Seed Protein of Lentils: Current Status, Progress, and Food Applications

 ,
, 
 ,
, 
Abstract
1. Introduction
2. Protein Content
3. Lentil Seed Storage Proteins
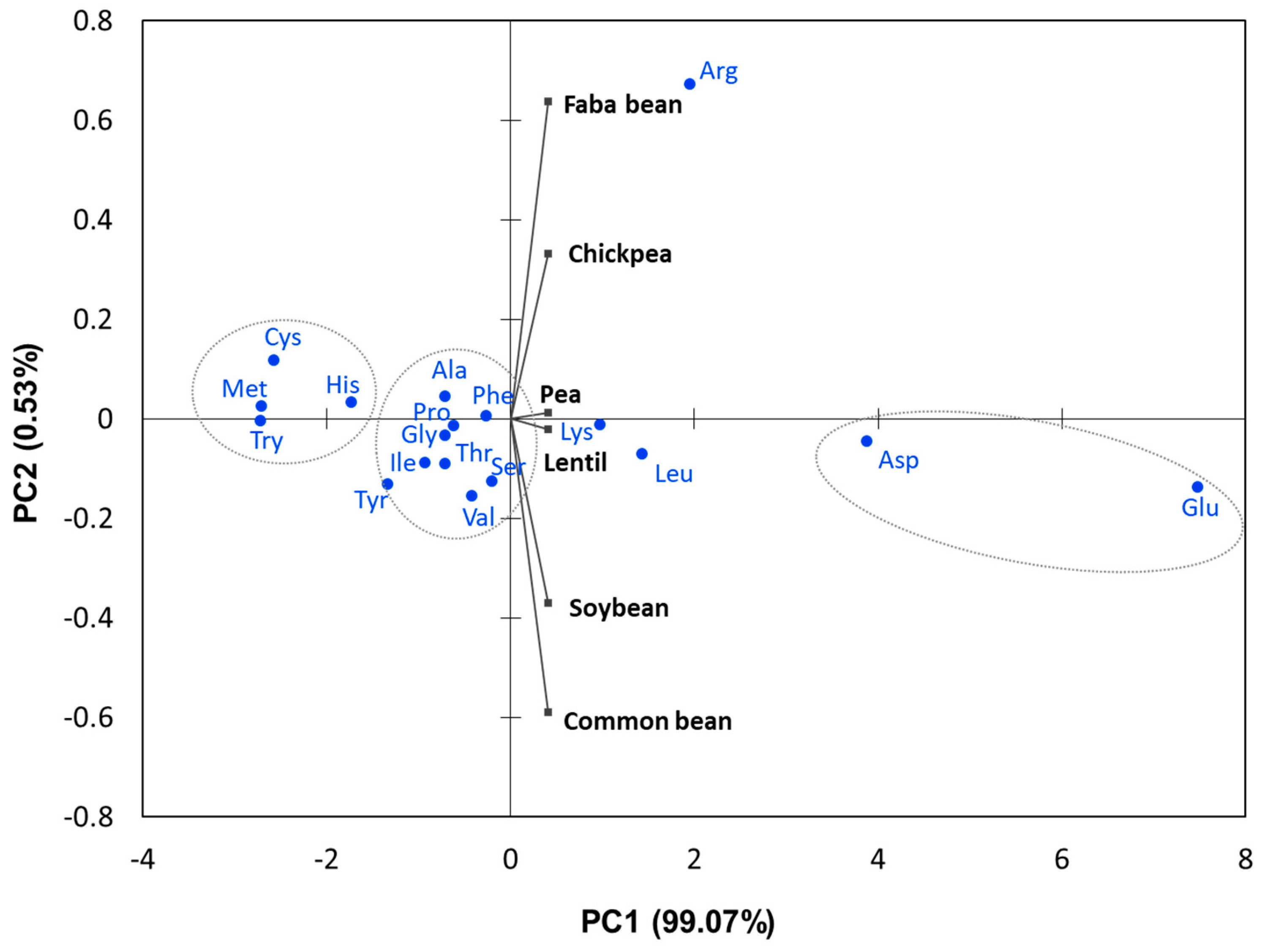
4. Amino Acids (AA)
5. Wild Lentil Taxa Protein Properties
6. Anti-Nutritional Factors Affecting Lentil Protein
7. Environmental Effects on Seed Proteins
8. Yield and Protein Relationships and Stability
9. Agronomic Protein Yield of Lentil
10. Seed Crude Protein Determination in Lentil
11. Protein Isolation Methods and Extraction
12. Bioactive Peptides
13. Food Applications of Lentil Proteins
13.1. Functionality
13.2. Challenges
13.3. Applications
14. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Krishnan, H.B.; Coe, E.B., Jr. Seed storage proteins. In Encyclopedia of Genetics; Science Direct: Amsterdam, The Netherlands, 2001; pp. 1782–1787. [Google Scholar]
- Nosworthy, M.G.; Neufeld, J.; Frohlich, P.; Young, G.; Malcolmson, L.; House, J.D. Determination of the protein quality of cooked Canadian pulses. Food Sci. Nutr. 2017, 5, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, A.; Slinkard, A.E. Genetics of seed coat color and pattern in lentil. J. Hered. 1990, 81, 484–488. [Google Scholar] [CrossRef]
- Vandenberg, A. Lentil: Improvement in developing countries. In The Lentil: Botany, Production and Uses; Erskine, W., Muehlbauer, F.J., Sarker, A., Sharma, B., Eds.; CAB International: Oxford, UK, 2009; pp. 391–424. [Google Scholar]
- Muehlbauer, F.J. Lentil: improvement in developing countries. In The Lentil: Botany, Production and Uses; Erskine, W., Muehlbauer, F.J., Sarker, A., Sharma, B., Eds.; CAB International: Oxford, UK, 2009; pp. 137–154. [Google Scholar]
- Khazaei, H.; Caron, C.T.; Podder, R.; Kundu, S.S.; Diapari, M.; Vandenberg, A.; Bett, K.E. Marker-trait association analysis of iron and zinc concentrations in lentils (Lens culinaris Medik.) seeds. Plant Genome. 2017, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Grela, E.R.; Kiczorowska, B.; Samolińska, W.; Matras, J.; Kiczorowski, P.; Rybiński, W.; Hanczakowska, E. Chemical composition of leguminous: Part I—Content of basic nutrients, amino acids, phytochemical compounds and antioxidant activity. Eur. Food Res. Technol. 2017, 243, 1385–1395. [Google Scholar] [CrossRef]
- Tahir, M.; Lindeboom, N.; Båga, M.; Vandenberg, A.; Chibbar, R.N. Composition and correlation between major seed constituents in selected lentil (Lens culinaris Medik). Can. J. Plant Sci. 2011, 91, 825–835. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTAT. Available online: http://faostat.fao.org (accessed on 2 September 2019).
- Semba, R.D. The rise and fall of protein malnutrition in global health. Ann. Nutr. Metab. 2016, 69, 79–88. [Google Scholar] [CrossRef]
- Arsenault, J.E.; Brown, K.H. Effects of protein or amino-acid supplementation on the physical growth of young children in low-income countries. Nutr. Rev. 2017, 75, 699–717. [Google Scholar] [CrossRef]
- Barulina, H. Lentils of the U.S.S.R. and of other countries: A botanic-agronomical monograph. In Bulletin of Applied Botany, of Genetics and Plant-Breeding; Leningrad Vsesoiuznaia akademiia sel’sko khoziaistvennyk im: Leningrad (St. Petersburg), Russia, 1930; (Suppl. S40), pp. 1–319. [Google Scholar]
- Hawtin, G.C.; Rachie, K.O.; Green, J.M. Breeding strategy for the nutritional improvement of pulses. In Nutritional Standards and Methods of Evaluation for Food Legume Breeders; Hulse, J.H., Rachie, K.O., Billingsley, L.W., Eds.; IDRC: Ottawa, ON, Canada, 1977; pp. 43–51. [Google Scholar]
- Hamdi, A.; Erskine, W.; Gates, P. Relationships among economic characters in lentil. Euphytica 1991, 57, 109–116. [Google Scholar] [CrossRef]
- Kumar, J.; Singh, J.; Kanaujia, R.; Gupta, S. Protein content in wild and cultivated taxa of lentil (Lens culinaris ssp. culinaris Medikus). Indian J. Genet. Plant Breed. 2016, 76, 631–634. [Google Scholar]
- Erskine, W.; Williams, P.C.; Nakkoul, H. Genetic and environmental variation in the seed size, protein, yield, and cooking quality of lentils. Field Crops Res. 1985, 12, 153–161. [Google Scholar] [CrossRef]
- Stoddard, F.L.; Marshall, D.R.; Ali, S.M. Variability in grain protein concentration of peas and lentils grown in Australia. Aust. J. Agric. Res. 1993, 44, 1415–1419. [Google Scholar] [CrossRef]
- Hedley, C.L. Carbohydrate in Grain Legume Seeds; CAB International: Wallingford, UK, 2001; pp. 1–11. [Google Scholar]
- Wang, N.; Daun, J.K. Effects of variety and crude protein content on nutrients and anti-nutrients in lentils (Lens culinaris). Food Chem. 2006, 95, 493–502. [Google Scholar] [CrossRef]
- Karaköy, T.; Erdem, H.; Baloch, F.S.; Toklu, F.; Eker, S.; Kilian, B.; Özkan, H. Diversity of macro-and micronutrients in the seeds of lentil landraces. Sci. World J. 2012, 2012, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zaccardelli, M.; Lupo, F.; Piergiovanni, A.R.; Laghetti, G.; Sonnante, G.; Daminati, M.G.; Sparvoli, F.; Lioi, L. Characterization of Italian lentil (Lens culinaris Medik.) germplasm by agronomic traits, biochemical and molecular markers. Genet. Resour. Crop Evol. 2012, 59, 727–738. [Google Scholar] [CrossRef]
- Alghamdi, S.S.; Khan, A.M.; Ammar, M.H.; El-Harty, E.H.; Migdadi, H.M.; Abd El-Khalik, S.M.; Al-Shameri, A.M.; Javed, M.M.; Al-Faifi, S.A. Phenological, nutritional and molecular diversity assessment among 35 introduced lentil (Lens culinaris Medik.) genotypes grown in Saudi Arabia. Int. J. Mol. Sci. 2014, 15, 277–295. [Google Scholar] [CrossRef]
- Heuzé, V.; Tran, G.; Sauvant, D.; Bastianelli, D.; Lebas, F. Lentil (Lens culinaris). Feedipedia, a programme by INRA, CIRAD, AFZ and FAO. 2015. Available online: https://www.feedipedia.org/node/284 (accessed on 2 September 2019).
- Shewry, P.R. Seed storage proteins: structures and biosynthesis. Plant Cell Online 1995, 7, 945–956. [Google Scholar]
- Sales, M.P.; Gerhardt, I.S.; Grossi-de-Sá, F.; Xavier-Filho, J. Do legume storage proteins play a role in Defending seeds against bruchids? Plant Physiol. 2000, 124, 512–522. [Google Scholar] [CrossRef]
- Cândido Ede, S.; Pinto, M.F.; Pelegrini, P.B.; Lima, T.B.; Silva, O.N.; Pogue, R.; Grossi-de-Sá, M.F.; Franco, O.L. Plant storage proteins with antimicrobial activity: Novel insights into plant defense mechanisms. FASEB J. 2011, 25, 3290–3305. [Google Scholar] [CrossRef]
- Osborne, T.B.; Campbell, G.F. Proteins of lentils. J. Am. Chem. Soc. 1898, 20, 362–375. [Google Scholar] [CrossRef][Green Version]
- Danielson, C.E. An electrophoretic investigation of vicilin and legumin from seeds of peas. Acta Chem. Scand. 1950, 4, 762–771. [Google Scholar] [CrossRef]
- Siddiqi, S.H. Chromatography on DEAE-cellulose and electrophoresis in polyacrylamide gel of protein fractions from lentil seeds. Pak. J. Sci. Ind. Res. 1982, 25, 167–169. [Google Scholar]
- Bhatty, R.S. Composition and quality of lentil (Lens culinaris): A review. Can. Inst. Food Sci. Technol. J. 1988, 21, 144–160. [Google Scholar] [CrossRef]
- Scippa, G.S.; Rocco, M.; Ialicicco, M.; Trupiano, D.; Viscosi, V.; Di Michele, M.; Arena, S.; Chiatante, D.; Scaloni, A. The proteome of lentil (Lens culinaris Medik.) seeds: Discriminating between landraces. Electrophoresis 2010, 31, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Neves, V.A.; Lourenco, E.J. Isolation and in vitro hydrolysis of globulin G1 from lentils (Lens culinaris Medik). J. Food Biochem. 1995, 19, 109–120. [Google Scholar] [CrossRef]
- Bourgeois, M.; Jacquin, F.; Savois, V.; Sommerer, N.; Labas, V.; Henry, C.; Burstin, J. Dissecting the proteome of pea mature seeds reveals the phenotypic plasticity of seed protein composition. Proteomics 2009, 9, 254–271. [Google Scholar] [CrossRef] [PubMed]
- Saint-Clair, P.M. Responses of Lens Esculenta Moench to Controlled Environmental Factors. Ph.D. Thesis, Committee of Agriculture, Wangeningen University, Wangeningen, The Netherlans, 1972; pp. 72–77. [Google Scholar]
- Bhatty, R.S.; Slinkard, A.E.; Sosulski, F.W. Chemical composition and protein characteristics of lentils. Can. J. Plant Sci. 1976, 56, 787–794. [Google Scholar] [CrossRef]
- Bhatty, R.S. In vitro hydrolysis of pea, faba bean and lentil meals and isolated protein fractions by pepsin and trypsin. Can. Inst. Food Sci. Technol. J. 1988, 21, 66–71. [Google Scholar] [CrossRef]
- Sulieman, M.A.; Amro, B.H.; Gamaa, A.O.; Mohamed, M.E.T.; Elhadi, A.I.E.K.; Abdullahi, H.E.T.; Elfadil, E.B. Changes in total protein digestibility, fractions content and structure during cooking of lentil cultivars. Pak. J. Nutr. 2008, 7, 801–805. [Google Scholar]
- Boye, J.I.; Aksay, A.; Roufik, S.; Ribereau, S.; Mondor, M.; Farnworth, E.; Rajamohamed, S.H. Comparison of the functional properties of pea, chickpea and lentil protein concentrates processed using ultrafiltration and isoelectric precipitation techniques. Food Res. Int. 2010, 43, 537–546. [Google Scholar] [CrossRef]
- Jarpa-Parra, M. Lentil protein: A review of functional properties and food application. An overview of lentil protein functionality. Int. J. Food Sci. Technol. 2018, 53, 892–903. [Google Scholar] [CrossRef]
- World Health Organization/Food and Agriculture—Organization of the United Nations/United Nations University. Protein and Amino Acid Requirements in Human Nutrition; Report of a Joint WHO/FAO/UNU Expert Consultation; WHO Technical Report Series 935; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Kahn, N.A.; Baker, B.E. The amino-acid composition of some Pakistani pulses. J. Sci. Food Agric. 1957, 8, 301–305. [Google Scholar] [CrossRef]
- Chatterjee, S.R.; Abrol, Y.P. Amino acid composition of new varieties of cereals and pulses and nutritional potential of cereal-pulse combinations. J. Food Sci. Technol. 1975, 12, 221–227. [Google Scholar]
- Sosulski, F.W. Legume protein concentration by air classification. In Developments in Food Proteins—2; Hudson, B.J.F., Ed.; Applied Science Publishers: Essex, UK, 1983. [Google Scholar]
- Bhatty, R.S.; Christison, G.I. Composition and nutritional quality of pea (Pisum sativum L.), faba bean (Vicia faba L. spp. minor) and lentil (lens culinaris Medik) meals, protein concentrates and isolates. Qual. Plant Plant Foods Hum. Nutr. 1984, 34, 41–51. [Google Scholar] [CrossRef]
- Shekib, L.A.H.; Zoueil, M.E.; Youssef, M.M.; Mohamed, M.S. Amino acid composition and in vitro digestibility of lentil and rice proteins and their mixture (Koshary). Food Chem. 1986, 20, 61–67. [Google Scholar] [CrossRef]
- Pirman, T.; Stibilj, V.; Stekar, J.M.A.; Combe, E. Amino acid composition of beans and lentil. Zb. Bioteh. Fak. Univ. Ljubl. Kmet. Zooteh. 2001, 78, 57–68. [Google Scholar]
- Porres, J.M.; Urbano, G.; Fernández-Fígares, I.; Prieto, C.; Pérez, L.; Aguilera, J.F. Digestive utilisation of protein and amino acids from raw and heated lentils by growing rats. J. Sci. Food Agric. 2002, 82, 1740–1747. [Google Scholar] [CrossRef]
- Zia-Ul-Haq, M.; Ahmad, S.; Shad, M.A.; Iqbal, S.; Qayum, M.; Ahmad, A.; Luthria, D.L.; Amarowicz, R. Compositional studies of lentil (Lens culinaris Medik.) cultivars commonly grown in Pakistan. Pak. J. Bot. 2011, 43, 1563–1567. [Google Scholar]
- Kaldy, N.S.; Kasting, R. Amino acid composition and protein quality of eight faba bean cultivars. Can. J. Plant Sci. 1974, 54, 869–871. [Google Scholar] [CrossRef]
- Lisiewska, Z.; Kmiecik, W.; Słupski, J. Content of amino acids in raw and frozen broad beans (Vicia faba var. major) seeds at milk maturity stage, depending on the processing method. Food Chem. 2007, 105, 1468–1473. [Google Scholar] [CrossRef]
- Schumacher, H.; Paulsen, H.M.; Gau, A.E.; Link, W.; Jürgens, H.U.; Sass, O.; Dieterich, R. Seed protein amino acid composition of important local grain Lupinus angustifolius L., Lupinus luteus L., Pisum sativum L. and Vicia faba L. Plant Breed. 2011, 130, 156–164. [Google Scholar] [CrossRef]
- Leterme, P.; Monmart, T.; Baudart, E. Amino acid composition of pea (Pisum sativum) proteins and protein profile of pea flour. J. Sci. Food Agric. 1990, 53, 107–110. [Google Scholar] [CrossRef]
- Pownall, T.L.; Udenigwe, C.C.; Aluko, R.E. Amino acid composition and antioxidant properties of pea seed (Pisum sativum L.) enzymatic protein hydrolysate fractions. J. Agric. Food Chem. 2010, 58, 4712–4718. [Google Scholar] [CrossRef] [PubMed]
- Kuiken, K.; Lyman, C.M.; Bradford, M.; Trant, M.; Dieterich, S. Essential amino acid composition of soy bean meals prepared from twenty strains of soy beans. J. Biol. Chem. 1949, 177, 29–36. [Google Scholar] [PubMed]
- Tkachuk, H.; Irvine, G.N. Amino acid composition of cereals and oilseed meals. Cereal Chem. 1969, 16, 206–218. [Google Scholar]
- Cho, C.Y.; Bayley, H.S. Evaluations of rapeseed and soybean meals as protein sources for swine: Apparent digestibilities of amino acids. Can. J. Anim. Sci. 1970, 50, 521–528. [Google Scholar] [CrossRef]
- Cavins, J.F.; Kwolek, W.E.; Inglett, G.E.; Cowan, J.C. Amino acid analysis of soybean meal: Interlaboratory study. J. Asso. Off. Anal. Chem. 1972, 55, 686–691. [Google Scholar]
- Wang, H.L.; Cavins, J.F. Yield and amino acid composition of fractions obtained during tofu production. Cereal Chem. 1989, 66, 359–361. [Google Scholar]
- Zarkadas, C.G.; Gagnon, C.; Gleddie, S.; Khanizadeh, S.; Cober, E.R.; Guillemette, R.J.D. Assessment of the protein quality of fourteen soybean (Glycine max (L.) Merr.) cultivars using amino acid analysis and two-dimensional electrophoresis. Food Res. Int. 2007, 40, 129–146. [Google Scholar] [CrossRef]
- Sotak-Peper, K.M.; González-Vega, J.C.; Stein, H.H. Amino acid digestibility in soybean meal sourced from different regions of the United States and fed to pigs. J. Anim. Sci. 2017, 95, 771–778. [Google Scholar] [CrossRef]
- Wang, N.; Daun, J.K. The Chemical Composition and Nutritive Value of Canadian Pulses; Canadian Grain Commission Report; Canadian Commision: Winnipeg, MB, Canada, 2004; pp. 19–29. [Google Scholar]
- Alajaji, S.A.; El-Adawy, T.A. Nutritional composition of chickpea (Cicer arietinum L.) as affected by microwave cooking and other traditional cooking methods. J. Food Comp. Anal. 2006, 19, 806–812. [Google Scholar] [CrossRef]
- Wang, X.; Gao, W.; Zhang, J.; Zhang, H.; Li, J.; He, X.; Ma, H. Subunit, amino acid composition and in vitro digestibility of protein isolates from Chinese kabuli and desi chickpea (Cicer arietinum L.) cultivars. Food Res. Int. 2010, 43, 567–572. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; El-Senousi, N.A.; Ali, Z.A.; Omran, A.A. The impact of using chickpea flour and dried carp fish powder on pizza quality. PLoS ONE 2017, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Williams, W.P.; Kunkel, M.E.; Acton, J.C.; Huang, Y.; Wardlaw, F.B.; Grimes, L.W. Amino acid availability and availability-corrected amino acid score of red kidney beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 1996, 44, 1296–1301. [Google Scholar] [CrossRef]
- Słupski, J. Effect of cooking and sterilisation on the composition of amino acids in immature seeds of flageolet bean (Phaseolus vulgaris L.) cultivars. Food Chem. 2010, 121, 1171–1176. [Google Scholar]
- Djemel, N.; Guedon, D.; Lechevalier, A.; Salon, C.; Miquel, M.; Prosperi, J.M.; Rochat, C.; Boutin, J.P. Development and composition of the seeds of nine genotypes of the Medicago truncatula species complex. Plant Physiol. Biochem. 2005, 43, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Van Vliet, S.; Burd, N.A.; van Loon, L.J. The skeletal muscle anabolic response to plant-versus animal-based protein consumption. J. Nutr. 2015, 145, 1981–1991. [Google Scholar] [CrossRef]
- Gorissen, S.H.M.; Crombag, J.J.R.; Senden, J.M.G.; Waterval, W.A.H.; Bierau, J.; Verdijk, L.B.; van Loon, L.J.C. Protein content and amino acid composition of commercially available plant-based protein isolates. Amino Acids 2018, 50, 1685–1695. [Google Scholar] [CrossRef]
- Rozan, P.; Kuo, Y.H.; Lambein, F. Amino acids in seeds and seedlings of the genus Lens. Phytochemistry 2001, 58, 281–289. [Google Scholar] [CrossRef]
- Bell, E.A.; Lyddiard, J.R.A.; Malik, M.A.; Motevalli, M.; Nunn, P.B.; O’Brien, P.; Perera, K.P.W.C. X-ray single-crystal structure of 2(S),4(R)-4-hydroxyarginine from Lens culinaris seeds. Phytochemistry 1999, 50, 1201–1204. [Google Scholar] [CrossRef]
- Sulser, H.; Sager, F. Identification of uncommon amino acids in the lentil seed (Lens culinaris Med.). Experientia 1976, 32, 422–423. [Google Scholar] [CrossRef]
- Wong, M.M.L.; Gujaria-Verma, N.; Ramsay, L.; Yuan, H.Y.; Caron, C.; Diapari, M.; Vandenberg, A.; Bett, K.E. Classification and characterization of species within the genus Lens using genotyping-by-sequencing (GBS). PLoS ONE 2015, 10, e0122025. [Google Scholar] [CrossRef] [PubMed]
- Bhatty, R.S. Protein subunits and amino acid composition of wild lentil. Phytochemistry 1986, 25, 641–644. [Google Scholar] [CrossRef]
- Gatel, F. Protein-quality of legume seeds for non-ruminant animals—A literature-review. Anim. Feed Sci. Technol. 1994, 45, 317–348. [Google Scholar] [CrossRef]
- Davis, K.R. Effect of processing on composition and Tetrahymena relative nutritive value on green and yellow peas, lentils and white pea beans. Cereal Chem. 1981, 58, 454–460. [Google Scholar]
- Matus, A.; Slinkard, A.E.; Vandenberg, A. The potential of zero tannin lentil. In New Crops; Janick, J., Simon, J.E., Eds.; Wiley: New York, NY, USA, 1993; pp. 279–282. [Google Scholar]
- Mirali, M.; Purves, R.W.; Stonehouse, R.; Song, R.; Bett, K.; Vandenberg, A. Genetics and biochemistry of zero-tannin lentils. PLoS ONE 2016, 11, e0164624. [Google Scholar] [CrossRef] [PubMed]
- Savage, G.P. Antinutritional factors in peas. In Recent Advances of Research in Antinutritional Factors in Legume Seeds; Huisman, J., van der Poel, T.F.B., Liener, I.E., Eds.; Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1989; pp. 342–350. [Google Scholar]
- Liener, I.E.; Kakade, M.L. Protease inhibitors. In Toxic Constituents in Plant Feedstuff; Liener, I.E., Ed.; Academic Press: New York, NY, USA, 1980; pp. 7–71. [Google Scholar]
- Pisulewska, E.; Pisulewski, P.M. Trypsin inhibitor activity of legume seed (peas, chickling vetch, lentils, and soya beans) as affected by the technique of harvest. Anim. Feed Sci. Technol. 2000, 86, 261–265. [Google Scholar] [CrossRef]
- Norton, G. Proteinase inhibitors. In Toxic Substances in Crop Plants; D’Mello, J.P.F., Duffus, C.M., Duffus, J.H., Eds.; The Royal Society of Chemistry: Cambridge, UK, 1991; pp. 68–106. [Google Scholar]
- Bhatty, R.S. Trypsin inhibitor activity in faba bean (Vicia faba var. minor), changes during germination and distribution. Can. J. Plant Sci. 1977, 57, 979–982. [Google Scholar] [CrossRef]
- Elkowicz, K.; Sosulski, F.W. Antinutritional factors in eleven legumes and their air-classified protein and starch fractions. Food Sci. 1982, 47, 1301–1304. [Google Scholar] [CrossRef]
- Shi, L.; Mu, K.; Arntfield, S.D.; Nickerson, M.T. Changes in levels of enzyme inhibitors during soaking and cooking for pulses available in Canada. J. Food Sci. Technol. 2017, 54, 1014–1022. [Google Scholar] [CrossRef]
- Zinselmeier, C.; Jeong, B.R.; Boyer, J.S. Starch and the control of kernel number in maize at low water potentials. Plant Physiol. 1999, 121, 25–35. [Google Scholar] [CrossRef]
- Gooding, M.J.; Ellis, R.H.; Shewry, P.R.; Schofield, J.D. Effects of restricted water availability and increased temperature on the grain filling, drying and quality of winter wheat. J. Cereal Sci. 2003, 37, 295–309. [Google Scholar] [CrossRef]
- Rakszegi, M.; Darkó, E.; Lovegrove, A.; Molnár, I.; Láng, L.; Bedő, Z.; Molnár-Láng, M.; Shewry, P. Drought stress affects the protein and dietary fiber content of wholemeal wheat flour in wheat/Aegilops addition lines. PLoS ONE 2019, 14, e0211892. [Google Scholar] [CrossRef] [PubMed]
- Behboudian, M.H.; Ma, Q.; Turner, N.C.; Palta, J.A. Reactions of chickpea to water stress: Yield and seed composition. J. Sci. Food Agric. 2001, 81, 1288–1291. [Google Scholar] [CrossRef]
- Dornbos, D.L.; Mullen, R.E. Soybean seed protein and oil contents and fatty acid composition adjustments by drought and temperature. J. Am. Oil Chem. Soc. 1992, 69, 228–231. [Google Scholar] [CrossRef]
- Mertz-Henning, L.M.; Ferreira, L.C.; Henning, F.A.; Mandarino, J.M.G.; Santos, E.D.; Oliveira, M.C.N.D.; Nepomuceno, A.E.L.; Farias, J.R.B.; Neumaier, N. Effect of water deficit-induced at vegetative and reproductive stages on protein and oil content in soybean grains. Agronomy 2018, 8, 3. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed]
- Sita, K.; Sehgal, A.; Bhandari, K.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.M.; Nayyar, H. Impact of heat stress during seed filling on seed quality and seed yield in lentil (Lens culinaris Medikus) genotypes. J. Sci. Food Agric. 2018, 98, 5134–5141. [Google Scholar] [CrossRef]
- Liu, T.; Staden, J. Partitioning of carbohydrates in salt-sensitive and salt-tolerant soybean cultures under salinity stress and its subsequent relief. Plant Growth Regul. 2001, 33, 13–17. [Google Scholar] [CrossRef]
- Zhao, H.; Dai, T.; Jiang, D.; Cao, W. Effects of high temperature on key enzymes involved in starch and protein formation in grains of two wheat cultivars. J. Agron. Crop Sci. 2008, 194, 47–54. [Google Scholar] [CrossRef]
- Mossé, J.; Huet, J.C.; Baudet, J. The amino acid composition of wheat grain as a function of nitrogen content. J. Cereal Sci. 1985, 3, 115–130. [Google Scholar] [CrossRef]
- Lizarazo, C.I.; Lampi, A.M.; Liu, J.; Sontag-Strohm, T.; Piironen, V.; Stoddard, F.L. Nutritive quality and protein production from grain legumes in a boreal climate. J. Sci. Food Agric. 2015, 95, 2053–2064. [Google Scholar] [CrossRef] [PubMed]
- Warsame, A.; O’Sullivan, D.M.; Tosi, P. Seed storage proteins of faba bean (Vicia faba L): Current status and prospects for genetic improvement. J. Agric. Food Chem. 2018, 66, 12617–12626. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, N.N. The causes of chemical variability in the seeds of chickpea in different geographical areas. In Monograph of the Genius; Quoted in Van der Maesen, 1972; Cicer, L.A., Ed.; Agriculture University Communication: Wageningen, The Netherlands, 1933. [Google Scholar]
- Khatun, A.M.; Bhuiyan, A.H.; Dey, T.K. Nitrogen uptake and protein yield in lentil as influenced by seed collection from different parts of plants. Bangladesh J. Agric. Res. 2010, 35, 515–523. [Google Scholar] [CrossRef]
- Ali-Khan, S.T. Seed yield, seed weight, percent protein and protein yield of field peas as affected by seeding dates. Can. J. Plant Sci. 1977, 57, 17–20. [Google Scholar] [CrossRef]
- Karjalainen, R.; Hovinen, S. Variation in protein content of peas under Finnish conditions. J. Sci. Agric. Soc. Finl. 1981, 53, 228–238. [Google Scholar] [CrossRef]
- Krotz, L.; Ciceri, E.; Giazzi, G. Protein Determination in Cereals and Seeds. Food Quality and Safety. 2008. Available online: http/Foodqualityandsafety.com (accessed on 2 September 2019).
- Moore, J.C.; de Vries, W.; Lipp, M.; Grifiths, J.C.; Abernethy, D.R. Total protein methods and their potential utility to reduce the risk of food protein adulteration. Compr. Rev. Food Sci. Food Saf. 2010, 9, 330–351. [Google Scholar] [CrossRef]
- FAO. Pulses. 2016. Available online: www.fao.com (accessed on 2 September 2019).
- Mihaljev, A.Ž.; Jakšić, M.S.; Prica, N.B.; Ćupić, Ž.N.; Živkov-Baloš, M. Comparison of the Kjeldahl method, Dumas method and NIR method for total nitrogen determination in meat and meat products. J. Agro Proc. Technol. 2015, 21, 365–370. [Google Scholar]
- Subedi, M.; Willenborg, C.J.; Vandenberg, A. Influence of harvest aid herbicides on seed germination, seedling vigor and milling quality traits of red lentil (Lens culinaris L.). Front. Plant Sci. 2017, 8, 311. [Google Scholar] [CrossRef] [PubMed]
- Swanson, B.G. Pea and lentil protein extraction and functionality. J. Am. Oil Chem. Soc. 1990, 67, 276–279. [Google Scholar] [CrossRef]
- Aryee, A.N.A.; Boye, J.I. Improving the digestibility of lentil flours and protein isolate and characterization of their enzymatically prepared hydrolysates. Int. J. Food Prop. 2016, 19, 2649–2665. [Google Scholar] [CrossRef]
- Joshi, M.; Timilsena, Y.; Adhikari, B. Global production, processing and utilization. J. Integr. Agric. 2017, 16, 2898–2913. [Google Scholar] [CrossRef]
- Vose, J.R. Production and functionality of starches and protein isolates form legume seeds (field peas and horsebeans). Cereal Chem. 1980, 57, 406–410. [Google Scholar]
- Gueguen, J. Legume seed protein extraction, processing, and end product characteristics. Plant Foods Hum. Nutr. 1993, 32, 267–303. [Google Scholar] [CrossRef]
- Tyler, R.T.; Youngs, C.G.; Sosulski, F.W. Air classification of legumes. 1. Separation efficiency, yield, and composition of the starch and protein fractions. Cereal Chem. 1981, 58, 144–148. [Google Scholar]
- Reichert, R.D. Quantitative isolation and estimation of cell wall material from dehulled pea (Pisum sativum) flours and concentrates. Cereal Chem. 1981, 58, 266–270. [Google Scholar]
- Lee, H.C.; Htoon, A.K.; Uthayakumaran, S.; Paterson, J.L. Chemical and functional quality of protein isolated from alkaline extraction of Australian lentil cultivars: Matilda and Digger. Food Chem. 2007, 102, 1199–1207. [Google Scholar] [CrossRef]
- Fan, T.Y.; Sosulski, F.W. Dispersibility and isolation of proteins from legume flours. Can. Inst. Food Sci. Technol. J. 1974, 7, 256–259. [Google Scholar] [CrossRef]
- Alsohaimy, S.A.; Sitohy, M.Z.; El-Masry, R.A. Isolation and partial characterization of chickpea, lupine and lentil seed proteins. World J. Agric. Sci. 2007, 3, 123–129. [Google Scholar]
- Joshi, M.; Adhikari, B.; Aldred, P.; Panozzo, J.F.; Kasapis, S. Physicochemical and functional properties of lentil protein isolates prepared by different drying methods. Food Chem. 2011, 129, 1513–1522. [Google Scholar] [CrossRef]
- Joshi, M.; Adhikari, B.; Aldred, P.; Panozzo, J.F.; Kasapis, S.; Barrow, C.J. Interfacial and emulsifying properties of lentil protein isolate. Food Chem. 2012, 134, 1343–1353. [Google Scholar] [CrossRef]
- Kaur, M.; Singh, N.; Sandhu, K.S. Preparation and characterization of protein isolates from different lentil (Lens culinaris) cultivars. J. Food Sci. Technol. 2007, 44, 327–329. [Google Scholar]
- Johnston, S.P.; Nickerson, M.T.; Nicholas, H.L. The physicochemical properties of legume protein isolates and their ability to stabilize oil-in-water emulsions with and without genipin. J. Food Sci. Technol. 2015, 52, 4135–4145. [Google Scholar] [CrossRef] [PubMed]
- Jarpa-Parra, M.; Bamdad, F.; Wang, Y.; Tian, Z.; Temelli, F.; Han, J.; Chen, L. Optimization of lentil protein extraction and the influence of process pH on protein structure and functionality. LWT Food Sci. Technol. 2014, 57, 461–469. [Google Scholar] [CrossRef]
- Matsusaka, S.; Maruyama, H.; Matsuyama, T.; Ghadiri, M. Triboelectric charging of powders: A review. Chem. Eng. Sci. 2010, 65, 5781–5807. [Google Scholar] [CrossRef]
- Jafari, M.; Rajabzadeh, A.R.; Tabtabaei, S.; Marsolais, F.; Legge, R.L. Physicochemical characterization of a navy bean (Phaseolus vulgaris) protein fraction produced using a solvent-free method. Food Chem. 2016, 208, 35–41. [Google Scholar] [CrossRef]
- Assatory, A.; Vitellia, M.; Rajabzdeh, A.R.; Legge, R.L. Dry fractionation methods for plant protein, starch and fiber enrichment: A review. Trends Food Sci. Technol. 2019, 86, 340–351. [Google Scholar] [CrossRef]
- Tabtabaei, S.; Jafari, M.; Rajabzadeh, A.R.; Legge, R.L. Solvent-free production of protein-enriched fractions from navy bean flour using a triboelectrification-based approach. J. Food Eng. 2016, 174, 21–28. [Google Scholar] [CrossRef]
- Tabtabaei, S.; Konakbayeva, D.; Rajabzadeh, A.R.; Legge, R.L. Analysis of protein enrichment during single- and multi-stage triboelectrostatic bioseparation processes for dry fractionation of legume flour. Sep. Purif. Technol. 2017, 176, 48–58. [Google Scholar] [CrossRef]
- Tabtabaei, S.; Konakbayeva, D.; Rajabzadeh, A.R.; Legge, R.L. Functional properties of navy bean (Phaseolus vulgaris) protein concentrates obtained by pneumatic tribo-electrostatic separation. Food Chem. 2019, 283, 101–110. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Food protein-derived bioactive peptides: Production, processing, and potential health benefits. J. Food Sci. 2012, 77, R11–R24. [Google Scholar] [CrossRef]
- Garcia-Mora, P.; Peñas, P.; Frias, J.; Martinez-Villaluenga, C. Savinase, the most suitable enzyme for releasing peptides from lentil (Lens culinaris var. castellana) protein concentrates with multifunctional properties. J. Agric. Food Chem. 2014, 62, 4166–4174. [Google Scholar] [CrossRef]
- Barbana, C.; Boye, J.I. Angiotensin I-converting enzyme inhibitory properties of lentil protein hydrolysates: Determination of the kinetics of inhibition. Food Chem. 2011, 127, 94–101. [Google Scholar] [CrossRef]
- Natesh, R.; Schwager, S.L.; Sturrock, E.D.; Acharya, K.R. Crystal structure of the human angiotensin-converting enzyme-Lysinopril complex. Nature 2003, 421, 551–554. [Google Scholar] [CrossRef] [PubMed]
- García-Mora, P.; Martín-Martínez, M.; Bonache, M.A.; González-Múniz, R.; Peñas, E.; Frias, J.; Martinez-Villaluenga, C. Identification, functional gastrointestinal stability and molecular docking studies of lentil peptides with dual antioxidant and angiotensin I converting enzyme inhibitory activities. Food Chem. 2017, 221, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural requirements of angiotensin I converting enzyme inhibitory peptides: Quantitative structure-activity relationship study of di and tripeptides. J. Agric. Food Chem. 2006, 54, 732–738. [Google Scholar] [CrossRef]
- Rizzello, C.B.; Verni, M.; Bordignon, S.; Gramaglia, V.; Gobbetti, M. Hydrolysate from a mixture of legume flours with antifungal activity as an ingredient for prolonging the shelf-life of wheat bread. Food Microbiol. 2017, 64, 72–82. [Google Scholar] [CrossRef]
- Wang, H.X.; Ng, T.B. An antifungal peptide from red lentil seeds. Peptides 2007, 28, 547–552. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Ulug, S.K.; Hong, H.; Wu, J. Regulatory requirements of bioactive peptides (protein hydrolysates) from food proteins. J. Funct. Foods 2019, 58, 123–129. [Google Scholar] [CrossRef]
- Mamilla, R.K.; Mishra, V.K. Effect of germination on antioxidant and ACE inhibitory activities of legumes. LWT Food Sci. Technol. 2017, 75, 51–58. [Google Scholar] [CrossRef]
- Boschin, G.; Scigliuolo, G.M.; Resta, D.; Arnoldi, A. ACE-inhibitory activity of enzymatic protein hydrolysates from lupin and other legumes. Food Chem. 2014, 145, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Jakubczyk, A.; Baraniak, B. Activities and sequences of the angiotensin I-converting enzyme (ACE) inhibitory peptides obtained from the digested lentil (Lens culinaris) globulins. Int. J. Food Sci. Technol. 2013, 48, 2363–2369. [Google Scholar] [CrossRef]
- Akıllıoğlu, H.G.; Karakaya, S. Effects of heat treatment and in vitro digestion on the Angiotensin converting enzyme inhibitory activity of some legume species. Eur. Food Res. Technol. 2009, 229, 915–921. [Google Scholar] [CrossRef]
- Pelgrom, P.J.M.; Boom, R.M.; Schutyser, M.A.I. Method development to increase protein enrichment during dry fractionation of starch-rich legumes. Food Bioprocess Technol. 2015, 8, 1495–1502. [Google Scholar] [CrossRef]
- Ladjal-Ettoumi, Y.; Boudries, H.; Chibane, M.; Romero, A. Pea, chickpea and lentil protein isolates: Physicochemical characterization and emulsifying properties. Food Biophys. 2016, 11, 43–51. [Google Scholar] [CrossRef]
- Can Karaca, A.; Low, N.; Nickerson, M. Emulsifying properties of chickpea, faba bean, lentil and pea proteins produced by isoelectric precipitation and salt extraction. Food Res. Int. 2011, 444, 2742–2750. [Google Scholar] [CrossRef]
- Boye, J.I.; Roufik, S.; Pesta, N.; Barbana, C. Angiotensin I-converting enzyme inhibitory properties and SDS-PAGE of red lentil protein hydrolysates. Food Sci. Technol. 2010, 43, 987–991. [Google Scholar] [CrossRef]
- Chang, C.; Tu, S.; Ghosh, S.; Nickerson, M.T. Effects of pH on the inter-relationships between the physicochemical, interfacial and emulsifying properties for pea, soy, lentil and canola protein isolates. Food Res. Int. 2015, 77, 360–367. [Google Scholar] [CrossRef]
- Primozic, M.; Duchek, A.; Nickerson, M.T.; Ghosh, S. Formation, stability and in vitro digestibility of nanoemulsions stabilized by high-pressure homogenized lentil protein isolates. Food Hydrocoll. 2018, 77, 126–141. [Google Scholar] [CrossRef]
- Gumus, C.E.; Decker, E.A.; Mc Clements, D.J. Impact of legume protein type and location on lipid oxidation in fish oil-in-water emulsions: Lentil, peas and faba bean proteins. Food Res. Int. 2017, 100, 175–185. [Google Scholar] [CrossRef]
- Aryee, A.N.A.; Boye, J.I. Comparative study of the effects of processing on the nutritional, physicochemical and functional properties of lentil. J. Food Proc. Preserv. 2017, 41, e12824. [Google Scholar] [CrossRef]
- Toews, R.; Wang, N. Physicochemical and functional properties of protein concentrates from pulses. Food Res. Int. 2013, 52, 445–451. [Google Scholar] [CrossRef]
- Stantiall, S.E.; Dale, K.J.; Calizo, F.S.; Serventi, L. Applications of pulses cooking water as functional ingredients: The foaming and gelling abilities. Eur. Food Res. Technol. 2018, 244, 97–104. [Google Scholar] [CrossRef]
- Chang, C.; Stone, A.K.; Green, R.; Nickerson, M.T. Reduction of off-flavours and the impact on the functionalities of lentil protein isolate by acetone, ethanol, and isopropanol treatments. Food Chem. 2019, 277, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Shariati-Ievari, S.; Ryland, D.; Edel, A.; Nicholson, T.; Suh, M.; Aliani, M. Sensory and physicochemical studies of thermally micronized chickpea (Cicer arietinum) and green lentil (Lens culinaris) flours as binders in low-fat beef burgers. J. Food Sci. 2016, 81, S1230–S1242. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Boye, J.I.; Azarnia, S.; Simpson, B.K. Volatile flavor profile of Saskatchewan grown pulses as affected by different thermal processing treatments. Int. J. Food Prop. 2016, 19, 2251–2271. [Google Scholar] [CrossRef]
- Eckert, E.; Wismer, W.; Waduthanthiri, K.; Babii, O.; Yang, J.; Chen, L. Application of barley-and lentil-protein concentrates in the production of protein-enriched doughnuts. J. Am. Oil Chem. Soc. 2018, 95, 1027–1040. [Google Scholar] [CrossRef]
- Jarpa-Parra, M.; Wong, L.; Wismer, W.; Temelli, F.; Han, J.; Huang, W.; Eckhart, E.; Tian, Z.; Shi, K.; Sun, T.; et al. Quality characteristics of angle3 food cake and muffin using lentil protein as egg/milk replacer. Int. J. Food Sci. Technol. 2017, 52, 1604–1613. [Google Scholar] [CrossRef]
- Han, J.; Janz, J.A.M.; Gerlat, M. Development of gluten-free cracker snacks using pulse flours and fractions. Food Res. Int. 2010, 43, 627–633. [Google Scholar] [CrossRef]
- Baugreet, S.; Kery, J.P.; Botinestean, C.; Allen, P.; Hamill, R.M. Development of novel fortified beef patties with added functional protein ingredients for elderly. Meat Sci. 2016, 122, 40–47. [Google Scholar] [CrossRef]
- Baugreet, S.; Kerry, J.P.; Brodkorb, A.; Gomex, C.; Auty, M.; Allen, P.; Hamill, R.M. Optimisation of plant protein and transglutaminase content in novel beef restructured steaks for older adults by central composite design. Meat Sci. 2018, 142, 65–77. [Google Scholar] [CrossRef]
- Ma, Z.; Boye, J.I.; Simpson, B.K. Preparation of salad dressing emulsions using lentil, chickpea and pea protein isolates: A response surface methodology study. J. Food Qual. 2016, 39, 274–291. [Google Scholar] [CrossRef]
- Primozic, M.; Duchek, A.; Nickerson, M.T.; Ghosh, S. Effect of lentil protein isolate concentration on the formation, stability and rheological behavior of oil-in-water nanoemulsions. Food Chem. 2017, 234, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Nickerson, M.T. Stability and in vitro release behaviour of encapsulated omega fatty acid-rich oils in lentil protein isolate-based microcapsules. Int. J. Food Sci. Nutr. 2018, 69, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Varankovich, N.; Nickerson, M.T. Microencapsulation of canola oil by lentil protein isolate-based wall materials. Food Chem. 2016, 212, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Boyaci, D.; Yemenicioglu, A. Expanding horizons of active packaging: Design of consumer-controlled release systems helps risk management of susceptible individuals. Food Hydrocoll. 2018, 79, 291–300. [Google Scholar] [CrossRef]
- Tam, N.; Oguz, S.; Aydogdu, A.; Sumnu, G.; Sahin, S. Influence of solution properties and pH on the fabrication of electro spun lentil flour/HPMC blend nanofibers. Food Res. Int. 2017, 102, 616–624. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Crop | Global Production | Global Growth Trends | ||||
|---|---|---|---|---|---|---|
| 1960–1965 | 2012–2017 | |||||
| Annual Mean (Mt) | % of Total | Annual Mean (Mt) | % of Total | 50 Years % Growth | Annual % Growth | |
| Lentil | 1.0 | 2.8 | 5.9 | 9.2 | 515 | 10.3 |
| Faba bean | 5.1 | 14.7 | 4.5 | 7.0 | −12 | −0.2 |
| Pea | 10.5 | 30.0 | 13.1 | 20.5 | 25 | 0.5 |
| Chickpea | 6.3 | 17.9 | 12.7 | 19.8 | 104 | 2.1 |
| Common bean | 12.1 | 34.5 | 27.9 | 43.5 | 132 | 2.6 |
| Total pulses 1,2 | 34.9 | 100 | 64.2 | 90 | 84 | 1.7 |
| Soybean (total) | 28.6 | 319.0 | 1115 | 22.3 | ||
| Soybean eaten | 10.0 | 67.0 | 670 | 13.4 | ||
| directly 3 | ||||||
| People (1967) 4 | 3465 million | 7600 million | 119 | 2.4 | ||
| Protein Content (Range, % DM 1) | Number of Accessions | Environment | Method | Reference |
|---|---|---|---|---|
| 27.5–31.7 | - | Russia | A micro Kjeldahl Method | Barulina [12] |
| 23.4–36.4 | 1688 | Egypt | A micro Kjeldahl Method | Hawtin et al. [13] |
| 25.5–28.9 | 24 | Lebanon and Syria | NIR 2 using a Neotec model FQA51 A analyzer | Erskine et al. [16] |
| 19.6–29.8 and 18.6–30.2 | 829 and 987 | Tel Hadya (Syria) | NIR | Hamdi et al. [14] |
| 23.9–26.3 | 58 | Australia | A Kjeldahl N × 6.25 on an oven-dry basis method | Stoddard et al. [17] |
| 23.0–32.0 | - | - | - | Hedley [18] |
| 24.3–30.2 | 4 | - | NIR using a NIR Systems 6500 analyzer calibrated against the Dumas method | Wang and Daun [19] |
| 23.8–29.3 | 22 | Saskatoon, Canada | A Dumus Combustion method to determine Nitrogen percentage using a method in 46-30.01 | Tahir et al. [8] |
| 22.7–31.88 | 46 | Turkey | A Kjeldahl method AOAC, Official Method of Analysis | Karaköy et al. [20] |
| 21.8–27.1 | 14 | Italy | A Kjeldahl method | Zaccardelli et al. [21] |
| 25.3–29.3 | 35 | Saudi Arabia | A Kjeldahl method of Association of Official Analytical Chemists (AOAC) | Alghamdi et al. [22] |
| 24.6–30.0 | 23 | Multiple | Multiple methods | Heuzé et al. [23] |
| 10.5–27.1 | 45 | India | A Kjeldahl method | Kumar et al. [15] |
| Salt Soluble | Water Soluble | Acid Soluble | Ethanol Soluble | Reference |
|---|---|---|---|---|
| Globulins | Albumins | Glutelins | Prolamins | |
| 44% | 26% | 20% | 2% | Saint-Clair [34] |
| 47% | 4% | 15% | 3% | Bhatty et al. [35] 1 |
| 54% | 20% | - | - | Bhatty [36] |
| 42% | 11% | 47% 2 | - | Neves and Lourenco [32] |
| 27% | 61% | 3% | 2% | Sulieman et al. [37] |
| 70% | 16% | 11% | 3% | Boye et al. [38] |
| Amino Acid | Lentil 1 | Faba Bean 2 | Pea 3 | Soybean 4 | Chickpea 5 | Common Bean 6 | SEM 7 |
|---|---|---|---|---|---|---|---|
| Alanine | 4.2 ± 0.4 | 4.1 ± 0.2 | 4.3 ± 0.2 | 4.2 ± 0.3 | 4.1 ± 0.5 | 3.8 ± 0.3 | 0.07 |
| Arginine | 7.8 ± 1.0 | 10.2 ± 1.1 | 8.2 ± 0.7 | 7.4 ± 0.6 | 9.0 ± 1.2 | 6.5 ± 0.7 | 0.51 |
| Aspartic acid | 10.7 ± 1.1 | 11.0 ± 1.6 | 11.3 ± 0.5 | 11.3 ± 0.7 | 11.6 ± 0.7 | 10.6 ± 1.3 | 0.15 |
| Cysteine | 1.1 ± 0.3 | 1.4 ± 0.3 | 1.3 ± 0.3 | 1.3 ± 0.4 | 1.3 ± 0.1 | 1.1 ± 0.2 | 0.06 |
| Glutamic acid | 16.1 ± 2.6 | 16.7 ± 2.2 | 16.4 ± 0.7 | 17.8 ± 1.2 | 16.8 ± 2.2 | 15.6 ± 2.1 | 0.31 |
| Glycine | 4.1 ± 0.7 | 4.3 ± 0.2 | 4.3 ± 0.2 | 4.1 ± 0.4 | 3.6 ± 0.6 | 4.2 ± 0.5 | 0.10 |
| Histidine | 2.4 ± 0.5 | 2.6 ± 0.2 | 2.3 ± 0.3 | 2.6 ± 0.1 | 2.6 ± 0.5 | 2.7 ± 0.2 | 0.05 |
| Isoleucine | 4.1 ± 0.5 | 4.0 ± 0.4 | 4.1 ± 0.5 | 4.4 ± 0.6 | 3.8 ± 0.4 | 4.2 ± 0.3 | 0.09 |
| Leucine | 7.2 ± 0.4 | 7.7 ± 0.6 | 7.3 ± 0.8 | 7.5 ± 0.4 | 7.0 ± 0.4 | 7.5 ± 0.7 | 0.11 |
| Lysine | 6.7 ± 0.6 | 6.4 ± 0.1 | 7.6 ± 1.2 | 6.4 ± 0.6 | 6.5 ± 0.8 | 6.3 ± 0.5 | 0.19 |
| Methionine | 0.9 ± 0.2 | 0.7 ± 0.1 | 1.0 ± 0.1 | 1.3 ± 0.3 | 1.4 ± 0.3 | 1.0 ± 0.4 | 0.11 |
| Phenylalanine | 5.0 ± 0.6 | 4.2 ± 0.2 | 4.8 ± 0.5 | 4.8 ± 0.3 | 5.5 ± 0.5 | 4.4 ± 0.7 | 0.18 |
| Proline | 3.8 ± 0.4 | 4.1 ± 0.5 | 4.4 ± 0.9 | 5.1 ± 0.3 | 4.4 ± 0.4 | 3.8 ± 0.4 | 0.19 |
| Serine | 4.7 ± 0.7 | 4.6 ± 0.4 | 4.9 ± 0.5 | 5.1 ± 0.5 | 4.8 ± 0.9 | 5.2 ± 0.7 | 0.08 |
| Threonine | 3.7 ± 0.4 | 3.5 ± 0.2 | 3.8 ± 0.3 | 3.9 ± 0.4 | 3.7 ± 0.6 | 4.0 ± 0.2 | 0.07 |
| Tryptophan | 0.8 ± 0.1 | 0.9 ± 0.1 | 1.2 ± 0.6 | 1.4 ± 0.3 | 1.0 ± 0.1 | 1.1 ± 0.4 | 0.09 |
| Tyrosine | 2.5 ± 0.7 | 3.1 ± 0.3 | 3.3 ± 0.5 | 3.4 ± 0.6 | 2.9 ± 0.5 | 3.7 ± 0.6 | 0.18 |
| Valine | 4.7 ± 0.4 | 4.4 ± 0.4 | 4.5 ± 0.5 | 4.7 ± 0.5 | 4.0 ± 0.4 | 4.9 ± 0.5 | 0.13 |
| Conditions for Lentil Protein Extraction | % Protein Yield | % Protein in Final Extracts | Reference |
|---|---|---|---|
| pH: 6, 7, 8, 9, 10, 11, and 12 Temperature: room Solid to solvent ratio: 5:100 | 80.0 | 21.5 | Alsohaimy et al. [117] |
| pH: 8, 8.5, 9, 9.5 Temperature: 22 °C, 30 °C, 35 °C, and 40 °C Solid to solvent ratio: 1:10 | 56.6–59.3 | - | Lee et al. [115] |
| pH: 9 Temperature: 25 °C Solid to solvent ratio: 1:10 | 50.3–69.1 | - | Boye et al. [38] |
| pH: 8 Temperature: room Solid to solvent ratio: 1:10 | - | - | Joshi et al. [119] |
| pH: 4.6 Temperature: room Solid to solvent ratio: 1:10 | 82.0 | 14.5 | Johnston et al. [121] |
| pH: 8, 9 and 10 Temperature: room Solid to solvent ratio: 1:10 | 70.3–85.7 | 12.3–16.5 | Jarpa-Parra et al. [122] |
| Biological Activity | Raw Material | Processing Conditions | Peptide Sequence | Effect Observed | Reference |
|---|---|---|---|---|---|
| Antioxidant and antihypertensive | Protein concentrate | Enzymatic hydrolysis with Savinase 16 L (0.1 U/mg protein, pH 8, 40 °C, 2 h) | LLSGTQNQPSFLSGF 1 | ACE 2 inhibition: IC50 3 = 120 µM ORAC 4: 0.013 µmol Trolox eq./µmol | Garcia-Mora et al. [133] |
| NSLTLPILRYL | ACE inhibition: IC50 = 77.14 µM ORAC: 1.432 µmol Trolox eq./µmol | ||||
| TLEPNSVFLPVLLH | ACE inhibition: IC50 = 117.81 µM ORAC: 0.139 µmol Trolox eq./µmol | ||||
| Antihypertensive | Sprouts | Germination (30–40 °C for 5 days, 98% humidity) | Unknown | ACE inhibition: IC50 = 0.044 and 0.034 mg/mL | Mamilla and Mishra [138] |
| Antifungal | Flour | Fermentation with Lactobacillus plantarum (7 log cfu/g) and enzymatic hydrolysis with Veron PS (E/S 5 of 1/400, 30 °C, 24 h) | HIGIDVNSIK | Inhibition of germination of Penicillium roqueforti DPPMAF1 conidia: MIC 6 = 7–9 mg/mL | Rizzello et al. [135] |
| NLIFQGDGYTTK | |||||
| FSPDQQNLIFQGDGYTTK | |||||
| HIGIDVNSIK | |||||
| Antihypertensive | Protein isolate | Enzymatic hydrolysis with pepsin (E/S of 1/100, pH 2, 37 °C for 18 h) | Unknown | ACE inhibition: IC50 = 606 µg/mL | Boschin et al. [139] |
| Antihypertensive | Protein isolate | Pepsin (250 U/mg, pH 2, 37 °C, 2 h) and pancreatin (0.7%, pH 7, 37 °C, 1 h) | KLRT | ACE inhibition: IC50 = 0.13–0.02 mg/mL for different peptide fractions | Jakubczyk and Baraniak [140] |
| TLHGMV | |||||
| VNRLM | |||||
| Antihypertensive | Red protein concentrates | Pepsin (E/S of 1/250, for 2 h, pH 2, 37 °C) + Trypsin and α-chymotrypsin (E/S of 1/250 for each enzyme, 2.5 h, pH 6.5, 37 °C) | Unknown | ACE inhibition: IC50 = 0.090 mg/mL | Barbana and Boye [131] |
| Papain (E/S of 1/25, pH 6.5, 4 h, 40 °C) | Unknown | IC50 = 0.086 mg/mL | |||
| Alcalase (1/8 for E/S ratio, pH 7, 1 h, 50 °C) + Flavourzyme (E/S of 1/10, pH 8, 1.5 h at 50 °C) | Unknown | IC50 = 0.154 mg/mL | |||
| Bromelain (E/S of 1/4, pH 8, 8 h, 40 °C) | Unknown | IC50 = 0.190 mg/mL | |||
| Green protein concentrates | Pepsin (pepsin (E/S of 1/250, for 2 h, pH 2, 37 °C) + Trypsin and α-chymotrypsin (E/S of 1/250 for each enzyme, 2.5 h, pH 6.5, 37 °C) | Unknown | ACE inhibition: IC50 = 0.053 mg/mL | ||
| Papain (E/S of 1/25, pH 6.5, 4 h, 40 °C) | Unknown | IC50 = 0.080 mg/mL | |||
| Alcalase (1/8 for E/S ratio, pH 7, 1 h, 50 °C) + Flavourzyme (E/S of 1/10, pH 8, 1.5 h at 50 °C) | Unknown | IC50 = 0.152 mg/mL | |||
| Bromelain (E/S of 1/4, pH 8, 8 h, 40 °C) | Unknown | IC50 = 0.174 mg/mL | |||
| Antihypertensive | Red protein isolates | Pepsin (E/S of 4/10, pH 2, 37 °C, 2 h) and pancreatin (E/S of 0.5/10, pH 7, 37 °C, 2 h) | Unknown | ACE inhibition: IC50 = 0.008–0.33 mg/mL in gastric phase | Akıllıoğlu and Karakaya [141] |
| IC50 = 0.26–0.89 mg/mL in intestinal phase | |||||
| Antifungal | Red lentil extract | Chromatographic fractionation | TETNSFSITKFSPDGNKLIFQGDGYTTKGK | Inhibition of mycelial growth in Mycosphaerella arachidicola: IC50 = 36 μM | Wang and Ng [136] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khazaei, H.; Subedi, M.; Nickerson, M.; Martínez-Villaluenga, C.; Frias, J.; Vandenberg, A. Seed Protein of Lentils: Current Status, Progress, and Food Applications. Foods 2019, 8, 391. https://doi.org/10.3390/foods8090391
Khazaei H, Subedi M, Nickerson M, Martínez-Villaluenga C, Frias J, Vandenberg A. Seed Protein of Lentils: Current Status, Progress, and Food Applications. Foods. 2019; 8(9):391. https://doi.org/10.3390/foods8090391
Chicago/Turabian StyleKhazaei, Hamid, Maya Subedi, Mike Nickerson, Cristina Martínez-Villaluenga, Juana Frias, and Albert Vandenberg. 2019. "Seed Protein of Lentils: Current Status, Progress, and Food Applications" Foods 8, no. 9: 391. https://doi.org/10.3390/foods8090391
APA StyleKhazaei, H., Subedi, M., Nickerson, M., Martínez-Villaluenga, C., Frias, J., & Vandenberg, A. (2019). Seed Protein of Lentils: Current Status, Progress, and Food Applications. Foods, 8(9), 391. https://doi.org/10.3390/foods8090391