Phytopharmacological Strategies in the Management of Type 2 Diabetes Mellitus





Abstract
1. Introduction
2. Conservative Treatment
Lifestyle Interventions: Diet and Physical Activity
3. Novel Therapeutic Targets for Naturally-Occurring Compounds
3.1. Adiponectin
3.2. Lipase Inhibition
3.2.1. Pharmacological Approach
3.2.2. Naturally-Occurring Lipase Inhibitors
Plants
Polyphenolic Compounds—Fruits, Vegetables and Plants
Polyphenolic Compounds—Seaweed
Alginates from Algae
Metabolic Products from Microorganisms
3.3. PTP1B Inhibition
3.4. DPP4 Inhibition
3.4.1. Incretins and DPP4
3.4.2. Pharmacological Approach
3.4.3. Naturally-Occurring DPP4 Inhibitors
Peptides
Polyphenolic Compounds: Fruits, Vegetables, and Plants
Polyphenolic Compounds: Seaweed
3.5. Alpha-Amylase and/or Alpha-Glucosidase Inhibition
3.5.1. Pharmacological Approach
3.5.2. Naturally-Occurring Alpha-Amylase and/or Alpha-Glucosidase Inhibitors
Peptides
Polyphenolic Compounds: Fruits, Vegetables, and Plants
Seaweed

{kind=link}
{kind=link}
| Compound | Source | Method | Results | Reference |
|---|---|---|---|---|
| Diphlorethohydroxycarmalol (DPHC) | Ishige okamure | In vitro | IC50 0.53 mM | [93] |
| - | Cladophora rupestris | Methanol (72%), ethanol (65%) and ethyl acetate (70%) extracts | [94] | |
| - | Palmaria sp. | Complete inhibition at 2–5 μg/mL, IC50 ≈ 0.1 μg/mL | [95] | |
| Alaria sp. | ||||
| Ascophyllum sp | ||||
| - | Aqueous extract of Ulva reticulata | 89.1 ± 0.96% inhibition after 8 h of extraction | [96] | |
| - | Aqueous extract of Gracilaria edulis | 87.86 ± 3.4% inhibition after 8 h of extraction | ||
| - | Grape seed extract | IC50 8.7 ± 0.8 μg/mL | [79] | |
| - | Green tea extract | IC50 34.9 ± 0.9 μg/mL | ||
| - | White tea extract | IC50 378 ± 134 μg/mL | ||
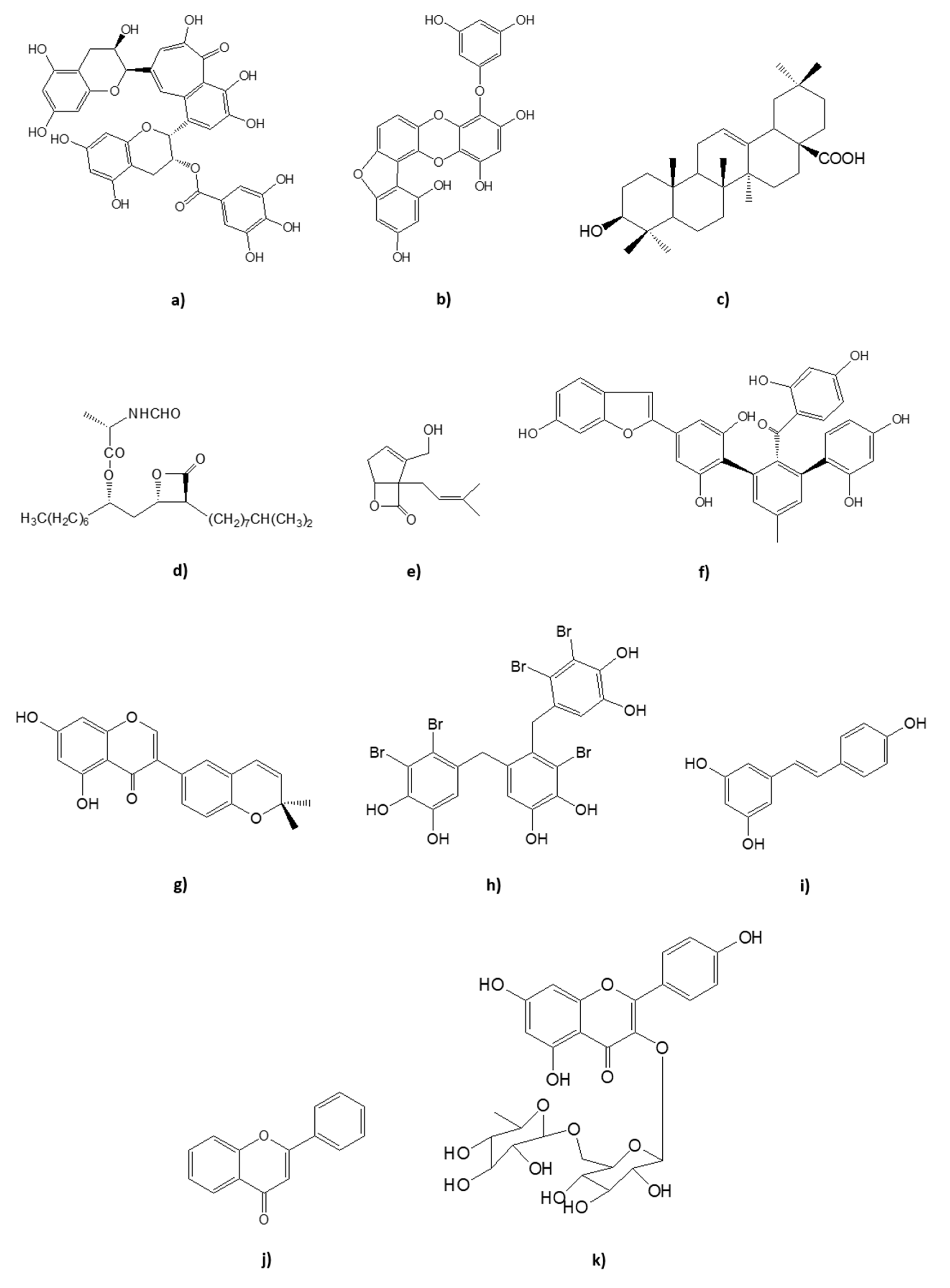
| Kaempferol diglycoside (Figure 1k) | Camelia sinensis | IC50 0.09 ± 0.02 μM | [80] | |
| - | Aqueous extracts of Morinda lucida | IC50 2.30 mg/mL | [81] | |
| - | Ethanolic extract from Senna surattensi | IC50 123.95 μg/mL | [82] | |
| - | Acetone extracts from Picralima nitida | IC50 6.50 mg/mL | [81] | |
| Hydroxytyrosol derivate from the phenolic compound oleuropein | Olea europea | IC50 150 μM at 600 μM | [84] | |
| - | Ethanol-free extract of Abutilon indicum | IC50 191.64 mcg/mL | [85] | |
| - | Aqueous methanol extracts of Ononis angustissima | IC50 2.01 mg/mL | [86] | |
| - | Distilled water extracts of Ononis angustissima | IC50 2.52 mg/mL | ||
| - | Root ethanolic extract of Cissus cornifolia | IC50 22.75 ± 1.23 μg/mL | [87] | |
| - | Aqueous root extract of Cissus cornifolia | IC50 33.70 ± 3.75 μg/mL | ||
| - | Ethanolic extracts (70%) of Hibiscus sabdariffa | IC50 41.77 μg/mL | [88] | |
| - | Aqueous ethanolic extract (70% v/v) of Capsicum annuum | 58% at 5 mg/mL | [89] | |
| - | Purple flesh potatoes extract | IC50 25 μg/mL | [90] | |
| Cys-Ser-Ser-Val | Protein hydrolysate from Andrias davidianus | IC50 13.76 × 103 μg/mL | [78] | |
| Tyr-Ser-Phe-Arg | Protein hydrolysate from Andrias davidianus | IC50 10.82 × 103 μg/mL | ||
| Ser-Ala-Ala-Pro | Protein hydrolysate from Andrias davidianus | IC50 4.46 × 103 μg/mL | ||
| Pro-Gly-Gly-Pro | Protein hydrolysate from Andrias davidianus | IC50 4.23 × 103 μg/mL | ||
| Leu-Gly-Gly-Gly-Asn | Protein hydrolysate from Andrias davidianus | IC50 2.86 × 103 μg/mL | ||
| - | Hexane extracts of leaf essential oil of Juniperus phonicea | IC50 30.15 μg/mL | [25] | |
| - | Methanol extract of Hibiscus sabdariffa | IC50 29.3 ± 0.5 μg/mL | [34] | |
| - | Aqueous extract of Tamarindus indica | IC50 139.4 ± 9 μg/mL | ||
| - | Aqueous extract of Aframomum danielli, Hypodapnis zenkeri, Echinops giganteus, Aframomum citratum, Xylopia aethiopica | >75% inhibition | [35] | |
| Polyphenols | Polyphenol rich extracts of Phaseolus vulgaris | IC50 values ranged from 69 ± 1.9 to 126 ± 3.2 μg/mL and from 107.01 ± 4.5 to 184.20 ± 5.7 μg/mL before and after cooking | [38] | |
| (S)-malic acid | Ethanol and methanol extracts from Flacourtia inermis | IC50 from 1021 to 1949 ppm | [40] | |
| Syringic acid, o-coumaric acid and quercetin | Aqueous extract of Psidium guajava | before the exposure to gastric fluid 14410.60 ± 38 inhibited enzyme unit in μmol·min−1·g−1 | [43] | |
| - | Argyranthemum pinnatifidum (leaves) | IC50 1.55 ± 0.04 mg/mL | [44] | |
| - | Argyranthemum pinnatifidum (flowers) | IC50 2.18 ± 0.05 mg/mL | ||
| - | Artemisia argentea (leaves) | IC50 1.81 ± 0.03 mg/mL | ||
| - | Artemisia argentea (flowers) | IC50 2.48 ± 0.04 mg/mL | ||
| - | Helichrysum devium (leaves) | IC50 1.85 ± 0.06 mg/mL | ||
| - | Helichrysum devium | IC50 2.39 ± 0.03 mg/mL, | ||
| - | Helichrysum melaleucum (leaves) | IC50 1.71 ± 0.02 mg/mL | ||
| - | Helichrysum melaleucum (flowers) | IC50 2.15 ± 0.05 mg/mL | ||
| - | Ethyl acetate extracts pits from Tunisian date palm variety Kentichi | IC50 25.4 ± 0.6 μg/mL | [45] | |
| - | Methanol extracts pits from Tunisian date palm variety Kentichi | IC50 0.072 ± 0.003 μg/mL | ||
| - | Ethanol extract of fresh Euchema denticulatum | 88% inhibition | [50] | |
| Saccharina japonica fermented by Monascus purpureus | IC50 0.98 ± 0.10 mg/mL | [53] |
| Compound | Source | Method | Results | Reference |
|---|---|---|---|---|
| 2,4,6-tribromophenol | Purified from Grateloupia elliptica | In vitro | IC50 60.3 μM | [92] |
| 2,4-dibromophenol | Purified from Grateloupia elliptica | IC50 110.4 μM | ||
| diphlorethohydroxycarmalol (DPHC) | Ishige okamure | IC50 0.16 mM | [83] | |
| - | Cladophora rupestris | Methanol (67%) and ethyl acetate (61%) extracts | [94] | |
| - | Ascophyllum sp. | IC50 ≈ 19 μg/mL | [95] | |
| - | Aqueous extract of Ulva reticulata | 76.02 ± 0.83% inhibition after 8 h of extraction | [96] | |
| - | Aqueous extract of Gracilaria edulis | 79.55 ± 3.08% inhibition after 8 h of extraction | ||
| - | Grape seed extract | IC50 1.2 ± 0.2 μg/mL | [79] | |
| - | Green tea extract | IC50 0.5 ± 0.1 μg/mL | ||
| - | White tea extract | IC50 2.5 ± 0.4 μg/mL | ||
| Kaempferol monoglycoside | Camelia sinensis | IC50 40.02 ± 4.61 μM | [80] | |
| - | Aqueous extracts of Morinda lucida | IC50 2.00 mg/mL | [81] | |
| - | Acetone extracts from Picralima nitida | IC50 3.00 μg/mL | ||
| - | Ethanol-free extract of Abutilon indicum | IC50 = 207.13 mcg/mL | [85] | |
| - | Aqueous methanol extracts of Ononis angustissima | IC50 0.94 mg/mL | [86] | |
| - | N-butanol extracts of Ononis angustissima | IC50 0.99 mg/mL | ||
| - | Root ethanolic extract of Cissus cornifolia | IC50 2.81 ± 0.97 μg/mL | [87] | |
| - | Aqueous root extract of Cissus cornifolia | IC50 37.48 ± 2.35 μg/mL | ||
| - | Ethanolic extracts (70%) of Hibiscus sabdariffa | IC50 18.09 μg/mL | [88] | |
| - | Aqueous ethanolic extract (70% v/v) of Capsicum annuum | 66% at 5 mg/mL | [89] | |
| - | Purple flesh potatoes extract | IC50 42 μg/mL | [90] | |
| Cys-Ser-Ser-Val | Protein hydrolysate from Andrias davidianus | IC50 206 μg/mL | [78] | |
| Tyr-Ser-Phe-Arg | IC50 162 μg/mL | |||
| Ser-Ala-Ala-Pro | IC50 66.90 μg/mL | |||
| Pro-Gly-Gly-Pro | IC50 63.50 μg/mL | |||
| Leu-Gly-Gly-Gly-Asn | IC50 42.93 μg/mL | |||
| 5-methoxy-7-hydroxy-9,10-dihydro-1,4-phenanthrenequinone | Methanol extract of Dendobium formosum | IC50 126.88 ± 0.66μM | [24] | |
| Polyphenols | Polyphenol rich extracts of Phaseolus vulgaris | IC50 values ranged from 39.3 ± 4.4 to 74.13 ± 6.9 μg/mL and from 51 ± 7.7 to 122.1 ± 5.2 μg/mL before and after cooking | [38] | |
| (S)-malic acid | Ethanol and methanol extracts from Flacourtia inermis | IC50 from 549 to 710 ppm | [40] | |
| Catechin | Aqueous extract of Psidium guajava | Before gastric fluid exposure 28.82 ± 0.02 inhibited enzyme unit in μmol·min−1·g−1 and after exposure 2.59 ± 0.06 inhibited enzyme unit in μmol·min−1·g−1 | [43] | |
| - | Argyranthemum pinnatifidum (leaves) | IC50 0.57 ± 0.03 mg/mL | [44] | |
| - | Argyranthemum pinnatifidum (flowers) | IC50 0.81 ± 0.02 mg/mL | ||
| - | Saccharina japonica fermented by Monascus purpureus | Maltose IC50 0.02 ± 0.07 mg/mL and sucrose IC50 0.08 ± 0.13 mg/mL | [53] | |
| Polyphenols | Winemaking generates by-products | From 75.6 ± 2.5% to 93.7 ± 0.5%, samples treated with pronase and from 84.5 ± 0.5% to 96.5 ± 2.9% viscozyme | [48] |
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sharifuddin, Y.; Chin, Y.X.; Lim, P.E.; Phang, S.M. Potencial bioactive compounds from seaweed for diabetes management. Mar. Drugs 2015, 13, 5447–5491. [Google Scholar] [CrossRef] [PubMed]
- Gago, A.S. Compostos Bioativos de Microalgas com Interesse no Tratamento da Diabetes. Master’s Thesis, Faculdade de Ciências e Tecnologia, Algarve, Portugal, 2016. [Google Scholar]
- Motshakeri, M.; Ebrahimi, M.; Goh, Y.M.; Othman, H.H.; Hair-Bejo, M.; Mohamed, S. Effects of brown seaweed (Sargassum polycystum) extracts on kidney, liver, and pancreas of type 2 diabetic rat model. Evid. Based Complment. Altern. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Hsu, M.J.; Agoramoorthy, G. Understanding the molecular dynamics of type-2 diabetes drug target DPP-4 and its interaction with sitagliptin and inhibitor diprotin-A. Cell Biochem. Biophys. 2014, 70, 907–922. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Khunti, K.; Davies, M.J. Type 2 diabetes. Lancet 2017, 389, 2239–2251. [Google Scholar] [CrossRef]
- Lee, S.H.; Jeon, Y.J. Anti-diabetic effects of brown algae derived phlorotannins marine polyphenols through diverse mechanisms. Fitoterapia 2013, 86, 129–136. [Google Scholar] [CrossRef]
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2010, 33, S62–S69. [Google Scholar] [CrossRef]
- Hussain, A.; Claussen, B.; Ramachandran, A.; Williams, R. Prevention of type 2 diabetes: A review. Diabetes Res. Clin. Pract. 2007, 76, 317–326. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, H.C.; Vitek, L.; Nam, C.M. Algae Consumption and Risk of Type 2 Diabetes: Korean National Health and Nutrition Examination Survey in 2005. J. Nutr. Sci. Vitaminol. 2010, 56, 13–18. [Google Scholar] [CrossRef]
- Fowler, M.J. Microvascular e macrovascular complications of diabetes. Clin. Diabetes 2008, 26, 77–82. [Google Scholar] [CrossRef]
- Duarte, R. Inibidores da DPP-4 (gliptinas)-10 anos depois (2007–2017). RPD 2017, 12, 62–67. [Google Scholar]
- Liu, D.; Gao, H.; Tang, W.; Nie, S. Plant non-starch polysaccharides that inhibit key enzymes linked to type 2 diabetes mellitus. Ann. N. Y. Acad. Sci. 2017, 1401, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.L.; Kitajima, C.; Ito, H.; Miyazaki, T.; Baba, M.; Okuyama, T.; Okada, Y. Antidiabetic effect of polyphenols from brown alga Ecklonia kurome in genetically diabetic KK-AY mice. Pharm. Biol. 2012, 50, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, B.T.; Jarvis, J.; Khunti, K.; Davies, M.J. Recent advances in the management of type 2 diabetes mellitus: A review. Postgrad. Med. J. 2008, 84, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.A.; Kushi, L.H.; Jacobs, D.; Slavin, J.; Sellers, T.A.; Folsom, A.R. Carbohydrates, dietary fiber, and incident type 2 diabetes in wolder woman. Am. J. Clin. Nutr. 2000, 71, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Forouhi, N.G.; Misra, A.; Mohan, V.; Taylor, R.; Yancy, W. Dietary and nutritional approaches for prevention and management of type 2 diabetes. BMJ 2018, 361, 1–9. [Google Scholar] [CrossRef]
- Caprita, R.; Caprita, A.; Julean, C. Biochemical aspects of non-starch polysaccharides. Sci. Pap. Anim. Sci. Biotechnol. 2010, 43, 368–375. [Google Scholar]
- Colberg, S.R.; Sigal, R.J.; Fernhall, B.; Regensteiner, J.G.; Blissmer, B.J.; Rubin, R.R.; Chasan-Taber, L.; Albright, A.L.; Braun, B. Exercise and type 2 diabetes. Diabetes Care 2010, 33, 147–167. [Google Scholar] [CrossRef]
- Achari, A.E.; Jain, S.K. Adiponectin, a therapeutic target for obesity, diabetes, and endothelial dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef]
- Weyer, C.; Funahashi, T.; Tanaka, S.; Hotta, K.; Matsuzawa, Y.; Pratley, R.E.; Tataranni, P.A. Hypoadiponectinemia in obesity and type 2 diabetes: Close association with insulin resistance and hyperinsulinemia. J. Clin. Endocrinol. Metab. 2001, 86, 1930–1935. [Google Scholar] [CrossRef]
- Ríos, J.L.; Francini, F.; Schinella, G. Natural products for the treatment of type 2 diabetes mellitus. Planta Med. 2015, 81, 975–994. [Google Scholar] [CrossRef]
- De la Garza, A.L.; Milagro, F.I.; Boque, N.; Campión, J.; Martínez, J.A. Natural inhibitors of pancreatic lipase as new players in obesity treatment. Planta Med. 2011, 77, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Roh, C.; Jung, U. Screening of Crude Plant Extracts with Anti-Obesity Activity. Int. J. Mol. Sci. 2012, 13, 1710–1719. [Google Scholar] [CrossRef] [PubMed]
- Inthongkaew, P.; Chatsumpun, N.; Supasuteekul, C.; Kitisripanya, T.; Putalun, W.; Likhitwitayawuid, K.; Sritularak, B. α-Glucosidase and pancreatic lipase inhibitory activities and glucose uptake stimulatory effect of phenolic compounds from Dendrobium formosum. Rev. Bras. Farmacogn. 2017, 27, 480–487. [Google Scholar] [CrossRef]
- Keskes, H.; Mnafgui, K.; Hamden, K.; Damak, M.; El-Feki, A.; Allouche, N. In vitro anti-diabetic, anti-obesity and antioxidant proprieties of Juniperus phoenicea L. leaves from Tunisia. Asian Pac. J. Trop. Biomed. 2014, 4, S649–S655. [Google Scholar] [CrossRef]
- Briones, A.T.; Chichioco-Hernandez, C.L. Lipase inhibitory activity of Carica papaya, Chrysophyllum cainito, Corcorus olitorius, Cympogon citrates and Syzygium cumini extracts. Food Res. 2017, 2, 51–55. [Google Scholar] [CrossRef]
- Moon, S.H.; Kim, M.Y. Phytochemical profile, antioxidant, antimicrobial and antipancreatic lipase activities of fermented Camellia japonica L leaf extracts. Trop. J. Pharm. Res. 2018, 17, 905–912. [Google Scholar] [CrossRef]
- Jamous, R.M.; Abu-Zaitoun, S.Y.; Akkawi, R.J.; Ali-Shtayeh, M.S. Antiobesity and antioxidant potentials of selected Palestinian medicinal plants. Evid. Based Complement. Altern. Med. 2018, 2018, 1–21. [Google Scholar] [CrossRef]
- Glisan, S.L.; Grove, K.A.; Yennawar, N.H.; Lambert, J.D. Inhibition of pancreatic lipase by black tea theaflavins: Comparative enzymology and In silico modeling studies. Food Chem. 2017, 216, 296–300. [Google Scholar] [CrossRef]
- Zhu, Y.T.; Ren, X.Y.; Yuan, L.; Liu, Y.M.; Liang, J.; Liao, X. Fast identification of lipase inhibitors in oolong tea by using lipase functionalised Fe3O4 magnetic nanoparticles coupled with UPLC–MS/MS. Food Chem. 2015, 173, 521–526. [Google Scholar] [CrossRef]
- Yu, H.; Dong, S.; Wang, L.; Yun, L. The effect of triterpenoid saponins on pancreatic lipase In vitro: Activity, conformation, kinetics, thermodynamics and morphology. Biochem. Eng. J. 2017, 125, 1–9. [Google Scholar] [CrossRef]
- Oliveira, V.B.; Araújo, R.L.B.; Eidenberger, T.; Brandão, M.G.L. Chemical composition and inhibitory activities on dipeptidyl peptidase IV and pancreatic lipase of two underutilized species from the Brazilian Savannah: Oxalis cordata A.St.-Hil. and Xylopia aromatica (Lam.) Mart. Food Res. Int. 2018, 105, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Belfeki, H.; Mnasser, H.; Mondher, M. Antioxidant and anti-lipases activities In vitro of Mentha viridis and Eucalyptus globulus extracts. Ind. Crop. Prod. 2016, 89, 514–521. [Google Scholar] [CrossRef]
- Buchhols, T.; Melzig, M.F. Medicinal plants traditionally used for treatment of obesity and diabetes mellitus-screening for pancreatic lipase and α-amylase inhibition. Phytother. Res. 2016, 30, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Etoundi, C.B.; Kuate, D.; Ngondi, J.L.; Oben, J. Anti-amylase, anti-lipase and antioxidant effects of aqueous extracts of some Cameroonian spices. J. Nat. Prod. 2010, 3, 165–171. [Google Scholar]
- Maqsood, M.; Ahmed, D.; Atique, I.; Malik, W. Lipase inhibitory activity of Lagenaria siceraria fruit as a strategy to treat obesity. Asian Pac. J. Trop. Biomed. 2017, 10, 305–310. [Google Scholar] [CrossRef]
- Liu, T.T.; He, X.R.; Xu, R.X.; Wu, X.B.; Qi, Y.X.; Huang, J.Z.; Chen, Q.H.; Chen, Q.X. Inhibitory mechanism and molecular analysis of furoic acid and oxalic acid on lipase. Int. J. Biol. Macromol. 2018, 120, 1925–1934. [Google Scholar] [CrossRef]
- Ombra, M.N.; d’Acierno, A.; Nazzaro, F.; Spigno, P.; Riccardi, R.; Zaccardelli, M.; Pane, C.; Coppola, R.; Fratianni, F. Alpha-amylase, a-glucosidase and lipase inhibiting activities of polyphenol-rich extracts from six common bean cultivars of Southern Italy, before and after cooking. Int. J. Food Sci. Nutr. 2018, 69, 824–834. [Google Scholar] [CrossRef]
- Austin, C.; Stewart, D.; Allwood, J.W.; Mcdougall, G.J. Extracts from the edible seaweed, Ascophyllum nodosum, inhibit lipase activity In vitro: Contributions of phenolic and polysaccharide components. Food Funct. 2018, 9, 502–510. [Google Scholar] [CrossRef]
- Alakolanga, A.G.; Kumar, N.S.; Jayasinghe, L.; Fujimoto, Y. Antioxidant property and α -glucosidase, α -amylase and lipase inhibiting activities of Flacourtia inermis fruits: Characterization of malic acid as an inhibitor of the enzymes. J. Food Sci. Technol. 2015, 52, 8383–8388. [Google Scholar] [CrossRef]
- Ikarashi, N.; Takeda, R.; Ito, K.; Ochiai, W.; Suigiyama, K. The inhibition of lipase and glucosidase activities by Acacia polyphenol. Evid. Based Complement. Altern. Med. 2011, 2011, 1–8. [Google Scholar] [CrossRef]
- Han, L.; Li, W.; Narimatsu, S.; Liu, L.; Fu, H.; Okuda, H.; Koike, K. Inhibitory effects of compounds isolated from fruit of Juglans mandshurica on pancreatic lipase. J. Nat. Med. 2006, 61, 184–186. [Google Scholar] [CrossRef]
- Simão, A.A.; Marques, T.R.; Marcussi, S.; Corrêa, A.D. Aqueous extract of Psidium guajava leaves: Phenolic compounds and inhibitory potential on digestive enzymes. An. Acad. Bras. Cienc. 2017, 89, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Spínola, V.; Castilho, P.C. Evaluation of Asteraceae herbal extracts in the management of diabetes and obesity. Contribution of caffeoylquinic acids on the inhibition of digestive enzymes activity and formation of advanced glycation end-products (In vitro). Phytochemistry 2017, 143, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Masmoudi-Allouche, F.; Touati, S.; Mnafgui, K.; Gharsallah, N.; El Feki, A.; Allouche, N. Phytochemical profile, antioxidant, antibacterial, antidiabetic and anti-obesity activities of fruits and pits from date palm (Phoenix dactylifera L.) grown in south of Tunisia. J. Pharmacogn. Phytochem. 2016, 5, 15–22. [Google Scholar]
- Jang, D.S.; Lee, G.Y.; Kim, J.; Lee, Y.M.; Kim, J.M.; Kim, Y.S. A New Pancreatic Lipase Inhibitor Isolated from the Roots of Actinidia arguta. Arch. Pharm. Res. 2008, 31, 666–670. [Google Scholar] [CrossRef]
- Wan, L.H.; Jiang, X.L.; Liu, Y.M.; Hu, J.J.; Liang, J.; Liao, X. Screening of lipase inhibitors from Scutellaria baicalensis extract using lipase immobilized on magnetic nanoparticles and study on the inhibitory mechanism. Anal. Bioanal. Chem. 2016, 408, 2275–2283. [Google Scholar] [CrossRef]
- de Camargo, A.C.; Regitano-d’Arce, M.A.B.; Biasoto, A.C.T.; Shahidi, F. Enzyme-assisted extraction of phenolics from winemaking by-products: Antioxidant potential and inhibition of alpha-glucosidase and lipase activities. Food Chem. 2016, 212, 395–402. [Google Scholar] [CrossRef]
- Wong, X.; Madrid, A.M.; Tralma, K.; Castillo, R.; Carrasco-Pozo, C.; Navarrete, P.; Beltrán, C.; Pastene, E.; Gotteland, M. Polyphenol extracts interfere with bacterial lipopolysaccharide In vitro and decrease postprandial endotoxemia in human volunteers. J. Funct. Foods 2016, 26, 406–417. [Google Scholar] [CrossRef]
- Balasubramaniam, V.; Mustar, S.; Khalid, N.M.; Rashed, A. Inhibitory activities of three Malaysian edible seaweeds on lipase and α-amylase. J. Appl. Phycol. 2013, 25, 441–445. [Google Scholar] [CrossRef]
- Bitou, N.; Ninomiya, M.; Tsujita, T.; Okuda, H. Screening of lipase inhibitors from marine algae. Lipids 1999, 34, 441–445. [Google Scholar] [CrossRef]
- Eom, S.H.; Lee, M.S.; Lee, E.W.; Kim, Y.M.; Kim, T.H. Pancreatic lipase inhibitory activity of phlorotannins isolated from Eisenia bicyclis. Phytother. Res. 2012, 27, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Suraiya, S.; Lee, J.M.; Cho, H.J.; Jang, W.J.; Kim, D.G.; Kim, Y.O.; Kong, I.S. Monascus spp. fermented brown seaweeds extracts enhance bio-functional activities. Food Biosci. 2018, 21, 90–99. [Google Scholar] [CrossRef]
- Birari, R.B.; Bhutani, K.K. Pancreatic lipase inhibitors from natural sources: Unexplored potential. Drug Discov. Today 2007, 12, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, T.; Melzig, M.F. Polyphenolic compounds as pancreatic lipase inhibitors. Planta Med. 2015, 81, 771–783. [Google Scholar] [CrossRef]
- Wilcox, M.D.; Brownlee, I.A.; Richardson, J.C.; Dettmar, P.W.; Pearson, J.P. The modulation of pancreatic lipase activity by alginates. Food Chem. 2014, 146, 479–484. [Google Scholar] [CrossRef]
- Bower, A.M.; Real Hernandez, L.M.; Berhow, M.A.; de Mejia, E.G. Bioactive compounds from culinary herbs inhibit a molecular target for type 2 diabetes management, dipeptidyl peptidase IV. J. Agric. Food Chem. 2014, 62, 6147–6158. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, C.S.; Gao, L.X.; Wang, Z.H.; Li, J.Y.; Li, J.; Li, X.W.; Guo, Y.W. Design, synthesis and In vitro activity of phidianidine B derivatives as novel PTP1B inhibitors with specific selectivity. Bioorg. Med. Chem. Lett. 2016, 26, 778–781. [Google Scholar] [CrossRef]
- Huang, Q.H.; Lei, C.; Wang, P.P.; Li, J.Y.; Li, J.; Hou, A.J. Isoprenylated phenolic compounds with PTP1B inhibition from Morus alba. Fitoterapia 2017, 122, 138–143. [Google Scholar] [CrossRef]
- Zhang, J.; Sasaki, T.; Li, W.; Nagata, K.; Higai, K.; Feng, F.; Wang, J.; Cheng, M.; Koike, K. Identification of caffeoylquinic acid derivatives as natural protein tyrosine phosphatase 1B inhibitors from Artemisia princeps. Bioorg. Med. Chem. Lett. 2018, 28, 1194–1197. [Google Scholar] [CrossRef]
- Trinh, B.T.D.; Jäger, A.K.; Staerk, D. High-resolution inhibition profiling combined with HPLC-HRMS-SPE-NMR for identification of PTP1B inhibitors from Vietnamese plants. Molecules 2017, 22, 1228. [Google Scholar] [CrossRef]
- Ezzat, S.M.; Bishbishy, M.H.E.; Habtemariam, S.; Salehi, B.; Sharifi-Rad, M.; Martins, N.; Sharifi-Rad, J. Looking at marine-derived bioactive molecules as upcoming anti-diabetic agents: A special emphasis on PTP1B inhibitors. Molecules 2018, 23, 3334. [Google Scholar] [CrossRef] [PubMed]
- dos Reis, M.S.R. Inibidores da DPP-4: Uma Esperança na Nefropatia Diabética? Master’s Thesis, Faculdade de Medicina da Universidade de Coimbra, Coimbra, Portugal, 2015. [Google Scholar]
- Chin, Y.X.; Lim, P.E.; Maggs, C.A.; Phang, S.M. Anti-diabetic potencial of selected Malaysian seaweeds. J. Appl. Phycol. 2014, 27, 2137–2148. [Google Scholar] [CrossRef]
- Unnikrishnan, P.S.; Suthindhiran, K.; Jayasri, M.A. Antidiabetic potential of marine algae by inhibiting key metabolic enzymes. Front. Life Sci. 2015, 8, 148–159. [Google Scholar] [CrossRef]
- Power, O.; Nongonierma, A.B.; Jakeman, P.; FitzGerald, R.J. Food protein hydrolysates as a source od dipeptidyl peptidase for the management of type 2 diabetes. Proc. Nutr. Soc. 2014, 73, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Tulipano, G.; Sibilia, V.; Caroli, A.M.; Cocchi, D. Whey proteins as source of dipeptidyl dipeptidase IV (dipeptidyl peptidase-4) inhibitors. Peptides 2011, 32, 835–838. [Google Scholar] [CrossRef] [PubMed]
- Suman, R.K.; Mohanty, I.R.; Maheshwari, U.; Borde, M.K.; Deshmukh, Y.A. Natural dipeptidyl peptidase-IV inhibitor mangiferin mitigates diabetes-and metabolic syndrome-Induced changes in experimental rats. Diabetes Metab. Syndr. Obes. 2016, 9, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Evaluation of the potential of dietary proteins as precursors of dipeptidyl peptidase (DPP)-IV inhibitors by an In silico approach. J. Funct. Foods 2012, 4, 403–422. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Dipeptidyl peptidase IV inhibitory and antioxidative properties of milk protein-derived dipeptides and hydrolysates. Peptides 2013, 39, 157–163. [Google Scholar] [CrossRef]
- Fan, J.; Johnson, M.H.; Lila, M.A.; Yousef, G.; de Mejia, E.G. Berry and citrus phenolic compounds inhibit dipeptidyl peptidase IV: Implications in diabetes management. Evid. Based Complement. Altern. Med. 2013, 2013, 1–13. [Google Scholar] [CrossRef]
- Unnikrishnan, P.S.; Suthindhiran, K.; Jayasri, M.A. Inhibitory potencial of Turbinaria ornata against key metabolic enzymes linked to diabetes. Biomed. Res. Int. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Pascual, I.; Lopéz, A.; Gómez, H.; Chappe, M. Screening of inhibitors of porcine dipeptidyl peptidase IV activity in aqueous extracts from marine organisms. Enzyme. Microb. Technol. 2007, 40, 414–419. [Google Scholar] [CrossRef]
- Jao, C.L.; Hung, C.C.; Tung, Y.S.; Lin, P.Y.; Chen, M.C.; Hsu, K.C. The development of bioactive peptides from dietary proteins as a dipeptidyl peptidase IV inhibitor for the management of type 2 diabetes. Biomedicine 2015, 5, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Food-derived dipeptidyl-peptidase IV inhibitors as a potential approach for glycemic regulation-current knowledge and future research considerations. Trends Food Sci. Technol. 2016, 54, 1–16. [Google Scholar] [CrossRef]
- Pereira, L.; Neto, J.M. Marine Algae-Biodiversity, Taxonomy, Environmental Assessment, and Biotechnology, 1st ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: Boca Raton, FL, USA, 2015; pp. 1–2. [Google Scholar]
- Sun, Z.; Chen, F. Evaluation of green alga Chlorella pyrenoidosa for management of diabetes. J. Food Drug Anal. 2012, 20, 246–249. [Google Scholar]
- Ramadhan, A.H.; Nawas, T.; Zhang, X. Purification and identification of a novel antidiabetic peptide from Chinese giant salamander (Andrias davidianus) protein hydrolysate against α-amylase and α-glucosidase. Int. J. Food Prop. 2018, 20, S3360–S3372. [Google Scholar] [CrossRef]
- Yilmazer-Musa, M.; Griffith, A.M.; Michels, A.J.; Schneider, E.; Frei, B. Inhibition of α-amylase and α-glucosidase activity by tea and grape seed extracts and their constituent catechins. J. Agric. Food Chem. 2012, 60, 8924–8929. [Google Scholar] [CrossRef]
- Hua, F.; Zhou, P.; Wu, H.Y.; Chu, G.X.; Xie, Z.W.; Bao, G.H. Inhibition of flavonoid glycosides from Lu’an GuaPian tea on α-glucosidase and α-amylase: Molecular docking and interaction mechanism. Food Funct. 2018, 9, 4173–4183. [Google Scholar] [CrossRef]
- Kazeem, M.I.; Adamson, J.O.; Ogunwande, I.A. Modes of inhibition of α-amylase and α-glucosidase by aqueous extract of Morinda lucida benth leaf. Biomed. Res. Int. 2013, 2013, 1–6. [Google Scholar] [CrossRef]
- Thilagam, E.; Parimaladevi, B.; Kumarappan, C.; Mandal, S.C. α-Glucosidase and α-amylase inhibitory activity of Senna surattensis. J. Acupunct. Meridian Stud. 2013, 6, 24–30. [Google Scholar] [CrossRef]
- Kazeem, M.I.; Ogunbiyi, J.V.; Ashafa, A.O.T. In vitro studies on the inhibition of α-amylase and α-glucosidase by leaf extracts of Picralima nitida (staft). TJPR 2013, 12, 719–725. [Google Scholar] [CrossRef]
- Hadrich, F.; Bouallagui, Z.; Junkyu, H.; Isoda, H.; Sayadi, S. The α-glucosidase and α-amylase enzyme inhibitory of hidroxytyrosol and oleuropein. J. Oleo Sci. 2015, 64, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Arce, F.J.V.; Concepcion, J.E.D.; Mayol, K.M.C.; See, G.L.L. In vitro α-amylase and α-glucosidase inhibition activity of tabing Abutilon indicum (Linn 1836) root extracts. IJTPR 2016, 8, 391–396. [Google Scholar]
- Laoufi, H.; Nabila, B.; Adjdir, S.; Djaziri, R. In vitro α-amylase and α-glucosidase inhibitory activity of Ononis angustissima extracts. J. Appl. Pharm. Sci. 2017, 7, 191–198. [Google Scholar] [CrossRef]
- Chipiti, T.; Ibrahim, M.A.; Singh, M.; Islam, M.S. In vitro α-amylase and α-glucosidase inhibitory and cytotoxic activities of extracts from Cissus cornifolia planch parts. Pharmacogn. Mag. 2017, 13, S329–S333. [Google Scholar] [CrossRef]
- Gondokesumo, M.E.; Kusuma, H.S.W.; Widowati, W. α-/β-glucosidase and α-amylase inhibitory activities of roselle (Hibiscus sabdariffa L.) ethanol extract. MCBS 2017, 1, 34–40. [Google Scholar] [CrossRef]
- Watcharachaisoponsiri, T.; Sornchan, P.; Charoenkiatkul, S.; Suttisansanee, U. The α-glucosidase and α-amylase inhibitory activity from different chili pepper extracts. Int. Food Res. J. 2016, 23, 1439–1445. [Google Scholar]
- Kalita, D.; Holm, D.G.; LaBarbera, D.V.; Petrash, J.M.; Jayanty, S.S. Inhibition of α-glucosidase, α-amylase, and aldose reductase by potato polyphenolic compounds. PLoS ONE 2018, 13, e0191025. [Google Scholar] [CrossRef]
- Okada, Y.; Ishimaru, A.; Suzuki, R.; Okuyama, T. A new phloroglucinol derivative from the brown alga Eisenia bibyclis: Potencial for the effective treatment of diabetic complications. J. Nat. Prod. 2004, 67, 103–105. [Google Scholar] [CrossRef]
- Kim, K.Y.; Nam, K.A.; Kurihara, H.; Kim, S.M. Potent α-glucosidase inhibitors purifief from the red alga Grateloupia elliptica. Phytochemistry 2008, 69, 2820–2825. [Google Scholar] [CrossRef]
- Heo, S.J.; Hwang, J.Y.; Choi, J.I.; Han, J.S.; Kim, H.J.; Jeon, Y.J. Diphlorethohydroxycarmalol isolated from Ishige okamure, a brown algae, a potent α-glucosidase and α-amylase inhibitor, alleviates postprandial hyperglycemia in diabetic mice. Eur. J. Pharmacol. 2009, 615, 252–256. [Google Scholar] [CrossRef]
- Krish, S.; Das, A. In-vitro bioactivity of marine seaweed, Cladophora rupestris. Int. J. Pharma Bio Sci. 2014, 5, B898–B908. [Google Scholar] [CrossRef]
- Nwosu, F.; Morris, J.; Lund, V.A.; Stewart, D.; Ross, H.A.; McDougall, G.J. Anti-proliferative and potential anti-diabetic effects of phenolic-rich extracts from edible marine algae. Food Chem. 2011, 126, 1006–1012. [Google Scholar] [CrossRef]
- Reka, P.; Banu, T.; Seethalakshmi, M. Alpha amylase and alpha glucosidase inhibition activity of selected edible seaweeds from south coast area of India. Int. J. Pharm. Pharm. Sci. 2017, 9, 64–68. [Google Scholar] [CrossRef]

| Compound | Source | Method | Results | Reference |
|---|---|---|---|---|
| - | Crude extracts of Rubi Fructus | In vitro | 32.5% inhibition at 100 μg/mL | [23] |
| - | Crude extracts of Corni Fructus | 34.8% inhibition at 100 μg/mL | ||
| - | Crude extracts of Salicis Radicis et Cortex | 38% inhibition at 100 μg/mL | ||
| - | Crude extracts of Geranium nepalense | 31.4% inhibition at 100 μg/mL | ||
| 5-methoxy-7-hydroxy-9,10-dihydro-1,4-phenanthrenequinone | Methanol extract of Dendobium formosum | IC50 69.45 ± 10.14 μM | [24] | |
| - | Methanol extracts of leaf essential oil of Juniperus phonicea | IC50 60.22 μg/mL | [25] | |
| - | Ethanolic extracts of Chrysophyllum cainito | 74.91% inhibition and 92.11% for bioactive compounds concentrated in hexane | [26] | |
| - | Methanol extracts of fermented Camellia japonica | IC50 0.308 mg/mL | [27] | |
| - | Ethanol extracts of Camelia sinensis | IC50 0.5 mg/mL | [28] | |
| (−)-Epigallocatechin-3-gallate | Green Tea Camelia sinensis | IC50 7.5 μmol/L | [29] | |
| Theaflavin-3,3′-digallate | Black Tea Camelia sinensis | IC50 1.9 μmol/L | ||
| theaflavin-3′-gallate | Black Tea Camelia sinensis | IC50 4.2 μmol/L | ||
| theaflavin-3-gallate (Figure 1a) | Black Tea Camelia sinensis | IC50 3 μmol/L | ||
| theaflavin | Black Tea Camelia sinensis | IC50 >10 μmol/L | ||
| (−)-epigallocatechin-3-O-gallate | Oolong tea Camelia sinensis | IC50 54.97 ± 1.32 μM | [30] | |
| (−)-gallocatechin-3-O-gallate | Oolong tea Camelia sinensis | IC50 54.68 ± 2.23 μM | ||
| (−)-epicatechin-3-O-gallate | Oolong tea Camelia sinensis | IC50 38.22 ± 0.89 μM | ||
| Tea saponin | - | EC50 242 mM | [31] | |
| Oleanolic acid | - | EC50 0.895 mM | ||
| Betulin | - | EC50 0.958 mM | ||
| Ginsenoside Ro | - | EC50 0.885 mM | ||
| Ginsenoside Rd | - | EC50 388 mM | ||
| - | Ethanol extracts of Ceratonia siliqua | IC50 0.8 mg/mL | [28] | |
| - | Ethanol extracts of Curcuma longa | IC50 0.8 mg/mL | ||
| - | Ethanol extracts of Sarcopoterium spinosum | IC50 1.2 mg/mL | ||
| - | Ethanol extracts of Mentha spicata | IC50 1.2 mg/mL | ||
| - | Crude extract of Oxalis cordata | IC50 0.84 mg/mL | [32] | |
| - | Ethyl acetate extract of Oxalis cordata | IC50 0.88 mg/mL | ||
| - | Aqueous fraction of Oxalis cordata | IC50 0.63 mg/mL | ||
| Phenolic compounds | Methanol extracts of Eucalyptus globulus | 78.92% inhibition against lipase from Aspergillus niger and 82.65% against lipase from olive | [33] | |
| Phenolic compounds | Methanol extracts of Mentha viridis | 72.75% inhibition against lipase from Aspergillus niger and 75.75% against lipase from olive | ||
| - | Methanol extract of Hibiscus sabdariffa L. | IC50 35.8 ± 0.8 μg/mL | [34] | |
| - | Methanol extract of Tamarindus indica L. | IC50 152 ± 7 μg/mL | ||
| - | Aqueous extracts of Xylopia aethiopica | 92.25% inhibition | [35] | |
| - | Aqueous extracts of Scorodophloeus zenkeri | 56.39% inhibition | ||
| Hexadecanoic acid, methyl hexadecanoate, isopropryl palmitate, methyl 9,12-octadecadienate and methyl 9,12,15-octadecatrienoate | Chloroform fractions of Lagenaria siceraria | IC50 157.59 μg/mL | [36] | |
| Furoic acid | Compounds derived from the oxidation of vitamin C | IC50 2.12 ± 0.04 mM | [37] | |
| Oxalic acid | - | IC50 15.05 ± 0.78 mM | ||
| Polyphenols | Polyphenol rich extracts of Phaseolus vulgaris | IC50 values ranged from 63.11 ± 7.5 to 103.2 ± 5.9 μg/mL and from 92 ± 6.3 to 128.5 ± 7.4 μg/mL before and after cooking | [38] | |
| - | Polyphenol rich extracts of Ascophyllum nodosum | oil-based turbidimetric assay | IC50 200 μg gallic acid equivalents per assay | [39] |
| Purified phlorotannin-enriched fraction | Polyphenol rich extracts of Ascophyllum nodosum | IC50 60 μg gallic acid equivalents per assay | ||
| (S)-malic acid | Ethanol and methanol extracts of Lovi from Flacourtia inermis | In vitro | IC50 from 1290 to 2096 ppm | [40] |
| Catechin-like flavan-3-ols | Acacia mearnsii | IC50 0.95 mg/mL | [41] | |
| - | Water extract of Juglans mandshurica | IC50 2.3 mg/mL | [42] | |
| 1,4,8-trihydroxynaphthalene-1-O-β-D [6′-O-(3′’,4′’,5′’-trihydroxybenzoyl)] glucopyranoside | Water extract of Juglans mandshurica | 88% inhibition at 1mM | ||
| Epigallocatechin gallate and revesterol | Aqueous extract of Psidium guajava (Pedro Sato cultivar) | Before exposure to gastric fluid 36.45 ± 0.68 inhibited enzyme unit in μmol·min−1·g−1 and after exposure 43.33 ± 1.80 inhibited enzyme unit in μmol·min−1·g−1 | [43] | |
| - | Phagnalon lowei | IC50 3.05 ± 0.07 mg/mL | [44] | |
| - | Ethyl acetate extracts pits from Tunisian date palm variety Kentichi | IC50 5.4 ± 0.6 μg/mL | [45] | |
| - | Methanol extracts pits from Tunisian date palm variety Kentichi | IC50 1.21 ± 0.23 μg/mL | ||
| 3-O-trans-p-coumaroyl actinidic acid | Roots of Actinidia arguta | IC50 14.95 μM | [46] | |
| Baicalin | Scutellaria baicalensis | IC50 229.22 ± 12.67 μM | [47] | |
| Wogonin | Scutellaria baicalensis | IC50 153.71 ± 9.21 μM | ||
| Oroxylin A | Scutellaria baicalensis | IC50 56.07 ± 4.90 μM | ||
| Polyphenols | Winemaking generates by-products | From 35.2 ± 0.2% to 45.5 ± 1.2%, samples treated with pronase and from 86.2 ± 0.3% to 94.3 ± 1.5% viscozyme | [48] | |
| - | Grape extract | IC50 8.6 ± 1.1mg/mL | [49] | |
| - | Ethanol extract of dried Kappaphycus striatus | 92% inhibition | [50] | |
| Caulerpenyne | Ethyl acetate extract of Caulerpa taxifolia | 50% inhibition | [51] | |
| Fucofuroeckol A (Figure 1b) | Methanol extract of Eisenia bicyclis | IC50 37.2 ± 2.3 μM | [52] | |
| 7-phloroeckol | Methanol extract of Eisenia bicyclis | IC50 12.7 ± 1 μM | ||
| - | Saccharina japonica fermented by Monascus purpureus | IC50 4.98 ± 0.85 μg/mL | [53] | |
| Carnosic acid | Methanol extract of Salvia officialis | IC50 12 μg/mL at 36 μM | [54] | |
| Carnosol | Methanol extract of Salvia officialis | IC50 4.4 μg/mL | ||
| Roylenoic acid | Methanol extract of Salvia officialis | IC50 35 μg/mL | ||
| 7-methoxyrosmanol | Methanol extract of Salvia officialis | IC50 32 μg/mL | ||
| Triterpene oleanolic acid (Figure 1c) | Methanol extract of Salvia officialis | IC50 83 μg/mL | ||
| Crocin | Gardenia jasminoids | IC50 28.63 μmol | ||
| Lipstatin | Streptomyces toxytricini | IC50 0.14 μm | ||
| Panclicins A (Figure 1d) | Streptomyces sp. | IC502.9 μM | ||
| Panclicins B | Streptomyces sp. | IC50 2.6 μM | ||
| Panclicins C | Streptomyces sp. | IC50 0.62 μM | ||
| Panclicins D | Streptomyces sp. | IC50 0.66 μM | ||
| Panclicins E | Streptomyces sp. | IC50 0.89 μM | ||
| Valilactone | Streptomyces albolongus | IC50 0.14 nm | ||
| Ebelactones A | Streptomyces aburaviensis | IC50 3 nm/mL | ||
| Ebelactones B | Streptomyces aburaviensis | IC50 0.8 nm/mL | ||
| Esterastin | Streptomyces lavendulae | IC50 0.2 ng/mL | ||
| Vibralactone | Boreostereum virans | IC50 0.14 μg/mL | ||
| Percyquinnin (Figure 1e) | Basidiomycete stereum complicatum | IC50 2 μm |
| Compound | Source | Method | Results | Reference |
|---|---|---|---|---|
| - | Methanol extract from Rosemary | In vitro | IC50 40.9 ± 7.2 μM (commercial) | [57] |
| - | Methanol extract from Mexican oregano | IC50 37.3 ± 6.8 μM (commercial) | ||
| - | Methanol extract from Marjoram | IC50 32.4 ± 17.5 μM (commercial) | ||
| Mulberrofurans C | Morus alba | IC50 0.72 ± 0.09 μM | [59] | |
| Mulberrofurans J (Figure 1f) | Morus alba | IC50 0.60 ± 0.07 μM | ||
| Mulberrofurans F | Morus alba | IC50 0.57 ± 0.16 μM | ||
| Caffeoylquinic acid derivate chlorogenic acid | Leaves of Artemisia princeps | IC50 11.1 μM | [60] | |
| Isoderrone (Figure 1g) | Ethanol-water extract of Ficus racemosa | IC50 22.7 ± 1.7 μM | [61] | |
| Derrone | Ethanol-water extract of Ficus racemosa | IC50 12.6 ± 1.6 μM | ||
| Alpinumisoflavone | Ethanol-water extract of Ficus racemosa | IC50 21.2 ± 3.8 μM | ||
| Mucusisoflavone B | Ethanol-water extract of Ficus racemosa | IC50 2.5 ± 0.2 μM | ||
| Sulfircin | Ircinia | - | [62] | |
| 2,2′,3,3′-Tetrabromo-4,4′,5,5′-tetra-hydroxydiphenyl methane | Rhodomela confervoides | IC50 2.4 μM | ||
| 3-Bromo-4,5-Bis-(2,3-dibromo- 4,5-dihydroxybenzyl) pyrocatechol (Figure 1h) | Rhodomela confervoides | IC50 1.7 μM | ||
| Bis-(2,3-dibromo-4,5-dihydroxybenzyl) ether | Rhodomela confervoides | IC50 1.5 μM | ||
| 2,2′,3,3′-Tetrabromo-3′,4,4′,5-tetrahydroxy-6′-ethyloxymethyldiphenylmethane | Rhodomela confervoides | IC50 0.8 μM | ||
| 3,4-Dibromo-5-(2-bromo-3,4-dihydroxy-6-(ethoxymethyl)benzyl)benzene-1,2-diol | Rhodomela confervoides | IC50 0.8 μM | ||
| 3,4-Dibromo-5-(methoxymethyl)benzene-1,2-diol | Rhodomela confervoides | IC50 3.4 μM | ||
| 3-(2,3-Dibromo-4,5-dihydroxyphenyl)-2-methylpropanal | Rhodomela confervoides | IC50 4.5 μM | ||
| 3,4-Dibromo-5-(2-bromo-3,4-dihydroxy-6-(isobutoxymethyl)benzyl)benzene-1,2-diol | Rhodomela confervoides | IC50 2.4 μM | ||
| 7-Bromo-1-(2,3-dibromo-4,5-dihydroxy phenyl)- 2,3-dihydro-1H-indene-5,6-diol | Rhodomela confervoides | IC50 2.8 μM | ||
| 2,5,8-Tribromo-3-bromoamino-7-bromomethylnaphthalene | Laurencia similis | IC50 65.3 μM | ||
| 2,5,6-Tribromo-3-bromoamino-7-bromomethylnaphthalene | Laurencia similis | IC50 69.8 μM | ||
| 2′,5′,6′,5,6-Pentabromo-3′,4′,3,4-tetramethoxybenzo-phenone | Laurencia similis | IC50 2.7 μM | ||
| 2-(3′,5′-Dibromo-2′-methoxyphenoxy)-3,5-dibromophenol | Lamellodysidea herbacea | IC50 0.9 μM | ||
| 3,5-Dibromo-2-(3′,5′-dibromo-2′-methoxyphenoxy)-1-methoxybenzene | Lamellodysidea herbacea | IC50 1.7 μM | ||
| 3,5-Dibromo-2-(3′,5′-dibromo-2′ -methoxyphenoxy) phenylethanoate | Lamellodysidea herbacea | IC50 0.6 μM | ||
| 3,5-Dibromo-2-(3′,5′-dibromo-2′-methoxyphenoxy) phenylbutanoate | Lamellodysidea herbacea | IC50 0.7 μM | ||
| 3,5-Dibromo-2-(3′,5′-dibromo-2′ -methoxyphenoxy) phenylhexanoate | Lamellodysidea herbacea | IC50 0.7 μM | ||
| 3,5-Dibromo-2-(3′,5′-dibromo-2′-methoxyphenoxy) phenyl benzoate | Lamellodysidea herbacea | IC50 1 μM | ||
| 5,5′-(3-Bromo-4,5-dihydroxy-1,2-phenylene)-Bis-(methylene))Bis-(3,4-dibromobenzene-1,2-diol) | Rhodomela confervoides | IC50 1.7 μM | ||
| 3,4-Dibromo-5-(2-bromo-3,4-dihydroxy-6-(ethoxymethyl)benzyl)benzene-1,2-diol | Rhodomela confervoides | IC50 0.84 μM | ||
| 2-(30,50-Dibromo-20-methoxyphenoxy)-3,5-dibromophenol | Lamellodysidea herbacea | IC50 0.9 μM | ||
| 2-(30,50-Dibromo-20-methoxyphenoxy)-3,5-dibromophenol-methyl ether | Lamellodysidea herbacea | IC50 1.7 μM | ||
| 2,3,6-Tribromo-4,5-dihydroxybenzyl methyl ether | Symphyocladia latiuscula | IC50 3.9 μM | ||
| Bis-(2,3,6-tribromo-4,5-dihydroxyphenyl) methane | Symphyocladia latiuscula | IC50 4.3 μM | ||
| 1,2-Bis-(2,3,6-tribromo-4,5-dihydroxyphenyl)-ethane | Symphyocladia latiuscula | IC50 2.7 μM | ||
| 3′,5′,6′,6-Tetrabromo-2,4-dimethyldiphenyl ether | Laurencia similis | IC50 3 μM | ||
| 2′,5′,6′,5,6-Pentabromo-3′,4′,3,4-tetramethoxybenzo-phenone | Laurencia similis | IC50 2.7 μM | ||
| 3′,5′,6′6-Tetrabromo-2,4-dimethyldiphenyl ether | Laurencia similis | IC50 3 μM | ||
| 1,2,5-Tribromo-3-bromoamino-7-bromomethylnaphthalene | Laurencia similis | IC50 102 μM |
| Compound | Source | Method | Results | Reference |
|---|---|---|---|---|
| Caseins | Protein hydrolysates cow’s milk | In silico | Occurrence frequency 0.249 (beta-casein), 0.380 (bovine meat) and 0.305 (salmon). Gly-Ala, Gly-Pro and Pro-Gly were the most frequently occurring sequences | [69] |
| Collagens | Protein hydrolysates bovine meat and salmon | |||
| Gly-Pro-Gly-Ala | Protein hydrolysates Atlantic salmon skin gelatin | In vitro | IC50 49.6 μM | [66] |
| Gly-Pro-Ala-Glu | IC50 41.9 μM | |||
| Pro-Gly-Val-Gly-Gly-Pro-Leu-Gly-Pro-Ile-Gly-Pro-Cys-Tyr | Protein hydrolysates tuna cooking juice | IC50 116 μM | ||
| Cys-Ala-Tyr-Gln-Trp-Gln-Arg-Pro-Val-Asp-Arg-Ile-Arg | IC50 78 μM | |||
| Pro-Ala-Cys-Gly-Gly-Phe-Tyr-Ile-Ser-Gly-Arg-Pro-Gly | IC50 96.4 μM | |||
| Leu-Pro | Protein hydrolysates Japanese rice bran | IC50 2400 μM | ||
| Ile-Pro | IC50 410 μM | |||
| Met-Pro | IC50 870 μM | |||
| Val-Pro | IC50 880 μM | |||
| Arg-Pro | IC50 2240 μM | |||
| Thr-Pro | IC50 2370 μM | |||
| Leu-Pro | IC50 2370 μM | |||
| Lys-Pro | Protein hydrolysates Japanese rice bran | IC50 2540 μM | ||
| His-Pro | IC50 2820 μM | |||
| Tyr-Pro | IC50 3170μM | |||
| Phe-Pro | IC50 3630 μM | |||
| Trp-Pro | IC50 4530 μM | |||
| Pro-Pro | IC50 5860 μM | |||
| Ser-Pro | IC50 5980 μM | |||
| Ala-Pro | IC50 7950 μM | |||
| Leu-Pro-Gln-Asn-Ile-Pro-Pro-Leu | Protein hydrolysates gouda cheese | IC50 46 μM | ||
| Leu-Pro-Gln-Asn-Ile-Pro-Pro | IC50 160μM | |||
| Pro-Gln-Asn-Ile-Pro-Pro-Leu | IC50 1500 μM | |||
| Leu-Pro-Gln | IC50 82 μM | |||
| Val-Pro-Ile-Thr-Pro-Thr | IC50 130 μM | |||
| Val-Pro-Ile-Thr-Pro-Thr-Leu | IC50 110 μM | |||
| Phe-Pro-Gly-Pro-Ile-Pro-Asp | IC50 260 μM | |||
| Pro-Gly-Pro-Ile-His-Asp-Ser | IC50 1000 μM | |||
| Ile-Pro-Pro-Leu-The-Gln-Thr-Pro-Val | IC50 1300 μM | |||
| Val-Pro-Pro-Phe-Ile-Gln-Pro-Glu | IC50 2500 μM | |||
| Tyr-Pro-Phe-Pro-Gly-Pro-Ile-Pro-Asp | IC50 670 μM | |||
| Val-Ala-Gly-Thr-Trp-Tyr | Protein hydrolysates β-lactoglobulin | IC50 174 μM | ||
| Ile-Pro-Ala | IC50 49 μM | |||
| Ile-Pro-Ala-Val-Phe | IC50 45 μM | |||
| Ile-Pro-Ala-Val-Phe-Lys | IC50 143 μM | |||
| Val-Leu-Val-Leu-Asp-Thr-Asp-Tyr-Lys | IC50 424 μM | |||
| Thr-Pro-Glu-Val-Asp-Asp-Glu-Ala-Leu-Glu-Lys | IC50 320 μM | |||
| Glu-Lys | Protein hydrolysates milk protein | IC50 3216 μM | ||
| Gly-Leu | IC50 2615 μM | |||
| Ala-Leu | IC50 882 μM | |||
| Val-Ala | IC50 168 μM | |||
| Trp-Val | IC50 65 μM | |||
| Phe-Leu | IC50 399 μM | |||
| His-Leu | IC50 143 μM | |||
| Ser-Leu | IC50 2517 μM | |||
| Trp-Val | Synthetic dipeptides | IC50 0.020 ± 0.001 mg/mL | [70] | |
| Lactoferrin hydrolysate LFH1 | Milk | IC50 1.088 ± 0.106 mg/mL | ||
| Casein hydrolysate CasH2 | IC50 0.882 ± 0.057 mg/mL | |||
| Cirsimaritin, naringenin, hispidulin, eriodictyol and carnosol | Methanol extract of Rosemary | IC50 28.7 ± 3.1 μM (greenhouse-grown) 6.5 ± 0.4 μM (commercial) | [57] | |
| Methanol extract of Mexican oregano | IC50 25.3 ± 0.3 μM (greenhouse-grown) 3.9 ± 0.6 μM (commercial) | |||
| - | Methanol extract of Marjoram | IC50 37.7 ± 7.9 μM (greenhouse-grown) | ||
| Cirsimaritin | Purified compounds | IC50 0.43 ± 0.07 μM | ||
| Hispidulin | IC50 0.49 ± 0.06 μM | |||
| Naringenin | IC50 2.5 ± 0.29 μM | |||
| Anthocyanins | Blueberry-blackberry wine blends | IC50 0.07 ± 0.02 to >300 μM | [71] | |
| Resveratrol (Figure 1i) | Commonly found in citrus, berry, grape and soybean | IC50 0.6 ± 0.4 nM | ||
| Luteolin | IC50 0.12 ± 0.01 μM | |||
| Apigenin | IC50 0.14 ± 0.02 μM | |||
| Flavone (Figure 1j) | IC50 0.17 ± 0.01 μM | |||
| - | Ethanolic precipitate of Sargassum binderi | IC50 2.194 mg/mL | [64] | |
| - | Ethanolic precipitate of Padina sulcata | IC50 2.306 mg/mL | ||
| - | Ethanolic precipitate of Turbinaria conoides | IC50 3.594 mg/mL | ||
| - | Methanol extract of Sargassum wightii | IC50 38.27 μg/mL | [65] | |
| - | Methanol extract of Sargassum polycystum | IC50 36.94 μg/mL | ||
| - | Methanol extract of Turbinaria conoides | IC50 55.2 μg/mL | [72] | |
| Mangiferin | Mangifera indica | Rat model and ELISA | 89 ± 8% | [68] |
| - | Crude extract of marine sponge Xetospongia muta | In vitro | IC50 0.8 mg/mL (preincubation time: 10 min) Treatment with 2.5% TCA or heat (60 °C) increase 3.7 and 2.7 total inhibitory activity | [73] |
| - | Crude extracts of marine anemone Bunodosoma granulifera | IC50 1.2 mg/mL (preincubation time: 10 min) thermal treatment destroyed inhibitory activity | ||
| - | Crude extracts of marine anemone Bartholomea annulata | IC50 0.38 mg/mL (preincubation time: 3 min) heat or TCA treatment decreased inhibitory activity |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duarte, A.M.; Guarino, M.P.; Barroso, S.; Gil, M.M. Phytopharmacological Strategies in the Management of Type 2 Diabetes Mellitus. Foods 2020, 9, 271. https://doi.org/10.3390/foods9030271
Duarte AM, Guarino MP, Barroso S, Gil MM. Phytopharmacological Strategies in the Management of Type 2 Diabetes Mellitus. Foods. 2020; 9(3):271. https://doi.org/10.3390/foods9030271
Chicago/Turabian StyleDuarte, Ana M., Maria P. Guarino, Sónia Barroso, and Maria M. Gil. 2020. "Phytopharmacological Strategies in the Management of Type 2 Diabetes Mellitus" Foods 9, no. 3: 271. https://doi.org/10.3390/foods9030271
APA StyleDuarte, A. M., Guarino, M. P., Barroso, S., & Gil, M. M. (2020). Phytopharmacological Strategies in the Management of Type 2 Diabetes Mellitus. Foods, 9(3), 271. https://doi.org/10.3390/foods9030271