Phytochemicals of Minthostachys diffusa Epling and Their Health-Promoting Bioactivities

 , ,
, ,  , ,
, ,  , and
, and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Chemicals
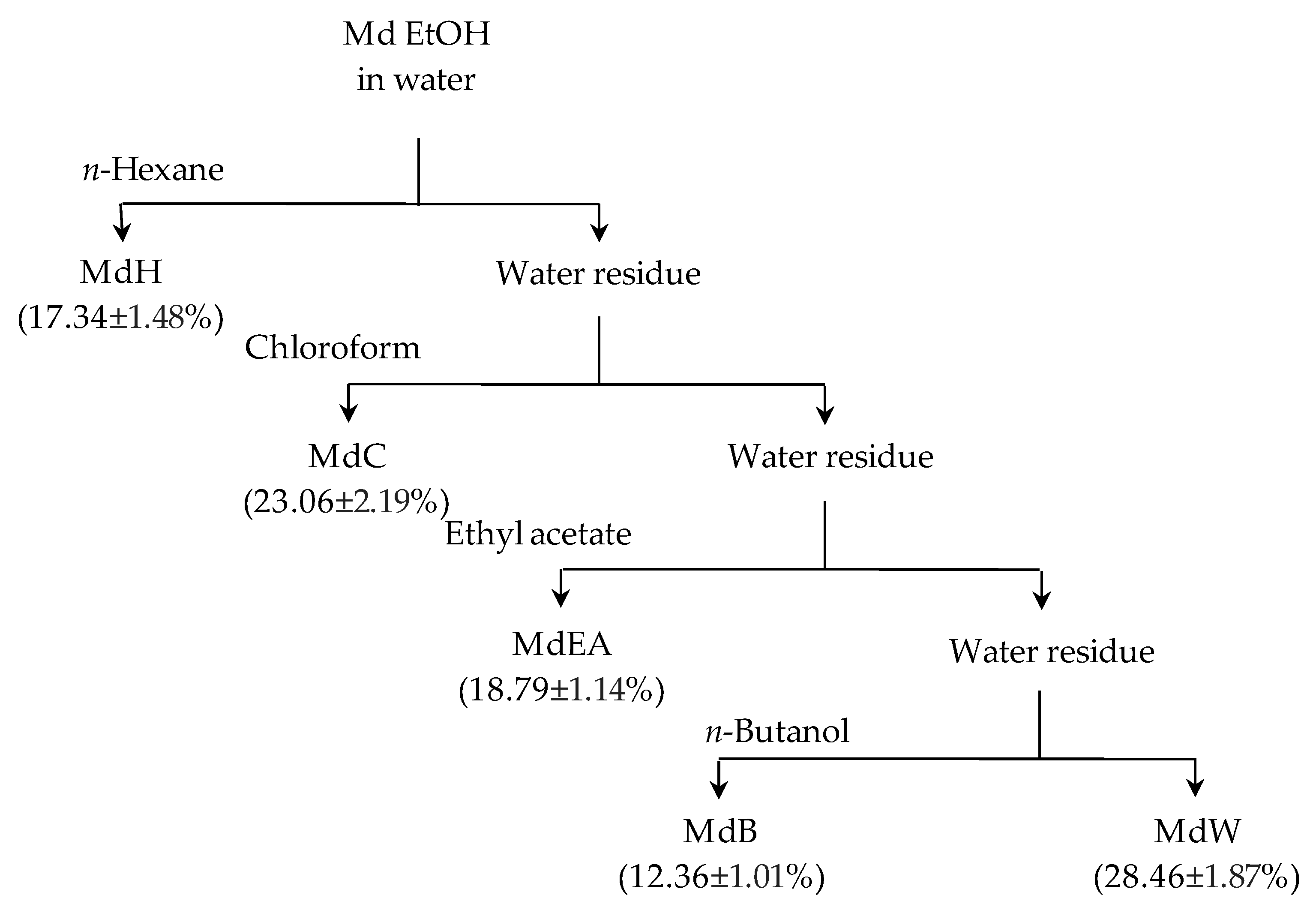
2.2. Plant Material and Samples Preparation
2.3. Total Phenolic, Flavonoid and Terpenoid Contents
2.4. Antioxidant Activity
2.4.1. Radical Scavenging Activity
2.4.2. Ferric Reducing Antioxidant Power Assay (FRAP)
2.4.3. β-carotene Bleaching Assay
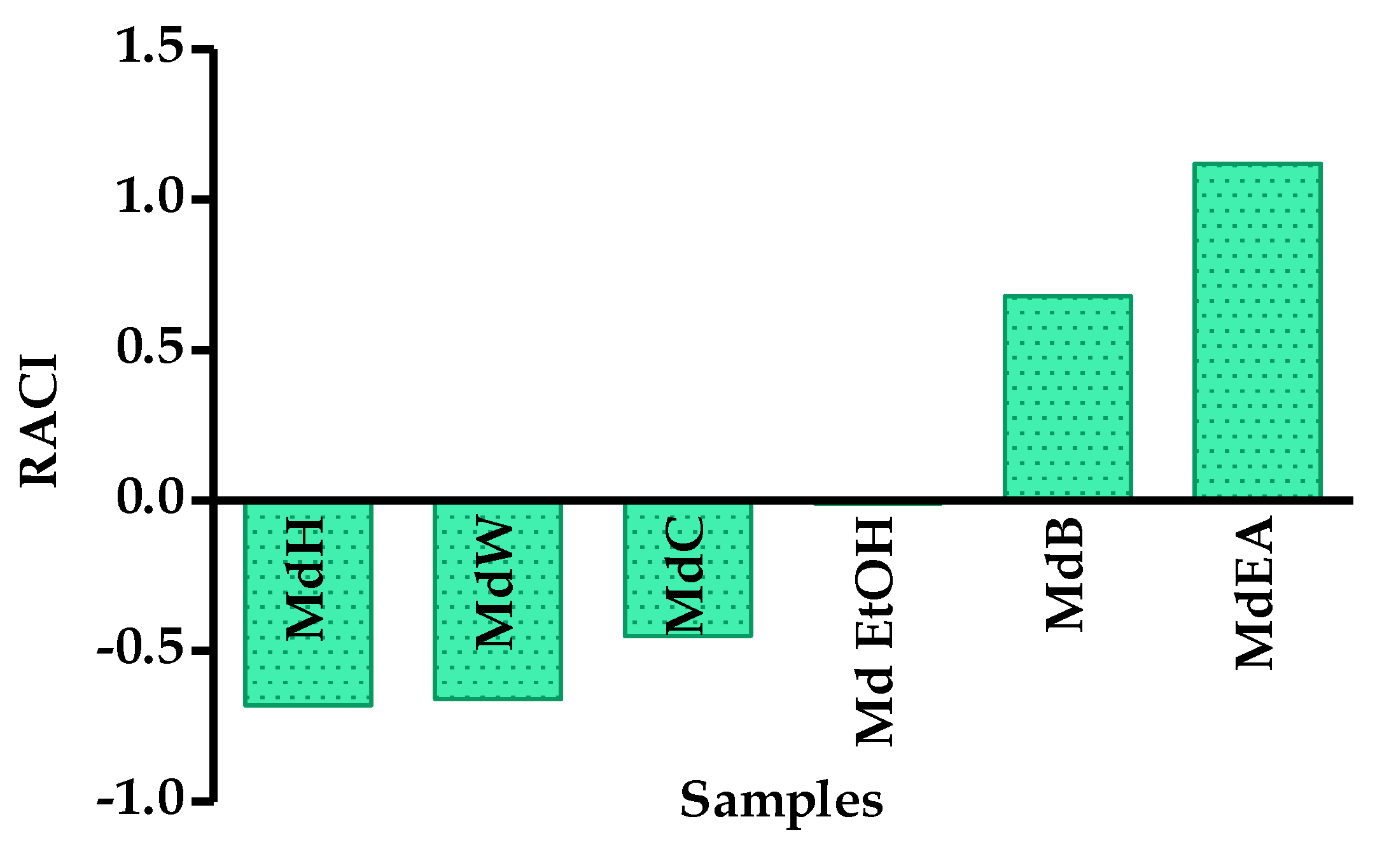
2.4.4. Relative Antioxidant Capacity Index (RACI)
2.5. Potential Antidiabetic Activity
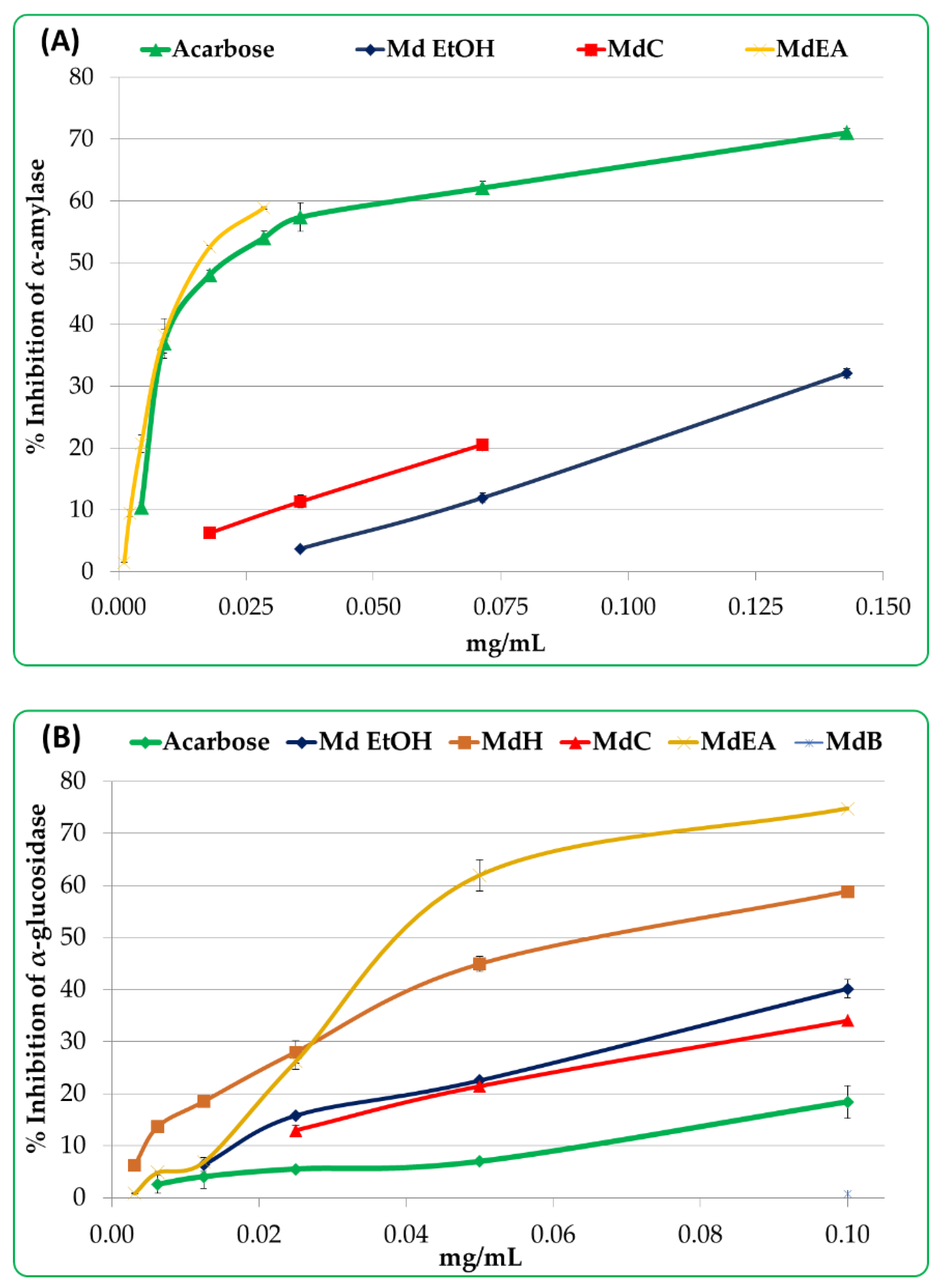
2.5.1. α-amylase Inhibition
2.5.2. α-glucosidase Inhibition
2.6. Anticholinesterase Activity
2.7. Liquid Chromatography Tandem Mass Spectrometry Analysis of Polyphenols
2.8. Molecular Docking
2.9. Statistical Analysis
3. Results and Discussion
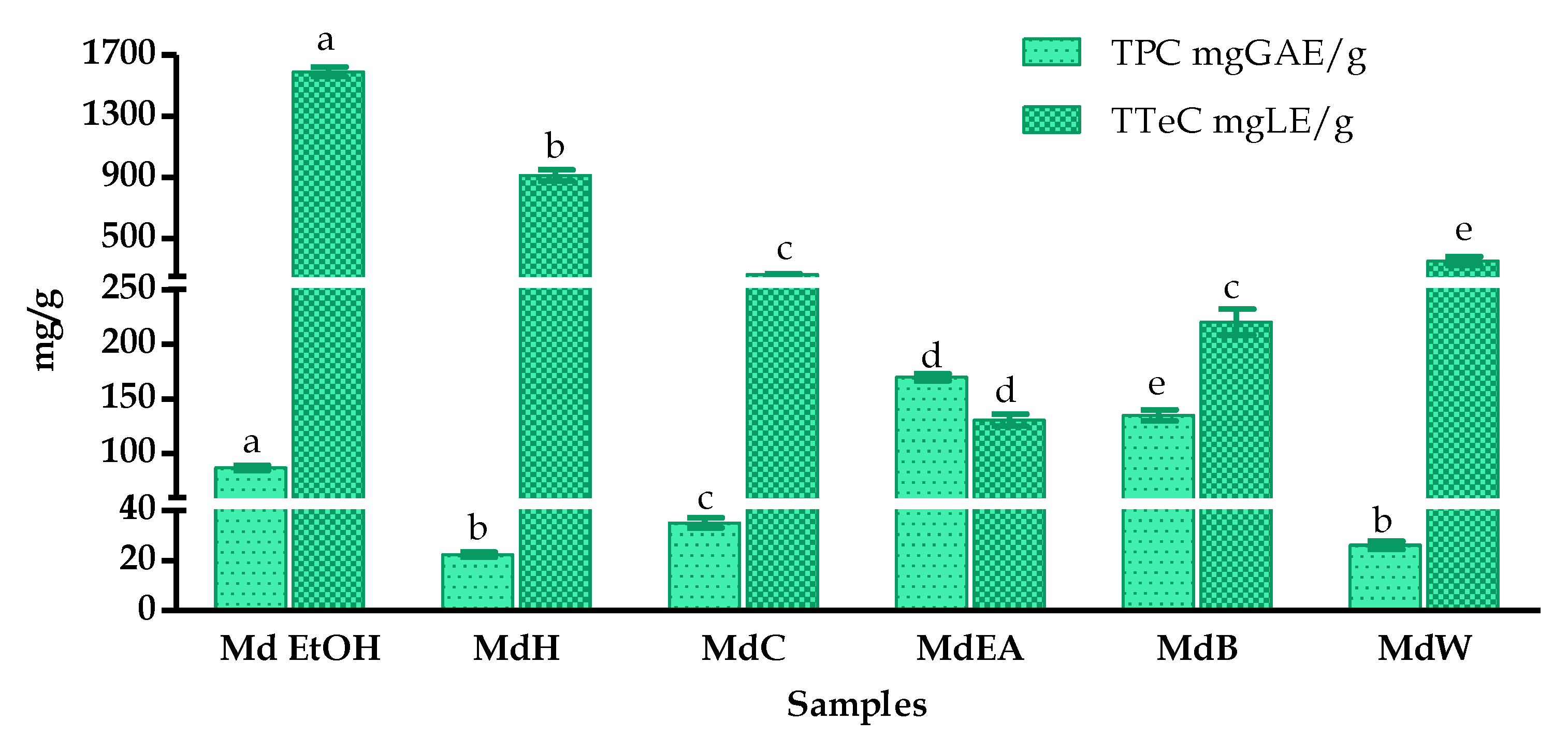
3.1. Extraction Yield and Influence of Solvents on Total Polyphenolic, Flavonoid and Terpenoid Contents
3.2. Antioxidant Activity
3.3. Potential Antidiabetic Activity
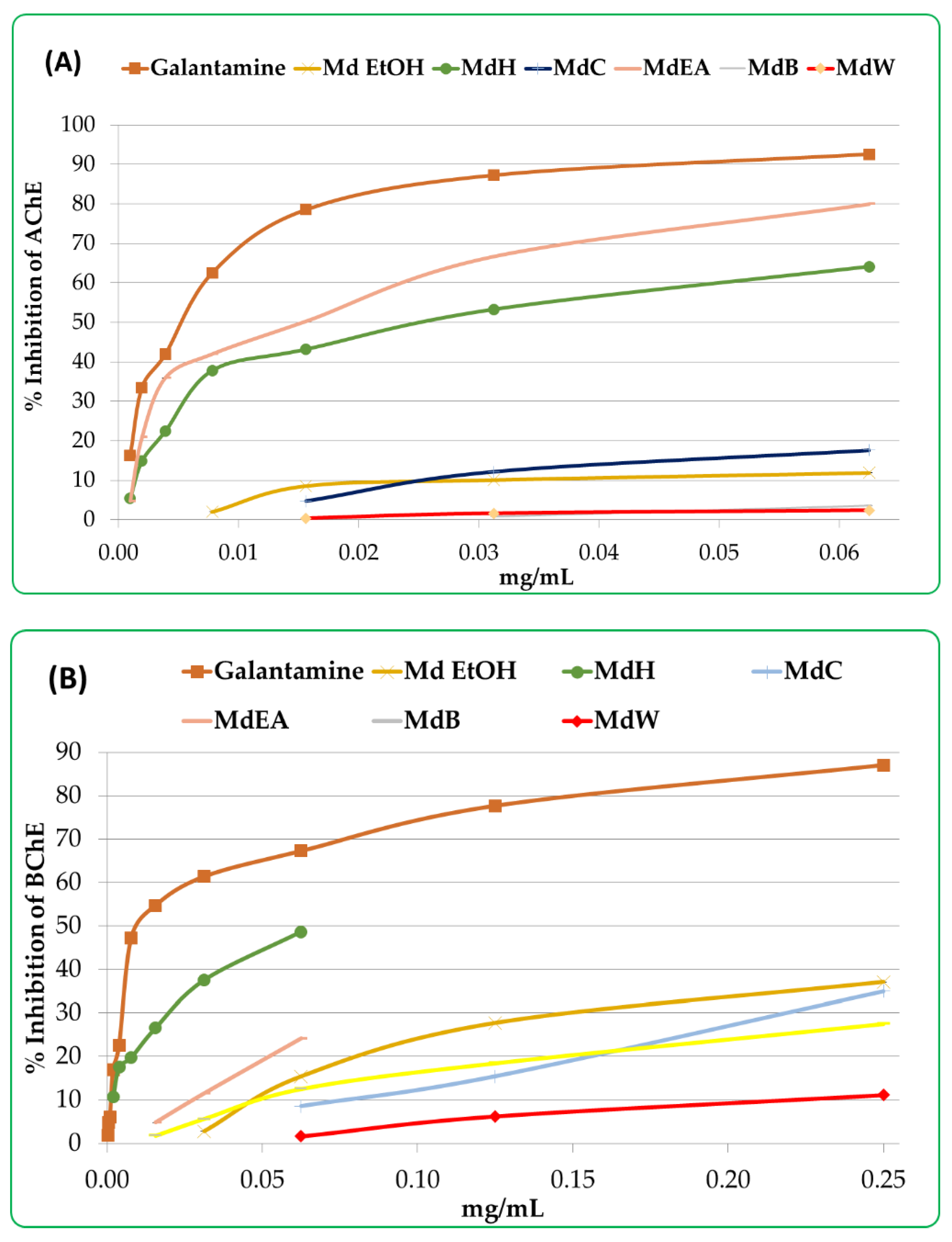
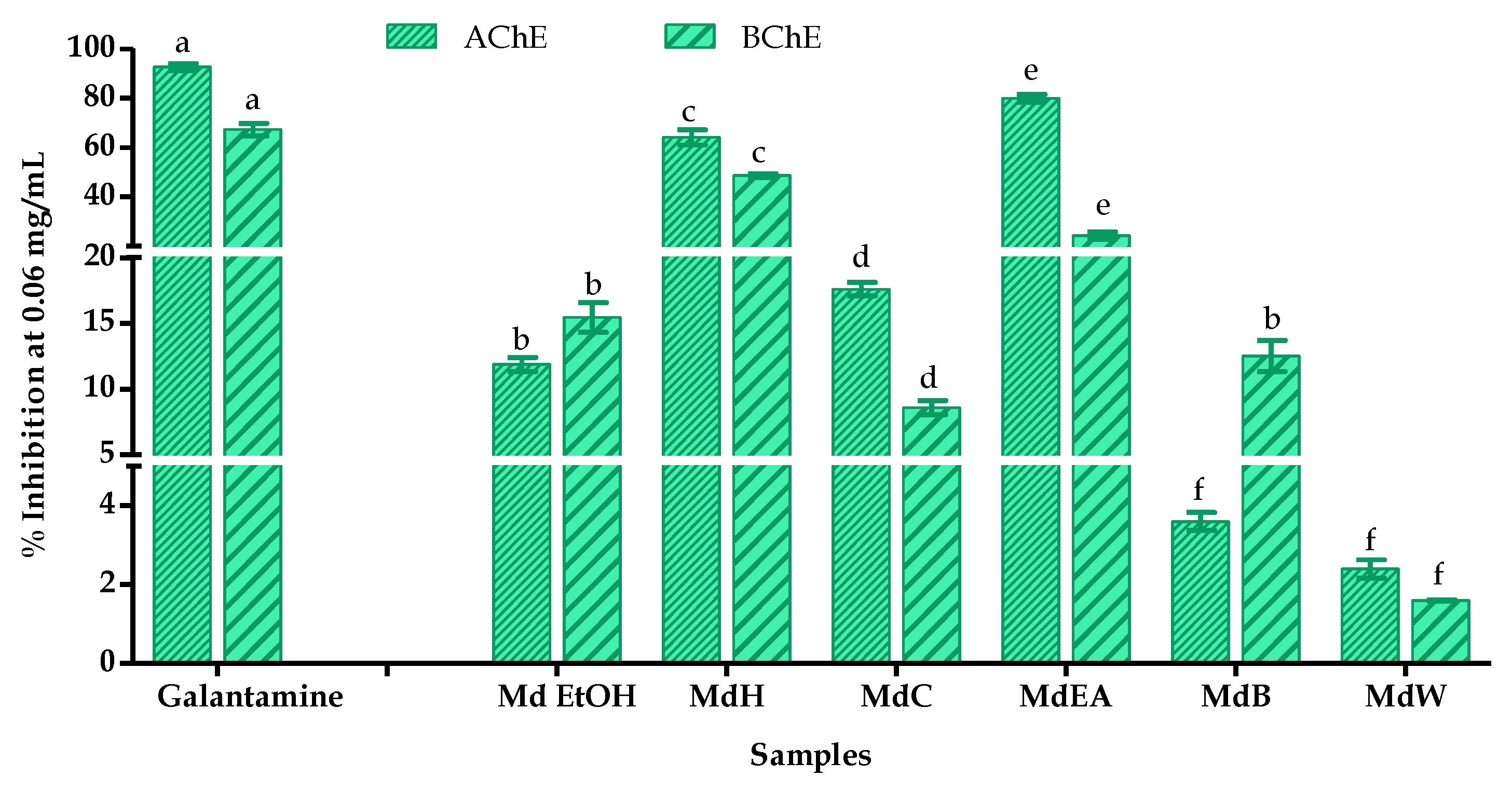
3.4. Determination of Anticholinesterase Activity
3.5. Identification and Quantification of Phytochemicals by Liquid Chromatography-Mass Spectrometry
3.6. Potential Anticholinesterase Activity of Identified Triterpenes from MdEA and MdH Fractions
3.7. Molecular Docking of the Identified Terpenes into AChE and BChE
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Buchanan, B.B.; Gruissem, W.; Jones, R.L. Biochemistry & Molecular Biology of Plants; American Society of Plant Physiologists: Rockville, MD, USA, 2000; Volume 40. [Google Scholar]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of plant secondary metabolites: A historical perspective. Plant Sci. 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Pinakin, D.J.; Kumar, V.; Suri, S.; Sharma, R.; Kaushal, M. Nutraceutical potential of tree flowers: A comprehensive review on biochemical profile, health benefits, and utilization. Food Res. Int. 2020, 127, 108724. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.-Y.; Meng, X.; Gan, R.-Y.; Zhao, C.-N.; Liu, Q.; Feng, Y.-B.; Li, S.; Wei, X.-L.; Atanasov, A.G.; Corke, H.; et al. Health functions and related molecular mechanisms of tea components: An update review. Int. J. Mol. Sci. 2019, 20, 6196. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Zidorn, C. The genus Tragopogon (asteraceae): A review of its traditional uses, phytochemistry, and pharmacological properties. J. Ethnopharmacol. 2020, 250, 112466. [Google Scholar] [CrossRef]
- Schmidt-Lebuhn, A.N. Ethnobotany, biochemistry and pharmacology of Minthostachys (Lamiaceae). J. Ethnopharmacol. 2008, 118, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Lock, O.; Perez, E.; Villar, M.; Flores, D.; Rojas, R. Bioactive compounds from plants used in Peruvian traditional medicine. Nat. Prod. Commun. 2016, 11, 315–337. [Google Scholar] [PubMed]
- Cariddi, L.; Escobar, F.; Moser, M.; Panero, A.; Alaniz, F.; Zygadlo, J.; Sabini, L.; Maldonado, A. Monoterpenes isolated from Minthostachys verticillata (Griseb.) Epling essential oil modulates immediate-type hypersensitivity responses in vitro and in vivo. Planta Med. 2011, 77, 1687–1694. [Google Scholar] [CrossRef]
- Montironi, I.D.; Cariddi, L.N.; Reinoso, E.B. Evaluation of the antimicrobial efficacy of Minthostachys verticillata essential oil and limonene against Streptococcus uberis strains isolated from bovine mastitis. Rev. Argent. Microbiol. 2016, 48, 210–216. [Google Scholar] [CrossRef]
- Mora, F.D.; Araque, M.; Rojas, L.B.; Ramirez, R.; Silva, B.; Usubillaga, A. Chemical composition and in vitro antibacterial activity of the essential oil of Minthostachys mollis (Kunth) Griseb Vaught from the Venezuelan Andes. Nat. Prod. Commun. 2009, 4, 997–1000. [Google Scholar] [CrossRef]
- Cantín, Á.; Lull, C.; Primo, J.; Miranda, M.A.; Primo-Yúfera, E. Isolation, structural assignment and insecticidal activity of (−)-(1S,2R,3R,4S)-1,2-Epoxy-1-methyl-4-(1-methylethyl)-cyclohex-3-yl acetate, a natural product from Minthostachys tomentosa. Tetrahedron Asymmetry 2001, 12, 677–683. [Google Scholar]
- Roberts, R.O.; Knopman, D.S.; Przybelski, S.A.; Mielke, M.M.; Kantarci, K.; Preboske, G.M.; Senjem, M.L.; Pankratz, V.S.; Geda, Y.E.; Boeve, B.F.; et al. Association of type 2 diabetes with brain atrophy and cognitive impairment. Neurology 2014, 82, 1132. [Google Scholar] [CrossRef]
- Aguirre-Acevedo, D.C.; Lopera, F.; Henao, E.; Tirado, V.; Muñoz, C.; Giraldo, M.; Bangdiwala, S.I.; Reiman, E.M.; Tariot, P.N.; Langbaum, J.B.; et al. Cognitive decline in a colombian kindred with autosomal dominant Alzheimer disease: A retrospective cohort study. JAMA Neurol. 2016, 73, 431–438. [Google Scholar] [CrossRef]
- Redondo, M.T.; Beltrán-Brotóns, J.L.; Reales, J.M.; Ballesteros, S. Executive functions in patients with Alzheimer’s disease, type 2 diabetes mellitus patients and cognitively healthy older adults. Exp. Gerontol. 2016, 83, 47–55. [Google Scholar] [CrossRef]
- Faraone, I.; Rai, D.; Chiummiento, L.; Fernandez, E.; Choudhary, A.; Prinzo, F.; Milella, L. Antioxidant activity and phytochemical characterization of Senecio clivicolus Wedd. Molecules 2018, 23, 2497. [Google Scholar] [CrossRef]
- Lamorte, D.; Faraone, I.; Laurenzana, I.; Milella, L.; Trino, S.; De Luca, L.; Del Vecchio, L.; Armentano, M.; Sinisgalli, C.; Chiummiento, L. Future in the past: Azorella glabra Wedd. as a source of new natural compounds with antiproliferative and cytotoxic activity on Multiple Myeloma cells. Int. J. Mol. Sci. 2018, 19, 3348. [Google Scholar] [CrossRef]
- Faraone, I.; Rai, D.K.; Russo, D.; Chiummiento, L.; Fernandez, E.; Choudhary, A.; Milella, L. Antioxidant, Aantidiabetic, and Aanticholinesterase Aactivities and Pphytochemical Pprofile of Azorella glabra Wedd. Plants 2019, 8, 265. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Lin, J.-Y.; Tang, C.-Y. Determination of total phenolic and flavonoid contents in selected fruits and vegetables, as well as their stimulatory effects on mouse splenocyte proliferation. Food Chem. 2007, 101, 140–147. [Google Scholar] [CrossRef]
- Chakraborty, S.; Guchhait, S.; Saha, S.; Biswas, S. Estimation of total terpenoids concentration in plant tissues using a monoterpene, linalool as standard reagent. Protoc. Exch. 2012, 5, 1038. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Stratil, P.; Klejdus, B.; Kubáň, V. Determination of total content of phenolic compounds and their antioxidant activity in vegetables evaluation of spectrophotometric methods. J. Agric. Food Chem. 2006, 54, 607–616. [Google Scholar] [CrossRef]
- Fidelis, Q.C.; Faraone, I.; Russo, D.; Aragão Catunda, F.E., Jr.; Vignola, L.; de Carvalho, M.G.; De Tommasi, N.; Milella, L. Chemical and biological insights of Ouratea hexasperma (A. St.-Hil.) Baill.: A source of bioactive compounds with multifunctional properties. Nat. Prod. Res. 2019, 33, 1500–1503. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Krieger, E.; Joo, K.; Lee, J.; Lee, J.; Raman, S.; Thompson, J.; Tyka, M.; Baker, D.; Karplus, K. Improving physical realism, stereochemistry, and side-chain accuracy in homology modeling: Four approaches that performed well in CASP8. Proteins Struct. Funct. Bioinform. 2009, 77, 114–122. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Palacios, S.M.; del Corral, S.; Carpinella, M.C.; Ruiz, G. Screening for natural inhibitors of germination and seedling growth in native plants from Central Argentina. Ind. Crops Prod. 2010, 32, 674–677. [Google Scholar] [CrossRef]
- Vaquero, M.R.; Serravalle, L.T.; De Nadra, M.M.; De Saad, A.S. Antioxidant capacity and antibacterial activity of phenolic compounds from Argentinean herbs infusions. Food Control 2010, 21, 779–785. [Google Scholar] [CrossRef]
- Solis-Quispe, L.; Tomaylla-Cruz, C.; Callo-Choquelvica, Y.; Solís-Quispe, A.; Rodeiro, I.; Hernández, I.; Fernández, M.D.; Pino, J.A. Chemical composition, antioxidant and antiproliferative activities of essential oil from Schinus areira L. and Minthostachys spicata (Benth.) Epl. grown in Cuzco, Peru. J. Essent. Oil Res. 2016, 28, 234–240. [Google Scholar] [CrossRef]
- Carpinella, M.C.; Andrione, D.G.; Ruiz, G.; Palacios, S.M. Screening for acetylcholinesterase inhibitory activity in plant extracts from Argentina. Phytother. Res. 2010, 24, 259–263. [Google Scholar] [CrossRef]
- Savelev, S.U.; Okello, E.J.; Perry, E.K. Butyryl-and acetyl-cholinesterase inhibitory activities in essential oils of Salvia species and their constituents. Phytother. Res. 2004, 18, 315–324. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Kumar, V.; Mal, M.; Houghton, P.J. Acetylcholinesterase inhibitors from plants. Phytomedicine 2007, 14, 289–300. [Google Scholar] [CrossRef]
- Alkire, B.H.; Tucker, A.O.; Maciarello, M.J. Tipo, Minthostachys mollis (lamiaceae): An Ecuadorian mint. Econ. Bot. 1994, 48, 60–64. [Google Scholar] [CrossRef]
- Senatore, F. Composition of the essential oil of Minthostachys spicata (Benth.) Epl. Flavour Fragr. J. 1995, 10, 43–45. [Google Scholar] [CrossRef]
- López-Lázaro, M. Distribution and biological activities of the flavonoid luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Uma Devi, P.; Ganasoundari, A.; Vrinda, B.; Srinivasan, K.; Unnikrishnan, M. Radiation protection by the Ocimum flavonoids orientin and vicenin: Mechanisms of action. Radiat. Res. 2000, 154, 455–460. [Google Scholar] [CrossRef]
- Leal, A.S.M. Preparation and biological evaluation of new triterpene derivates of ursolic and oleanolic acid. Ph.D. Thesis, Universidade de Coimbra, Coimbra, Portugal, 2012. [Google Scholar]
- Sultana, N. Clinically useful anticancer, antitumor, and antiwrinkle agent, ursolic acid and related derivatives as medicinally important natural product. J. Enzym. Inhib. Med. Chem. 2011, 26, 616–642. [Google Scholar] [CrossRef]
- Jamila, N.; Khairuddean, M.; Yeong, K.K.; Osman, H.; urugaiyah, V. Cholinesterase inhibitory triterpenoids from the bark of Garcinia hombroniana. J. Enzym. Inhib. Med. Chem. 2015, 30, 133–139. [Google Scholar] [CrossRef]
- Ruhal, P.; Dhingra, D. Ameliorative effect of betulinic acid on ageing and scopolamine-induced learning and memory impairment in rats. Asian J. Pharm. Pharmacol. 2018, 4, 825–841. [Google Scholar] [CrossRef]
- Öztürk, M.; Kolak, U.; Topçu, G.; Öksüz, S.; Choudhary, M.I. Antioxidant and anticholinesterase active constituents from Micromeria cilicica by radical-scavenging activity-guided fractionation. Food Chem. 2011, 126, 31–38. [Google Scholar] [CrossRef]
- Bahadori, M.B.; Dinparast, L.; Valizadeh, H.; Farimani, M.M.; Ebrahimi, S.N. Bioactive constituents from roots of Salvia syriaca L.: Acetylcholinesterase inhibitory activity and molecular docking studies. South Afr. J. Bot. 2016, 106, 1–4. [Google Scholar] [CrossRef]
- Geromichalos, G.D.; Lamari, F.N.; Papandreou, M.A.; Trafalis, D.T.; Margarity, M.; Papageorgiou, A.; Sinakos, Z. Saffron as a source of novel acetylcholinesterase inhibitors: Molecular docking and in vitro enzymatic studies. J. Agric. Food Chem. 2012, 60, 6131–6138. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | ABTS (mgTE/g) | DPPH (mgTE/g) | SO IC25 (mg/mL) | FRAP (mgTE/g) | BCB %AA |
|---|---|---|---|---|---|
| Md EtOH | 146.07 ± 4.20 a | 113.05 ± 3.51 a | 0.92 ± 0.08 a | 224.67 ± 3.37 a | 10.12 ± 0.52 a |
| MdH | nc | nc | nc | 27.27 ± 0.99 b | 11.01 ± 0.33 a |
| MdC | 29.61 ± 0.74 b | 17.64 ± 0.03 b | 1.03 ± 0.08 a,b | 22.18 ± 0.90 b | 13.03 ± 0.49 b |
| MdEA | 444.76 ± 28.24 c | 281.64 ± 7.93 c | 0.62 ± 0.04 c | 574.86 ± 9.14 c | 19.13 ± 0.93 c |
| MdB | 218.56 ± 9.38 d | 122.30 ± 2.77 a | 0.26 ± 0.01 d | 297.75 ± 5.71 d | nc |
| MdW | 45.85 ± 0.42 b | 37.01 ± 1.63 d | 1.14 ± 0.09 b | 64.67 ± 1.63 e | nc |
| TPC | TTeC | ABTS | DPPH | SO | NO | FRAP | BCB | |
|---|---|---|---|---|---|---|---|---|
| TPC | 1.00 | |||||||
| TTeC | −0.26 | 1.00 | ||||||
| ABTS | 0.96 | −0.31 | 1.00 | |||||
| DPPH | 0.94 | −0.23 | 0.99 | 1.00 | ||||
| SO | 0.69 | −0.40 | 0.50 | 0.44 | 1.00 | |||
| NO | 0.44 | −0.31 | 0.21 | 0.13 | 0.91 | 1.00 | ||
| FRAP | 0.96 | −0.24 | 1.00 | 0.99 | 0.50 | 0.22 | 1.00 | |
| BCB | 0.26 | 0.06 | 0.40 | 0.43 | −0.40 | −0.58 | 0.38 | 1.00 |
| Pk. no. | RT (min) | [M-H]−m/z Observed | [M-H]−m/z Calculated | Predicted Molecular Formula | MS/MS (m/z) | Compound Identity | mg/g DW |
|---|---|---|---|---|---|---|---|
| 1 | 6.18 | 179.0367 | 179.0344 | C9H8O4 | 135, 79 | Caffeic acid | 2.49 ± 0.01 |
| 2 | 6.84 | 461.1769 | 461.1753 | C31H26O4 | 329, 301 | unknown | nq |
| 3 | 6.71 | 625.1441 | 625.1405 | C27H30O17 | 343, 301, 271, 255, 179, 151 | Quercetin-3,4′-di-glucoside | 0.03 ± 0.02 |
| 4 | 7.17 | 609.1473 | 609.1456 | C27H30O16 | 300, 285, 271, 255, 179, 151 | Rutin | 1.63 ± 0.07 |
| 5 | 7.37 | 463.0876 | 463.0877 | C21H20O12 | 300, 271, 255, 179, 151 | Quercetin-3-O-glucoside | 22.87 ± 0.25 |
| 6 | 7.67 | 505.0982 | 505.098 | C23H22O13 | 300, 271, 255, 179, 161, 151 | Quercetin derivative | Nq |
| 7 | 7.70 | 433.0742 | 433.0771 | C20H18O11 | 300, 271, 255, 179,151 | Quercetin-3-O-arabinoside | 1.92 ± 0.02 |
| 8 | 7.87 | 447.0921 | 447.0927 | C21H20O11 | 284, 255, 227 | Kaempferol-3-O-glucoside | 2.07± 0.15 |
| 9 | 8.00 | 609.1848 | 609.1819 | C28H34O15 | 325, 301, 286, 242, 199, 164, 125 | Hesperidin | 1.10 ± 0.09 |
| 10 | 431.0981 | 431.0978 | C21H20O10 | 269, 239, 224 | Apigenin-7-O-glycoside | 0.54 ± 0.05 | |
| 11 | 433.1125 | 433.1135 | C21H22O10 | 151, 107 | Naringenin-7-O-glucoside | 0.17 ± 0.01 | |
| 12 | 8.24 | 359.0766 | 359.0767 | C18H16O8 | 197, 179, 161, 135, 133, 123, 73 | Rosmarinic acid | 69.64 ± 1.53 |
| 13 | 8.73 | 609.1457 | 609.1456 | C27H30O16 | 463, 323, 300, 285, 271, 255, 179, 161, 151 | Quercetin-O-glucoside- rhamnoside | nq |
| 14 | 8.79 | 533.1882 | 533.1870 | C23H34O14 | 387, 374, 207, 163, 145, 119, 101 | Coumaric acid derivative | nq |
| 15 | 471.1216 | 471.1232 | C31H20O5 | 307, 205, 163, 145, 119, 101 | Coumaric acid derivative | nq | |
| 16 | 163.0392 | 163.0395 | C9H8O3 | 119, 93 | 4-Coumaric acid | 0.23 ± 0.02 | |
| 17 | 9.15 | 593.1697 | 593.1506 | C27H30O15 | 327, 309, 285, 270, 241, 164, 151 | Luteolin-rutinoside | nq |
| 18 | 285.0384 | 285.0399 | C15H10O6 | 151, 133 | Luteolin | 1.00 ± 0.00 | |
| 19 | 9.39 | 301.0362 | 301.0348 | C15H10O7 | 179, 151 | Quercetin | 2.07 ± 0.09 |
| 20 | 9.69 | 373.0908 | 373.0923 | C19H18O8 | 197, 179, 161, 135, 117, 107 | Rosmarinic acid methyl ester | nq |
| 21 | 11.34 | 487.3447 | 487.3424 | C30H48O5 | 469, 441, 405, 397,389, 85, 73 | Asiatic acid type | nq |
| 22 | 11.50 | 487.3447 | 487.3424 | C30H48O5 | 469, 441, 405, 397,389, 85, 73 | Asiatic acid type | nq |
| 23 | 12.03 | 829.4156 | 829.4163 | C48H62O12 | 811, 789, 667, 649, 553, 359, 179, 161, 135 | Rosmarinic acid derivative | nq |
| 24 | 12.82 | 471.3475 | 471.3474 | C30H48O4 | 427, 425, 409, 353, 337, 57 | Corosolic type triterpenoid | nq |
| 25 | 13.81 | 501.3549 | 501.3580 | C31H50O5 | 469, 421, 407, 389 | Asiatic acid methyl ester | nq |
| 26 | 14.18 | 813.4201 | 813.4214 | C48H62O11 | 651, 453, 359, 197, 179, 161, 135, 73 | Rosmarinic acid derivative | nq |
| 27 | 14.51 | 455.3527 | 455.3525 | C30H48O3 | 411, 393, 381, 351, 83, 71, 57 | Oleanolic type triterpenoid | nq |
| 28 | 15.22 | 469.3342 | 469.3318 | C30H46O4 | 451, 425, 421, 407, 391, 377, 353, 337, 137 | Corosolic type triterpenoid | nq |
| 29 | 15.67 | 471.3453 | 471.3474 | C30H48O4 | 453, 411, 353, 337, 121, 113, 97, 71, 57 | Corosolic type triterpenoid | nq |
| 30 | 16.38 | 469.3342 | 469.3318 | C30H46O4 | 451, 425, 421, 407, 391, 377, 353, 337, 137 | Corosolic type triterpenoid | nq |
| 31 | 17.04 | 469.3342 | 469.3318 | C30H46O4 | 451, 425, 421, 407, 391, 377, 353, 337, 137 | Corosolic type triterpenoid | nq |
| 32 | 17.41 | 471.3453 | 471.3474 | C30H48O4 | 453, 411, 353, 337, 121, 113, 97, 71, 57 | Corosolic acid | 4.06 ± 2.46 |
| 33 | 20.18 | 453.3460 | 453.3369 | C30H46O3 | 405, 391, 389, 371, 337, 97 | Oleanolic type triterpenoid | nq |
| 34 | 20.84 | 455.3521 | 455.3525 | C30H48O3 | 452, 407, 391, 389, 375, 373, 189, 183, 137 | Betulinic acid | 3.93 ± 0.63 |
| 35 | 21.33 | 455.3539 | 455.3525 | C30H48O3 | 407, 391, 389, 375, 373, 189, 183, 137, 97 | Oleanolic acid | 7.26 ± 1.56 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faraone, I.; Russo, D.; Chiummiento, L.; Fernandez, E.; Choudhary, A.; Monné, M.; Milella, L.; Rai, D.K. Phytochemicals of Minthostachys diffusa Epling and Their Health-Promoting Bioactivities. Foods 2020, 9, 144. https://doi.org/10.3390/foods9020144
Faraone I, Russo D, Chiummiento L, Fernandez E, Choudhary A, Monné M, Milella L, Rai DK. Phytochemicals of Minthostachys diffusa Epling and Their Health-Promoting Bioactivities. Foods. 2020; 9(2):144. https://doi.org/10.3390/foods9020144
Chicago/Turabian StyleFaraone, Immacolata, Daniela Russo, Lucia Chiummiento, Eloy Fernandez, Alka Choudhary, Magnus Monné, Luigi Milella, and Dilip K. Rai. 2020. "Phytochemicals of Minthostachys diffusa Epling and Their Health-Promoting Bioactivities" Foods 9, no. 2: 144. https://doi.org/10.3390/foods9020144
APA StyleFaraone, I., Russo, D., Chiummiento, L., Fernandez, E., Choudhary, A., Monné, M., Milella, L., & Rai, D. K. (2020). Phytochemicals of Minthostachys diffusa Epling and Their Health-Promoting Bioactivities. Foods, 9(2), 144. https://doi.org/10.3390/foods9020144