The Effect of Sleep Curtailment on Hedonic Responses to Liquid and Solid Food


Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Development of Stimuli
2.3. Study Timeline
2.4. Consent Visit
2.5. Laboratory Visits
2.6. Sensory Evaluation
2.7. Statistical Analysis
3. Results
3.1. Participants
3.2. Summary of Curtailment
3.3. Summary of Affect, Cravings and Hunger
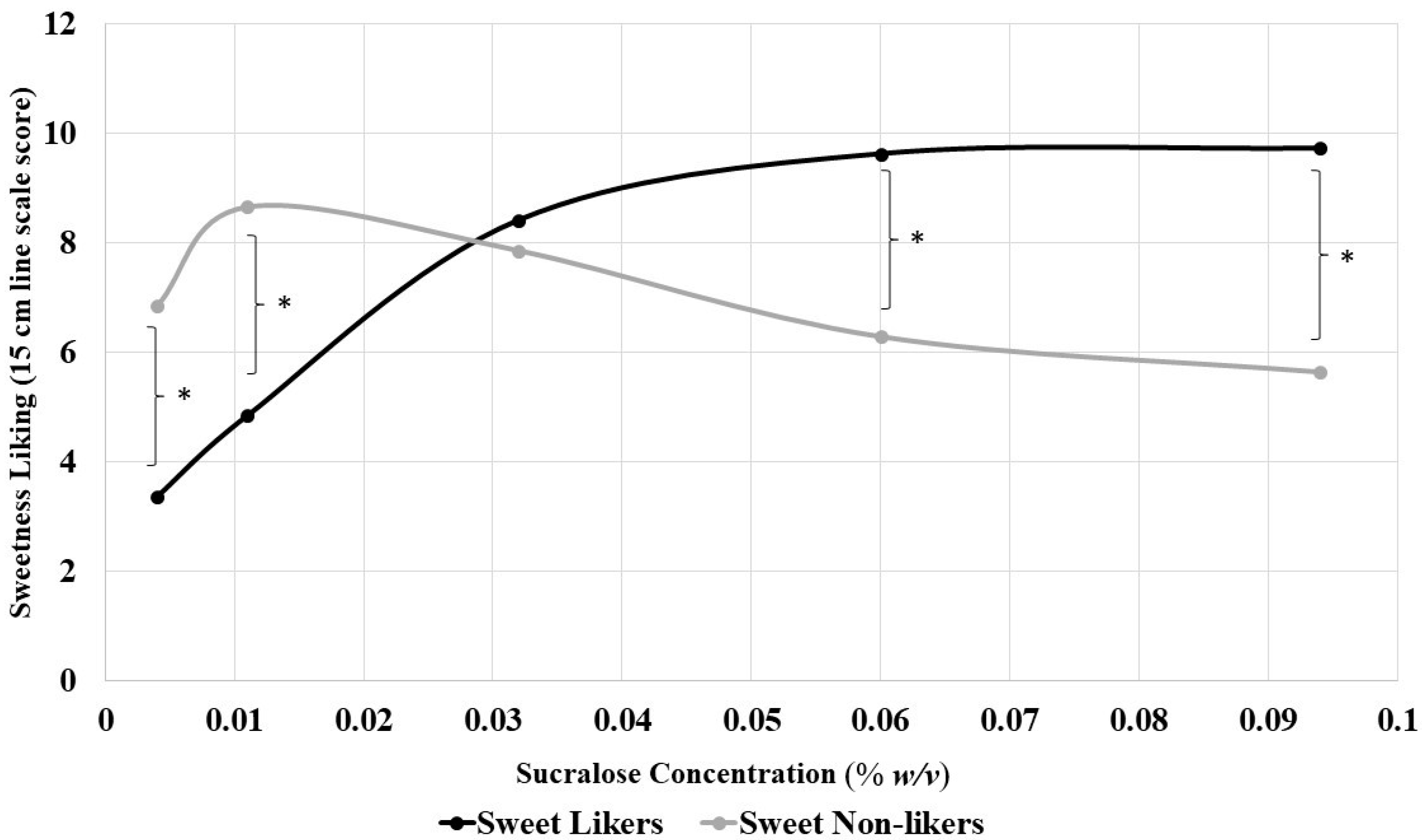
3.4. Sweet Liking Phenotypes
3.5. Sweetness Perception in the Model System
3.5.1. Model System Sweet Preference
3.5.2. Model System Sweet Liking Slopes
3.5.3. Model System Sweet Liking by Concentration
3.6. Hedonic Response in the Oat Product Systems
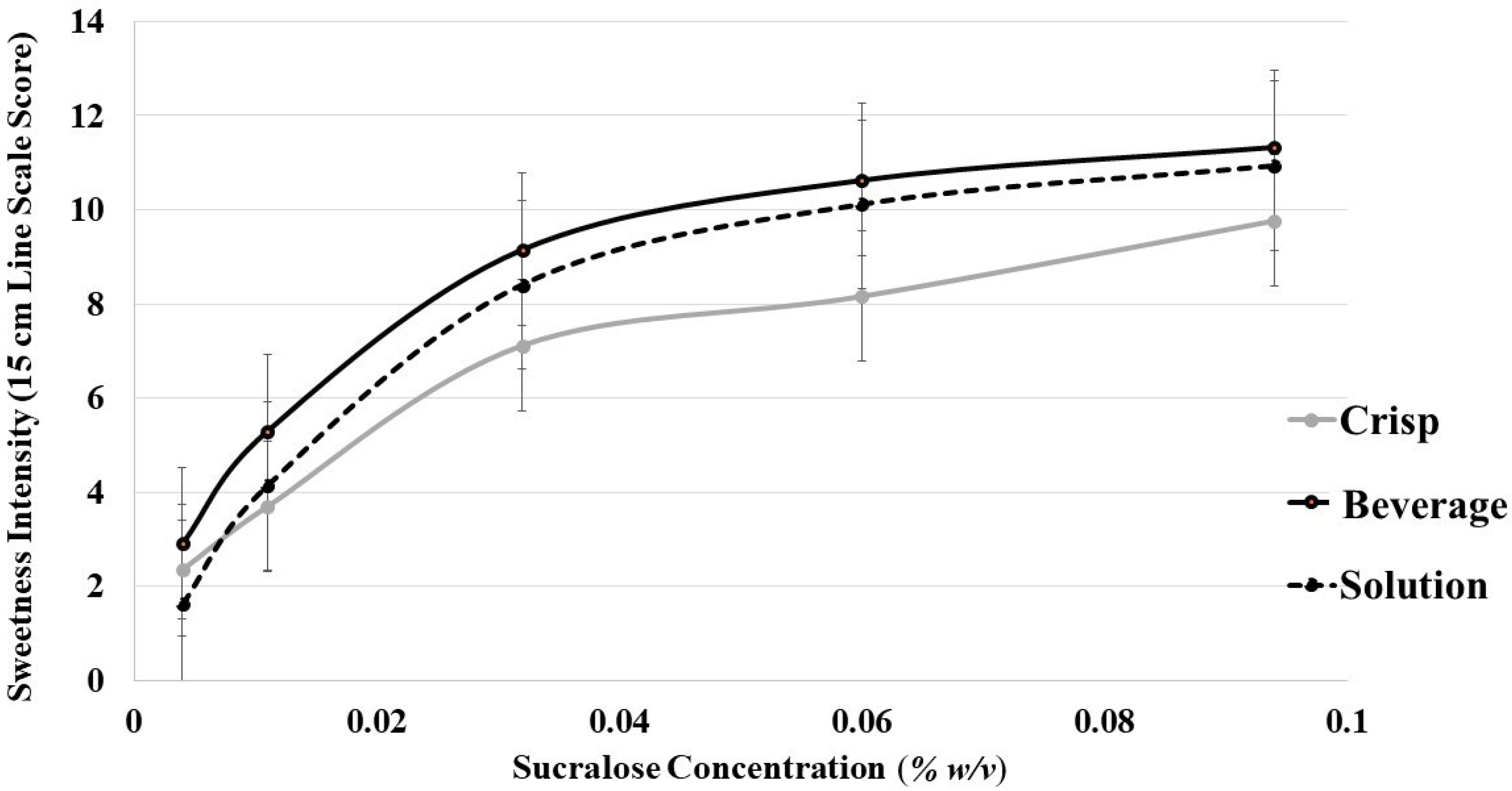
3.6.1. Oat Product Sweetness Intensity
3.6.2. Oat Product Liking Slopes
3.6.3. Oat Product Liking by Concentration
4. Discussion
Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Szczygiel, E.J.; Cho, S.; Tucker, R.M. Characterization of the relationships between sleep duration, quality, architecture and chemosensory function in non-obese females. Chem. Senses 2018, 43, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Szczygiel, E.J.; Cho, S.; Snyder, M.K.; Tucker, R.M. Associations between chemosensory function, sweet taste preference, and the previous night’s sleep in non-obese males. Food Qual. Prefer. 2019, 75, 105–112. [Google Scholar] [CrossRef]
- Smith, S.L.; Ludy, M.-J.; Tucker, R.M. Changes in taste preference and steps taken after sleep curtailment. Physiol. Behav. 2016, 163, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Lv, W.; Finlayson, G.; Dando, R. Sleep, food cravings and taste. Appetite 2018, 125, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.L.; Field, J.; Miller, L.E.; DiFrancesco, M.; Beebe, D.W. Sweet/dessert foods are more appealing to adolescents after sleep restriction. PLoS ONE 2015, 10, e0115434. [Google Scholar] [CrossRef]
- Nedeltcheva, A.V.; Kilkus, J.M.; Imperial, J.; Kasza, K.; Schoeller, D.A.; Penev, P.D. Sleep curtailment is accompanied by increased intake of calories from snacks. Am. J. Clin. Nutr. 2009, 89, 126–133. [Google Scholar] [CrossRef]
- Markwald, R.R.; Melanson, E.L.; Smith, M.R.; Higgins, J.; Perreault, L.; Eckel, R.H.; Wright, K.P. Impact of insufficient sleep on total daily energy expenditure, food intake, and weight gain. Proc. Natl. Acad. Sci. USA 2013, 110, 5695–5700. [Google Scholar] [CrossRef]
- Hanlon, E.C.; Andrzejewski, M.E.; Harder, B.K.; Kelley, A.E.; Benca, R.M. The effect of REM sleep deprivation on motivation for food reward. Behav. Brain Res. 2005, 163, 58–69. [Google Scholar] [CrossRef]
- Greer, S.M.; Goldstein, A.N.; Walker, M.P. The impact of sleep deprivation on food desire in the human brain. Nat. Commun. 2013, 4, 2259. [Google Scholar] [CrossRef]
- Benedict, C.; Brooks, S.J.; O’Daly, O.G.; Almèn, M.S.; Morell, A.; Åberg, K.; Gingnell, M.; Schultes, B.; Hallschmid, M.; Broman, J.-E.; et al. Acute sleep deprivation enhances the brain’s response to hedonic food stimuli: An fMRI study. J. Clin. Endocrinol. Metab. 2012, 97, E443–E447. [Google Scholar] [CrossRef]
- Demos, K.E.; Sweet, L.H.; Hart, C.N.; McCaffery, J.M.; Williams, S.E.; Mailloux, K.A.; Trautvetter, J.; Owens, M.M.; Wing, R.R. The effects of experimental manipulation of sleep duration on neural response to food cues. Sleep 2017, 40, zsx125. [Google Scholar] [CrossRef] [PubMed]
- St-Onge, M.-P.; Wolfe, S.; Sy, M.; Shechter, A.; Hirsch, J. Sleep restriction increases the neuronal response to unhealthy food in normal-weight individuals. Int. J. Obes. 2014, 38, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Drewnowski, A. Intense sweeteners and energy density of foods: Implications for weight control. Eur. J. Clin. Nutr. 1999, 53, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Drewnowski, A.; Mennella, J.A.; Johnson, S.L.; Bellisle, F. Sweetness and food preference. J. Nutr. 2012, 142, 1142S–1148S. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T. Brain Mechanisms of sweetness and palatability of sugars. Nutr. Rev. 2003, 61, S5–S9. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Gelaye, B.; Williams, M.A. Sleep characteristics and health-related quality of life among a national sample of American young adults: Assessment of possible health disparities. Qual. Life Res. 2014, 23, 613–625. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of Obesity among Adults and Youth: United States, 2015–2016; U.S Department of Health & Human Services: Atlanta, GA, USA, 2017.
- Furihata, R.; Uchiyama, M.; Suzuki, M.; Konno, C.; Konno, M.; Takahashi, S.; Kaneita, Y.; Ohida, T.; Akahoshi, T.; Hashimoto, S.; et al. Association of short sleep duration and short time in bed with depression: A Japanese general population survey. Sleep Biol. Rhythm. 2015, 13, 136–145. [Google Scholar] [CrossRef]
- Lin, C.-L.; Lin, C.-P.; Chen, S.-W.; Wu, H.-C.; Tsai, Y.-H. The association between sleep duration and overweight or obesity in Taiwanese adults: A cross-sectional study. Obes. Res. Clin. Pract. 2018, 12, 384–388. [Google Scholar] [CrossRef]
- Hogenkamp, P.S.; Nilsson, E.; Chapman, C.D.; Cedernaes, J.; Vogel, H.; Dickson, S.L.; Broman, J.-E.; Schiöth, H.B.; Benedict, C. Sweet taste perception not altered after acute sleep deprivation in healthy young men. Somnologie 2013, 17, 111–114. [Google Scholar] [CrossRef][Green Version]
- Szczygiel, E.J.; Cho, S.; Tucker, R.M. Multiple dimensions of sweet taste perception altered after sleep curtailment. Nutrients 2019, 11, 2015. [Google Scholar] [CrossRef]
- Tanaka, T.; Hong, G.; Tominami, K.; Kudo, T. Oral fat sensitivity is associated with social support for stress coping in young adult men. Tohoku J. Exp. Med. 2018, 244, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Huber, J. The Psychophysics of Taste: Perceptions of Bitterness and Sweetness in Iced Tea; Advances in Consumer Research: Ann Arbor, MI, USA, 1974; Volume NA-01, pp. 166–181. [Google Scholar]
- Mazur, J.; Drabek, R.; Goldman, A. Hedonic contrast effects in multi-product food evaluations differing in complexity. Food Qual. Prefer. 2018, 63, 159–167. [Google Scholar] [CrossRef]
- Drewnowski, A.; Shrager, E.E.; Lipsky, C.; Stellar, E.; Greenwood, M.R.C. Sugar and fat: Sensory and hedonic evaluation of liquid and solid foods. Physiol. Behav. 1989, 45, 177–183. [Google Scholar] [CrossRef]
- Tan, S.-Y.; Tucker, R.M. Sweet taste as a predictor of dietary intake: A systematic review. Nutrients 2019, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, J.; Running, C.A.; Tucker, R.M.; Mattes, R.D. Effects of food form on appetite and energy balance. Food Qual. Prefer. 2016, 48, 368–375. [Google Scholar] [CrossRef]
- Gujar, N.; Yoo, S.-S.; Hu, P.; Walker, M.P. Sleep deprivation amplifies reactivity of brain reward networks, biasing the appraisal of positive emotional experiences. J. Neurosci. 2011, 31, 4466–4474. [Google Scholar] [CrossRef] [PubMed]
- Krause, A.J.; Simon, E.B.; Mander, B.A.; Greer, S.M.; Saletin, J.M.; Goldstein-Piekarski, A.N.; Walker, M.P. The sleep-deprived human brain. Nat. Rev. Neurosci. 2017, 18, 404. [Google Scholar] [CrossRef]
- Kamiyama, H.; Iida, T.; Nishimori, H.; Kubo, H.; Uchiyama, M.; Laat, A.D.; Lavigne, G.; Komiyama, O. Effect of sleep restriction on somatosensory sensitivity in the oro-facial area: An experimental controlled study. J. Oral Rehabil. 2019, 46, 303–309. [Google Scholar] [CrossRef]
- Schloegl, H.; Percik, R.; Horstmann, A.; Villringer, A.; Stumvoll, M. Peptide hormones regulating appetite—Focus on neuroimaging studies in humans. Diabetes/Metab. Res. Rev. 2011, 27, 104–112. [Google Scholar] [CrossRef]
- Auvray, M.; Spence, C. The multisensory perception of flavor. Conscious. Cogn. 2008, 17, 1016–1031. [Google Scholar] [CrossRef]
- Roehrs, T.; Hyde, M.; Blaisdell, B.; Greenwald, M.; Roth, T. Sleep loss and REM sleep loss are Hyperalgesic. Sleep 2006, 29, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Barr, R.G.; Pantel, M.S.; Young, S.N.; Wright, J.H.; Hendricks, L.A.; Gravel, R. The response of crying newborns to sucrose: Is it a “sweetness” effect? Physiol. Behav. 1999, 66, 409–417. [Google Scholar] [CrossRef]
- Iatridi, V.; Hayes, J.E.; Yeomans, M.R. Reconsidering the classification of sweet taste liker phenotypes: A methodological review. Food Qual. Prefer. 2019, 72, 56–76. [Google Scholar] [CrossRef]
- Iatridi, V.; Hayes, J.E.; Yeomans, M.R. Quantifying sweet taste liker phenotypes: Time for some consistency in the classification criteria. Nutrients 2019, 11, 129. [Google Scholar] [CrossRef] [PubMed]
- Yeomans, M.R.; Tepper, B.J.; Rietzschel, J.; Prescott, J. Human hedonic responses to sweetness: Role of taste genetics and anatomy. Physiol. Behav. 2007, 91, 264–273. [Google Scholar] [CrossRef]
- Asao, K.; Miller, J.; Arcori, L.; Lumeng, J.C.; Han-Markey, T.; Herman, W.H. Patterns of sweet taste liking: A pilot study. Nutrients 2015, 7, 7298–7311. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Prescott, J.; Kim, K.-O. Patterns of sweet liking in sucrose solutions and beverages. Food Qual. Prefer. 2014, 36, 96–103. [Google Scholar] [CrossRef]
- Mennella, J.A.; Pepino, M.Y.; Reed, D.R. Genetic and environmental determinants of bitter perception and sweet preferences. Pediatrics 2005, 115, e216–e222. [Google Scholar] [CrossRef]
- Bachmanov, A.A.; Bosak, N.P.; Floriano, W.B.; Inoue, M.; Li, X.; Lin, C.; Murovets, V.O.; Reed, D.R.; Zolotarev, V.A.; Beauchamp, G.K. Genetics of sweet taste preferences. Flavour Fragr. J. 2011, 26, 286–294. [Google Scholar] [CrossRef]
- Garneau, N.L.; Nuessle, T.M.; Mendelsberg, B.J.; Shepard, S.; Tucker, R.M. Sweet liker status in children and adults: Consequences for beverage intake in adults. Food Qual. Prefer. 2018, 65, 175–180. [Google Scholar] [CrossRef]
- Holt, S.H.A.; Cobiac, L.; Beaumont-Smith, N.E.; Easton, K.; Best, D.J. Dietary habits and the perception and liking of sweetness among Australian and Malaysian students: A cross-cultural study. Food Qual. Prefer. 2000, 11, 299–312. [Google Scholar] [CrossRef]
- Wiet, S.G.; Beyts, P.K. Sensory characteristics of sucralose and other high intensity sweeteners. J. Food Sci. 1992, 57, 1014–1019. [Google Scholar] [CrossRef]
- Kim, M.-Y.; Cho, H.-Y.; Park, J.-Y.; Lee, S.-M.; Suh, D.-S.; Chung, S.-J.; Kim, H.-S.; Kim, K.-O. Relative sweetness of sucralose in beverage systems and sensory properties of low calorie beverages containing sucralose. Korean J. Food Sci. Technol. 2005, 37, 425–430. [Google Scholar]
- Reis, F.; De Andrade, J.; Deliza, R.; Ares, G. Comparison of two methodologies for estimating equivalent sweet concentration of high-intensity sweeteners with untrained assessors: Case study with orange/pomegranate juice. J. Sens. Stud. 2016, 31, 341–347. [Google Scholar] [CrossRef]
- Dinges, D.F.; Pack, F.; Williams, K.; Gillen, K.A.; Powell, J.W.; Ott, G.E.; Aptowicz, C.; Pack, A.I. Cumulative sleepiness, mood disturbance, and psychomotor vigilance performance decrements during a week of sleep restricted to 4–5 hours per night. Sleep 1997, 20, 267–277. [Google Scholar]
- Buysse, D.J.; Reynolds, C.F.; Monk, T.H.; Berman, S.R.; Kupfer, D.J. The Pittsburgh sleep quality index: A new instrument for psychiatric practice and research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef]
- Cohen, S.; Kamarck, T.; Mermelstein, R. A Global measure of perceived stress. J. Health Soc. Behav. 1983, 24, 385–396. [Google Scholar] [CrossRef]
- Cepeda-Benito, A.; Gleaves, D.H.; Williams, T.L.; Erath, S.A. The development and validation of the state and trait food-cravings questionnaires. Behav. Ther. 2000, 31, 151–173. [Google Scholar] [CrossRef]
- Meule, A.; Kübler, A. Double trouble. Trait food craving and impulsivity interactively predict food-cue affected behavioral inhibition. Appetite 2014, 79, 174–182. [Google Scholar] [CrossRef]
- Kaplan, R.F.; Wang, Y.; Loparo, K.A.; Kelly, M.R.; Bootzin, R.R. Performance evaluation of an automated single-channel sleep–wake detection algorithm. Nat. Sci. Sleep 2014, 6, 113–122. [Google Scholar] [CrossRef]
- Kaida, K.; Takahashi, M.; Åkerstedt, T.; Nakata, A.; Otsuka, Y.; Haratani, T.; Fukasawa, K. Validation of the Karolinska sleepiness scale against performance and EEG variables. Clin. Neurophysiol. 2006, 117, 1574–1581. [Google Scholar] [CrossRef]
- Watson, D.; Clark, L.A.; Tellegen, A. Development and validation of brief measures of positive and negative affect: The PANAS scales. J. Personal. Soc. Psychol. 1988, 54, 1063–1070. [Google Scholar] [CrossRef]
- Merrill, E.P.; Kramer, F.M.; Cardello, A.; Schutz, H. A comparison of satiety measures. Appetite 2002, 39, 181–183. [Google Scholar] [CrossRef]
- Franzen, P.L.; Siegle, G.J.; Buysse, D.J. Relationships between affect, vigilance, and sleepiness following sleep deprivation. J. Sleep Res. 2008, 17, 34–41. [Google Scholar] [CrossRef]
- Noel, C.; Dando, R. The effect of emotional state on taste perception. Appetite 2015, 95, 89–95. [Google Scholar] [CrossRef]
- Popper, R.; Rosenstock, W.; Schraidt, M.; Kroll, B.J. The effect of attribute questions on overall liking ratings. Food Qual. Prefer. 2004, 15, 853–858. [Google Scholar] [CrossRef]
- Mennella, J.A.; Lukasewycz, L.D.; Griffith, J.W.; Beauchamp, G.K. Evaluation of the Monell forced-choice, paired-comparison tracking procedure for determining sweet taste preferences across the lifespan. Chem. Senses 2011, 36, 345–355. [Google Scholar] [CrossRef]
- Glickman, M.E.; Rao, S.R.; Schultz, M.R. False discovery rate control is a recommended alternative to Bonferroni-type adjustments in health studies. J. Clin. Epidemiol. 2014, 67, 850–857. [Google Scholar] [CrossRef]
- Binns, N.M. Sucralose—All sweetness and light. Nutr. Bull. 2003, 28, 53–58. [Google Scholar] [CrossRef]
- Cheer, R.L.; Lelievre, J. Effects of sucrose on the rheological behavior of wheat starch pastes. J. Appl. Polym. Sci. 1983, 28, 1829–1836. [Google Scholar] [CrossRef]
- Van Boekel, M.A.J.S. Formation of flavour compounds in the Maillard reaction. Biotechnol. Adv. 2006, 24, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Ashoor, S.H.; Zent, J.B. Maillard browning of common amino acids and sugars. J. Food Sci. 1984, 49, 1206–1207. [Google Scholar] [CrossRef]
- Frank, G.K.W.; Oberndorfer, T.A.; Simmons, A.N.; Paulus, M.P.; Fudge, J.L.; Yang, T.T.; Kaye, W.H. Sucrose activates human taste pathways differently from artificial sweetener. NeuroImage 2008, 39, 1559–1569. [Google Scholar] [CrossRef] [PubMed]
- Sylvetsky, A.C.; Rother, K.I. Trends in the consumption of low-calorie sweeteners. Physiol. Behav. 2016, 164, 446–450. [Google Scholar] [CrossRef] [PubMed]
- MacFie, H.J.H.; Meiselman, H.L. Food Choice, Acceptance and Consumption; Springer Science & Business Media: New York, NY, USA, 2012; ISBN 978-1-46-131221-5. [Google Scholar]
- Green, E.; Murphy, C. Altered processing of sweet taste in the brain of diet soda drinkers. Physiol. Behav. 2012, 107, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Cappuccio, F.P.; Taggart, F.M.; Kandala, N.-B.; Currie, A.; Peile, E.; Stranges, S.; Miller, M.A. Meta-analysis of short sleep duration and obesity in children and adults. Sleep 2008, 31, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Alley, R.L.; Alley, T.R. The influence of physical state and color on perceived sweetness. J. Psychol. 1998, 132, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Calvino, A.M. Perception of sweetness: The effects of concentration and temperature. Physiol. Behav. 1986, 36, 1021–1028. [Google Scholar] [CrossRef]
- Schiffman, S.S.; Sattely-Miller, E.A.; Graham, B.G.; Bennett, J.L.; Booth, B.J.; Desai, N.; Bishay, I. Effect of temperature, pH, and ions on sweet taste. Physiol. Behav. 2000, 68, 469–481. [Google Scholar] [CrossRef]
- Kundermann, B.; Spernal, J.; Huber, M.T.; Krieg, J.-C.; Lautenbacher, S. Sleep deprivation affects thermal pain thresholds but not somatosensory thresholds in healthy volunteers. Psychosom. Med. 2004, 66, 932–937. [Google Scholar] [CrossRef]
- Engelen, L.; Bilt, A.V.D. Oral physiology and texture perception of semisolids. J. Texture Stud. 2008, 39, 83–113. [Google Scholar] [CrossRef]
- De Graaf, C. Why liquid energy results in overconsumption. Proc. Nutr. Soc. USA 2011, 70, 162–170. [Google Scholar] [CrossRef]
- James, B. Oral processing and texture perception influences satiation. Physiol. Behav. 2018, 193, 238–241. [Google Scholar] [CrossRef]
- Moskowitz, H.R.; Krieger, B. The contribution of sensory liking to overall liking: An analysis of six food categories. Food Qual. Prefer. 1995, 6, 83–90. [Google Scholar] [CrossRef]
- De Kermadec, F.H.; Durand, J.F.; Sabatier, R. Comparison between linear and nonlinear PLS methods to explain overall liking from sensory characteristics. Food Qual. Prefer. 1997, 8, 395–402. [Google Scholar] [CrossRef]
- Van Dongen, H.P.A.; Maislin, G.; Mullington, J.M.; Dinges, D.F. The cumulative cost of additional wakefulness: Dose-response effects on neurobehavioral functions and sleep physiology from chronic sleep restriction and total sleep deprivation. Sleep 2003, 26, 117–126. [Google Scholar] [CrossRef]
- Wever, R.A.; Polášek, J.; Wildgruber, C.M. Bright light affects human circadian rhythms. Pflug. Arch. 1983, 396, 85–87. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Oat Beverage | Oat Crisp | |
|---|---|---|
| Macronutrient | 100 kcal | 100 kcal |
| Fat | 2 g | 2 g |
| Carbohydrates | 18 g | 17 g |
| Protein | 3 g | 3 g |
| Crude Fiber | <1 g | <1 g |
| Moisture | 189 g | 1 g |
| Ash | <1 g | <1 g |
| Sex | n | % |
|---|---|---|
| Male | 15 | 37% |
| Female | 26 | 63% |
| Race | ||
| White | 27 | 66% |
| Asian | 13 | 32% |
| Other/More than 1 | 1 | 2% |
| Anthropometrics | Mean ± SD | Range |
| BMI (kg/m2) | 23.1 ± 3.0 | 16.4–29.2 |
| BF (%) | 24.8 ± 11.8 | 9.1–35.5 |
| Age (y) | 24.1 ± 5.0 | 18–41 |
| Traits/Habits | ||
| G-FCQ-T (Score) | 52.5 ± 18.5 | 23–117 |
| PSS (Score) | 12.1 ± 4.6 | 3–23 |
| PSQI (Score | 3.9 ± 1.1 | 1–5 |
| Habitual | Curtailed | % Reduction | p-Value | q-Value | ||
|---|---|---|---|---|---|---|
| Objective Sleep Measures (h) | Time in Bed | 8.3 ± 0.7 | 5.4 ± 0.7 | 34.90% | <0.001 | <0.001 |
| Total Sleep Time | 7.2 ± 0.7 | 4.5 ± 1.0 | 37.50% | <0.001 | <0.001 | |
| Light Sleep | 3.8 ± 0.5 | 2.0 ± 0.8 | 47.40% | <0.001 | <0.001 | |
| REM Sleep | 1.9 ± 0.5 | 1.2 ± 0.4 | 36.90% | <0.001 | <0.001 | |
| Slow Wave Sleep | 1.5 ± 0.4 | 1.4 ± 0.4 | 6.70% | 0.043 a | 0.053 | |
| Sleepiness (10 pt) | Karolinska Sleepiness Scale | 3.5 ± 1.4 | 5.7 ± 1.6 | <0.001 | <0.001 | |
| Subjective Previous Night’s Sleep Measures (5 pt) | Subjective Sleep Total | 13.5 ± 2.0 | 10.3 ± 2.4 | <0.001 | <0.001 | |
| How much sleep did you obtain last night? | 3.1 ± 0.4 | 1.5 ± 0.5 | <0.001 | <0.001 | ||
| How deeply did you sleep? | 3.6 ± 0.9 | 3.3 ± 1.0 | 0.243 | 0.268 | ||
| How would you rate the quality of your sleep | 3.8 ± 0.8 | 2.6 ± 1.0 | <0.001 | <0.001 | ||
| Compared to an average night, how comfortable were you when sleeping last night? | 3.0 ± 0.7 | 2.9 ± 1.0 | 0.593 | 0.593 |
| Measure | Factor | Habitual | Curtailed | p-Value | q-Value |
|---|---|---|---|---|---|
| Hunger | Hunger (100 mm VAS) | 67.1 ± 10.24 | 65.5 ± 10.3 | 0.916 | 0.916 |
| G-FCQ-S (0–15 per factor) | Total | 44.2 ± 9.7 | 46.2 ± 12.3 | 0.429 | 0.687 |
| F1-Desire to Eat | 6.1 ± 2.0 | 6.1 ± 2.2 | 0.948 | 0.916 | |
| F2-Anticipation to positive reinforcement | 8.9 ± 2.0 | 9.5 ± 2.7 | 0.232 | 0.618 | |
| F3-Anticipation to negative reinforcement | 11.2 ± 1.8 | 11.1 ± 2.6 | 0.859 | 0.916 | |
| F4-Obsessive preoccupation | 6.6 ± 2.4 | 7.4 ± 3.0 | 0.124 | 0.496 | |
| F5-Craving as a physiological state | 9.1 ± 2.0 | 9.4 ± 2.7 | 0.405 | 0.687 | |
| PANAS | Positive Affect | 23.6 ± 2.0 | 17.6 ± 6.4 | <0.001 | <0.001 |
| Negative Affect | 12.8 ± 3.9 | 13.2 ± 4.3 | 0.539 | 0.719 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczygiel, E.J.; Cho, S.; Tucker, R.M. The Effect of Sleep Curtailment on Hedonic Responses to Liquid and Solid Food. Foods 2019, 8, 465. https://doi.org/10.3390/foods8100465
Szczygiel EJ, Cho S, Tucker RM. The Effect of Sleep Curtailment on Hedonic Responses to Liquid and Solid Food. Foods. 2019; 8(10):465. https://doi.org/10.3390/foods8100465
Chicago/Turabian StyleSzczygiel, Edward J., Sungeun Cho, and Robin M. Tucker. 2019. "The Effect of Sleep Curtailment on Hedonic Responses to Liquid and Solid Food" Foods 8, no. 10: 465. https://doi.org/10.3390/foods8100465
APA StyleSzczygiel, E. J., Cho, S., & Tucker, R. M. (2019). The Effect of Sleep Curtailment on Hedonic Responses to Liquid and Solid Food. Foods, 8(10), 465. https://doi.org/10.3390/foods8100465