems. Anal. Biochem. 1996, 16, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Góth, L. A Simple Method for Determination of Serum Catalase Activity and Revision of Reference Range. Clin. Chim. Acta 1991, 196, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Derikvandy, A.; Pourkhabbaz, H.R.; Banaee, M.; Sureda, A.; Haghi, N. Genotoxicity and oxidative damage in zebrafish (Danio rerio) after exposure to effluent from ethyl alcohol industry. Chemosphere 2020, 251, 126609. [Google Scholar] [CrossRef]
- Capoluongo, E.; Giardina, B.; Minucci, A. Glucose-6-Phosphate Dehydrogenase (G6PD) Deficiency. In Brenner’s Encyclopedia of Genetics, 2nd ed.; Stanley Maloy, K.H., Ed.; Elsevier/Academic Press: London, UK, 2013; pp. 340–342. [Google Scholar] [CrossRef]
- Banaee, M.; Sureda, A.; Taheri, S.; Hedayatzadeh, F. Sub-lethal effects of dimethoate alone and in combination with cadmium on biochemical parameters in freshwater snail, Galba truncatula. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 220, 62–70. [Google Scholar] [CrossRef]
- Cicik, B.; Engin, K. The Effects of Cadmium on Levels of Glucose in Serum and Glycogen Reserves in the Liver and Muscle Tissues of Cyprinus carpio (L., 1758). Turk. J. Vet. Anim Sci. 2005, 29, 113–117. [Google Scholar]
- Hussein, S.Y.; Mekkawy, I.A.A. The effect of lead-exposure and lead-clay interaction on the growth performance, biochemical and physiological characteristics and histopathology of Tilapia zillii. Bull. Fac. Sci. Assiut Univ. 2001, 30, 65–97. [Google Scholar]
- Shaikh, Z.A.; Vu, T.T.; Zaman, K. Oxidative Stress as a Mechanism of Chronic Cadmium-Induced Hepatotoxicity and Renal Toxicity and Protection by Antioxidants. Toxicol. Appl. Pharmacol. 1999, 154, 256–263. [Google Scholar] [CrossRef]
- Mekkawy, I.A.A.; Lashein, F.E. The effect of lead and cadmium on LDH and G-6-PDH isozyme patterns exhibited during the early embryonic development of the teleost fish, Ctenopharyngodon idellus with emphasis on the corresponding morphological variations. In Proceedings of the 26th Annual Larval Fish Conference (LFC2002), Bergen, Norway, 22–26 July 2002; pp. 275–292. [Google Scholar]
- Karadeniz, A.; Cemek, M.; Simsek, N. The effects of Panax ginseng and Spirulina platensis on hepatotoxicity induced by cadmium in rats. Ecotoxicol. Environ. Saf. 2009, 72, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Mekkawy, I.A.A.; Mahmoud, U.M.; Wassif, E.T.; Naguib, M. Effects of cadmium on some haematological and biochemical characteristics of Oreochromis niloticus (Linnaeus, 1758) dietary supplemented with tomato paste and vitamin E. Fish Physiol. Biochem. 2010, 37, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Philippe, C.; Grégoir, A.F.; Thoré, E.S.J.; Brendonck, L.; De Boeck, G.; Pinceel, T. Acute sensitivity of the killifish Nothobranchius furzeri to a combination of temperature and reference toxicants (cadmium, chlorpyrifos and 3,4-dichloroaniline). Environ. Sci. Pollut. Res. 2018, 25, 10029–10038. [Google Scholar] [CrossRef]
- Philippe, C.; Hautekiet, P.; Grégoir, A.F.; Thoré, E.S.J.; Pinceel, T.; Stoks, R.; Brendonck, L.; De Boeck, G. Combined effects of cadmium exposure and temperature on the annual killifish (Nothobranchius furzeri). Environ. Toxicol. Chem. 2018, 37, 2361–2371. [Google Scholar] [CrossRef]
- Jyoti, D.; Sinha, R.; Faggio, C. Advances in biological methods for the sequestration of heavy metals from water bodies: A review. Environ. Toxicol. Pharmacol. 2022, 94, 103927. [Google Scholar] [CrossRef] [PubMed]
- Shahjahan, M.; Taslima, K.; Rahman, M.S.; Emran, A.; Alam, S.I.; Faggio, C. Effects of heavy metals on fish physiology—A review. Chemosphere 2022, 300, 134519. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, Y.; Ikeda, H.; Haramaki, N.; Yoshida, N.; Imaizumi, T. Oxidative stress is related to exercise intolerance in patients with heart failure. Am. Hear. J. 1998, 135, 115–120. [Google Scholar] [CrossRef]
- Al-Asgah, N.A.; Abdel-Warith, A.-W.A.; Younis, E.-S.M.; Allam, H.Y. Haematological and biochemical parameters and tissue accumulations of cadmium in Oreochromis niloticus exposed to various concentrations of cadmium chloride. Saudi J. Biol. Sci. 2015, 22, 543–550. [Google Scholar] [CrossRef]
- Üner, N.; Oruç, E.; Sevgiler, Y.; Şahin, N.; Durmaz, H.; Usta, D. Effects of diazinon on acetylcholinesterase activity and lipid peroxidation in the brain of Oreochromis niloticus. Environ. Toxicol. Pharmacol. 2006, 21, 241–245. [Google Scholar] [CrossRef]
- Heydarnejad, M.S.; Khosravian-Hemamai, M.; Nematollahi, A. Effects of cadmium at sub-lethal concentration on growth and biochemical parameters in rainbow trout (Oncorhynchus mykiss). Ir. Vet. J. 2013, 66, 11. [Google Scholar] [CrossRef]
- Abdelkhalek, N.K.M.; Eissa, I.A.M.; Ahmed, E.; Kilany, O.E.; El-Adl, M.; Dawood, M.A.; Hassan, A.M.; Abdel-Daim, M.M. Protective role of dietary Spirulina platensis against diazinon-induced Oxidative damage in Nile tilapia; Oreochromis niloticus. Environ. Toxicol. Pharmacol. 2017, 54, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Levesque, H.M.; Moon, T.W.; Campbell, P.G.C.; Hontela, A. Seasonal variation in carbohydrate and lipid metabolism of yellow perch (Perca flavescens) chronically exposed to metals in the field. Aquat. Toxicol. 2002, 60, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Wooster, G.A.; Bowser, P.R. Comparative blood chemistry and histopathology of tilapia infected with Vibrio vulnificus or Streptococcus iniae or exposed to carbon tetrachloride, gentamicin, or copper sulfate. Aquaculture 2004, 239, 421–443. [Google Scholar] [CrossRef]
- Shalaby, A.M.E. Effect of EDTA on Toxicity Reduction of Cadmium in Relation to Growth, Some Haematological and Biochemical Profiles of Nile Tilapia (Oreochromis niloticus). J. Fish. Aquat. Sci. 2007, 2, 100–109. [Google Scholar] [CrossRef]
- Banaee, M.; Akhlaghi, M.; Soltanian, S.; Sureda, A.; GholamHosseini, A.; Rakhshaninejad, M. Combined effects of exposure to sub-lethal concentration of the insecticide chlorpyrifos and the herbicide glyphosate on the biochemical changes in the freshwater crayfish Pontastacus leptodactylus. Ecotoxicology 2020, 29, 1500–1515. [Google Scholar] [CrossRef]
- Banaei, M.; Forouzanfar, M.; Jafarinia, M. Toxic effects of polyethylene microplastics on transcriptional changes, biochemical response, and oxidative stress in common carp (Cyprinus carpio). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 261, 109423. [Google Scholar] [CrossRef]
- Banaee, M.; Tahery, S.; Haghi, B.N.; Shahafve, S.; Vaziriyan, M. Blood biochemical changes in common carp (Cyprinus carpio) upon co-exposure to titanium dioxide nanoparticles and paraquat. Iran. J. Fish. Sci. 2019, 18, 242–255. [Google Scholar] [CrossRef]
- Hatami, M.; Banaee, M.; Haghi, B.N. Sub-lethal toxicity of chlorpyrifos alone and in combination with polyethylene glycol to common carp (Cyprinus carpio). Chemosphere 2019, 219, 981–988. [Google Scholar] [CrossRef]
- Han, S.-H.; Park, J.-C.; Byun, M.S.; Yi, D.; Lee, J.H.; Lee, D.Y.; Mook-Jung, I. Blood acetylcholinesterase level is a potential biomarker for the early detection of cerebral amyloid deposition in cognitively normal individuals. Neurobiol. Aging 2019, 73, 21–29. [Google Scholar] [CrossRef]
- Perrault, J.R.; Bauman, K.D.; Greenan, T.M.; Blum, P.C.; Henry, M.S.; Walsh, C.J. Maternal transfer and sublethal immune system effects of brevetoxin exposure in nesting loggerhead sea turtles (Caretta caretta) from western Florida. Aquat. Toxicol. 2016, 180, 131–140. [Google Scholar] [CrossRef]
- Atli, G.; Canli, M. Enzymatic responses to metal exposures in a freshwater fish Oreochromis niloticus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 145, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Faheem, M.; Qayyum, A.; Sulehria, K.; Tariq, M.; Khadjia, I.; Fiaz, A.; Saeed, M. Effect of sublethal dose of CdCl2 on biochemical profile and catalase activity in fresh water fish Oreochromis niloticus. Environ. Sci. 2012, 58, 73–79. [Google Scholar]
- Cao, L.; Huang, W.; Liu, J.; Yin, X.; Dou, S. Accumulation and oxidative stress biomarkers in Japanese flounder larvae and juveniles under chronic cadmium exposure. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2010, 151, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Umamaheswari, S.; Priyadarshinee, S.; Bhattacharjee, M.; Kadirvelu, K.; Ramesh, M. Exposure to polystyrene microplastics induced gene modulated biological responses in zebrafish (Danio rerio). Chemosphere 2021, 281, 128592. [Google Scholar] [CrossRef] [PubMed]
- Song, J.A.; Choi, C.Y.; Park, H.-S. Exposure of bay scallop Argopecten irradians to micro-polystyrene: Bioaccumulation and toxicity. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 236, 108801. [Google Scholar] [CrossRef]
- Messaoudi, I.; Barhoumi, S.; Saïd, K.; Kerken, A. Study on the sensitivity to cadmium of marine fish Salaria basilisca (Pisces: Blennidae). J. Environ. Sci. 2009, 21, 1620–1624. [Google Scholar] [CrossRef]
- Ibrahim, A.T.A.; Banaee, M.; Sureda, A. Genotoxicity, oxidative stress, and biochemical biomarkers of exposure to green synthesized cadmium nanoparticles in Oreochromis niloticus (L.). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 242, 108942. [Google Scholar] [CrossRef]
- Solomando, A.; Capó, X.; Alomar, C.; Álvarez, E.; Compa, M.; Valencia, J.M.; Pinya, S.; Deudero, S.; Sureda, A. Long-term exposure to microplastics induces oxidative stress and a pro-inflammatory response in the gut of Sparus aurata Linnaeus, 1758. Environ. Pollut. 2020, 266, 115295. [Google Scholar] [CrossRef]
- Li, Z.; Chang, X.; Hu, M.; Fang, J.K.-H.; Sokolova, I.M.; Huang, W.; Xu, E.G.; Wang, Y. Is microplastic an oxidative stressor? Evidence from a meta-analysis on bivalves. J. Hazard. Mater. 2021, 423, 127211. [Google Scholar] [CrossRef]
- Hamidi, S.; Banaee, M.; Pourkhabbaz, H.R.; Sureda, A.; Khodadoust, S. Effect of petroleum wastewater treated with gravity separation and magnetite nanoparticles adsorption methods on the blood biochemical response of mrigal fish (Cirrhinus cirrhosus). Environ. Sci. Pollut. Res. 2021, 29, 3718–3732. [Google Scholar] [CrossRef]
- Banihashemi, E.A.; Soltanian, S.; Gholamhosseini, A.; Banaee, M. Effect of microplastics on Yersinia ruckeri infection in rainbow trout (Oncorhynchus mykiss). Environ. Sci. Pollut. Res. 2022, 29, 11939–11950. [Google Scholar] [CrossRef] [PubMed]
- Fırat, Ö.; Çogun, H.Y.; Aslanyavrusu, S.; Kargın, F. Antioxidant responses and metal accumulation in tissues of Nile tilapia Oreochromis niloticus under Zn, Cd and Zn + Cd exposures. J. Appl. Toxicol. 2009, 29, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Scully, C. Metabolic disorders. In Scully’s Medical Problems in Dentistry, 7th ed.; Elsevier/Churchill Livingstone: London, UK, 2014; pp. 594–606. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S. The Role of Spirulina (Arthrospira) in the Mitigation of Heavy-Metal Toxicity: An Appraisal. J. Environ. Pathol. Toxicol. Oncol. 2020, 39, 149–157. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).