Properties of Peanut (KAC431) Protein Hydrolysates and Their Impact on the Quality of Gluten-Free Rice Bread

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Extraction of Peanut Proteins
2.2.2. Analysis of Proximate Composition
2.2.3. Analysis of Amino Acid Profiles
2.2.4. Production of Protein Hydrolysates (PHs)
2.2.5. Chemical and Physical Properties
Protein Patterns
Antioxidant Properties
Foaming and Emulsifying Properties
2.2.6. Gluten-Free Bread Preparation
2.2.7. Bread Quality Determinations
Specific Volume and Weight Loss
Crumb Firmness and Relative Elasticity
2.2.8. Statistical Analysis
3. Results and Discussion
3.1. Proximate Composition and Protein Extraction Yield
3.2. Amino Acid Profiles
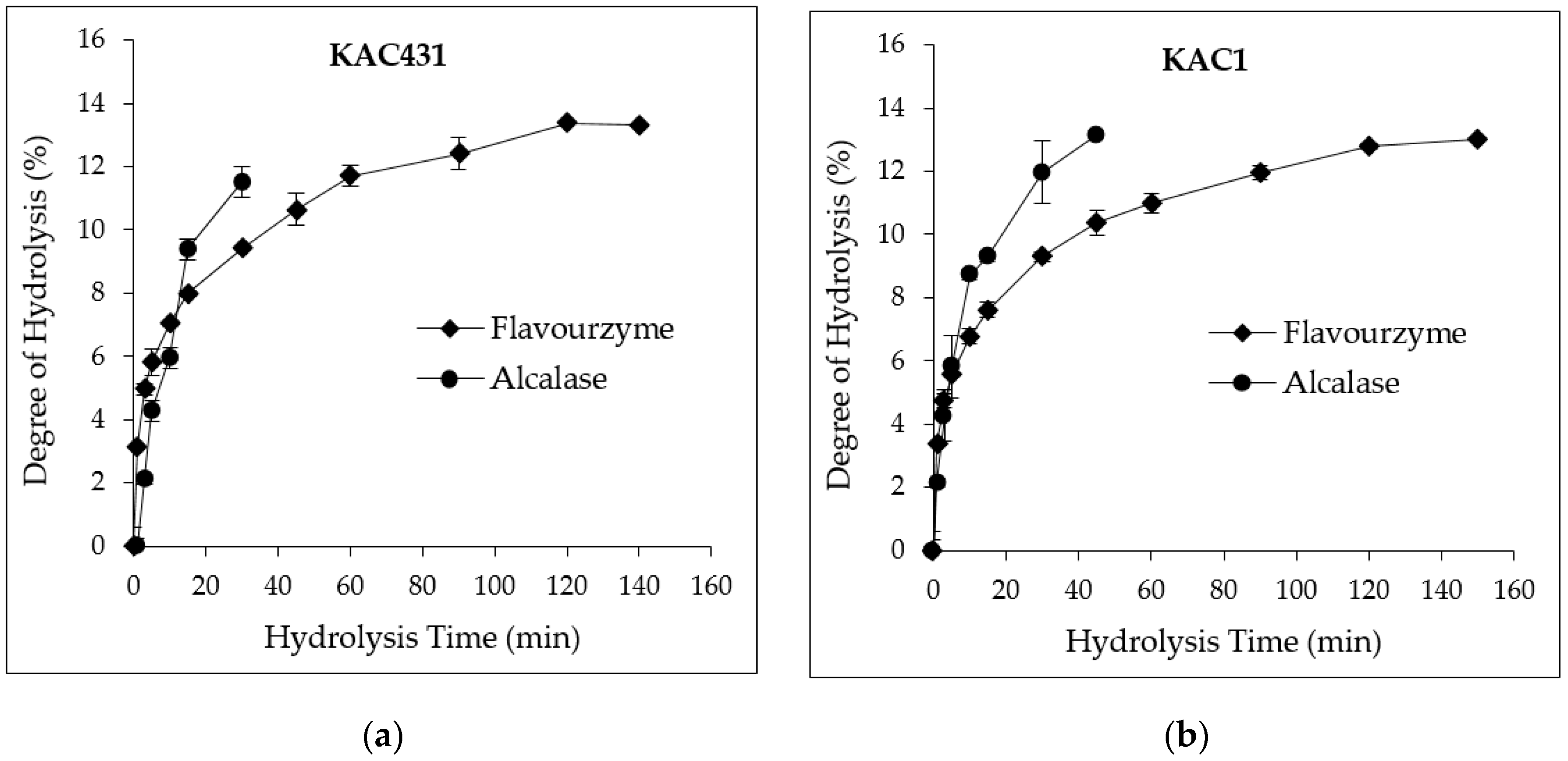
3.3. DH and PH Yields
3.4. Properties of PHs
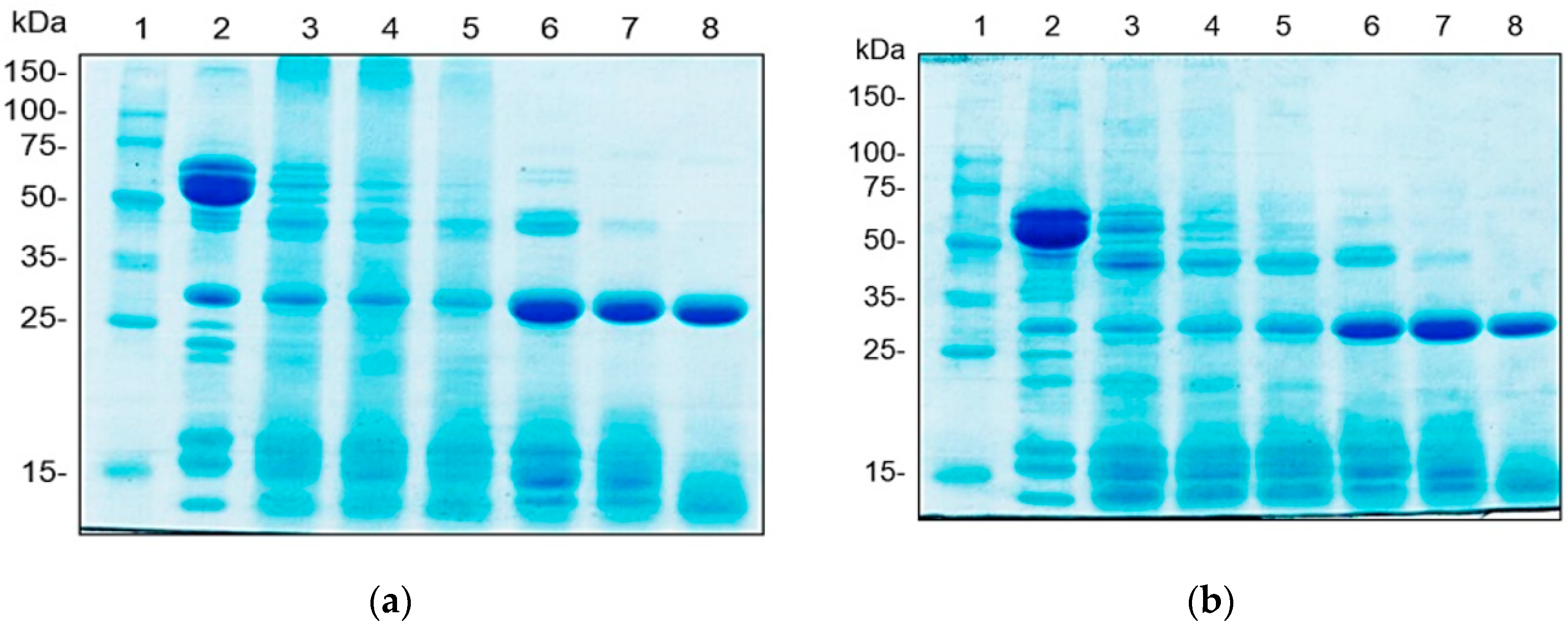
3.4.1. Protein Patterns
3.4.2. Antioxidant Properties
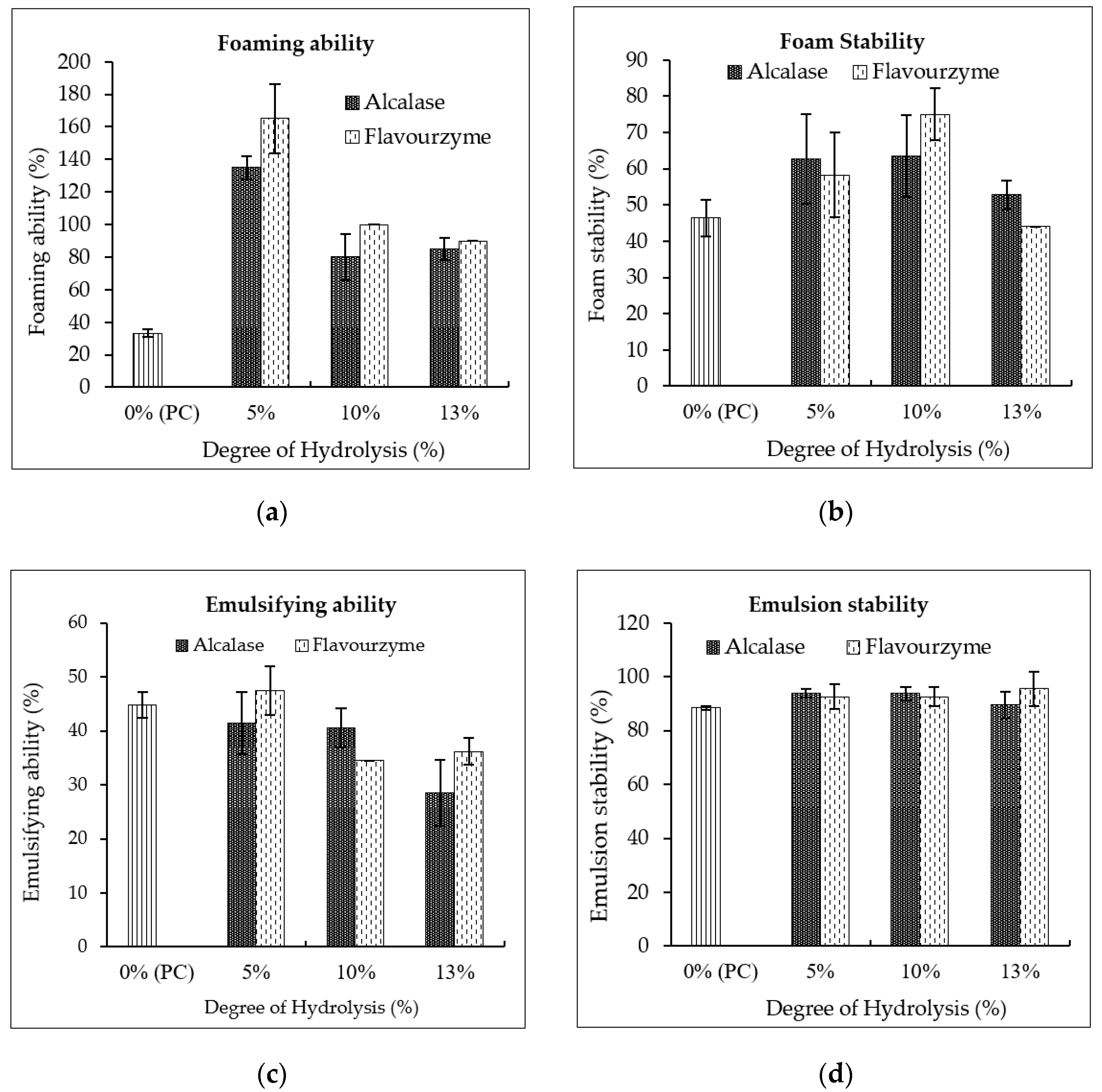
3.4.3. Functional Properties
3.5. Effect of PHs on Gluten-Free Bread Qualities
3.5.1. Weight loss and Specific Volume
3.5.2. Crumb Firmness and Relative Elasticity
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gutowski, E.D.; Weiten, D.; Green, K.H.; Rigaux, L.N.; Bernstein, C.N.; Graff, L.A.; Walker, J.R.; Duerksen, D.R.; Silvester, J.A. Can individuals with celiac disease identify gluten-free foods correctly? Clin. Nutr. ESPEN 2020, 36, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Han, A.; Romero, H.M.; Nishijima, N.; Ichimura, T.; Handa, A.; Xu, C.; Zhang, Y. Effect of egg white solids on the rheological properties and bread making performance of gluten-free batter. Food Hydrocoll. 2019, 87, 287–296. [Google Scholar] [CrossRef]
- Kiumarsi, M.; Shahbazi, M.; Yeganehzad, S.; Majchrzak, D.; Lieleg, O.; Winkeljann, B. Relation between structural, mechanical and sensory properties of gluten-free bread as affected by modified dietary fibers. Food Chem. 2019, 277, 664–673. [Google Scholar] [CrossRef]
- Chakrabortya, S.K.; Kotwaliwalea, N.; Navale, S.A. Selection and incorporation of hydrocolloid for gluten-free leavened millet breads and optimization of the baking process thereof. LWT—Food Sci. Technol. 2020, 119, 108878. [Google Scholar] [CrossRef]
- Phongthai, S.; D’Amico, S.; Schoenlechner, R.; Rawdkuen, S. Comparative study of rice bran protein concentrate and egg albumin on gluten-free bread properties. J. Cereal Sci. 2016, 72, 38–45. [Google Scholar] [CrossRef]
- Phongthai, S.; Lim, S.T.; Rawdkuen, S. Optimization of ultrasonic-assisted extraction of rice bran protein using response surface methodology. J. Food Biochem. 2016, 41, e12314. [Google Scholar] [CrossRef]
- Zheng, Z.; Li, J.; Li, J.; Sun, H.; Liu, Y. Physicochemical and antioxidative characteristics of black bean protein hydrolysates obtained from different enzymes. Food Hydrocoll. 2019, 97, 105222. [Google Scholar] [CrossRef]
- Liang, G.; Chen, W.; Qie, X.; Zeng, M.; Qin, F.; He, Z.; Chen, J. Modification of soy protein isolates using combined pre-heat treatment and controlled enzymatic hydrolysis for improving foaming properties. Food Hydrocoll. 2020, 105, 105764. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 17th ed.; AOAC: Arlington, VA, USA, 2000. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Phongthai, S.; Rawdkuen, S. Purification and characterization of antioxidant peptides from rice bran protein hydrolysate stimulated by in vitro gastrointestinal digestion. Cereal Chem. 2019, 97, 316–325. [Google Scholar] [CrossRef]
- AACC. Approved Methods of the American Association of Cereal Chemists; AACC: St. Paul, MN, USA, 2000. [Google Scholar]
- Torruco-Uco, J.; Chel-Guerrero, L.; Martı’nez-Ayala, A.; Da’vila-Ortı´z, G.; Betancur-Ancona, D. Angiotensin-I converting enzyme inhibitory and antioxidant activities of protein hydrolysates from Phaseolus lunatus and Phaseolus vulgaris seeds. LWT—Food Sci. Technol. 2009, 42, 1597–1604. [Google Scholar] [CrossRef]
- WHO (World Health Organization). Protein and Amino Acid Requirements in Human Nutrition: Report of a Joint FAO/WHO/UNU Expert Consultation. Available online: https://apps.who.int/iris/handle/10665/43411 (accessed on 10 June 2020).
- Li, P.; Jia, J.; Fang, M.; Zhang, L.; Guo, M.; Xie, J.; Xia, Y.; Zhou, L.; Wei, D. In vitro and in vivo ACE inhibitory of pistachio hydrolysates and in silico mechanism of identified peptide binding with ACE. Process Biochem. 2014, 49, 898–904. [Google Scholar] [CrossRef]
- Phongthai, S.; D’Amico, S.; Schoenlechner, R.; Rawdkuen, S. Fractionation and antioxidant properties of rice bran protein hydrolysates stimulated by in vitro gastrointestinal digestion. Food Chem. 2018, 240, 156–164. [Google Scholar] [CrossRef]
- Bamdad, F.; Wu, J.; Chen, L. Effects of enzymatic hydrolysis on molecular structure and antioxidant activity of barley hordein. J. Cereal Sci. 2011, 54, 20–28. [Google Scholar] [CrossRef]
- Liu, H.; Shi, A.; Liu, L.; Wu, H.; Ma, T.; He, X.; Lin, W.; Feng, X.; Liu, Y. Peanut protein processing technology. In Peanuts, Processing Technology and Product Development; Wang, Q., Ed.; Academic Press: London, UK, 2016; p. 107. [Google Scholar]
- Segura-Campos, M.R.; Espinosa-García, L.; Chel-Guerrero, L.A.; Betancur-Ancona, D.A. Effect of enzymatic hydrolysis on solubility, hydrophobicity, and in vivo digestibility in cowpea (Vigna unguiculata). Int. J. Food Prop. 2012, 15, 770–780. [Google Scholar] [CrossRef]
- Vaštag, Ž.; Popović, L.; Popović, S.; Krimer, V.; Peričin, D. Production of enzymatic hydrolysates with antioxidant and angiotensin-I converting enzyme inhibitory activity from pumpkin oil cake protein isolate. Food Chem. 2011, 124, 1316–1321. [Google Scholar] [CrossRef]
- Moure, A.; Sineiro, J.; Dominguez, H.; Parajo, J.C. Functionality of oilseed protein products: A review. Food Res. Int. 2006, 39, 945–963. [Google Scholar] [CrossRef]
- Guan, X.; Yao, H.; Chen, Z.; Shan, L.; Zhang, M. Some functional properties of oat bran protein concentrate modified by trypsin. Food Chem. 2007, 101, 163–170. [Google Scholar] [CrossRef]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Shahidi, F. Antioxidative activity and functional properties of protein hydrolysate of yellow stripe trevally (Selaroides leptolepis) as influenced by the degree of hydrolysis and enzyme type. Food Chem. 2007, 102, 1317–1327. [Google Scholar] [CrossRef]
- Jamdar, S.N.; Rajalakshmi, V.; Pednekar, M.D.; Juan, F.; Yardi, V.; Sharma, V. Influence of degree of hydrolysis on functional properties, antioxidant activity and ACE inhibitory activity of peanut protein hydrolysate. Food Chem. 2010, 121, 178–184. [Google Scholar] [CrossRef]
- Rodriguez Furlán, L.T.; Padilla, A.P.; Campderrós, M.E. Improvement of gluten-free bread properties by the incorporation of bovine plasma proteins and different saccharides into the matrix. Food Chem. 2015, 170, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Bender, D.; Schönlechner, R. Innovative approaches towards improved gluten-free bread properties. J. Cereal Sci. 2020, 91, 102904. [Google Scholar] [CrossRef]
- Masure, H.G.; Wouters, A.G.B.; Fierens, E.; Delcour, J.A. Impact of egg white and soy proteins on structure formation and crumb firming in gluten-free breads. Food Hydrocoll. 2019, 95, 406–417. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KAC431 | KAC1 | |
|---|---|---|
| Defatted Peanuts | ||
| Protein (g/100 g) | 42.39 ± 0.18 a | 35.16 ± 1.15 b |
| Carbohydrate (g/100 g) | 29.42 ± 0.11 a | 27.03 ± 0.96 b |
| Oil (g/100 g) | 15.01 ± 1.22 b | 18.77 ± 0.51 a |
| Crude fiber (g/100 g) | 5.31 ± 0.82 b | 10.83 ± 1.52 a |
| Ash (g/100 g) | 4.04 ± 0.03 a | 3.46 ± 0.07 b |
| Moisture (g/100 g) | 3.83 ± 0.10 a | 4.75 ± 0.01 b |
| Protein Concentrate | ||
| Extraction yield (%) | 43.26 ± 1.78 a | 37.25 ± 2.39 b |
| Protein content (g/100 g) | 77.76 ± 0.31 b | 74.14 ± 0.22 a |
| Amino Acids | KAC431 | KAC1 | Amino Acids | KAC431 | KAC1 |
|---|---|---|---|---|---|
| Hydrophobic | Electrical Charged | ||||
| Valine | 5007 ± 144 a | 4366 ± 38 b | Histidine | 1736 ± 0.00 a | 1621 ± 0.00 b |
| Isoleucine | 4244 ± 157 a | 3816 ± 231 b | Lysine | 2559 ± 0.00 b | 4493 ± 0.00 a |
| Leucine | 7885 ± 55 a | 7130 ± 144 b | Aspartic acid | 9267 ± 464 a | 8336 ± 373 b |
| Methionine | 777 ± 25 a | 817 ± 87 a | Glutamic acid | 21,108 ± 0.00 b | 21,310 ± 0.00 a |
| Phenylalanine | 6554 ± 0.00 a | 5883 ± 0.00 b | Polar Uncharged | ||
| Tyrosine | 4437 ± 0.00 a | 3670 ± 0.00 b | Serine | 2638 ± 16 a | 2424 ± 57 b |
| Tryptophan | 1838 ± 0.00 a | 1732 ± 0.00 b | Threonine | 1426 ± 39 a | 1229 ± 99 b |
| Alanine | 3325 ± 0.00 a | 3051 ± 0.00 b | Others | ||
| Special Cases | Hydroxylysine | <20 | <20 | ||
| Proline | 4594 ± 151 a | 4119 ± 168 b | Hydroxyproline | 69 ± 3 b | 80 ± 6 a |
| Glycine | 3893 ± 0.00 b | 4072 ± 0.00 a | Essential Amino Acid | 30,290 | 29,466 |
| Cystine | 846 ± 0.00 b | 3467 ± 0.00 a | Total Amino Acid | 82,223 | 81,636 |
| Recipes | Weight Loss (%) | Specific Volume (cm3/g) | Crumb Firmness (N) | Relative Elasticity (%) |
|---|---|---|---|---|
| Control | 12.56 ± 0.12 b | 1.98 ± 0.01 d | 40.16 ± 1.82 e | 52.88 ± 0.33 e |
| Egg albumen | 12.72 ± 0.23 b | 2.07 ± 0.02 c | 65.58 ± 0.73 a | 63.56 ± 1.37 a |
| Protein concentrate | 12.57 ± 0.26 b | 2.06 ± 0.02 c | 50.41 ± 0.95 b | 54.66 ± 1.22 d |
| ADH5% | 13.41 ± 0.24 a | 2.14 ± 0.05 b,c | 47.46 ± 1.03 c | 56.18 ± 0.20 c |
| ADH10% | 13.59 ± 0.29 a | 2.15 ± 0.04 b | 48.35 ± 1.28 b,c | 58.69 ± 0.54 b |
| FDH5% | 13.34 ± 0.18 a | 2.22 ± 0.02 a | 43.65 ± 1.38 d | 57.98 ± 0.66 b |
| FDH10% | 13.37 ± 0.29 a | 2.12 ± 0.06 b,c | 45.71 ± 2.84 c,d | 59.11 ± 0.98 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phongthai, S.; Singsaeng, N.; Nhoo-ied, R.; Suwannatrai, T.; Schönlechner, R.; Unban, K.; Klunklin, W.; Laokuldilok, T.; Phimolsiripol, Y.; Rawdkuen, S. Properties of Peanut (KAC431) Protein Hydrolysates and Their Impact on the Quality of Gluten-Free Rice Bread. Foods 2020, 9, 942. https://doi.org/10.3390/foods9070942
Phongthai S, Singsaeng N, Nhoo-ied R, Suwannatrai T, Schönlechner R, Unban K, Klunklin W, Laokuldilok T, Phimolsiripol Y, Rawdkuen S. Properties of Peanut (KAC431) Protein Hydrolysates and Their Impact on the Quality of Gluten-Free Rice Bread. Foods. 2020; 9(7):942. https://doi.org/10.3390/foods9070942
Chicago/Turabian StylePhongthai, Suphat, Nuttapon Singsaeng, Rossarin Nhoo-ied, Thipubol Suwannatrai, Regine Schönlechner, Kridsada Unban, Warinporn Klunklin, Thunnop Laokuldilok, Yuthana Phimolsiripol, and Saroat Rawdkuen. 2020. "Properties of Peanut (KAC431) Protein Hydrolysates and Their Impact on the Quality of Gluten-Free Rice Bread" Foods 9, no. 7: 942. https://doi.org/10.3390/foods9070942
APA StylePhongthai, S., Singsaeng, N., Nhoo-ied, R., Suwannatrai, T., Schönlechner, R., Unban, K., Klunklin, W., Laokuldilok, T., Phimolsiripol, Y., & Rawdkuen, S. (2020). Properties of Peanut (KAC431) Protein Hydrolysates and Their Impact on the Quality of Gluten-Free Rice Bread. Foods, 9(7), 942. https://doi.org/10.3390/foods9070942