Changes in the Bacterial Diversity of Human Milk during Late Lactation Period (Weeks 21 to 48)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction
2.3. DNA Sequencing and Analysis
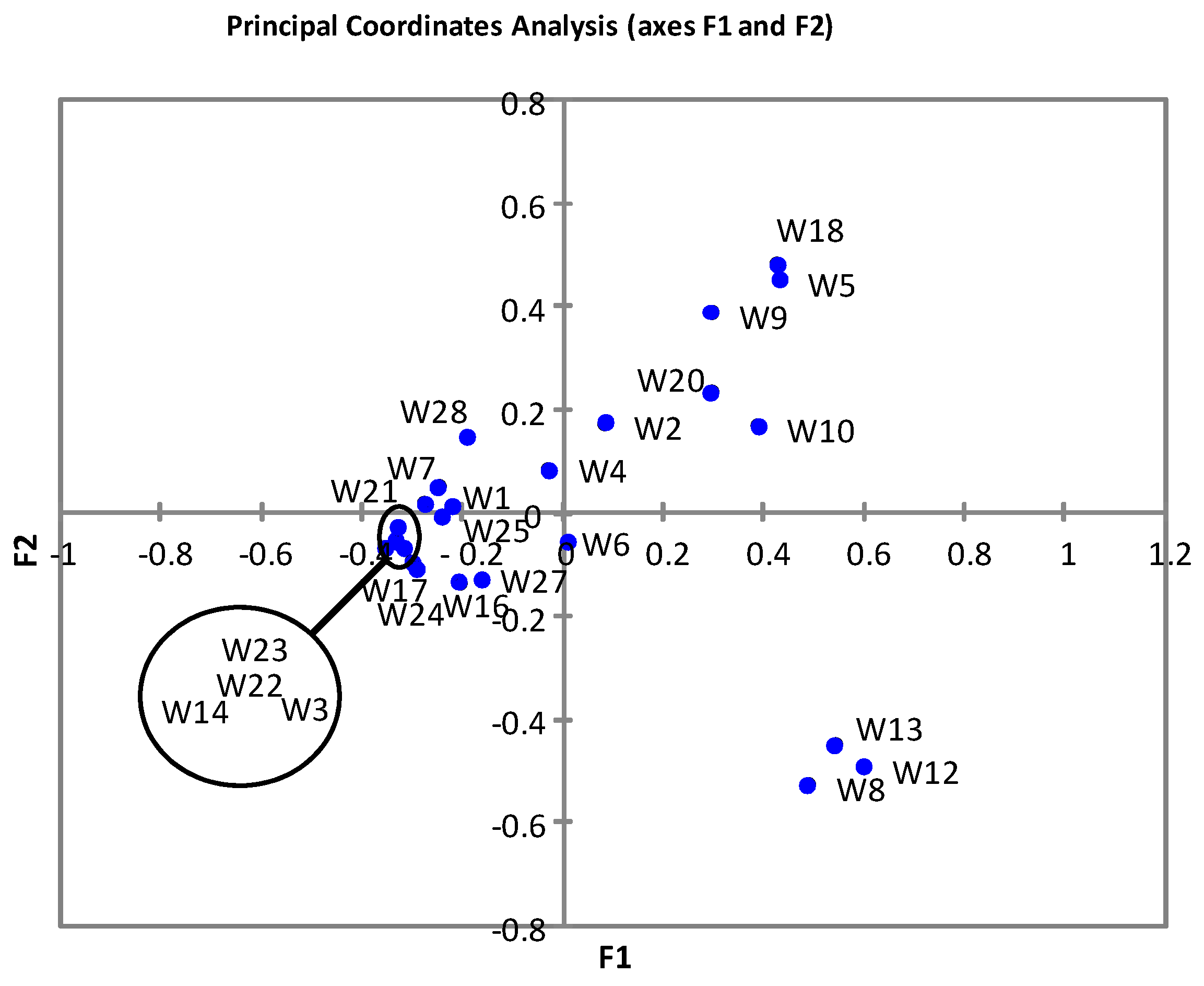
3. Results
3.1. Characteristics of Sequence Reads
3.2. Bacterial Diversity in Breast Milk Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heikkila, M.P.; Saris, P.E. Inhibition of Staphylococcus aureus by the commensal bacteria of human milk. J. Appl. Microbiol. 2003, 95, 471–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asan-Ozusaglam, M.; Gunyakti, A. Lactobacillus fermentum strains from human breast milk with probiotic properties and cholesterol-lowering effects. Food Sci. Biotechnol. 2018, 28, 501–509. [Google Scholar] [CrossRef]
- Cabrera-Rubio, R.; Mira-Pascual, L.; Mira, A.; Collado, M.C. Impact of mode of delivery on the milk microbiota composition of healthy women. J. Dev. Orig. Health 2016, 7, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Jost, T.; Lacroix, C.; Braegger, C.; Chassard, C. Impact of human milk bacteria and oligosaccharides on neonatal gut microbiota establishment and gut health. Nutr. Rev. 2015, 73, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.; Langa, S.; Martín, V.; Maldonado, A.; Jiménez, E.; Martín, R.; Rodríguez, J.M. The human milk microbiota: Origin and potential roles in health and disease. Pharmacol. Res. 2013, 69, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.; Rodríguez, J.M. Human milk microbiota: Origin and potential uses. Nestle Nutr. Inst. Workshop Ser. 2020, 94, 75–85. [Google Scholar] [CrossRef]
- Pannaraj, P.S.; Li, F.; Cerini, C.; Bender, J.M.; Yang, S.; Rollie, A.; Adisetiyo, H.; Zabih, S.; Lincez, P.J.; Bittinger, K.; et al. Association between breast milk bacterial communities and establishment and development of the infant gut. JAMA Pediatr. 2017, 171, 647–654. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Pang, B.; Liu, G.; Zhao, X.; Xu, X.; Jiang, C.; Yang, B.; Liu, Y.; Shi, J. Lactobacillus rhamnosus from human breast milk shows therapeutic function against foodborne infection by multi-drug resistant Escherichia coli in mice. Food Funct. 2020, 11, 435–447. [Google Scholar] [CrossRef]
- Fitzstevens, J.L.; Smith, K.C.; Hagadorn, J.I.; Caimano, M.J.; Matson, A.P.; Brownell, E.A. Systematic review of the human milk microbiota. Nutr. Clin. Pract. 2017, 32, 354–364. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. Breast milk microbiota: A review of the factors that influence composition. J. Infect. 2020, 81, 17–47. [Google Scholar] [CrossRef]
- Cabrera-Rubio, R.; Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E.; Mira, A. The human milk microbiome changes over lactation and is shaped by maternal weight and mode of delivery. Am. J. Clin. Nutr. 2012, 96, 544–551. [Google Scholar] [CrossRef] [Green Version]
- Khodayar-Pardo, P.; Mira-Pascual, L.; Collado, M.C.; Martínez-Costa, C. Impact of lactation stage, gestational age and mode of delivery on breast milk microbiota. J. Perinatol. 2014, 34, 599–605. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Mashima, I.; Theodorea, C.F.; Thaweboon, B.; Thaweboon, S.; Scannapieco, F.A.; Nakazawa, F. Exploring the salivary microbiome of children stratified by the oral hygiene index. PLoS ONE 2017, 12, e0185274. [Google Scholar] [CrossRef]
- Makino, H.; Kushiro, A.; Ishikawa, E.; Kubota, H.; Gawad, A.; Sakai, T.; Oishi, K.; Martin, R.; Ben-Amor, K.; Knol, J.; et al. Mother-to-infant transmission of intestinal bifidobacterial strains has an impact on the early development of vaginally delivered infant’s microbiota. PLoS ONE 2013, 8, e78331. [Google Scholar] [CrossRef]
- Rodríguez, J.M. The origin of human milk bacteria: Is there a bacterial entero-mammary pathway during late pregnancy and lactation? Adv. Nutr. 2014, 5, 779–784. [Google Scholar] [CrossRef] [Green Version]
- Boix-Amoros, A.; Collado, M.C.; Mira, A. Relationship between milk microbiota, bacterial load, macronutrients, and human cells during lactation. Front. Microbiol. 2016, 7, 492. [Google Scholar] [CrossRef] [Green Version]
- Wescombe, P.A.; Heng, N.C.; Burton, J.P.; Chilcott, C.N.; Tagg, J.R. Streptococcal bacteriocins and the case for Streptococcus salivarius as model oral probiotics. Future Microbiol. 2009, 4, 819–835. [Google Scholar] [CrossRef] [Green Version]
- Herrero, E.R.; Slomka, V.; Bernaerts, K.; Boon, N.; Hernandez-Sanabria, E.; Passonial, B.B.; Quirynen, M.; Teughels, W. Antimicrobial effects of commensal oral species are regulated by environmental factors. J. Dent. 2016, 47, 23–33. [Google Scholar] [CrossRef]
- Park, B.; Iwase, T.; Liu, G.Y. Intranasal application of S. epidermidis prevents colonization by methicillin-resistant Staphylococcus aureus in mice. PLoS ONE 2011, 6, e25880. [Google Scholar] [CrossRef] [Green Version]
- Ding, M.; Qi, C.; Yang, Z.; Jiang, S.; Bi, Y.; Lai, J.; Sun, J. Geographical location specific composition of cultured microbiota and Lactobacillus occurrence in human breast milk in China. Food Funct. 2019, 10, 554–564. [Google Scholar] [CrossRef]
- Dahaban, N.M.; Romli, M.F.; Roslan, N.R.; Kong, S.S.; Cheah, F.C. Bacteria in expressed breastmilk from mothers of premature infants and maternal hygienic status. Breastfeed. Med. 2013, 8, 422–423. [Google Scholar] [CrossRef]
- Cacho, N.T.; Harrison, N.A.; Parker, L.A.; Padgett, K.A.; Lemas, D.J.; Marcial, G.E.; Li, N.; Carr, L.E.; Neu, J.; Lorca, G.L. Personalization of the microbiota of donor human milk with mother’s own milk. Front. Microbiol. 2017, 8, 1470. [Google Scholar] [CrossRef]
- Patel, S.H.; Vaidya, Y.H.; Patel, R.J.; Pandit, R.J.; Joshi, C.G.; Kunjadiya, A.P. Culture independent assessment of human milk microbial community in lactational mastitis. Sci. Rep. 2017, 7, 7804. [Google Scholar] [CrossRef] [Green Version]
- Rigourd, V.; Barnier, J.P.; Ferroni, A.; Nicloux, M.; Hachem, T.; Magny, J.F.; Lapillonne, A.; Frange, P.; Nassif, X.; Bille, E. Recent actuality about Bacillus cereus and human milk bank: A new sensitive method for microbiological analysis of pasteurized milk. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1297–1303. [Google Scholar] [CrossRef]
- Moro, G.E.; Billeaud, C.; Rachel, B.; Calvo, J.; Cavallarin, L.; Christen, L.; Escuder-Vieco, D.; Gaya, A.; Lembo, D.; Wesolowska, A.; et al. Processing of donor human milk: Update and recommendations from the European Milk Bank Association (EMBA). Front. Pediatr. 2019, 7, 49. [Google Scholar] [CrossRef] [Green Version]
- Lewin, A.; Delage, G.; Bernier, F.; Germain, M. Banked human milk and quantitative risk assessment of Bacillus cereus infection in premature infants: A simulation study. Can. J. Infect. Dis. Med. Microbiol. 2019, 6348281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, N.K.; Kim, W.S.; Paik, H.D. Bacillus strains as human probiotics: Characterization, safety, microbiome, and probiotic carrier. Food Sci. Biotechnol. 2019, 28, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- US Food Drug Administration. GRAS Notice (GRN) No. 660. Notice to US Food and Drug Administration that Bacillus coagulans GBI-30, 6086 is Generally Recognized as Safe for Use in Non-Exempt Term Infant Formula. 2016. Available online: https://www.fda.gov/media/100025/download (accessed on 16 July 2020).
- Guo, Q.; Goldenberg, J.Z.; Humphrey, C.; El Dib, R.; Johnston, B.C. Probiotics for the prevention of pediatric antibiotic-associated diarrhea. Cochrane Database Syst. Rev. 2019, CD004827. [Google Scholar] [CrossRef] [PubMed]
- Van den Akker, C.H.P.; van Goudoever, J.B.; Szajewska, H.; Embleton, N.D.; Hojsak, I.; Reid, D.; Shamir, R. ESPGHAN Working Group for Probiotics, Prebiotics & Committee on Nutrition. Probiotics for preterm infants: A strain-specific systematic review and network meta-analysis. J. Pediatr. Gastroenterol. Nutr. 2018, 67, 103–122. [Google Scholar] [CrossRef] [PubMed]
- Boo, N.Y.; Nordiah, A.J.; Alfizah, H.; Nor-Rohaini, A.H.; Lim, V.K. Contamination of breast milk obtained by manual expression and breast pumps in mothers of very low birthweight infants. J. Hosp. Infect. 2001, 49, 274–281. [Google Scholar] [CrossRef]
- Eisenhofer, R.; Minich, J.J.; Marotz, C.; Cooper, A.; Knight, R.; Weyrich, L.A. Contamination in low microbial biomass microbiome studies: Issues and recommendations. Trends Microbiol. 2019, 27, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Salter, S.J.; Cox, M.J.; Turek, E.M.; Calus, S.T.; Cookson, W.O.; Moffatt, M.F.; Turner, P.; Parkhill, J.; Loman, N.J.; Walker, A.W. Reagent and laboratory contamination can critically impact sequence-based microbiome analyses. BMC Biol. 2014, 12, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | N° Reads | N° Observations | Chao1 | Shannon-Weaver | Simpson |
|---|---|---|---|---|---|
| W1 | 111,968.00 | 107.00 | 107.00 | 2.90 | 0.89 |
| W2 | 64,302.00 | 91.00 | 91.0 | 3.32 | 0.94 |
| W3 | 106,190.00 | 95.00 | 95.00 | 1.60 | 0.52 |
| W4 | 125,198.00 | 165.00 | 165.00 | 3.37 | 0.93 |
| W5 | 145,841.00 | 115.00 | 115.00 | 1.87 | 0.70 |
| W6 | 132,594.00 | 71.00 | 71.00 | 2.08 | 0.83 |
| W7 | 145,004.00 | 163.00 | 163.00 | 2.89 | 0.84 |
| W8 | 122,052.00 | 65.00 | 65.00 | 1.24 | 0.47 |
| W9 | 151,193.00 | 110.00 | 110.00 | 2.94 | 0.89 |
| W10 | 86,107.00 | 140.00 | 140.00 | 3.49 | 0.94 |
| W11 | 3245.00 | 9.00 | 9.00 | 1.34 | 0.66 |
| W12 | 55,527.00 | 65.00 | 65.00 | 2.02 | 0.78 |
| W13 | 138,427.00 | 118.00 | 118.00 | 2.27 | 0.81 |
| W14 | 141,003.00 | 93.00 | 93.00 | 1.86 | 0.65 |
| W15 | 143.00 | 3.00 | 3.00 | 0.32 | 0.14 |
| W16 | 102,600.00 | 102.00 | 102.00 | 2.45 | 0.80 |
| W17 | 98,764.00 | 81.00 | 81.00 | 2.00 | 0.68 |
| W18 | 107,346.00 | 1370.00 | 130.00 | 3.31 | 0.92 |
| W19 | 140.00 | 3.00 | 3.00 | 0.62 | 0.37 |
| W20 | 68,575.00 | 90.00 | 90.00 | 2.69 | 0.85 |
| W21 | 1,375,641.00 | 119.00 | 119.00 | 1.92 | 0.68 |
| W22 | 89,705.00 | 92.00 | 92.00 | 1.65 | 0.58 |
| W23 | 116,236.00 | 104.00 | 104.00 | 1.82 | 0.62 |
| W24 | 100,113.00 | 102.00 | 102.00 | 2.34 | 0.76 |
| W25 | 97,925.00 | 109.00 | 109.00 | 2.52 | 0.86 |
| W26 | 61,413.00 | 6.00 | 6.00 | 0.01 | 0.00 |
| W27 | 105,221.00 | 119.00 | 119.00 | 2.72 | 0.87 |
| W28 | 36,336.00 | 111.00 | 111.00 | 2.77 | 0.81 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marin-Gómez, W.; Grande, M.J.; Pérez-Pulido, R.; Galvez, A.; Lucas, R. Changes in the Bacterial Diversity of Human Milk during Late Lactation Period (Weeks 21 to 48). Foods 2020, 9, 1184. https://doi.org/10.3390/foods9091184
Marin-Gómez W, Grande MJ, Pérez-Pulido R, Galvez A, Lucas R. Changes in the Bacterial Diversity of Human Milk during Late Lactation Period (Weeks 21 to 48). Foods. 2020; 9(9):1184. https://doi.org/10.3390/foods9091184
Chicago/Turabian StyleMarin-Gómez, Wendy, Mᵃ José Grande, Rubén Pérez-Pulido, Antonio Galvez, and Rosario Lucas. 2020. "Changes in the Bacterial Diversity of Human Milk during Late Lactation Period (Weeks 21 to 48)" Foods 9, no. 9: 1184. https://doi.org/10.3390/foods9091184