Identification of Rice Koji Extract Components that Increase β-Glucocerebrosidase Levels in Human Epidermal Keratinocytes

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Rice Koji Extract
2.3. Effects of Fractions of Rice Koji Extracts on Cell Proliferation and β-GCase Activity in Cultured Human Keratinocytes
2.4. Effects of Fractions from Silica Gel Column Re-Fractionated Rice Koji Extracts on Cell Proliferation and β-GCase Activity in Cultured Human Keratinocytes
2.5. TLC Analysis of Components in Rice Koji That Affect β-GCase Activity
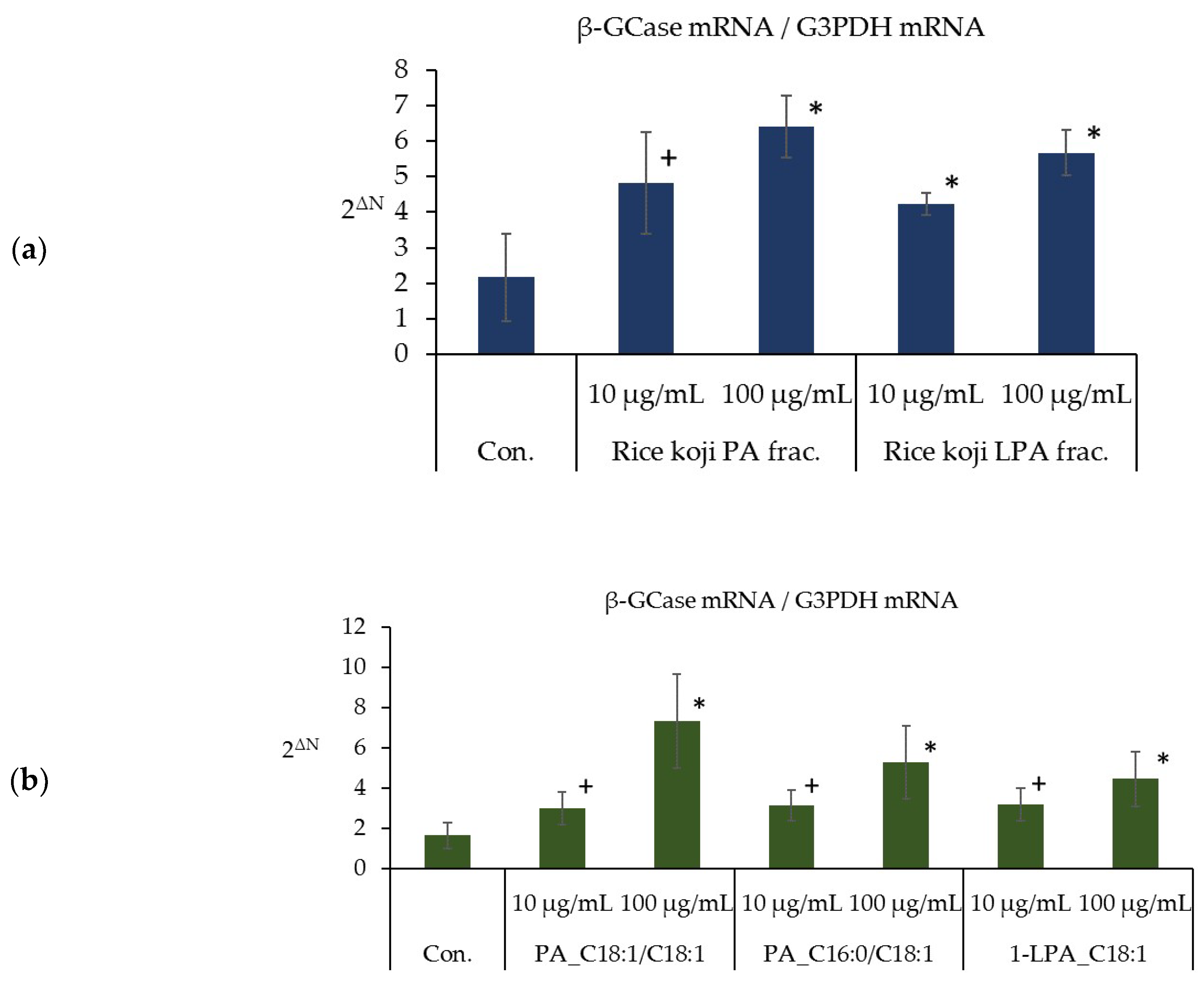
2.6. Effects of PA_C18:1/C18:1, PA_C16:0/C18:1, and 1-LPA_C18:1 on Cell Proliferation and β-GCase Activity in Cultured Human Keratinocytes
2.7. Effects of the PA and LPA Fractions of Rice Koji on TEWL on a Three-Dimensional Human Epidermis Model
2.8. Effects of PA and LPA on TEWL in a Three-Dimensional Human Epidermis Model
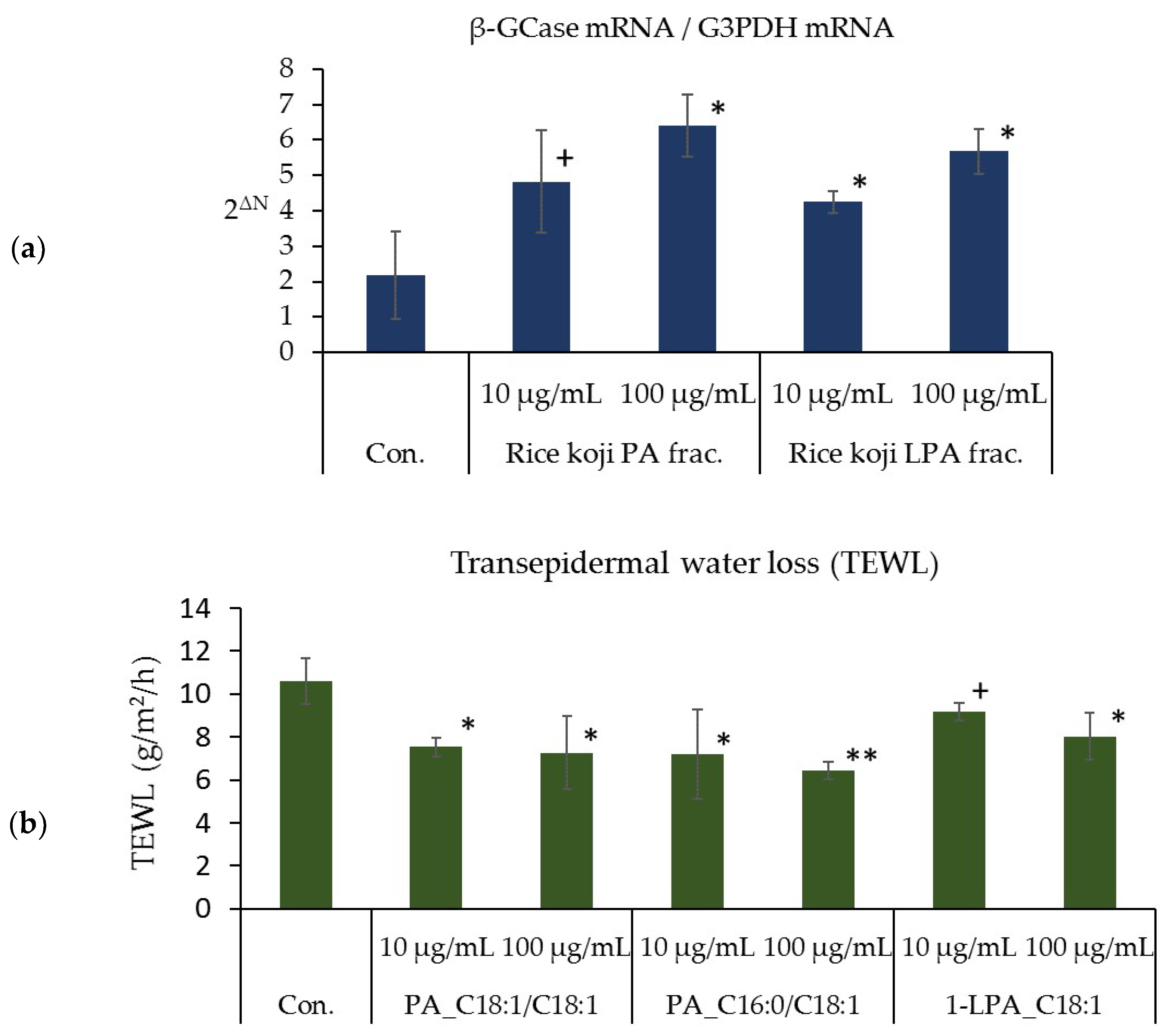
2.9. Effects of the PA and LPA Fractions of Rice Koji, PA_C18:1/C18:1, PA_C16:0/C18:1, and 1-LPA_C18:1 on the Amount of β-GCase mRNA in the Three-Dimensional Human Epidermis Model
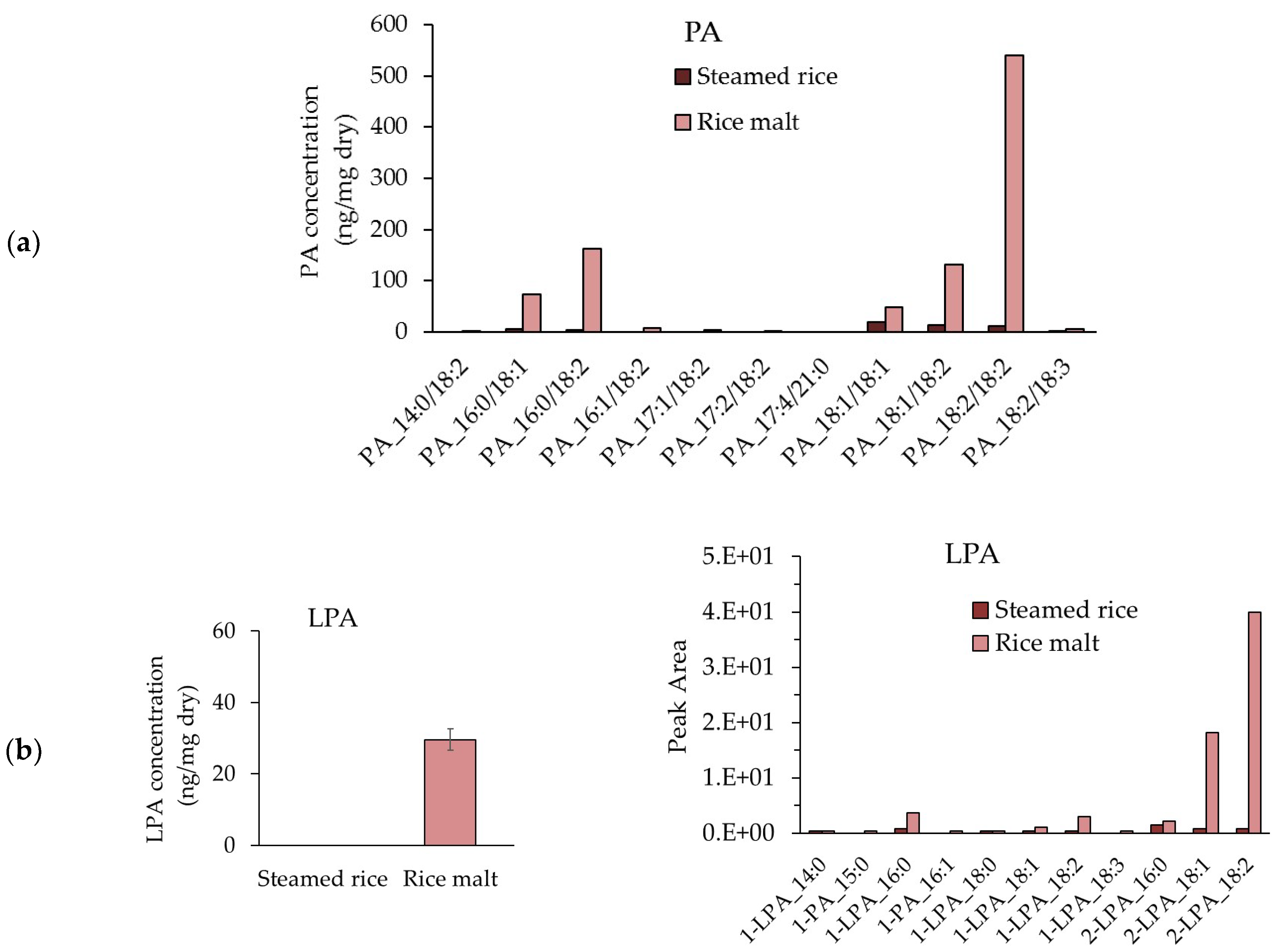
2.10. Measurement of the PA and LPA Contents in Steamed Rice and Rice Koji
| LC conditions | |
| Device | UltiMate 3000 BioRS (Thermo Fisher Scientific Inc.) |
| Column | L-column 2 ODS metal-free column (2 mm ID × 150 mm, 3 μm, Chemicals Evaluation and Research Institute, Tokyo, Japan) |
| Flow rate | 0.2 mL/min |
| Column temperature | 40 °C |
| Sampler temperature | 5 °C |
| Injection volume | 3 μL |
| MS conditions | |
| Device | 3200 QTRAP (AB Sciex Pte Ltd. Framingham, MA, USA) |
| Ionization method | Electrospray ionization |
| Measurement mode | SRM (negative) |
2.11. Measurement of the Relative LPA Contents in Steamed Rice and Rice Koji
2.12. Statistical Analysis
3. Results
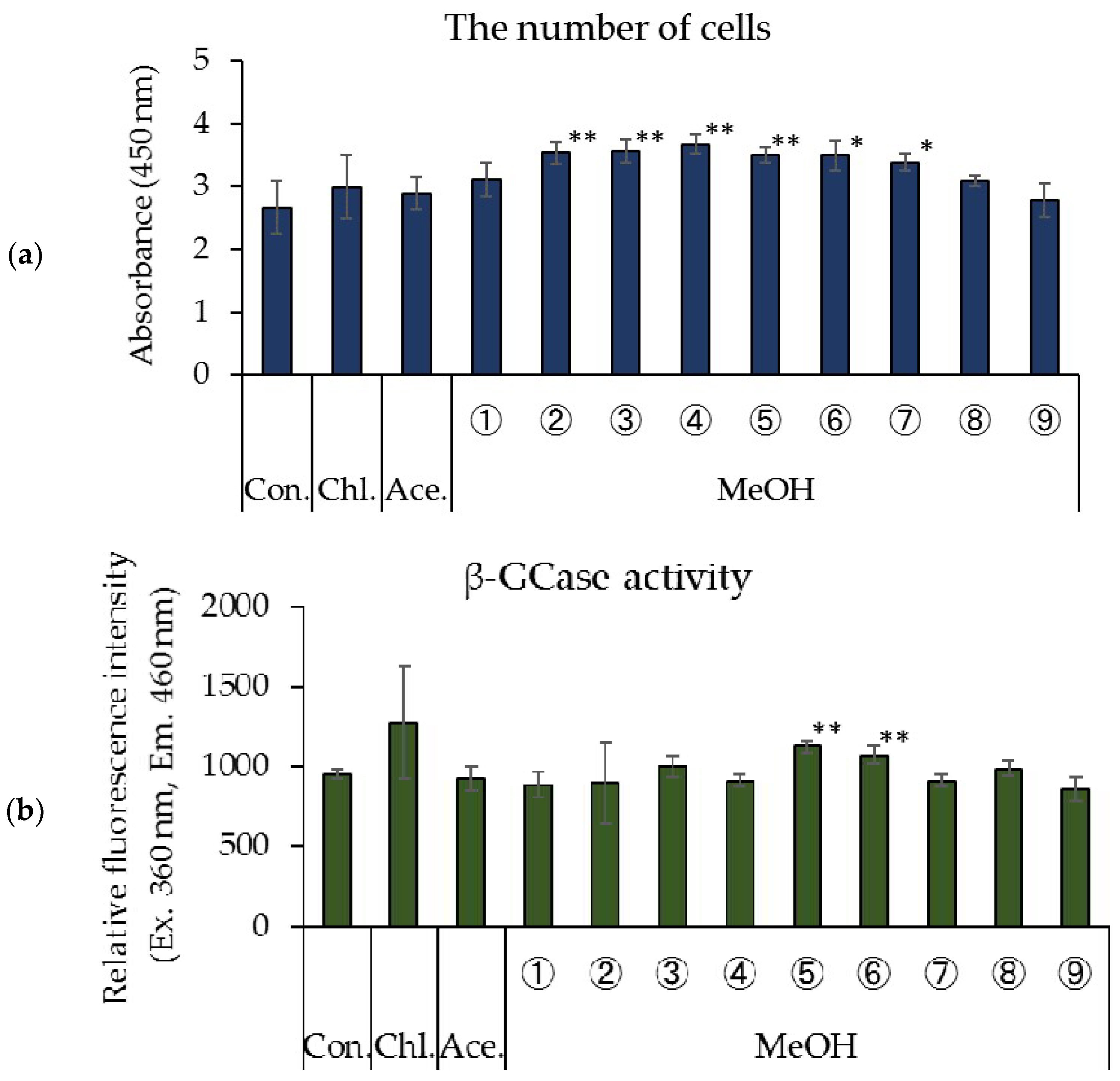
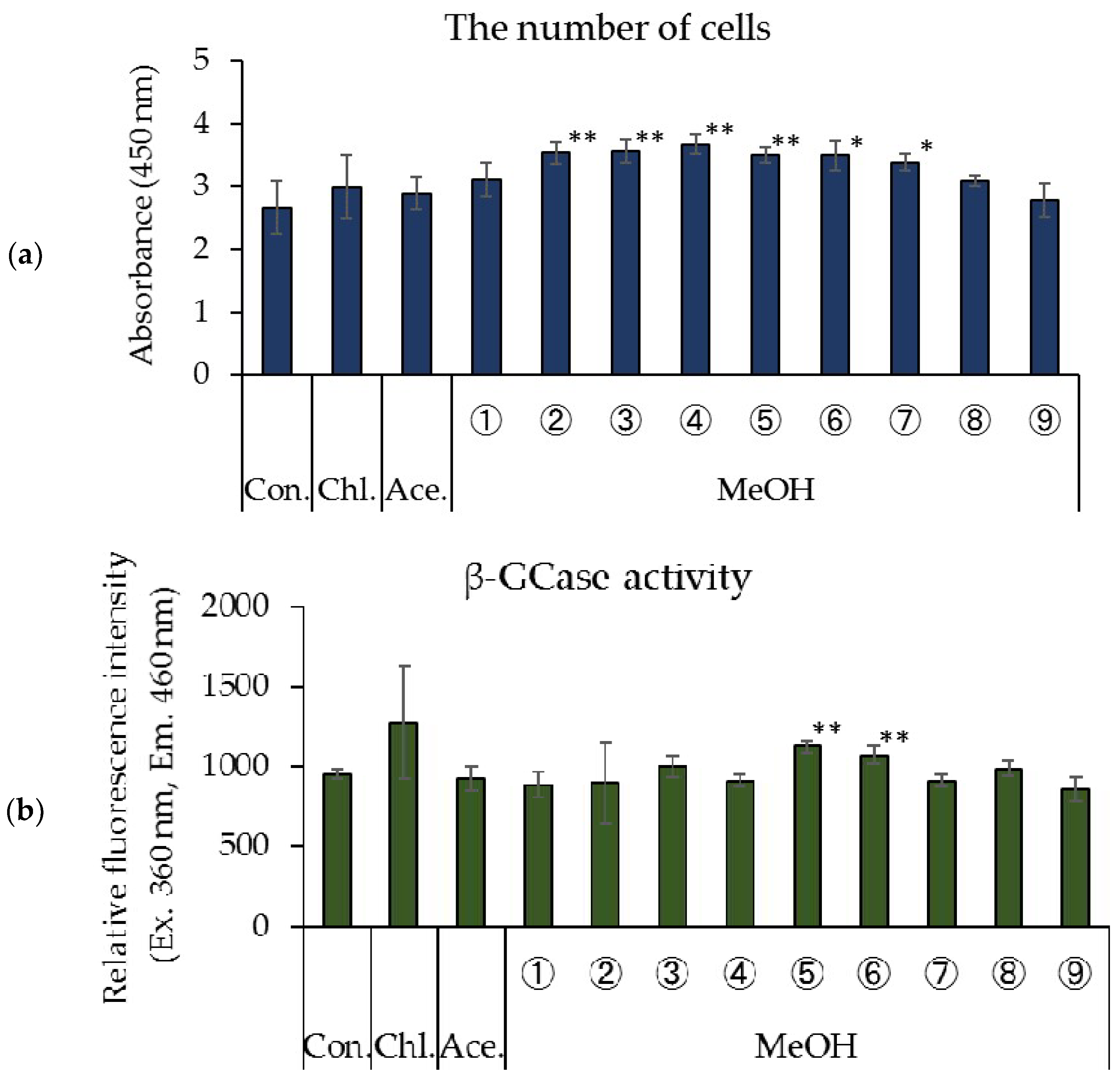
3.1. Effects of Rice Koji Extract Fractions on Cell Proliferation and β-GCase Activity in Cultured Human Keratinocytes
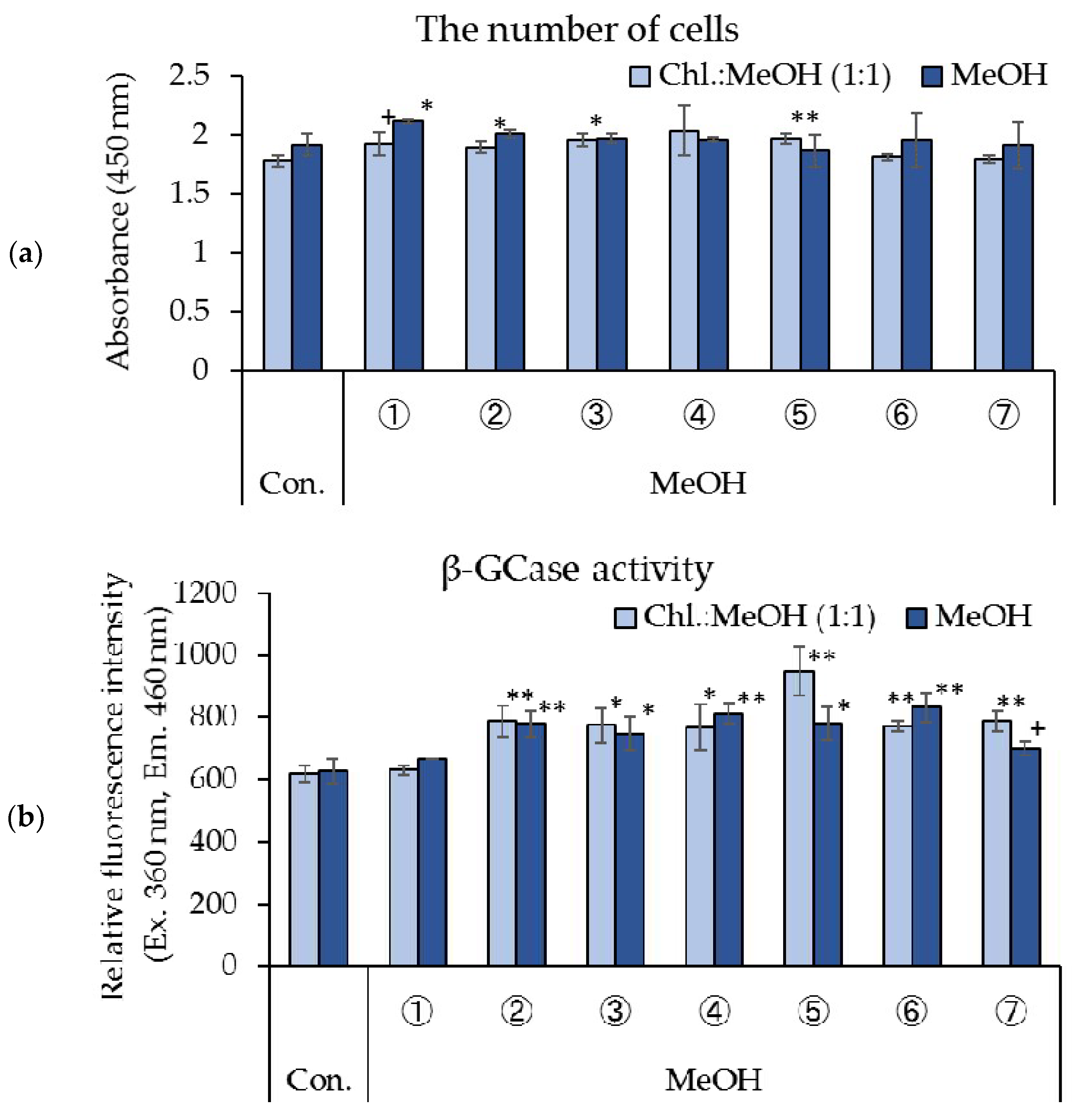
3.2. Effects of Fractions from Silica Gel Column Re-Fractionated Rice Koji Extracts on Cell Proliferation and β-GCase Activity in Cultured Human Keratinocytes
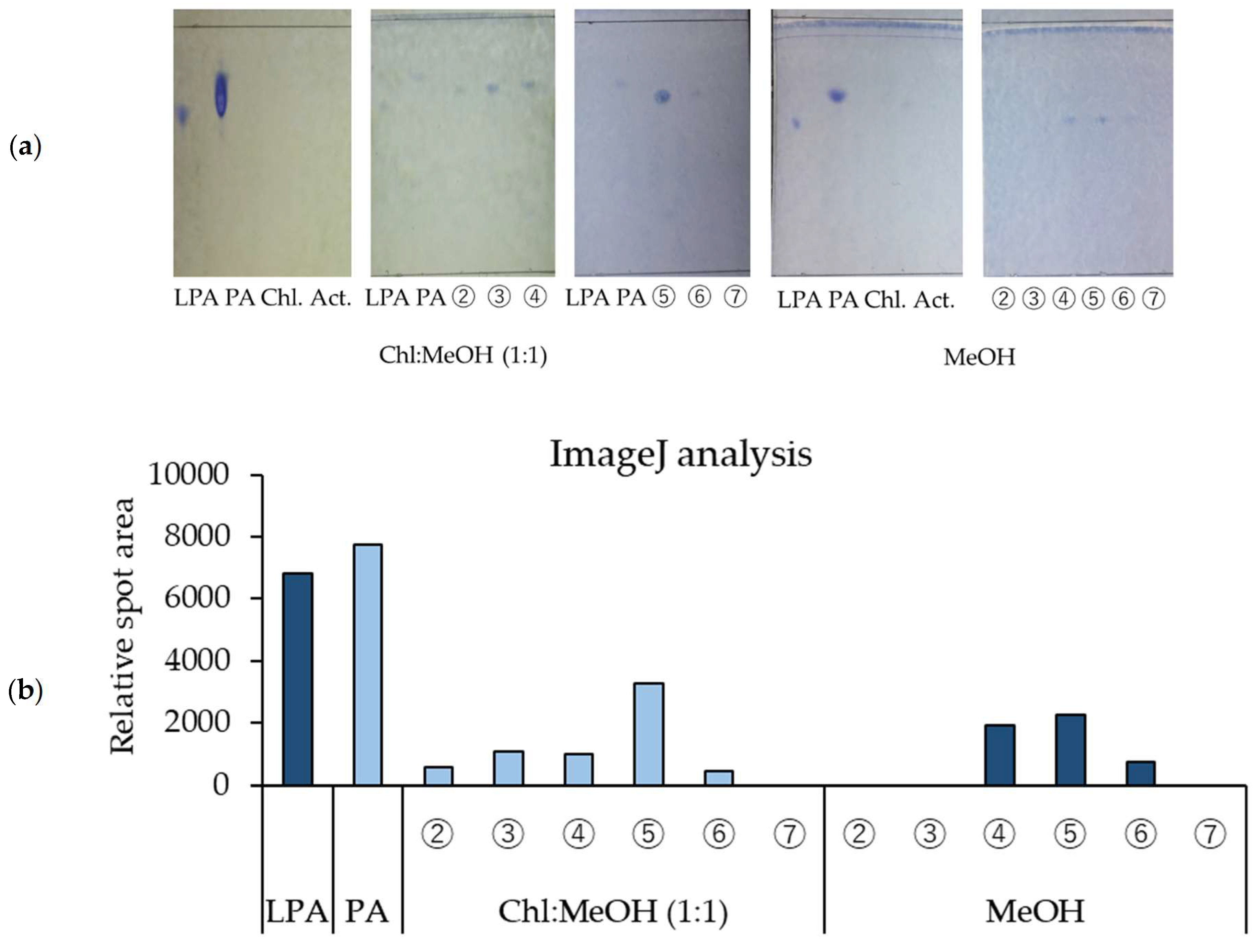
3.3. TLC Analysis of Components of Rice Koji Extract That Induce β-GCase Activity
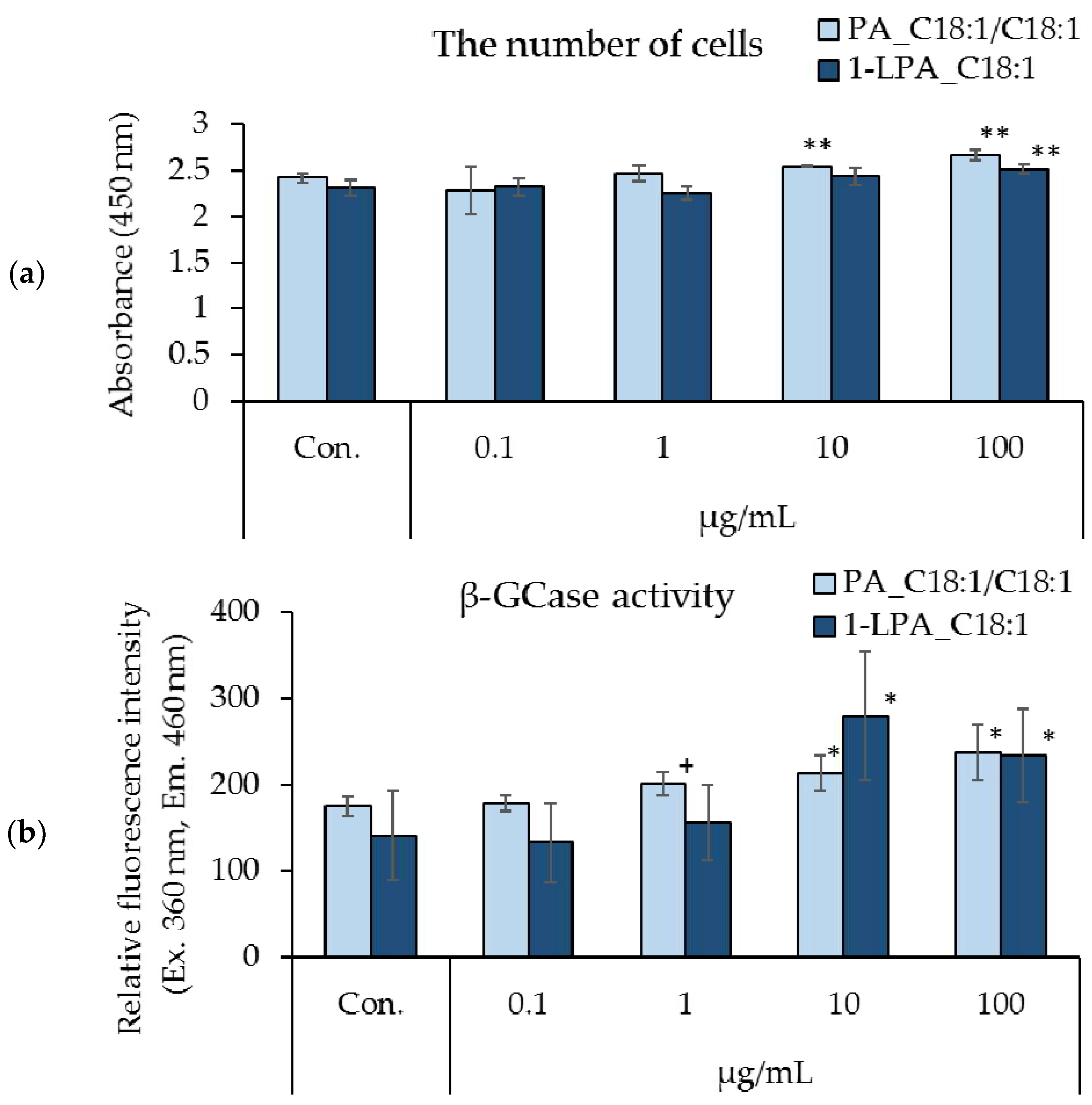
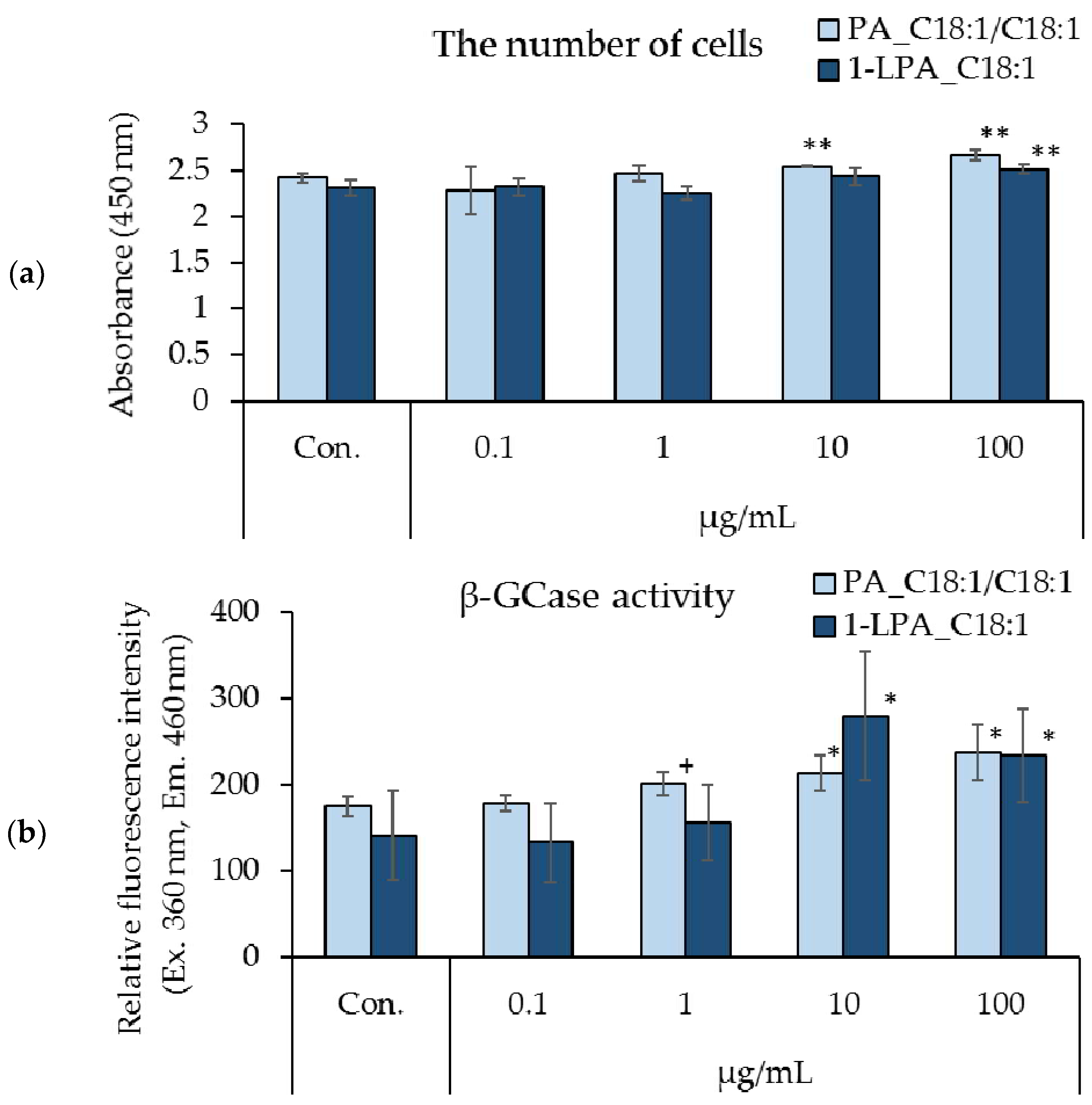
3.4. Effects of PA_C18:1/C18:1 and 1-LPA_C18:1 on Cell Proliferation and β-GCase Activity in Cultured Human Keratinocytes
3.5. Effects of the PA and LPA Fractions of Rice Koji Extract, PA_C18:1/C18:1, PA_C16:0/C18:1, and 1-LPA_C18:1 on the TEWL in the Three-Dimensional Human Epidermal Model
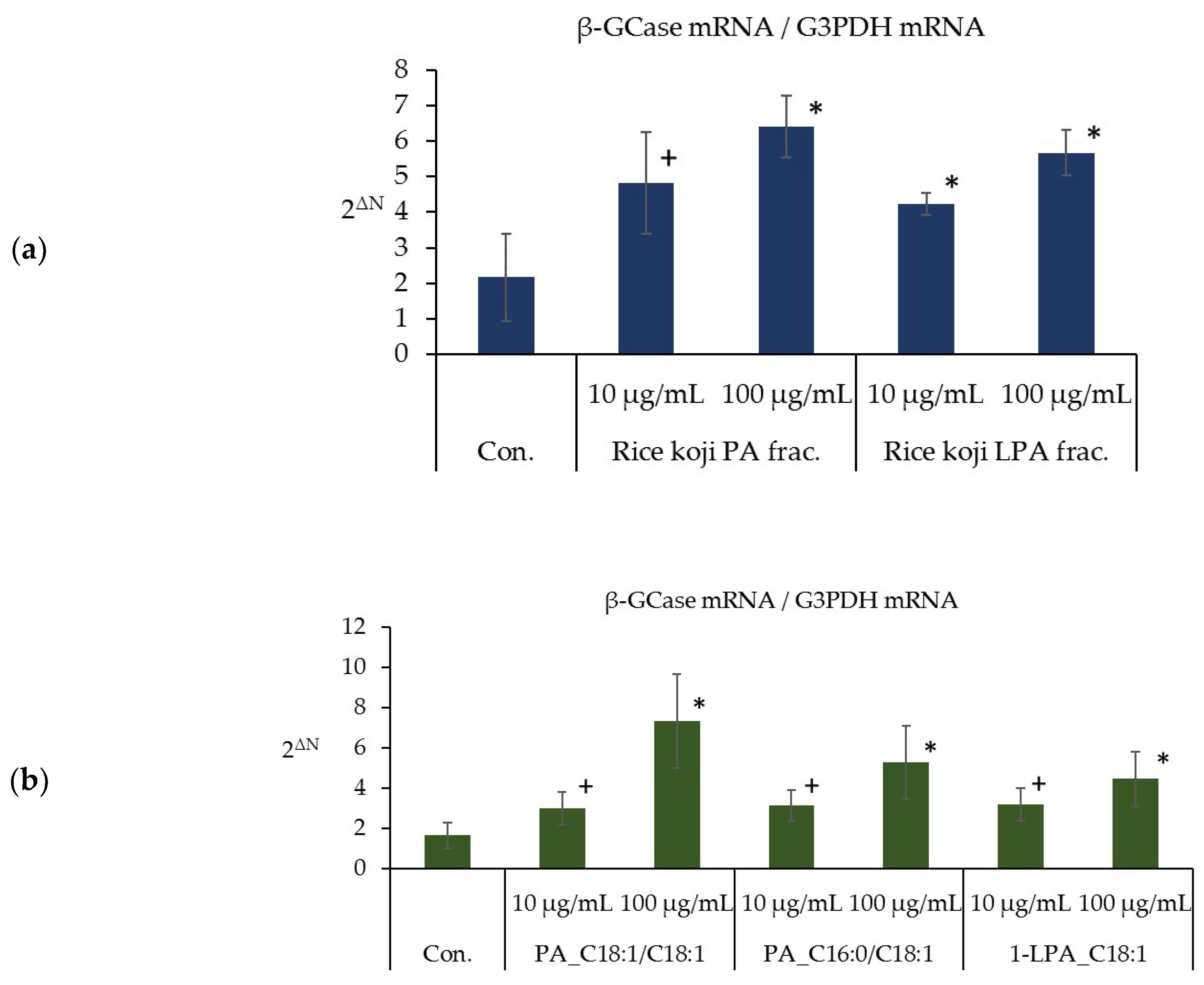
3.6. Effects of the PA and LPA Fractions of Rice Koji Extract, PA_C18:1/C18:1, PA_C16:0/C18:1, and 1-LPA_C18:1 on the Expression Level of β-GCase mRNA in the Three-Dimensional Human Epidermis Model
3.7. Measurement of the PA and LPA Contents in Steamed Rice and Rice Koji
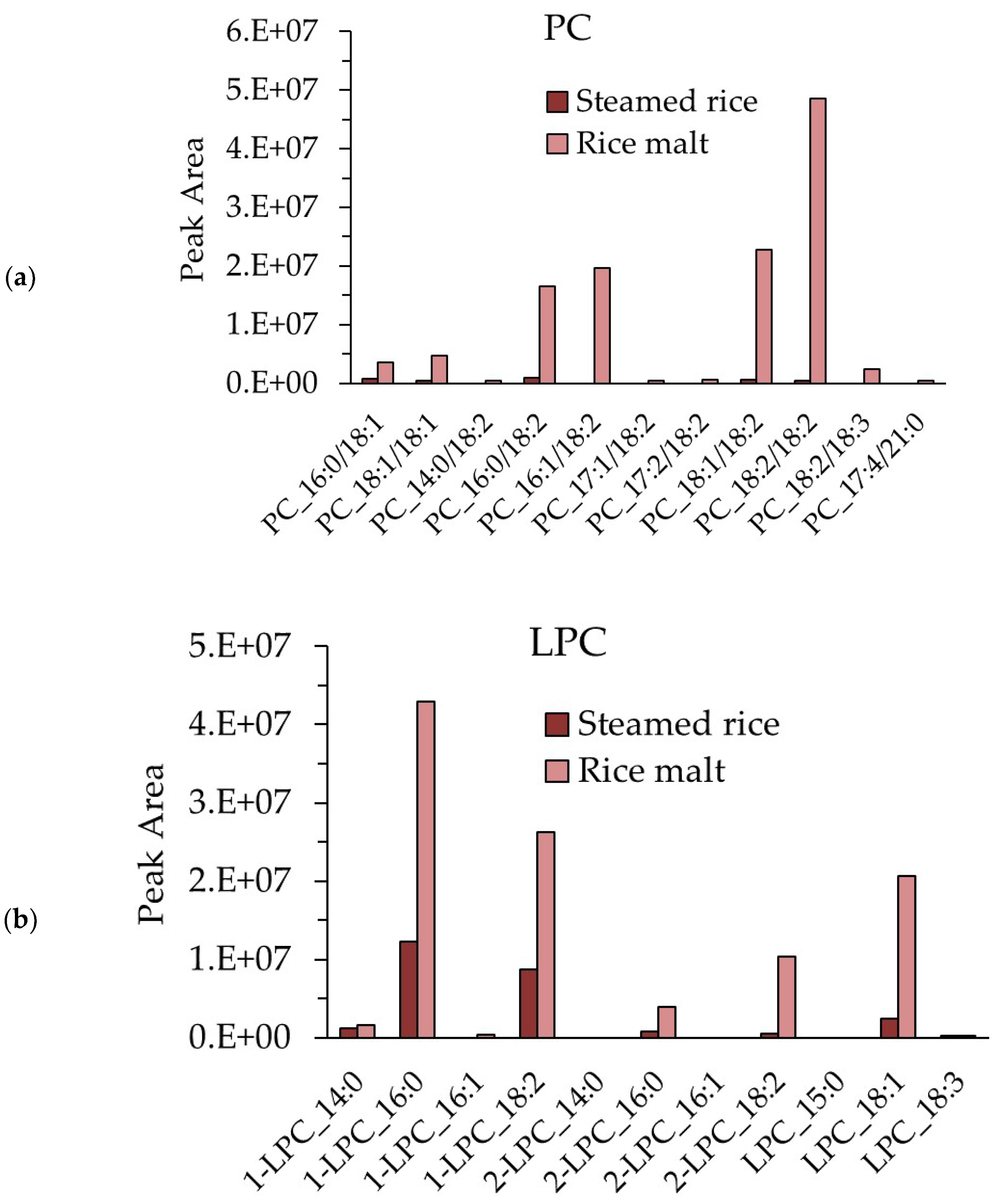
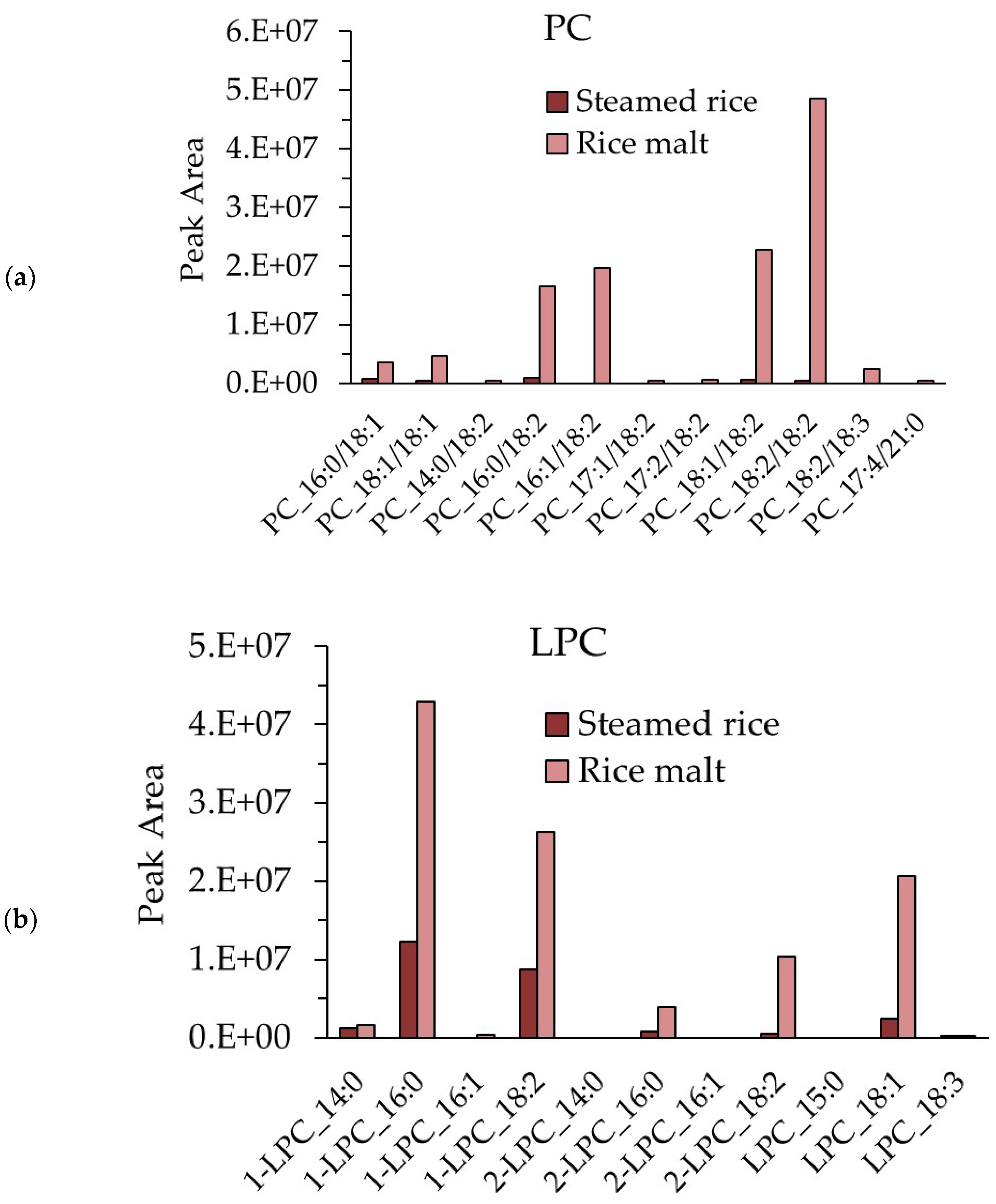
3.8. Measurement of the PC and LPC Contents in Steamed Rice and Rice Koji
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maeda, K.; Nakata, K.; Nakamura, A.; Kitagawa, M.; Ito, S. Improvement in skin conditions by consumption of traditional Japanese miso soup and its mechanism. J. Nutr. Food Sci. 2018, 8, 1. [Google Scholar] [CrossRef]
- Kitamoto, K. Bunshi Kojikin Gaku; Revised Version; The Brewing Society of Japan: Tokyo, Japan, 2012. (In Japanese) [Google Scholar]
- Iwano, K. Seisyu shuzou to kouso. J. Brew. Soc. Jpn. 1974, 74, 206–212. (In Japanese) [Google Scholar]
- Ministry of Education, Culture, Sports, Science and Technology, Japan. Standard Tables of Food Composition in Japan (Seventh Revised Version). 2015. Available online: http://www.mext.go.jp/en/policy/science_technology/policy/title01/detail01/1374030.htm (accessed on 14 April 2018).
- Ohnishi, K.; Yoshida, Y.; Toita, J.; Sekiguchi, J. Purification and characterization of a novel lipolytic enzyme from Aspergillus oryzae. J. Ferment. Bioeng. 1994, 78, 413–419. [Google Scholar] [CrossRef]
- Watanabe, T.; Owari, K.; Hori, K.; Takahashi, K. Selection of koji mold strain for making functional miso as rich antimutagenic activity. Nippon Shokuhin Kagaku Kogaku Kaishi 2004, 51, 698–702. (In Japanese) [Google Scholar] [CrossRef]
- Ohnishi, M.; Fujino, Y. Molecular species of free ceramides in Aspergillus oryzae. Agric. Biol. Chem. 1976, 40, 1419–1426. [Google Scholar] [CrossRef]
- Hosoyama, H.; Oisawa, M.; Hamano, M. Bifidobacterium growth promoting substance in rice bran koji extract. Nippon Shokuhin Kogyo Gakkaishi 1991, 38, 940–944. [Google Scholar] [CrossRef]
- Hamajima, H.; Matsunaga, H.; Fujikawa, A.; Sato, T.; Mitsutake, S.; Yanagita, T.; Nagao, K.; Nakayama, J.; Kitagaki, H. Japanese traditional dietary fungus koji Aspergillus oryzae functions as a prebiotic for Blautia coccoides through glycosylceramide: Japanese dietary fungus koji is a new prebiotic. Springerplus 2016, 5, 1321. [Google Scholar] [CrossRef] [PubMed]
- Kawasumi, T.; Sato, M.; Tsuriya, Y.; Ueno, S. Antioxidant activity of a medicine based on Aspergillus oryzae NK koji measured by a modified t-butyl peroxyl radical scavenging assay. Biosci. Biotechnol. Biochem. 1999, 63, 581–584. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Hiratake, J.; Aikawa, M.; Suizu, T.; Saito, Y.; Kawato, A.; Suginami, K.; Oda, J. Cysteine protease inhibitors produced by the industrial koji mold, Aspergillus oryzae O-1018. Biosci. Biotechnol. Biochem. 1998, 62, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, H.; Gu, L.; Zeng, H.; Maeda, K. Amino carbonylation of epidermal basement membrane inhibits epidermal cell function and is suppressed by methylparaben. Cosmetics 2017, 4, 38. [Google Scholar] [CrossRef]
- Feingold, K.R.; Elias, P.M. Role of lipids in the formation and maintenance of the cutaneous permeability barrier. Biochim. Biophys. Acta. 2014, 1841, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Groen, D.; Poole, D.S.; Gooris, G.S.; Bouwstra, J.A. Investigating the barrier function of skin lipid models with varying compositions. Eur. J. Pharm. Biopharm. 2011, 79, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Elias, P.M.; Menon, G.K. Structural and lipid biochemical correlates of the epidermal permeability barrier. Adv. Lipid Res. 1991, 24, 1–26. [Google Scholar] [PubMed]
- Norlén, L.; Nicander, I.; Lundh Rozell, B.; Ollmar, S.; Forslind, B. Inter- and intra- individual differences in human stratum corneum lipid content related to physical parameters of skin barrier function in vivo. J. Invest. Dermatol. 1999, 112, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Hamanaka, S.; Hara, M.; Nishio, H.; Otsuka, F.; Suzuki, A.; Uchida, Y. Human epidermal glucosylceramides are major precursors of stratum corneum ceramides. J. Invest. Dermatol. 2002, 119, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Van Smeden, J.; Hoppel, L.; van der Heijden, R.; Hankemeier, T.; Vreeken, R.J.; Bouwstra, J.A. LC/MS analysis of stratum corneum lipids: Ceramide profiling and discovery. J. Lipid Res. 2011, 52, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Holleran, W.M.; Takagi, Y.; Menon, G.K.; Jackson, S.M.; Lee, J.M.; Feingold, K.R.; Elias, P.M. Permeability barrier requirements regulate epidermal beta-glucocerebrosidase. J. Lipid Res. 1994, 35, 905–912. [Google Scholar] [PubMed]
- Hanley, K.; Jiang, Y.; Holleran, W.M.; Elias, P.M.; Williams, M.L.; Feingold, K.R. Glucosylceramide metabolism is regulated during normal and hormonally stimulated epidermal barrier development in the rat. J. Lipid Res. 1997, 38, 576–584. [Google Scholar] [PubMed]
- Uchida, Y.; Hara, M.; Nishio, H.; Sidransky, E.; Inoue, S.; Otsuka, F.; Suzuki, A.; Elias, P.M.; Holleran, W.M.; Hamanaka, S. Epidermal sphingomyelins are precursors for selected stratum corneum ceramides. J. Lipid Res. 2000, 41, 2071–2082. [Google Scholar] [PubMed]
- Jensen, J.M.; Fölster-Holst, R.; Baranowsky, A.; Schunck, M.; Winoto-Morbach, S.; Neumann, C.; Schütze, S.; Proksch, E. Impaired sphingomyelinase activity and epidermal differentiation in atopic dermatitis. J. Invest. Dermatol. 2004, 122, 1423–1431. [Google Scholar] [CrossRef] [PubMed]
- Oliveira Mdos, S.; Feddern, V.; Kupski, L.; Cipolatti, E.P.; Badiale-Furlong, E.; de Souza-Soares, L.A. Changes in lipid, fatty acids and phospholipids composition of whole rice bran after solid-state fungal fermentation. Bioresour. Technol. 2011, 102, 8335–8338. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Baranwal, V.; Shankar, A.; Kanwar, P.; Ranjan, R.; Yadav, S.; Pandey, A.; Kapoor, S.; Pandey, G.K. Rice phospholipase A superfamily: Organization, phylogenetic and expression analysis during abiotic stresses and development. PLoS ONE 2012, 7, e30947. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H. Phospholipase D of rice bran. I. Purification and characterization. Plant Sci. 1989, 59, 25–33. [Google Scholar] [CrossRef]
- Ueki, J.; Morioka, S.; Komari, T.; Kumashiro, T. Purification and characterization of phospholipase D (PLD) from rice (Oryza sativa L.) and cloning of cDNA for PLD from rice and maize (Zea mays L.). Plant Cell Physiol. 1995, 36, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, I.; Koishi, R.; Yao, Y.; Tsuji, T.; Serizawa, N. Molecular cloning and expression of the gene encoding a phospholipase A1 from Aspergillus oryzae. Biosci. Biotechnol. Biochem. 1999, 63, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Shiba, Y.; Ono, C.; Fukui, F.; Watanabe, I.; Serizawa, N.; Gomi, K.; Yoshikawa, H. High-level secretory production of phospholipaseA1 by Saccharomyces cerevisiae and Aspergillus oryzae. Biosci. Biotechnol. Biochem. 2001, 65, 94–101. [Google Scholar]
- Liu, L.; Waters, D.L.; Rose, T.J.; Bao, J.; King, G.J. Phospholipids in rice: significance in grain quality and health benefits: A review. Food Chem. 2013, 139, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.; Liu, L.; Waters, D.L.; Rose, T.J.; Bao, J.; King, G.J. Genotypic variation in lysophospholipids of milled rice. J. Agric. Food Chem. 2014, 62, 9353–9361. [Google Scholar] [CrossRef] [PubMed]
- AspGD. A. oryzae AO090005000433 Summary. The Board of Trustees, Leland Stanford Junior University. Available online: http://www.aspergillusgenome.org/cgi-bin/locus.pl?locus=AO090005000433#citations (accessed on 14 April 2018).
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maeda, K.; Ogino, Y.; Nakamura, A.; Nakata, K.; Kitagawa, M.; Ito, S. Identification of Rice Koji Extract Components that Increase β-Glucocerebrosidase Levels in Human Epidermal Keratinocytes. Foods 2018, 7, 94. https://doi.org/10.3390/foods7060094
Maeda K, Ogino Y, Nakamura A, Nakata K, Kitagawa M, Ito S. Identification of Rice Koji Extract Components that Increase β-Glucocerebrosidase Levels in Human Epidermal Keratinocytes. Foods. 2018; 7(6):94. https://doi.org/10.3390/foods7060094
Chicago/Turabian StyleMaeda, Kazuhisa, Yuuka Ogino, Ayano Nakamura, Keiji Nakata, Manabu Kitagawa, and Seiki Ito. 2018. "Identification of Rice Koji Extract Components that Increase β-Glucocerebrosidase Levels in Human Epidermal Keratinocytes" Foods 7, no. 6: 94. https://doi.org/10.3390/foods7060094
APA StyleMaeda, K., Ogino, Y., Nakamura, A., Nakata, K., Kitagawa, M., & Ito, S. (2018). Identification of Rice Koji Extract Components that Increase β-Glucocerebrosidase Levels in Human Epidermal Keratinocytes. Foods, 7(6), 94. https://doi.org/10.3390/foods7060094