The Influence of Selected Plant Essential Oils on Morphological and Physiological Characteristics in Pseudomonas Orientalis

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacteria Strains
2.2. Essential Oils
2.3. Chemicals
2.4. Estimation of Bioactive Compounds Content in Investigated Essential Oils by Gas Chromatography
2.5. Agar Disk Diffusion Assay
2.6. Determination of Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
2.7. Flow Cytometry Analysis
2.8. Observations of Morphological Changes in Bacterial Cells
2.9. Evaluation of Membrane Integrity
2.10. Statistical Analysis
3. Results and Discussion
3.1. Effect of Essential Oils on Bacterial Growth
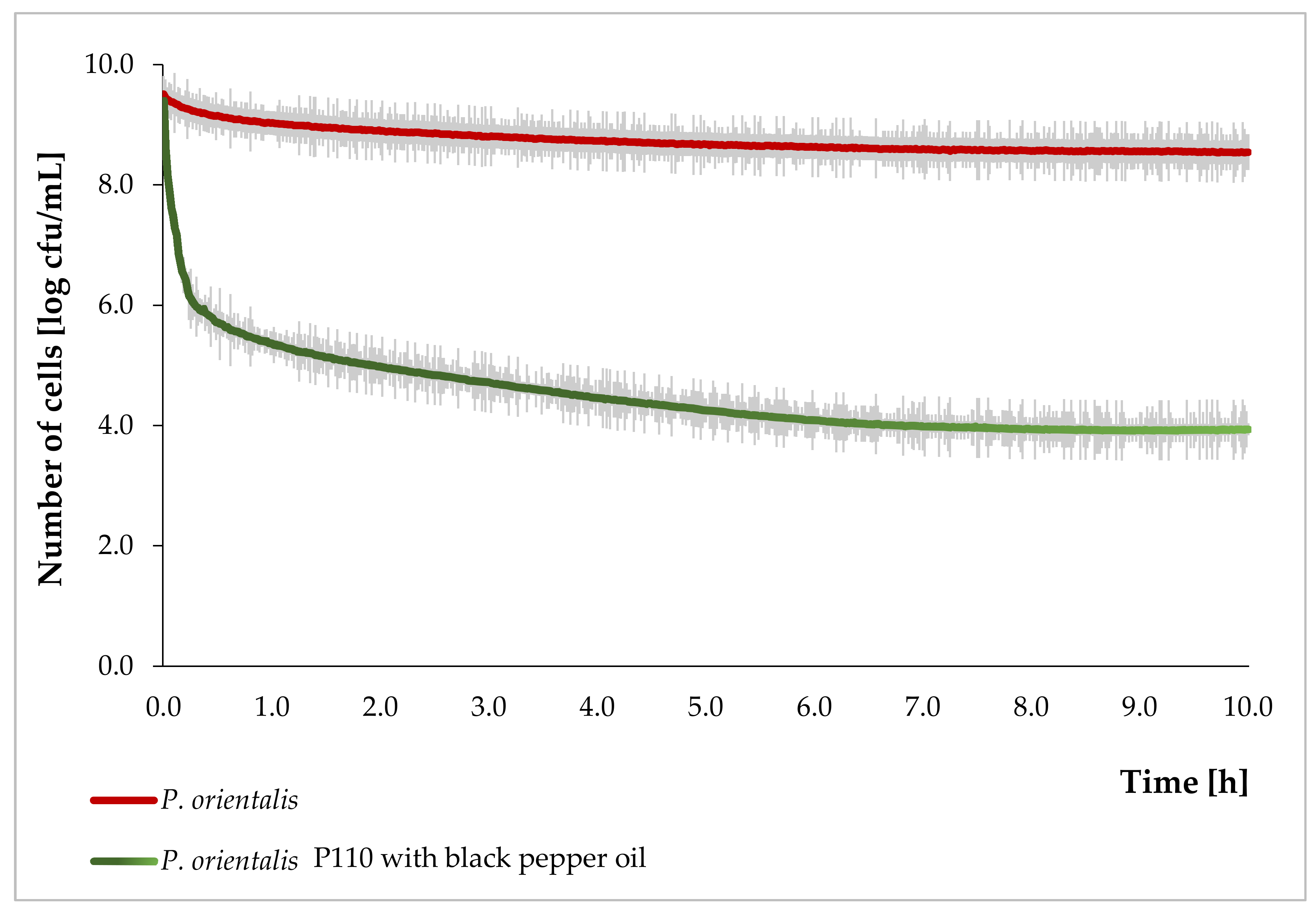
3.2. The Growth Curve of P. Orientalis P110
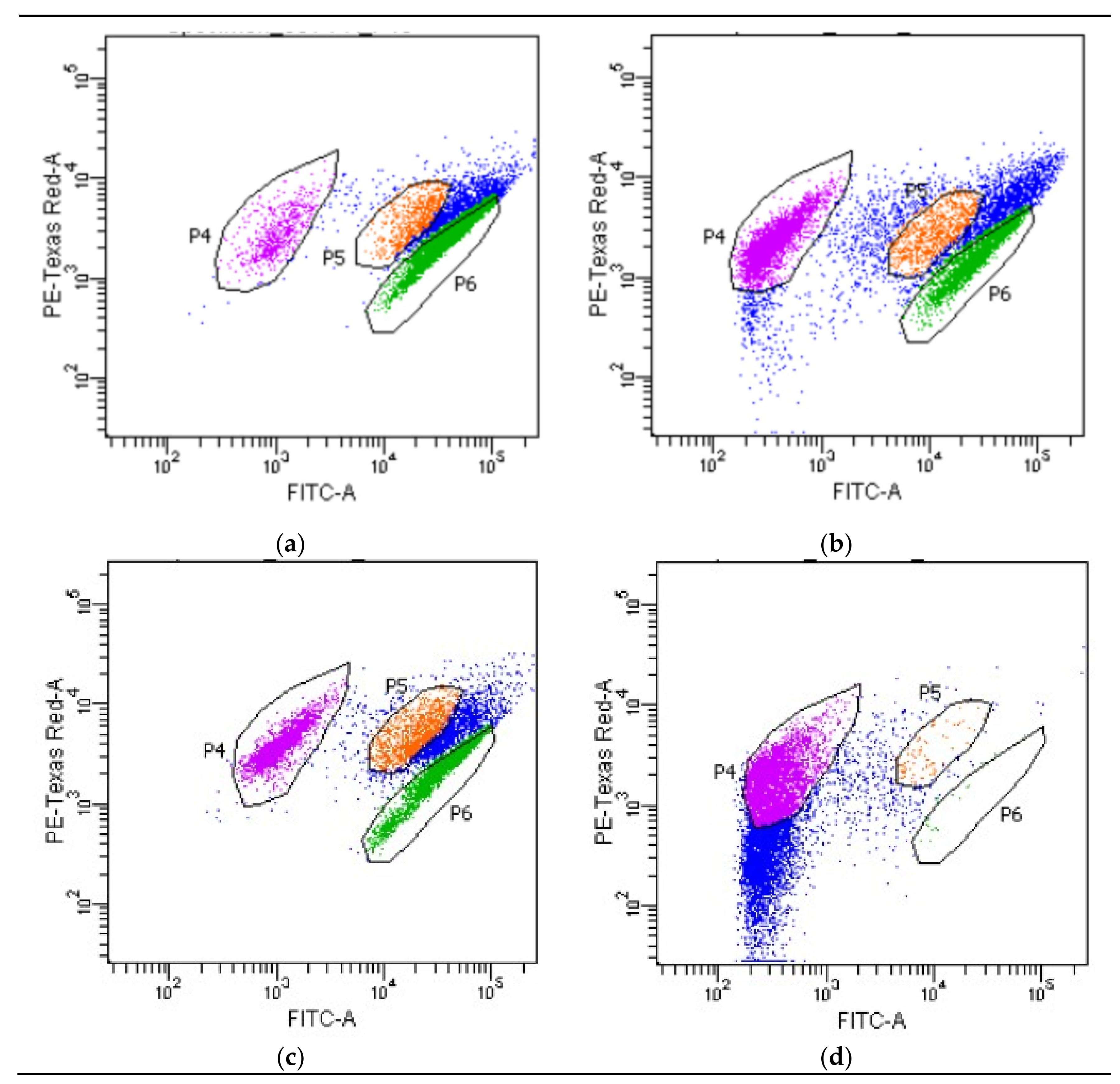
3.3. Investigation of Metabolic Activity of Cells Using a Flow Cytometer
3.4. Influence of the Temperature and Time Incubation on the Efficiency of Essential Oil Antibacterial Action
3.5. The Mode of Action of Essential Oils
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential Oils in Food Preservation: Mode of Action, Synergies, and Interactions with Food Matrix Components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Baj, T.; Korona-Głowniak, I.; Kowalski, R.; Malm, A. Chemical composition and microbiological evaluation of essential oil from Hyssopus officinalis L. with white and pink flowers. Open Chem. 2018, 16, 317–323. [Google Scholar] [CrossRef]
- Stappen, I.; Tabanca, N.; Ali, A.; Wanner, J.; Lal, B.; Jaitak, V.; Wedge, D.E.; Kaul, V.K.; Schmidt, E.; Jirovetz, L. Antifungal and repellent activities of the essential oils from three aromatic herbs from western Himalaya. Open Chem. 2018, 16, 306–316. [Google Scholar] [CrossRef]
- Aziz, M.; Karboune, S. Natural antimicrobial/antioxidant agents in meat and poultry products as well as fruits and vegetables: A review. Crit. Rev. Food Sci. Nutr. 2016, 58, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Smyth, C.; Brunton, N.; Fogarty, C.; Bolton, D. The Effect of Organic Acid, Trisodium Phosphate and Essential Oil Component Immersion Treatments on the Microbiology of Cod (Gadus morhua) during Chilled Storage. Foods 2018, 7, 200. [Google Scholar] [CrossRef]
- Tajkarimi, M.M.; Ibrahim, S.A.; Cliver, D.O. Antimicrobial herb and spice compounds in food. Food Control 2010, 21, 1199–1218. [Google Scholar] [CrossRef]
- Balpinar, N. The biological activities of Arabis alpina L. subsp. brevifolia (DC.) Cullen against food pathogens. Open Chem. 2018, 16, 930–936. [Google Scholar] [CrossRef]
- Peng, L.; Xiong, Y.; Wang, M.; Han, M.; Cai, W.; Li, Z. Chemical composition of essential oil in Mosla chinensis Maxim cv. Jiangxiangru and its inhibitory effect on Staphylococcus aureus biofilm formation. Open Life Sci. 2018, 13, 1–10. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of Essential Oils on Pathogenic Bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential Oils as Antimicrobial Agents—Myth or Real Alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef]
- Pandey, A.K.; Kumar, P.; Singh, P.; Tripathi, N.N.; Bajpai, V.K. Essential Oils: Sources of Antimicrobials and Food Preservatives. Front. Microbiol. 2017, 7, 2161. [Google Scholar] [CrossRef] [PubMed]
- Quendera, A.P.; Barreto, A.S.; Semedo-Lemsaddek, T. Antimicrobial activity of essential oils against foodborne multidrug-resistant enterococci and aeromonads in planktonic and biofilm state. Food Sci. Technol. Int. 2019, 25, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Mittal, R.P.; Rana, A.; Jaitak, V. Essential Oils: An Impending Substitute of Synthetic Antimicrobial Agents to Overcome Antimicrobial Resistance. Curr. Drug Targets 2019, 20, 605–624. [Google Scholar] [CrossRef] [PubMed]
- Restrepo, A.E.; Rojas, J.D.; García, O.R.; Sánchez, L.T.; Pinzón, M.I.; Villa, C.C. Mechanical, barrier, and color properties of banana starch edible films incorporated with nanoemulsions of lemongrass (Cymbopogon citratus) and rosemary (Rosmarinus officinalis) essential oils. Food Sci. Technol. Int. 2018, 24, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Prudent, D.; Perineau, F.; Bessiere, J.M.; Michel, G.M.; Baccou, J.C. Analysis of the Essential Oil of Wild Oregano from Martinique (Coleus aromaticus Benth.)—Evaluation of Its Bacteriostatic and Fungistatic Properties. J. Essent. Oil Res. 1995, 7, 165–173. [Google Scholar] [CrossRef]
- Calo, J.R.; Crandall, P.G.; O’Bryan, C.A.; Ricke, S.C. Essential oils as antimicrobials in food systems—A review. Food Control 2015, 54, 111–119. [Google Scholar] [CrossRef]
- Myszka, K.; Leja, K.; Majcher, M. A current opinion on the antimicrobial importance of popular pepper essential oil and its application in food industry. J. Essent. Oil Res. 2019, 31, 1–18. [Google Scholar] [CrossRef]
- Leja, K.; Szudera-Kończal, K.; Myszka, K.; Czaczyk, K. Antibacterial effect of natural oils—an opportunity to solve the problem of antibiotic resistance on the example of pseudomonas spp. Postep. Mikrobiol. 2019, 58, 177–190. [Google Scholar] [CrossRef]
- Dabboussi, F.; Hamze, M.; Izard, D.; Fouad, D. Analysis of the virulence characteristics of three new species of the genus Pseudomonas isolated from Lebanese spring waters. J. Eur. D’hydrologie 2000, 31, 239–247. [Google Scholar] [CrossRef]
- Kačániová, M.; Terentjeva, M.; Vukovic, N.; Puchalski, C.; Roychoudhury, S.; Kunová, S.; Klūga, A.; Tokár, M.; Kluz, M.; Ivanišová, E. The antioxidant and antimicrobial activity of essential oils against Pseudomonas spp. isolated from fish. Saudi Pharm. J. 2017, 25, 1108–1116. [Google Scholar] [CrossRef]
- Leja, K.; Drożdżyńska, A.; Majcher, M.; Kowalczewski, P.Ł.; Czaczyk, K. Influence of sub-inhibitory concentration of selected plant essential oils on physical and biochemical properties of Pseudomonas orientalis strains. Open Chem. 2019, 17, 492–505. [Google Scholar] [CrossRef]
- Bauer, A.W.; Perry, D.M.; Kirby, W.M. Single-disk antibiotic-sensitivity testing of staphylococci; an analysis of technique and results. AMA Arch. Intern. Med. 1959, 104, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Weerakkody, N.S.; Caffin, N.; Turner, M.S.; Dykes, G.A. In vitro antimicrobial activity of less-utilized spice and herb extracts against selected food-borne bacteria. Food Control 2010, 21, 1408–1414. [Google Scholar] [CrossRef]
- Kondratowicz, A.; Weiss, M.; Juzwa, W.; Majchrzycki, Ł.; Lewandowicz, G. Characteristics of liposomes derived from egg yolk. Open Chem. 2019. [Google Scholar] [CrossRef]
- Lv, F.; Liang, H.; Yuan, Q.; Li, C. In vitro antimicrobial effects and mechanism of action of selected plant essential oil combinations against four food-related microorganisms. Food Res. Int. 2011, 44, 3057–3064. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Borowska, D.; Jabłoński, A.; Pejsak, B. Disk diffusion method in veterinary diagnostics—practical data. Życie Weter. 2014, 89, 116–120. [Google Scholar]
- Prabuseenivasan, S.; Jayakumar, M.; Ignacimuthu, S. In vitro antibacterial activity of some plant essential oils. BMC Complement. Altern. Med. 2006, 6, 39. [Google Scholar] [CrossRef]
- Ghabraie, M.; Vu, K.D.; Tata, L.; Salmieri, S.; Lacroix, M. Antimicrobial effect of essential oils in combinations against five bacteria and their effect on sensorial quality of ground meat. LWT—Food Sci. Technol. 2016, 66, 332–339. [Google Scholar] [CrossRef]
- Alizadeh Sani, M.; Ehsani, A.; Hashemi, M. Whey protein isolate/cellulose nanofibre/TiO2 nanoparticle/rosemary essential oil nanocomposite film: Its effect on microbial and sensory quality of lamb meat and growth of common foodborne pathogenic bacteria during refrigeration. Int. J. Food Microbiol. 2017, 251, 8–14. [Google Scholar] [CrossRef]
- Elbanna, K.; Assiri, A.M.A.; Tadros, M.; Khider, M.; Assaeedi, A.; Mohdaly, A.A.A.; Ramadan, M.F. Rosemary (Rosmarinus officinalis) oil: Composition and functionality of the cold-pressed extract. J. Food Meas. Charact. 2018, 12, 1601–1609. [Google Scholar] [CrossRef]
- Wahab, A.; Haq, R.U.; Ahmed, A.; Khan, R.A.; Raza, M. Anticonvulsant activities of nutmeg oil of Myristica fragrans. Phyther. Res. 2009, 23, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Nabiha, B.; Abdelfatteh, E.O.; Faten, K.; Hervé, C.; Moncef, C.M. Chemical Composition of Bergamot (Citrus Bergamia Risso) Essential Oil Obtained by Hydrodistillation. J. Chem. Chem. Eng. 2010, 4, 29. [Google Scholar]
- Bassolé, I.H.N.; Lamien-Meda, A.; Bayala, B.; Obame, L.C.; Ilboudo, A.J.; Franz, C.; Novak, J.; Nebié, R.C.; Dicko, M.H. Chemical composition and antimicrobial activity of Cymbopogon citratus and Cymbopogon giganteus essential oils alone and in combination. Phytomedicine 2011, 18, 1070–1074. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.D.; Markham, J.L. Susceptibility and intrinsic tolerance of Pseudomonas aeruginosa to selected plant volatile compounds. J. Appl. Microbiol. 2007, 103, 930–936. [Google Scholar] [CrossRef] [PubMed]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial Properties of Plant Essential Oils against Human Pathogens and Their Mode of Action: An Updated Review. Evidence-Based Complement. Altern. Med. 2016, 2016, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Belitz, H.D.; Grosch, W.; Schieberle, P. Food Chemistry; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 978-3-540-69933-0. [Google Scholar]
- Dorman, H.J.D.; Deans, S.G. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef] [PubMed]
- O’ Keeffe, E.; Hughes, H.; McLoughlin, P.; Tan, S.P.; McCarthy, N. Methods of analysis for the in vitro and in vivo determination of the fungicidal activity of seaweeds: A mini review. J. Appl. Phycol. 2019, 1–18. [Google Scholar] [CrossRef]
- Hammer, K.A.; Carson, C.F.; Riley, T.V. Antimicrobial activity of essential oils and other plant extracts. J. Appl. Microbiol. 1999, 86, 985–990. [Google Scholar] [CrossRef]
- Naik, M.I.; Fomda, B.A.; Jaykumar, E.; Bhat, J.A. Antibacterial activity of lemongrass (Cymbopogon citratus) oil against some selected pathogenic bacterias. Asian Pac. J. Trop. Med. 2010, 3, 535–538. [Google Scholar] [CrossRef]
- Nguefack, J.; Budde, B.B.; Jakobsen, M. Five essential oils from aromatic plants of Cameroon: their antibacterial activity and ability to permeabilize the cytoplasmic membrane of Listeria innocua examined by flow cytometry. Lett. Appl. Microbiol. 2004, 39, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Ferreira, S.; Queiroz, J.A.; Domingues, F.C. Coriander (Coriandrum sativum L.) essential oil: its antibacterial activity and mode of action evaluated by flow cytometry. J. Med. Microbiol. 2011, 60, 1479–1486. [Google Scholar] [CrossRef] [PubMed]
- Paparella, A.; Taccogna, L.; Aguzzi, I.; Chaves-López, C.; Serio, A.; Marsilio, F.; Suzzi, G. Flow cytometric assessment of the antimicrobial activity of essential oils against Listeria monocytogenes. Food Control 2008, 19, 1174–1182. [Google Scholar] [CrossRef]
- Muñoz, M.; Guevara, L.; Palop, A.; Tabera, J.; Fernández, P.S. Determination of the effect of plant essential oils obtained by supercritical fluid extraction on the growth and viability of Listeria monocytogenes in broth and food systems using flow cytometry. LWT—Food Sci. Technol. 2009, 42, 220–227. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, C.; Xu, P.; Wang, X.C. Mechanisms of ultraviolet disinfection and chlorination of Escherichia coli: Culturability, membrane permeability, metabolism, and genetic damage. J. Environ. Sci. 2018, 65, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Wojtyś, A.; Jankowski, T. Effect of Temperature on the Rate of Penetration of Selected Essential Oils into Yeast Cells (in Polish). Żywność. Nauk. Technol. Jakość. 2004, 3, 77–86. [Google Scholar]
- Hassoun, A.; Emir Çoban, Ö. Essential oils for antimicrobial and antioxidant applications in fish and other seafood products. Trends Food Sci. Technol. 2017, 68, 26–36. [Google Scholar] [CrossRef]
- Kalchayanand, N.; Dunne, P.; Sikes, A.; Ray, B. Viability loss and morphology change of foodborne pathogens following exposure to hydrostatic pressures in the presence and absence of bacteriocins. Int. J. Food Microbiol. 2004, 91, 91–98. [Google Scholar] [CrossRef]
- Gennari, M.; Dragotto, F. A study of the incidence of different fluorescent Pseudomonas species and biovars in the microflora of fresh and spoiled meat and fish, raw milk, cheese, soil and water. J. Appl. Bacteriol. 1992, 72, 281–288. [Google Scholar] [CrossRef]
- Hall, C.W.; Hinz, A.J.; Gagnon, L.B.-P.; Zhang, L.; Nadeau, J.-P.; Copeland, S.; Saha, B.; Mah, T.-F. Pseudomonas aeruginosa Biofilm Antibiotic Resistance Gene ndvB Expression Requires the RpoS Stationary-Phase Sigma Factor. Appl. Environ. Microbiol. 2018, 84, e02762-17. [Google Scholar] [CrossRef]
- Devi, K.P.; Nisha, S.A.; Sakthivel, R.; Pandian, S.K. Eugenol (an essential oil of clove) acts as an antibacterial agent against Salmonella typhi by disrupting the cellular membrane. J. Ethnopharmacol. 2010, 130, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Y.; Shi, Y.; Pan, X.; Lu, Y.; Cao, P. Antibacterial effects of cinnamon (Cinnamomum zeylanicum) bark essential oil on Porphyromonas gingivalis. Microb. Pathog. 2018, 116, 26–32. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Essential Oil/Bioactive Compounds | Bacteria Strain | Zone of Growth Inhibition [mm] | MIC Value [μL/mL] 36 °C, 24 h | MIC Value [μL/mL] 15 °C, 72 h | MIC Value [mg/mL] 36 °C, 24 h | MIC Value [mg/mL] 15 °C, 72 h | The Degree of Inhibition of Bacterial Growth [%] 36 °C, 24 h | The Degree of Inhibition of Bacterial Growth [%] 15 °C, 72 h |
|---|---|---|---|---|---|---|---|---|
| Bergamot | P. orientalis P49 | 5.0 c ± 0.6 | 8.3 a ± 0.4 | 90.9 a ± 2.7 | 7.5 a ± 0.5 | 82.5 a ± 3.1 | 33.6 b ± 2.1 | 41.9 d ± 2.6 |
| P. orientalis P110 | 0 a | - | - | - | - | 0 a | 0 a | |
| Nutmeg | P. orientalis P49 | 0 a | - | - | - | - | 0 a | 0 a |
| P. orientalis P110 | 0 a | - | - | - | - | 0 a | 0 a | |
| bitter orange | P. orientalis P49 | 9.1 d ± 0.9 | 8.3 a ± 0.3 | 90.9 a ± 4.6 | 6.9 a ± 0.2 | 76.3 a ± 2.2 | 62.7 c ± 1.4 | 2.7 b ± 1.5 |
| P. orientalis P110 | 0 a | - | - | - | - | 0 a | 0 a | |
| Lime | P. orientalis P49 | 0 a | - | - | - | - | 0 a | 0 a |
| P. orientalis P110 | 19.2 d ± 1.0 | 90.9 b ± 7.6 | 166.7 a ± 8.9 | 81.5 b ± 6.5 | 149.5 a ± 10.2 | 85.0 d ± 0.2 | 25.7 c ± 2.4 | |
| Lemongrass | P. orientalis P49 | 11.1 d ± 1.1 | 8.3 a ± 0.4 | 90.9 a ± 8.3 | 7.6 a ± 0.2 | 83.7 a ± 7.6 | 48.5 c ± 1.8 | 30.3 c ± 3.1 |
| P. orientalis P110 | 18.1 d ± 1.9 | 8.3 a ± 0.6 | 166.7 a ± 11.2 | 7.6 a ± 0.3 | 153.5 a ± 9.8 | 62.7 c ± 5.7 | 13.4 b ± 4.5 | |
| Juniper | P. orientalis P49 | 2.0 b ± 0.3 | 90.9 b ± 5.1 | - | 76.0 b ± 8.1 | - | 40.7 c ± 2.5 | 0 a |
| P. orientalis P110 | 2.1 b ± 0.4 | 90.9 b ± 4.3 | - | 76.0 b ± 9.3 | - | 54.9 c ± 1.5 | 0 a | |
| black pepper | P. orientalis P49 | 2.0 b ± 0.2 | 90.9 b ± 5.9 | - | 78.7 b ± 2.6 | - | 49.9 c ± 0.7 | 0 a |
| P. orientalis P110 | 0 a | 8.3 a ± 0.2 | - | 7.2 a ± 0.3 | - | 64.5 c ± 2.9 | 0 a | |
| Rosemary | P. orientalis P49 | 0 a | - | - | - | - | 0 a | 0 a |
| P. orientalis P110 | 7.0 c ± 1.0 | 90.9 b ± 7.1 | 166.7 a ± 14.2 | 80.1 b ± 6.3 | 146.9 a ± 10.8 | 64.5 c ± 2.6 | 11.3 b ± 3.7 | |
| St. John’s wort | P. orientalis P49 | 2.0 b ± 0.3 | - | - | - | - | 0 a | 0 a |
| P. orientalis P110 | 2.0 b ± 0.1 | - | - | - | - | 0 a | 0 a | |
| Linalool | P. orientalis P49 | 0 a | - | - | - | - | 0 a | 0 a |
| P. orientalis P110 | 8.0 c ± 1.6 | 90.9 b ± 8.8 | - | 78.5 b ± 3.7 | - | 46.8 b ± 8.9 | 0 a | |
| Citral | P. orientalis P49 | 0 a | - | - | - | - | 0 a | 0 a |
| P. orientalis P110 | 0 a | - | - | - | - | 0 a | 0 a |
| Bioactive Compound/Oil Type | Rosemary [%] | Bitter Orange [%] | Black Pepper [%] | Bergamot [%] | Nutmeg [%] | Lemongrass [%] | Lime [%] | Juniper [%] |
|---|---|---|---|---|---|---|---|---|
| α-pinene | 19.0 ± 1.0 | 0.6 ± 0.2 | 10.0 ± 2.0 | nd | 17.5 ± 1.5 | nd | 1.75 ± 0.75 | 1.75 ± 0.75 |
| Camphene | 3.0 ± 1.0 | 0.5 ± 0.3 | nd | nd | nd | nd | nd | nd |
| ß-pinene | 9.5 ± 1.5 | 0.08 ± 0.07 | 9.5 ± 2.5 | 7.5 ± 2.0 | 14.5 ± 1.0 | nd | 21.5 ± 3.5 | 21.5 ± 3.5 |
| Myrcene | 1.75 ± 0.25 | 2.25 ± 0.75 | nd | nd | 2.2 ± 0.4 | nd | 1.75 ± 0.75 | 1.75 ± 0.75 |
| Limonene | 3.0 ± 0.5 | 94.5 ± 1.5 | 13.5 ± 3.5 | 34.5 ± 4.5 | nd | 1.85 ± 0.65 | nd | nd |
| Cineol | 19.0 ± 1.0 | nd | nd | nd | nd | nd | nd | nd |
| p-cymene | 1.6 ± 0.6 | nd | nd | nd | nd | nd | nd | nd |
| Camphor | 20.5 ± 0.5 | nd | nd | nd | nd | nd | nd | nd |
| Bornyl acetate | 0.7 ± 0.2 | nd | nd | nd | nd | nd | nd | nd |
| α-terpineol | 1.3 ± 0.3 | nd | nd | nd | 5.0 ± 1.0 | nd | nd | nd |
| Borneol | 3.75 ± 0.75 | nd | nd | nd | nd | nd | nd | nd |
| Verbenone | 1.2 ± 0.5 | nd | nd | nd | nd | nd | nd | nd |
| Linalool | nd | 0.42 ± 0.28 | nd | 12.0 ± 3.0 | nd | nd | nd | nd |
| Decanal | nd | 0.40 ± 0.30 | nd | nd | nd | 1.6 ± 0.1 | nd | nd |
| Neral | nd | 0.22 ± 0.19 | nd | nd | nd | 30.0 ± 2.0 | nd | nd |
| Geranial | nd | 0.13 ± 0.08 | nd | 0.36 ± 0.13 | nd | 37.0 ± 2.0 | nd | nd |
| ß-sinensal | nd | 0.21 ± 0.20 | 2.0 ± 1.5 | nd | nd | nd | nd | nd |
| Delta-3-Carene | nd | nd | 10.5 ± 1.5 | nd | 1.75 ± 0.95 | nd | nd | nd |
| ß-Caryophyllene | nd | nd | 24.5 ± 4.5 | nd | nd | nd | nd | nd |
| Linalyl acetate | nd | nd | 28.0 ± 6.0 | nd | nd | nd | nd | nd |
| γ-terpinene | nd | nd | nd | 8.0 ± 2.0 | 4.4 ± 0.8 | nd | 21.5 ± 3.5 | 21.5 ± 3.5 |
| β-bisabolene | nd | nd | nd | 0.43 ± 0.13 | nd | nd | nd | nd |
| Sabinene | nd | nd | nd | nd | 18.0 ± 1.0 | nd | nd | nd |
| Citronellol | nd | nd | nd | nd | nd | nd | nd | nd |
| Eugenol | nd | nd | nd | nd | 0.50 ± 0.01 | nd | nd | nd |
| Myristicin | nd | nd | nd | nd | 9.5 ± 1.5 | nd | nd | nd |
| α-thujene | nd | nd | nd | nd | 1.85 ± 0.25 | nd | nd | nd |
| Methyl heptanone | nd | nd | nd | nd | nd | 1.05 ± 0.95 | nd | nd |
| Citral | nd | nd | nd | nd | nd | 68.5 ± 1.5 | 10.0 ± 5.7 | 10.15 ± 5.85 |
| Geranyl acetate | nd | nd | nd | nd | nd | 2.0 ± 1.5 | nd | nd |
| Essential Oil | P. Orientalis Strain | Low Metabolic Activity [%] 36 °C, 24 h | Low Metabolic Activity [%] 15 °C, 72 h | Medium Metabolic Activity [%] 36 °C, 24 h | Medium Metabolic Activity [%] 15 °C, 72 h | High Metabolic Activity [%] 36 °C, 24 h | High Metabolic Activity [%] 15 °C, 72 h |
|---|---|---|---|---|---|---|---|
| control probe | P49 | 7.1 a ± 0.2 | 2.3 a ± 0.3 | 7.2 a ± 0.1 | 9.2 a ± 0.6 | 67.8 c ± 0.7 | 80.1 c ± 0.8 |
| P110 | 10.0 a ± 0.4 | 1.4 a ± 0.2 | 15.0 b ± 0.5 | 4.2 a ± 0.1 | 65.0 c ± 0.3 | 86.1 b ± 0.6 | |
| black pepper | P49 | 10.3 a ± 0.1 | 2.2 a ± 0.2 | 8.2 a ± 0.6 | 6.8 a ± 1.2 | 65.9 c ± 0.4 | 83.1 c ± 2.2 |
| P110 | 11.8 a ± 0.4 | 2.0 a ± 0.1 | 7.9 a ± 0.1 | 2.5 a ± 0.2 | 54.9 c ± 0.6 | 91.1 c ± 0.2 | |
| lemongrass | P49 | 29.0 b ± 0.3 | 36.7 b ± 0.5 | 13.2 b ± 0.2 | 24.6 c ± 0.6 | 47.8 b ± 0.5 | 25.9 a ± 0.4 |
| P110 | 52.b ± 0.7 | 6.8 a ± 0.5 | 10.6 a ± 0.1 | 5.8 a ± 0.4 | 6.4 a ± 0.2 | 78.3 a ± 0.5 | |
| Juniper | P49 | 9.2 a ± 0.2 | 4.2 a ± 0.1 | 8.1 a ± 0.2 | 18.1 b ± 2.6 | 67.3 c ± 1.0 | 62.8 b ± 3.3 |
| P110 | 16.4 a ± 0.2 | 2.7 a ± 0.1 | 10.1 a ± 0.1 | 6.0 a ± 0.6 | 46.8 b ± 0.3 | 82.4 b ± 1.1 | |
| bergamot | P49 | 31.2 b ± 0.4 | 2.5 a ± 0.3 | 9.2 a ± 0.5 | 11.7 b ± 1.0 | 29.8 a ± 0.9 | 73.2 b ± 1.0 |
| bitter orange | 5.2 a ± 0.1 | 3.8 a ± 0.7 | 5.3 a ± 0.2 | 15.8 b ± 1.4 | 60.4 c ± 1.2 | 72.1 a ± 1.1 | |
| Lime | P110 | 23.0 b ± 0.2 | 5.3 a ± 0.6 | 7.6 a ± 0.4 | 9.2 a ± 0.7 | 25.0 a ± 0.8 | 73.3 b ± 0.4 |
| rosemary | 47.8 b ± 0.6 | 3.2 a ± 0.3 | 10.6 a | 9.5 a ± 0.3 | 0.2 a ± 0.1 | 75.6 a ± 0.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leja, K.; Szudera-Kończal, K.; Świtała, E.; Juzwa, W.; Kowalczewski, P.Ł.; Czaczyk, K. The Influence of Selected Plant Essential Oils on Morphological and Physiological Characteristics in Pseudomonas Orientalis. Foods 2019, 8, 277. https://doi.org/10.3390/foods8070277
Leja K, Szudera-Kończal K, Świtała E, Juzwa W, Kowalczewski PŁ, Czaczyk K. The Influence of Selected Plant Essential Oils on Morphological and Physiological Characteristics in Pseudomonas Orientalis. Foods. 2019; 8(7):277. https://doi.org/10.3390/foods8070277
Chicago/Turabian StyleLeja, Katarzyna, Kamila Szudera-Kończal, Ewa Świtała, Wojciech Juzwa, Przemysław Łukasz Kowalczewski, and Katarzyna Czaczyk. 2019. "The Influence of Selected Plant Essential Oils on Morphological and Physiological Characteristics in Pseudomonas Orientalis" Foods 8, no. 7: 277. https://doi.org/10.3390/foods8070277
APA StyleLeja, K., Szudera-Kończal, K., Świtała, E., Juzwa, W., Kowalczewski, P. Ł., & Czaczyk, K. (2019). The Influence of Selected Plant Essential Oils on Morphological and Physiological Characteristics in Pseudomonas Orientalis. Foods, 8(7), 277. https://doi.org/10.3390/foods8070277